
Platycodon grandiflorus Polysaccharides Alleviate Cr(VI)-Induced Apoptosis in DF-1 Cells via ROS-Drp1 Signal Pathway
 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Cell Culture and Treatment
2.3. Western Blot Analysis
2.4. Measurement of ROS
2.5. Measurement of MMP
2.6. Annexin V-FITC/PI Staining for Apoptosis Detection
2.7. Statistical Analysis
3. Results
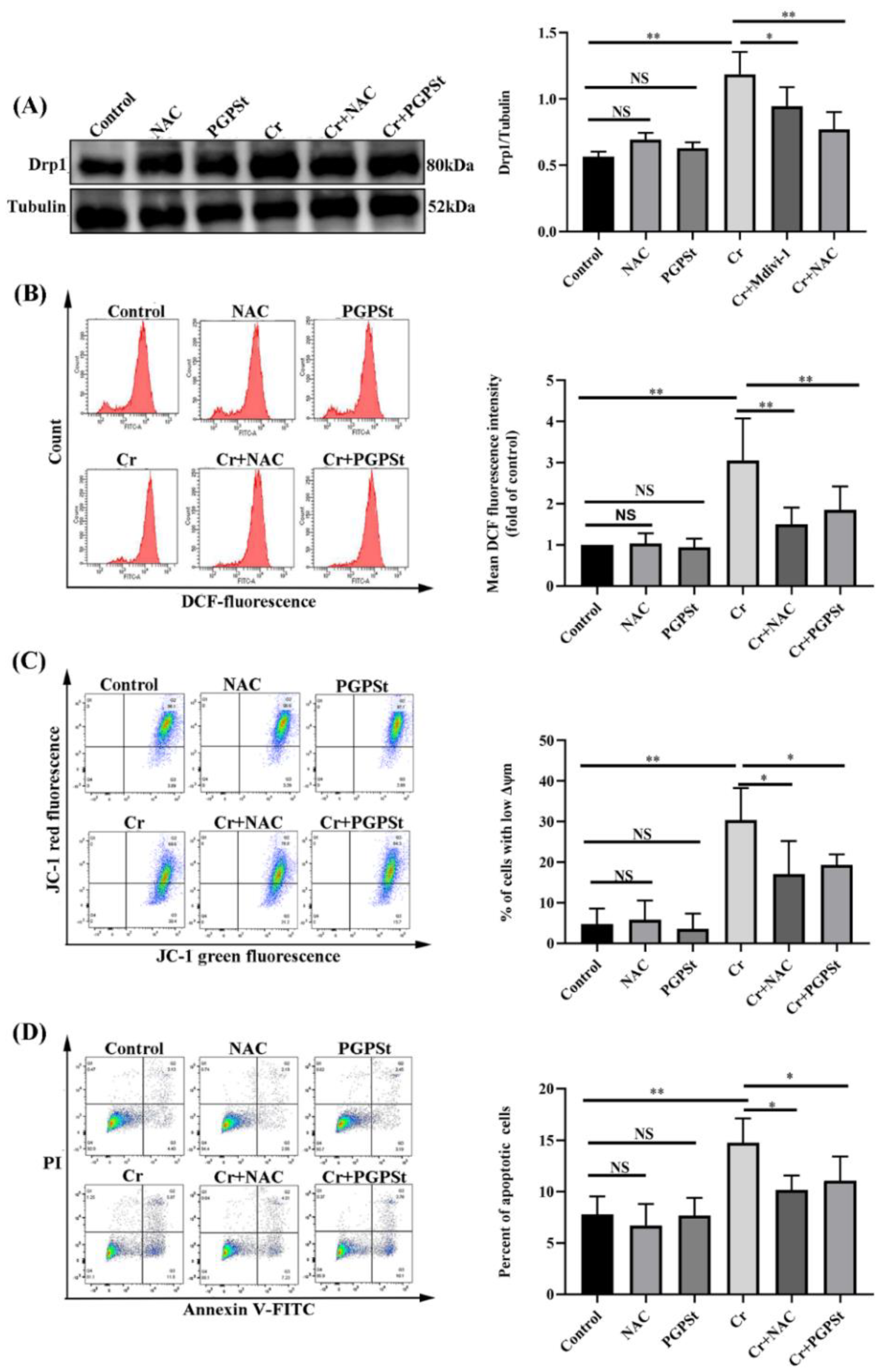
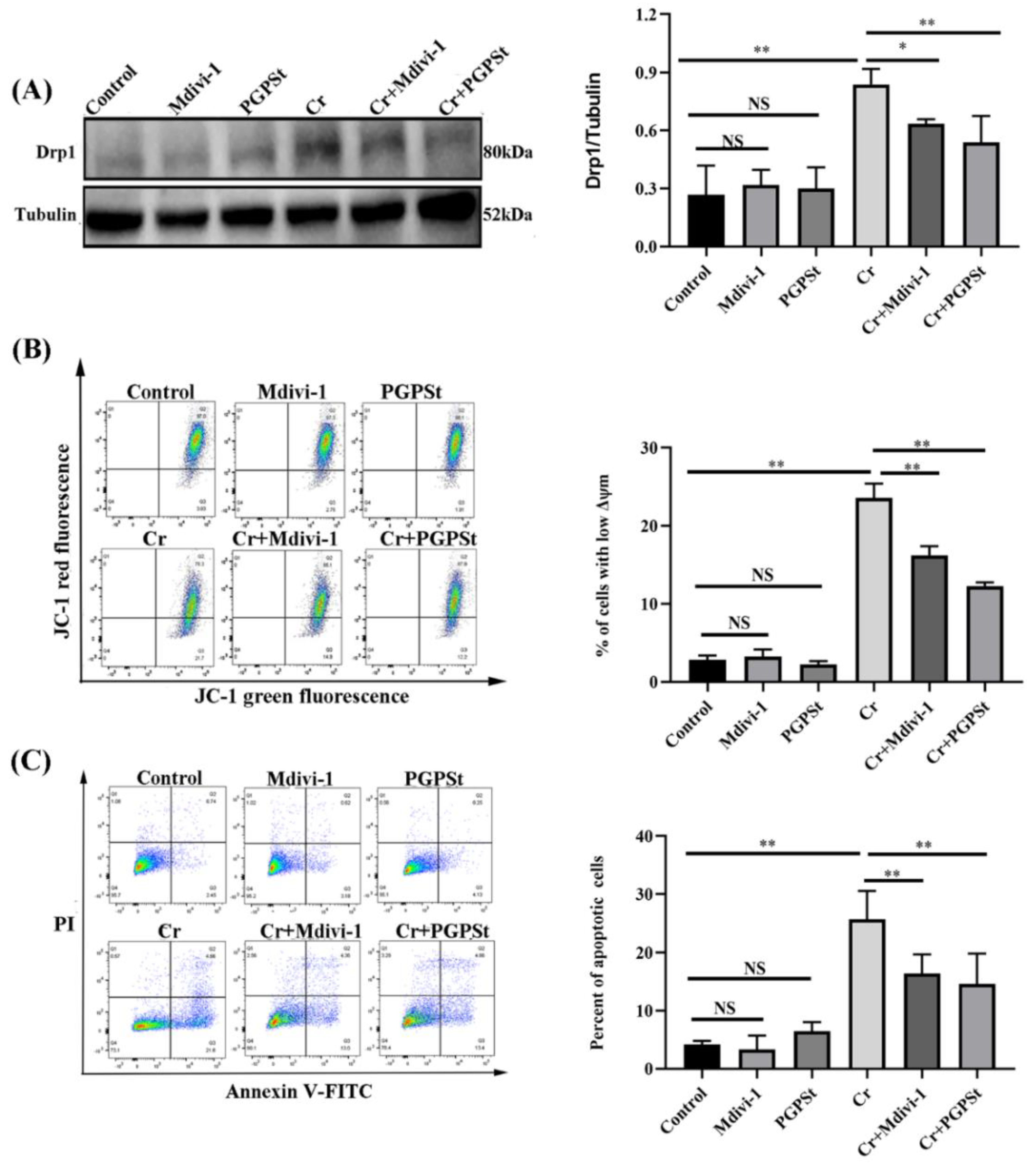
3.1. PGPSt Alleviated Cr(VI)-Induced Mitochondrial Damage
3.2. Inhibition of Cr(VI) Induction of Mitochondria-Dependent Apoptosis by PGPSt
3.3. Effect of NAC on Mitochondrial Damage and Cellular Apoptosis in Cr(VI)-Exposed DF-1 Cells
3.4. Effect of Midivi-1 on Mitochondrial Damage and Cellular Apoptosis in Cr(VI)-Exposed DF-1 Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Conflicts of Interest
References
- Son, Y.; Hitron, J.; Wang, X.; Chang, Q.; Pan, J.; Zhang, Z.; Liu, J.; Wang, S.; Lee, J.; Shi, X. Cr(VI) induces mitochondrial-mediated and caspase-dependent apoptosis through reactive oxygen species-mediated p53 activation in JB6 Cl41 cells. Toxicol. Appl. Pharmacol. 2010, 245, 226–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, S.C.; Liu, J.; Lee, K.; Tang, F.; Fang, K.; Yang, C.; Su, C.; Chen, H.; Hsu, R.; Chen, Y. Cr(VI) induces ROS-mediated mitochondrial-dependent apoptosis in neuronal cells via the activation of Akt/ERK/AMPK signaling pathway. Toxicol. In Vitro 2020, 65, 104795. [Google Scholar] [CrossRef] [PubMed]
- Stout, M.D.; Herbert, R.; Kissling, G.; Collins, B.; Travlos, G.; Witt, K.; Melnick, R.; Abdo, K.; Malarkey, D.; Hooth, M. Hexavalent chromium is carcinogenic to F344/N rats and B6C3F1 mice after chronic oral exposure. Environ. Health Perspect. 2009, 117, 716–722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, A.; Wang, Y.; Hao, J.; Wang, L.; Quan, L.; Duan, K.; Fakhar-e-AlamKulyar, M.; Ullah, K.; Zhang, J.; Wu, Y.; et al. Long-term hexavalent chromium exposure disturbs the gut microbial homeostasis of chickens. Ecotoxicol. Environ. Saf. 2022, 237, 113532. [Google Scholar] [CrossRef]
- Abreu, P.L.; Cunha-Oliveira, T.; Ferreira, L.; Urbano, A. Hexavalent chromium, a lung carcinogen, confers resistance to thermal stress and interferes with heat shock protein expression in human bronchial epithelial cells. Biometals 2018, 31, 477–487. [Google Scholar] [CrossRef]
- Yin, F.; Yan, J.; Zhao, Y.; Guo, K.; Zhang, Z.; Li, A.; Meng, C.; Guo, L. Bone marrow mesenchymal stem cells repair Cr (VI)- injured kidney by regulating mitochondria-mediated apoptosis and mitophagy mediated via the MAPK signaling pathway. Ecotoxicol. Environ. Saf. 2019, 176, 234–241. [Google Scholar] [CrossRef]
- Tang, S.; Ye, S.; Ma, Y.; Liang, Y.; Liang, N.; Xiao, F. Clusterin alleviates Cr(VI)-induced mitochondrial apoptosis in L02 hepatocytes via inhibition of Ca(2+)-ROS-Drp1-mitochondrial fission axis. Ecotoxicol. Environ. Saf. 2020, 205, 111326. [Google Scholar] [CrossRef]
- Zhai, R.; Dong, X.; Feng, L.; Li, S.; Hu, Z. The Effect of Heat Stress on Autophagy and Apoptosis of Rumen, Abomasum, Duodenum, Liver and Kidney Cells in Calves. Animals 2019, 9, 854. [Google Scholar] [CrossRef] [Green Version]
- Burin, S.M.; Ghisla, S.; Ouchida, A.; Aissa, A.; Coelho, M.; Costa, T.; Marsola, A.; Pinto-Simoes, B.; Antunes, L.; Curti, C.; et al. CR-LAAO antileukemic effect against Bcr-Abl(+) cells is mediated by apoptosis and hydrogen peroxide. Int. J. Biol. Macromol. 2016, 86, 309–320. [Google Scholar] [CrossRef]
- Wang, X.; Chen, C.; Zhou, G.; Ye, J.; Yin, R.; Feng, D.; Zhang, S.; Wang, X.; Zhao, X.; Zhang, Z. Sepia Ink Oligopeptide Induces Apoptosis of Lung Cancer Cells via Mitochondrial Pathway. Cell Physiol. Biochem. 2018, 45, 2095–2106. [Google Scholar] [CrossRef]
- Hu, C.; Huang, Y.; Li, L. Drp1-Dependent Mitochondrial Fission Plays Critical Roles in Physiological and Pathological Progresses in Mammals. Int. J. Mol. Sci. 2017, 18, 144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Hu, C.; Huang, J.; Liu, W.; Lai, W.; Leng, F.; Tang, Q.; Liu, Y.; Wang, Q.; Zhou, M.; et al. ROCK1 induces dopaminergic nerve cell apoptosis via the activation of Drp1-mediated aberrant mitochondrial fission in Parkinson’s disease. Exp. Mol. Med. 2019, 51, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Xiao, Y.; Ma, Y.; Liang, N.; Liang, Y.; Lu, C.; Xiao, F. ROS-mediated miR-21-5p regulates the proliferation and apoptosis of Cr(VI)-exposed L02 hepatocytes via targeting PDCD4. Ecotoxicol. Environ. Saf. 2020, 191, 110160. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Zhang, H.; Li, J.; Jiang, X.; Zhang, Y.; Wu, Q.; Shen, L.; Shi, J.; Gao, N. ROCK1 activation-mediated mitochondrial translocation of Drp1 and cofilin are required for arnidiol-induced mitochondrial fission and apoptosis. J. Exp. Clin. Cancer Res. 2020, 39, 37. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Zhang, Y.; Jiang, X.; Zhang, H.; Gao, Z.; Li, Y.; Fu, R.; Li, L.; Li, J.; Cui, H.; et al. ROS-mediated activation and mitochondrial translocation of CaMKII contributes to Drp1-dependent mitochondrial fission and apoptosis in triple-negative breast cancer cells by isorhamnetin and chloroquine. J. Exp. Clin. Cancer Res. 2019, 38, 225. [Google Scholar] [CrossRef]
- Li, L.; Xu, W.; Yi, C.; Cheng, Y.; Xin, H.; Xue, V.; Li, C.; Fang, X.; Yang, L.; Chen, C.; et al. Astragalus polysaccharide has a protective effect on hematopoiesis in an irradiated mouse model and decreases apoptosis in megakaryocytes. Mol. Med. Rep. 2021, 23, 1. [Google Scholar] [CrossRef]
- Cao, P.; Sun, J.; Sullivan, M.; Huang, X.; Wang, H.; Zhang, Y.; Wang, N.; Wang, K. Angelica sinensis polysaccharide protects against acetaminophen-induced acute liver injury and cell death by suppressing oxidative stress and hepatic apoptosis in vivo and in vitro. Int. J. Biol. Macromol. 2018, 111, 1133–1139. [Google Scholar] [CrossRef]
- Zou, Y.F.; Chen, M.; Fu, Y.; Zhu, Z.; Zhang, Y.; Paulsen, B.; Rise, F.; Chen, Y.; Yang, Y.; Jia, R.; et al. Characterization of an antioxidant pectic polysaccharide from Platycodon grandiflorus. Int. J. Biol. Macromol. 2021, 175, 473–480. [Google Scholar] [CrossRef]
- Xing, Y.; Wang, L.; Xu, G.; Guo, S.; Zhang, M.; Cheng, G.; Liu, Y.; Liu, J. Platycodon grandiflorus polysaccharides inhibit Pseudorabies virus replication via downregulating virus-induced autophagy. Res. Vet. Sci. 2021, 140, 18–25. [Google Scholar] [CrossRef]
- Zheng, P.; Fan, W.; Wang, S.; Hao, P.; Wang, Y.; Wan, H.; Hao, Z.; Liu, J.; Zhao, X. Characterization of polysaccharides extracted from Platycodon grandiflorus (Jacq.) A.DC. affecting activation of chicken peritoneal macrophages. Int. J. Biol. Macromol. 2017, 96, 775–785. [Google Scholar] [CrossRef]
- Wang, C.; Cheng, G.; Yang, S.; Li, L.; Zhang, Y.; Zhao, X.; Liu, J. Protective effects of Platycodon grandiflorus polysaccharides against apoptosis induced by carbonyl cyanide 3-chlorophenylhydrazone in 3D4/21 cells. Int. J. Biol. Macromol. 2019, 141, 1220–1227. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Song, Y.; Tian, B.; Qi, C.; Li, L.; Wang, L.; Xing, Y.; Zhao, X.; Liu, J. The role of ATF6 in Cr(VI)-induced apoptosis in DF-1 cells. J. Hazard. Mater. 2021, 410, 124607. [Google Scholar] [CrossRef]
- Hao, J.; Zhao, X.; Hao, J.; Zhu, Y.; Wang, Y.; Wang, L.; Guo, S.; Yi, H.; Liu, Y.; Liu, J. Platycodon grandifloras polysaccharides inhibit mitophagy injury induced by Cr (VI) in DF-1 cells. Ecotoxicol. Environ. Saf. 2020, 202, 110901. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Zhu, Y.; Wan, H.; Wang, Y.; Hao, P.; Cheng, Z.; Liu, Y.; Liu, J. Effects of the Oral Administration of K2Cr2O7 and Na2SeO3 on Ca, Mg, Mn, Fe, Cu, and Zn Contents in the Heart, Liver, Spleen, and Kidney of Chickens. Biol. Trace Elem. Res. 2017, 180, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Wilkinson, K.; Evans, A.; Rubin, P.; Henley, J. SENP3-mediated deSUMOylation of Drp1 facilitates interaction with Mff to promote cell death. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, C.Q.; Guo, Y.; Chu, X.Y. Neuropathic Pain: The Dysfunction of Drp1, Mitochondria, and ROS Homeostasis. Neurotox. Res. 2020, 38, 553–563. [Google Scholar] [CrossRef]
- Yang, J.; Chen, W.; Sun, Y.; Xia, P.; Liu, J.; Zhang, W. The role of microRNAs in regulating cadmium-induced apoptosis by targeting Bcl-2 in IEC-6 cells. Toxicol. Appl. Pharmacol. 2021, 432, 115737. [Google Scholar] [CrossRef]
- Sivakumar, K.K.; Stanley, J.; Behlen, J.; Wuri, L.; Dutta, S.; Wu, J.; Arosh, J.; Banu, S. Inhibition of Sirtuin-1 hyperacetylates p53 and abrogates Sirtuin-1-p53 interaction in Cr(VI)-induced apoptosis in the ovary. Reprod. Toxicol. 2022, 109, 121–134. [Google Scholar] [CrossRef]
- Shi, L.; Feng, L.; Tong, Y.; Jia, J.; Li, T.; Wang, J.; Jiang, Z.; Yu, M.; Xia, H.; Jin, Q.; et al. Genome wide profiling of miRNAs relevant to the DNA damage response induced by hexavalent chromium exposure (DDR-related miRNAs in response to Cr (VI) exposure). Environ. Int. 2021, 157, 106782. [Google Scholar] [CrossRef]
- Ge, H.; Li, Z.; Jiang, L.; Li, Q.; Geng, C.; Yao, X.; Shi, X.; Liu, Y.; Cao, J. Cr (VI) induces crosstalk between apoptosis and autophagy through endoplasmic reticulum stress in A549cells. Chem. Biol. Interact. 2019, 298, 35–42. [Google Scholar] [CrossRef]
- Liang, N.; Li, S.; Liang, Y.; Ma, Y.; Tang, S.; Ye, S.; Xiao, F. Clusterin inhibits Cr(VI)-induced apoptosis via enhancing mitochondrial biogenesis through AKT-associated STAT3 activation in L02 hepatocytes. Ecotoxicol. Environ. Saf. 2021, 221, 112447. [Google Scholar] [CrossRef] [PubMed]
- Maycotte, P.; Marin-Hernandez, A.; Goyri-Aguirre, M.; Anaya-Ruiz, M.; Reyes-Leyva, J.; Cortes-Hernandez, P. Mitochondrial dynamics and cancer. Tumour. Biol. 2017, 39, 8391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, L.L.; Shi, F.; Tan, Z.; Li, Y.; Bode, A.; Cao, Y. Mitochondrial network structure homeostasis and cell death. Cancer Sci. 2018, 109, 3686–3694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Y.; Ming, V.; Gong-Hua, H.; Lan, G.; Lu, D.; Peng, L.; Feng, J.; Cai-Gao, Z. Cr(VI) induces the decrease of ATP level and the increase of apoptosis rate mediated by ROS or VDAC1 in L-02 hepatocytes. Environ. Toxicol. Pharmacol. 2012, 34, 579–587. [Google Scholar] [CrossRef]
- Kim, D.; Sesaki, H.; Roy, S. Reduced Levels of Drp1 Protect against Desvelopment of Retinal Vascular Lesions in Diabetic Retinopathy. Cells 2021, 10, 1379. [Google Scholar] [CrossRef]
- Zhao, X.; Wang, Y.; Yan, P.; Cheng, G.; Wang, C.; Geng, N.; Wang, X.; Liu, J. Effects of Polysaccharides from Platycodon grandiflorum on Immunity-Enhancing Activity In Vitro. Molecules 2017, 22, 1918. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Zheng, P.; Qi, C.; Cui, Y.; Qi, Y.; Xue, K.; Yan, G.; Liu, J. Platycodon grandiflorus Polysaccharides Alleviate Cr(VI)-Induced Apoptosis in DF-1 Cells via ROS-Drp1 Signal Pathway. Life 2022, 12, 2144. https://doi.org/10.3390/life12122144
Zhang Z, Zheng P, Qi C, Cui Y, Qi Y, Xue K, Yan G, Liu J. Platycodon grandiflorus Polysaccharides Alleviate Cr(VI)-Induced Apoptosis in DF-1 Cells via ROS-Drp1 Signal Pathway. Life. 2022; 12(12):2144. https://doi.org/10.3390/life12122144
Chicago/Turabian StyleZhang, Zhuanglong, Pimiao Zheng, Changxi Qi, Yuehui Cui, Yijian Qi, Kun Xue, Guangwei Yan, and Jianzhu Liu. 2022. "Platycodon grandiflorus Polysaccharides Alleviate Cr(VI)-Induced Apoptosis in DF-1 Cells via ROS-Drp1 Signal Pathway" Life 12, no. 12: 2144. https://doi.org/10.3390/life12122144