1. Introduction
Huang-Qin (
Scutellaria baicalensis Georgi) is harvested for its fresh root with annual yield about 17 tons in Taiwan. The dry roots slice of Huang-Qin belongs to nutraceutical can be easily accessed from supermarkets is a popular use botanical herb, commonly prepared with four-entity {sì wù} chicken soup as a summer health-preserving formula in Chinese society. Recently, Huang-Qin is renowned as a critical ingredient in the Chinese herbal compound medicine NRICM101 (清冠一號) in treating COVID-19. The baicalein, baicalin and wogonin are the three major flavonoids identified in Huang-Qin that only wogonin exhibit potent anti-inflammatory property by inhibiting both gene expressions of
iNOS (induced nitric oxide synthase) and
COX-2 (cyclooxygenase-2) in LPS-activated RAW264.7 macrophages [
1]. Particularly, the COX-2 gene inhibition property has also been demonstrated in human lung epithelial cancer cells [
2], and wogonin has also shown to prevent LPS-induced lung inflammation via activation of PPARγ (peroxisome proliferation-activated receptor γ) [
3]. Nevertheless, wogonin also possess many other potent therapeutic functions in anti-cancer, anti-viral infection, and neuroprotection etc [
4,
5] in additional to its anti-inflammatory property.
Wogonin is currently under intensive studies for its anti-cancer activity primarily evidenced by induction of oxidative stress in attribution to cell apoptosis. Acting as an inhibitor of apoptosis, survivin is not only a normal cell cycle protein expressed in healthy mitotic cells, but also notoriously as a cancer marker highly expressed in progressive cancers [
6]. It has been ubiquitously reported wogonin is able to attenuate survivin activity in various cancer cells models [
7,
8,
9]. Nevertheless, survivin plays important role in immune system. It is essential for proliferation and differentiation of normal lymphocytes and macrophages. Lymphoblastogenesis is a fundamental effector response of antigen-activated lymphocytes in mediating acquired immunity. It has been reported survivin knocked out has completely blocked B cell development arrested at pre-B cell stage and impaired B cell clone expansion [
10]. Moreover, T cell-specific surviving-deficient mice have mainly immature T cells and diminished peripheral T cells population indicates an important role in T cell development [
11]. Survivin has also been reported to express in LPS or IL-4-activated macrophages with M2 polarization [
12]. Whether cytotoxic and apoptotic roles of wogonin in cancer cells could be deleterious to mitotic lymphoblastogenesis as fast proliferating lymphocytes in responding to antigen stimulation have to be monitored for potential immunotoxicity.
Although wogonin exhibit multiple therapeutic functions, its bioavailability is low as demonstrated in pharmacological study. The deglycosylated wogonin is not easily absorbed due to low solubility in water, oral bioavailability is only 1.1% [
13]. On the other hand, wogonosides of glycosylated form wogonin can quickly mount a high concentration within hours in rat post oral administration [
14]. To improve its proficiency, various hydrophilic carriers are available such as polyethylene glycol to improve the water insoluble compound such as wogonin. Hence, to improve wogonin bioavailability and optimizing delivery are under intensive studies and reported [
15,
16]. Using LC-MS/MS tracking wogonin absorption in beagle dogs shows limited blood uptake and can be improved by hydrophilic carrier arginine solution [
15]. Previously, chemically synthesized wogonin derivatives at the 7-
O-position of the A ring with different number of prenyl units exhibiting enhanced anti-cancer activity by arrest S phase via an apoptotic mechanism [
17]. The lipophilic property of the prenyl conjugation to wogonin has been claimed to contribute to the enhanced anti-cancer function. The outcomes in anti-cancer with promoting wogonin bioavailability by increasing lipophilic activity with flavonoid terpenylation has shown some promised results.
Chemically synthesized terpenylated wogonin derivatives has posed some new indications in its bioavailability and delivery. In addition, the terpenylation can also expand biological exploration of the category of natural scarce terpenylated flavonoids. Naturally, majority of flavonoids is
C-terpenylated, the
O-terpenylation is even rare and most of them occur as secondary metabolites synthesized in response to pathogenic microbial infection or challenge of abiotic elicitors in some plants [
18]. Regardless the structural diversified terpenylated flavonoids have been only found in limited plant families of Leguminosae, Moraceae, Asteraceae of mainly anatomical sections of roots and barks [
18], terpenylated flavonoids are also identified in anti-microbial sealant, propolis, of bee hive [
19,
20]. To learn from nature, chemically synthesized terpenylated flavonoids are of our interest in the perspective of a mimicry to nature and together to promote bioavailability of the anti-inflammatory nature of wogonin for better therapeutic applications. The chemically synthesized 7-
O-terpenylated wogonin are therefore deemed for further investigations. Currently, the immunotoxicity and anti-inflammatory characteristics are assessed in facilitating scientific basis for its future development in immunomodulatory purposes.
2. Materials and Methods
2.1. Purification of Wogonin and Synthesis of Terpenylated Wogonin Derivatives
Wogonin (W) was extracted from dried
Scutellaria baicalensis Georgi roots that were cut into small pieces, immersed, and extracted with 10-times
v/
w acetone twice at room temperature for 2 weeks. After filtration, the residues were reflux-extracted with 4-times
v/
w 50% aqueous ethanol twice. Acetone and 50% aqueous ethanol extracts were concentrated, then these extracts were subjected to column chromatography on silica gel eluted with CHCl
3 and CHCl
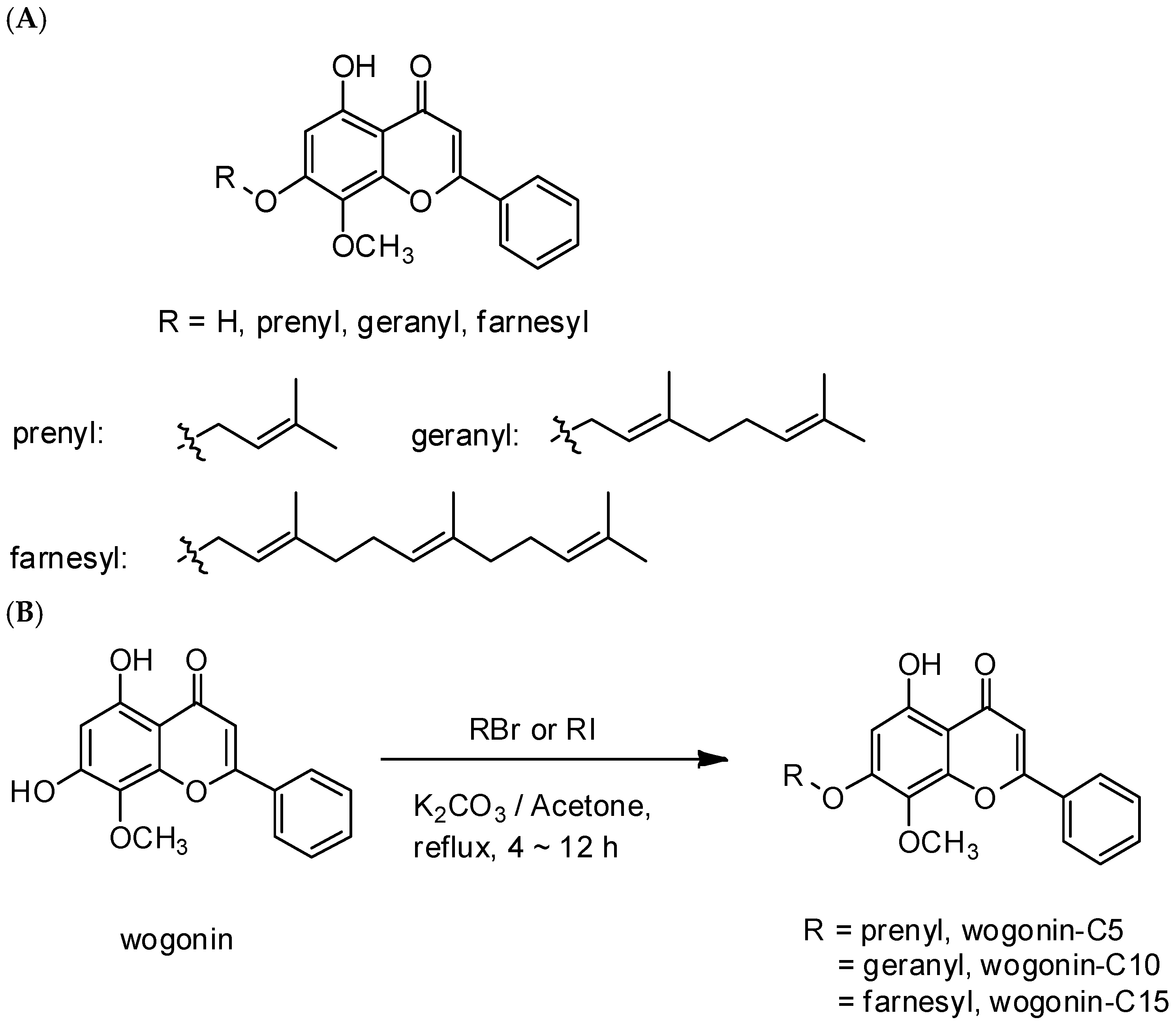
3-MeOH and rechromatographed on silica gel eluted with hexane-acetone to yield W. The readily available W was the starting material for the preparation of terpenylated wogonin derivatives with different units of isoprene. Treatment of W with prenyl bromide, geranyl bromide, and farnesyl bromide in the presence of K
2CO
3 gave hemiterpene (prenyl wogonin; W5), monoterpene (geranyl wogonin; W10), and sesquiterpene (farnesyl wogonin; W15) wogonin derivatives, respectively. A scheme for the chemical synthesizing 7-
O-prenyl derivatives of W is demonstrated in
Scheme 1.
2.2. Antioxidative Capacity
2.2.1. DPPH Assay
The quenching of free radicals by wogonin and wogonin derivatives evaluated spectrophotometrically at 517 nm against the absorbance of the DPPH (1,1-diphenyl-2-picrylhydrazyl) radical. A freshly prepared DPPH solution (10 mM) was used for the assay. Samples were dissolved in ethanol and the ethanolic solution of DPPH served as a control. The degree of decoloration indicates the free radical scavenging efficiency of the substances. Quercetin and trolox were used as a reference free radical scavenger.
2.2.2. TEAC Assay
ABTS (2,2′-azino-bis-(3-ethylbenthiazoline sulfonic acid)) was used in conjunction with peroxidase and hydroperoxide to generate an oxidized form (ABTS radical). The aliquot of W or respective synthesized derivatives were evaluated against the ABTS radical substrate. The inhibition on the blue greenish color formation was detected with UV-vis absorbance wave length = 734 nm. A trolox standard curve was plotted to obtained relative antioxidative capacity.
2.3. Immunotoxicity Assessment
2.3.1. Cell Culture
Murine cell lines of myeloid origin RAW264.7 macrophages (BCRC#60001) and lymphoid origin YAC-1 cells line (BCRC#60147l) and somatic cell line, BNL CL2 embryonic liver cell line (BCRC#60180) were purchased from Bioresource Center and Research Center of the Food and Industrial Research Institute, Taiwan. The cell line was maintained and sub-cultured as recommend. Sub-cultured cells used for the entire study were not exceeding 10th passages. Fresh culture medium was substituted together with transferring into 24-well or 96-well culture plates one day before assay. The seeding density was 1 × 106/well or 2 × 105/well, respectively, for cytotoxic assay and cell cycle analysis. Culture medium was DMEM (Dulbecco’s modified Eagle’s medium) or RPMI 1640 (Roswell Park Memorial Institute) basal medium, which was containing 2 mM L-glutamine and 10% fetal bovine serum (Sigma, Co., Ltd, St. Louis, MO, USA) for the anchorage-dependent or -independent cells uses.
2.3.2. Cell Viability Assay and Microscopic Examination
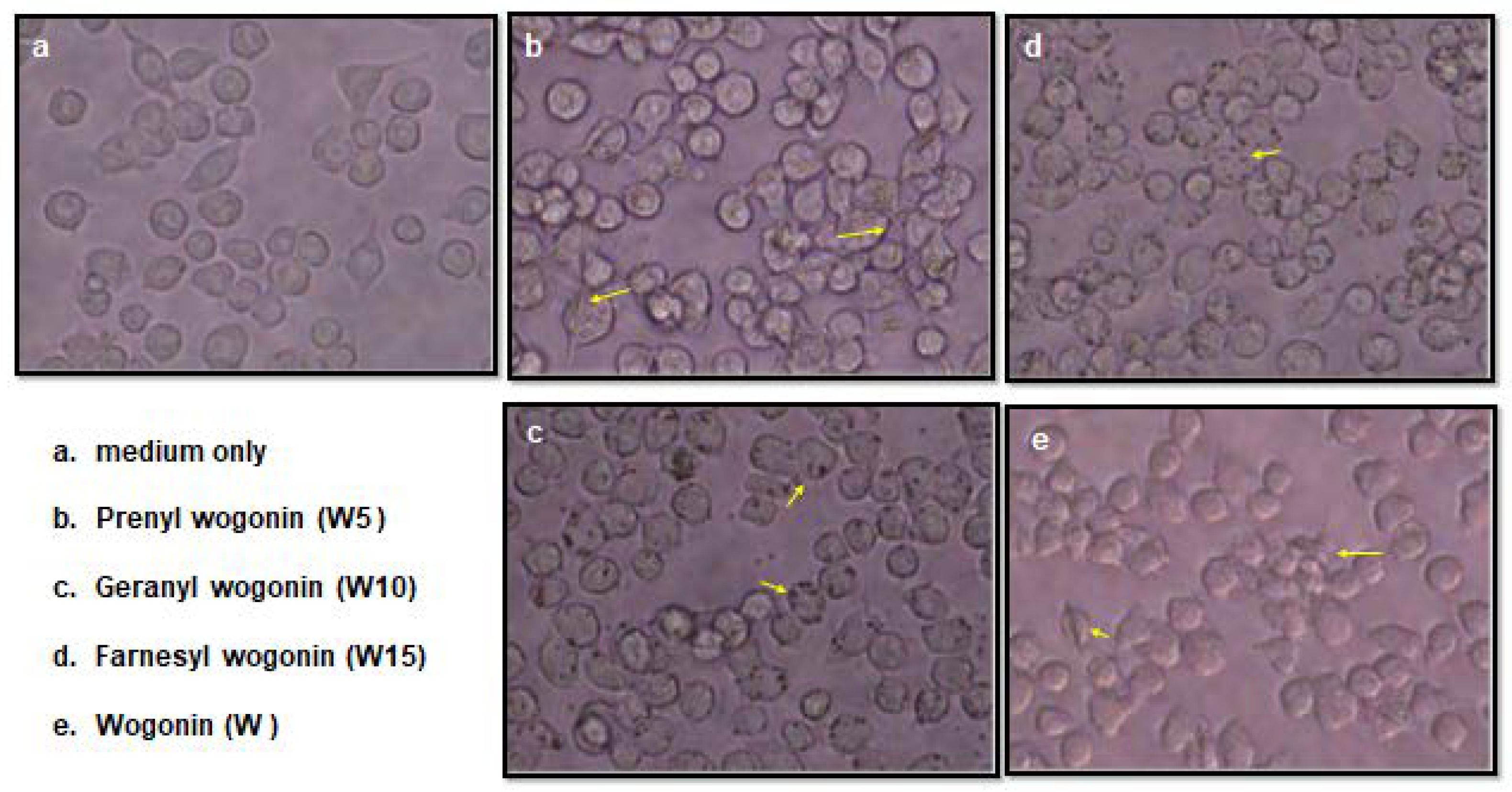
Cell viability was assessed by staining with 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT), the murine macrophages cell line, RAW 264.7 cell, was plated at a density of 1 × 105 cell/well into 96-well plates. After an overnight conditioning, cells were treated with different concentrations of terpenylated wogonin derivatives for 24 h. At the end of treatment, 20 μL MTT (5 mg/mL) was added, and cells were incubated for 3 h prior to the colorimetrical determination by ELISA plate reader at a 570 nm wavelength (Bio-Tek Instruments, Inc., Winooski, VT, USA). The results were obtained from three independent tests. Seeding in same density, macrophages cultured with 50 μM of terpenylated wogonin derivatives treatments for 1.5 h were subjected for microscopic examination. Photographs were taken at 400× magnification with a CCD equipped microscope (Olympus CKX 41, Japan). Rod and micelle formations of wogonin and terpenylated wogonin derivatives treatments were resolved over 3 h and thereafter monitoring micelle until last observation at 24 h cell culture.
2.4. Anti-Inflammatory Function Assessment
2.4.1. Nitric Oxide Production with Griess Test
Extracellular NO production was predicted as nitrite concentration in cell suspension by the Griess reaction. RAW264.7 were incubated with wogonin and its terpenylated derivatives at concentration of 50 μM and stimulated with 100 ng/mL LPS (lipopolysaccharides, Sigma Chemical Co., St. Louis, MO, USA) for 24 h at 5%, 37 °C, humidified incubator. After incubation, 50 μL supernatant was withdrawn from each well and transferred into another 96-well plate for quantification of NO by reaction with Griess reagent. To each well, 50 μL 1% sulphanilamide and 50 μL 0.1% naphthylethene diamine hydrochloride were added in order and then incubated at dark for 15 min at room temperature. Absorbance at 550 nm of the reactants was analyzed by a 96-well plate reader (Bio-Tek Instruments, Inc., Winooski, VT, USA). A standard curve was constructed by preparation of a series of sodium nitrite solutions from 0 to 200 μM.
2.4.2. Intracellular Oxidative Status of Macrophage
The protection of terpenylated wogonin derivatives on LPS-induced reactive oxygen intermediates productions was measured by DCFH (2′,7′-dichlorofluorescin) oxidized fluorescent products retained in RAW264.7 cells. Cells incubated with 50 μM terpenylated wogonin derivatives and 100 ng/mL LPS were loaded with DCFH-DA (dichlorofluorescin diacetate) dye for detecting the amount of intracellular reactive oxygen species. Analysis were performed at 3, 6, 24, and 48 h. Upon a 10 min incubation with DCFH-DA dye loading at dark, room temperature, cells were quickly immersed into ice-cold water incubation prior to flow cytometry analysis (Becton Dickson, Franklin Lakes, NJ, USA). Data analysis was completed by the CellQuest software (Becton Dickson, Franklin Lakes, NJ, USA). The percentage of cells population exerts fluorescence more than the second log values were considered and the geometrical means of fluorescent intensity were recorded.
2.5. Flow Cytometry Analysis for Cell Cycle of YAC-1
Cultured lymphoid YAC-1 cells were harvested at an exponential growth phase, and single cell suspensions containing 1 × 106 cells were treated as described previously in cell livability assay at concentrations of 10, 50 and 100 μM for 24 h. At the end of culture, cells were fixed with 70% alcohol. Cell cycle was monitored using propidium iodide (PI) staining of nuclei. The fluorescence of DNA-bound PI in cells was measured with a flow cytometer (FACScan®, Becton Dickson, Franklin Lakes, NJ, USA) and the results were analyzed with ModFit software (Becton Dickson, Franklin Lakes, NJ, USA).
2.6. mRNA Expression of Survivin and iNOS by Semi-Quantitative RT-PCR Analysis
Briefly, 1 × 106 cells were harvested at designated experimental stages for RNA extractions. Total RNA was extracted using the Trizol reagent following the manufacturer’s instruction. RNA was reverse transcribed using a cDNA synthesis kit (Promega, Madison, WI, USA). Synthesized cDNA was then amplified with Taq polymerase and specific primers for survivin, iNOS (inducible nitric oxide synthase) and GAPDH (glyceraldehyde-3-phosphate dehydrogenase) in a thermal cycler. Optimum conditions for RT-PCR were 54 °C for 1 min, 56 °C for 1 min, and 56 °C for 1 min for 30 cycles for survivin, 35 cycles for iNOS, and 30 cycles for GAPDH, respectively, and a final extension at 72 °C for 7 min. The primers used were 5′-CCG GTG AAG GAG ACA CCG T-3′ (sense) and 5′-GTG TTC TGA CCA GGC ACG AT-3′ (antisense) for survivin; and 5′-GTC AAC TGC AAG AGA ACG GAG AAC-3′ (sense) and 5′-GAG CTC CTC CAG AGG GTA GGC T-3′ (antisense) for iNOS; and 5′-CAG AGC TCC AAT CAA CTG TCG-3′(sense) and 5′-TAA GTT AGA TCC AGC TCC GAC T-3′ (antisense) for GAPDH. A total of 25 μL of final PCR products was analyzed by electrophoresis on 1.2% agarose gel. The bands were visualized with ethidium bromide and each band was measured and analyzed using Gel-Pro Analyzer® software (Media Cybernetics, Inc., Silver Spring, MD, USA).
2.7. Statistics
The statistical analysis was performed using Student’s t test where significance was specified as * mark at p-value less than 0.05. Except the DPPH and ABTS assays were results of duplicated samples in average. Data were presented as mean ± standard deviation. For flow cytometric analysis of cell cycle, the one-way ANOVA was used flowed by treatment comparison by LSD test (SAS software, 1996).
4. Discussion
Terpenylated flavonoids are mainly
C-prenylated. The
O-prenylated structures are unique chemicals naturally synthesized in limited plant families of Leuminosae, Moraceae, and Astreaceae, mainly as secondary metabolites in defense against microorganisms [
18]. Propolis for building bee hives also exist prenylated flavonoids [
20,
21], and they may exhibit multiple biological functions, including antioxidant, anti-microbial anti-inflammation, anti-cancer activities, etc. Due to their naturally scarce occurrence, chemically synthesized to promote anti-cancer activity by increasing lipophilic tendency of baicalein, wogonin, and chrysin of 7-
O-alkylated isoprene have been reported for specific comparisons [
17]. Recently, 7-
O-alkyl or 7-
O-acyloxy derivatives of natural flavonoids (naringenin and apigenin) have been synthesized chemically to avoid methylation, sulfation, glucuronidation, and phenolation during phase-two metabolism and to enhance their functions [
22,
23]. Currently, the 7-
O-alkylated wogonin with one to three isoprene units termed prenyl wogonin (W5), geranyl wogonin (W10), and farnesyl wogonin (W15) have been shown to increase lipophilic activity as observed the preferential membrane-bound rod or micelle-like entities (
Figure 2) along with the clog
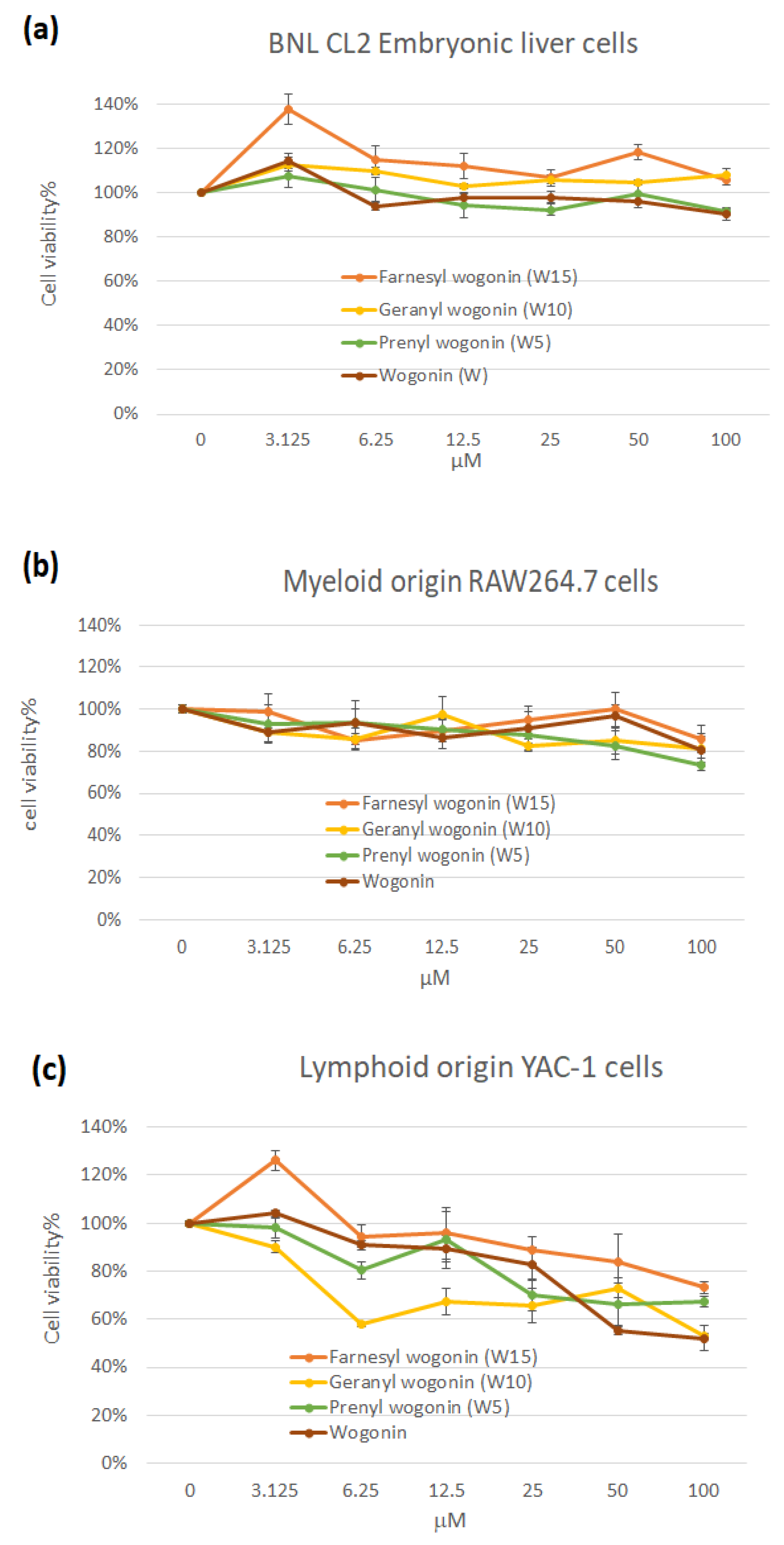
P values of the extending terpenyl moieties. A dose-dependently selective immunotoxicity in lymphoid origin YAC-1 cells but not myeloid (macrophage RAW264.7) and somatic (embryonic liver cells BNL CL2) origins are demonstrated (
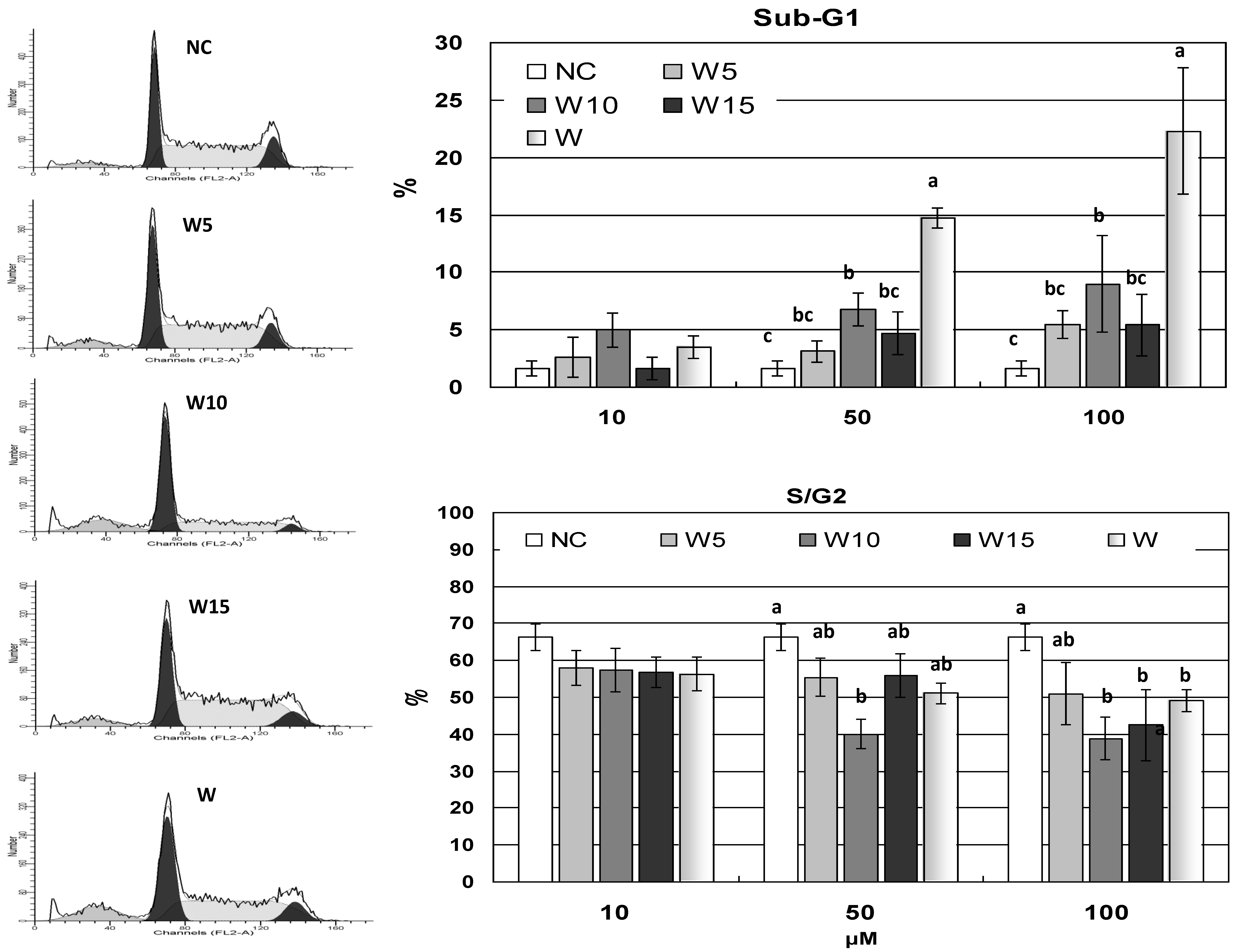
Figure 1). Nevertheless, the overall cell livability of YAC-1 cells remained above 50% at concentrations less than 100 μM of all treatments. The increased lipophilic property of terpenylated wogonin derivatives has not departed from wogonin in cytotoxicity. To further follow these results, cell cycle analysis on YAC-1 cells demonstrates that wogonin at higher concentrations of 50 and 100 μM significantly induced apoptosis by elevating sub-G1 proportion (
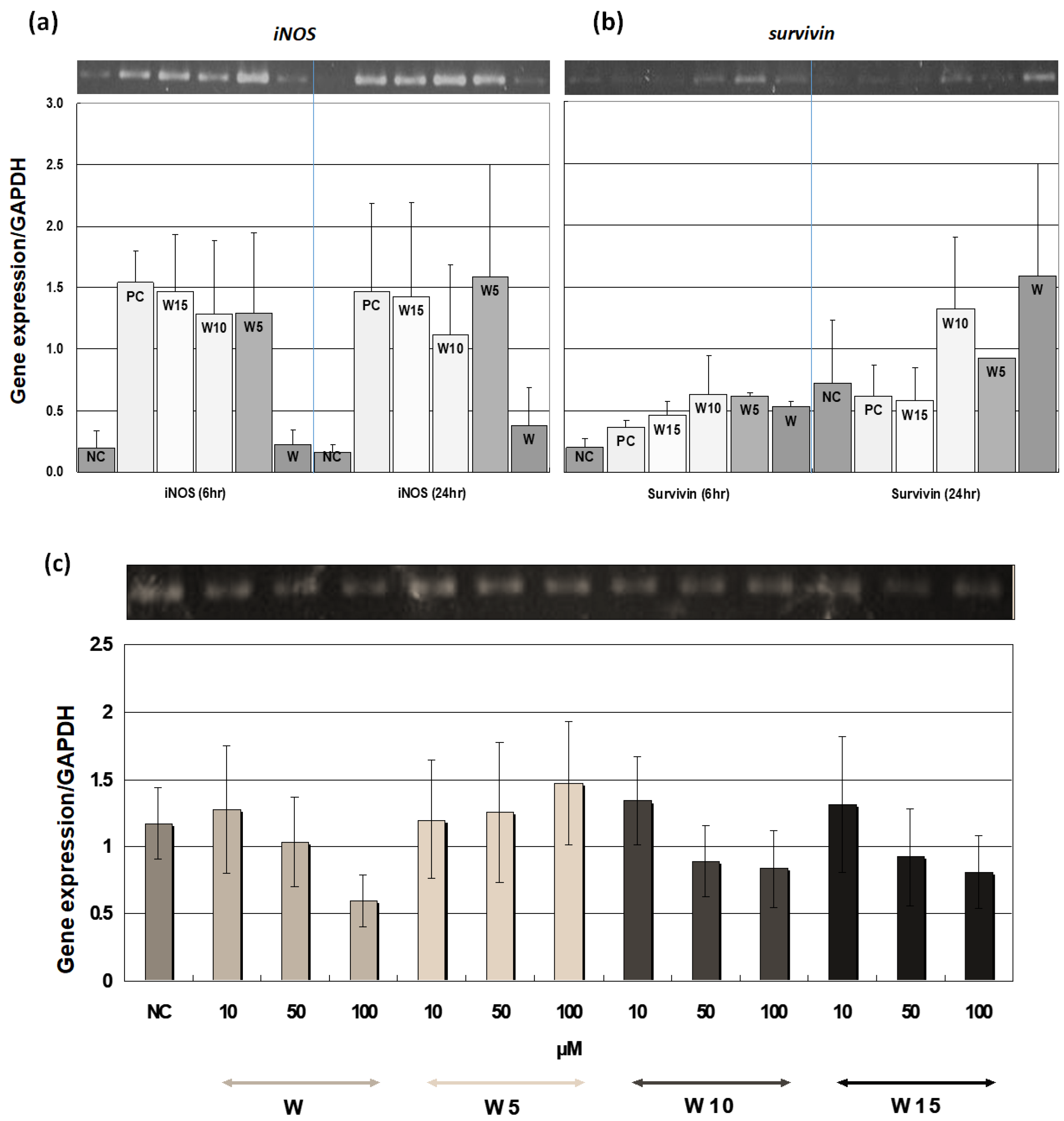
Figure 5). Wogonin and geranyl wogonin have also reduced the S/G2 proportions, while prenyl wogonin exhibits relatively less impact on the cell cycle, which is in agreement with the finding of sustained mitotic survivin gene expression of W5 (
Figure 6c). Wogonin has been shown to preferentially induce malignant lymphocyte apoptosis while no toxicity to normal lymphocytes [
24]. It is worth noting that YAC-1 is a cancer cell line that may not well-represent normal lymphoblastogenic lymphocytes. Antigens activate selected lymphocyte clonal proliferation and facilitate fundamental steps in acquired immunity. Hampered lymphocyte proliferation has been shown to disrupt both cell-mediated and humoral immunities. Terpenylated wogonin derivatives exhibit less impact in inducing apoptosis of proliferating lymphoid origin YAC-1 cells might imply additional advantages besides the well-recognized anti-cancer property of wogonin.
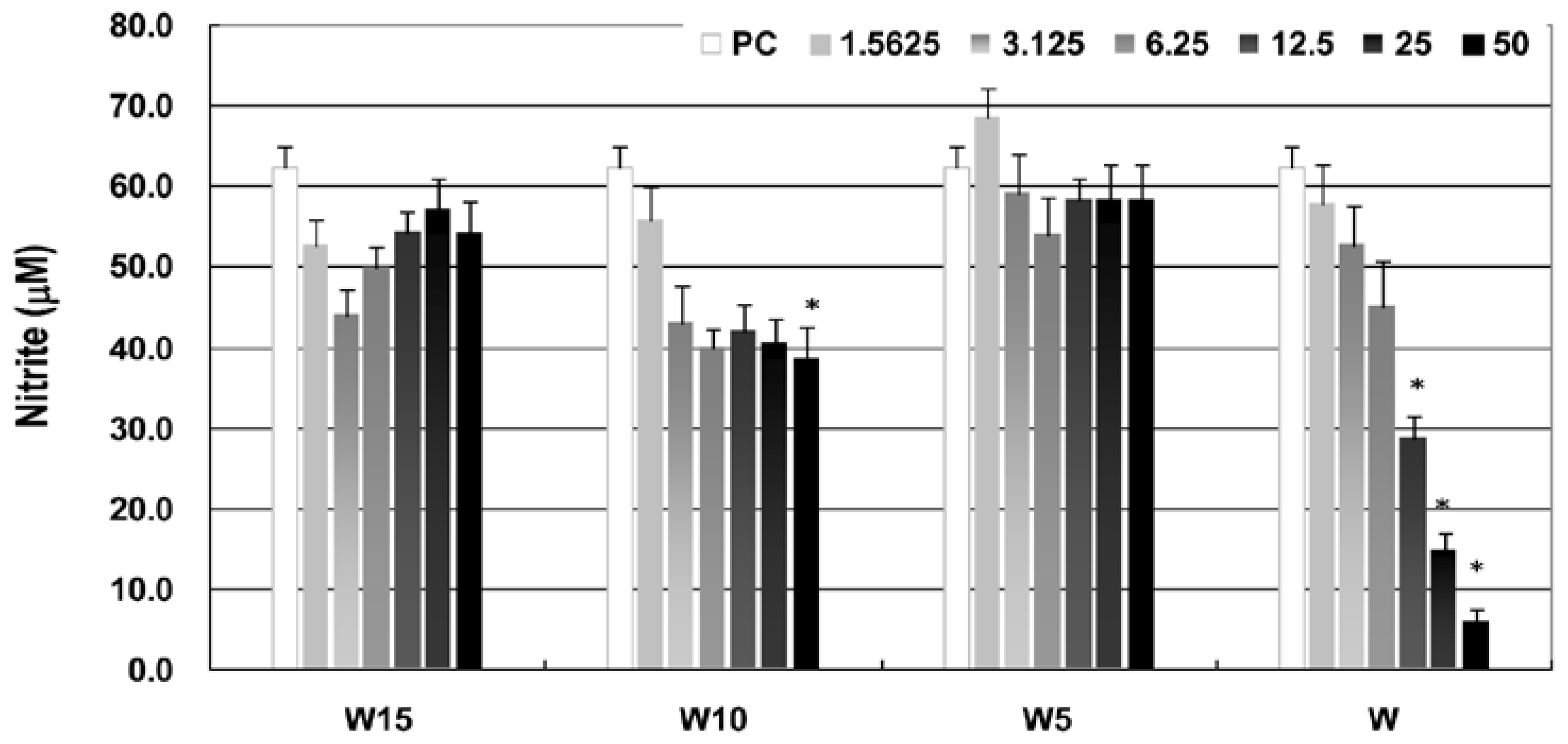
Moreover, the anti-inflammatory property as assessed by nitrite concentration of LPS-activated macrophages in wogonin and terpenylated wogonin derivatives shows different degrees of NO (nitric oxide) suppressions (
Figure 3), and wogonin exhibits a potent anti-inflammatory property in a dose-dependent manner at concentrations up to 50 μM. At similar concentration levels, Lee and Park [
1] have also demonstrated inhibitions of NO and other pro-inflammatory and inflammatory cytokines in polyinosinic-poly-cytidylic acid-induced RAW264.7 macrophages. Compared with wogonin, terpenylated wogonin derivatives show moderate or even no inhibitions on NO productions. Interestingly, at lower concentration levels (1.5625 to 6.25 μM), geranyl wogonin exhibits even faster-depressing NO production than wogonin, and the suppression has leveled at higher concentrations up to 50 μM. Moreover, while geranyl wogonin (W10) has significantly (
p < 0.05) inhibited NO production at a concentration of 50 μM as compared to the positive control (PC) of medium only of LPS-activated macrophages, prenyl (W5) and farnesyl (W15) wogonin exhibit no inhibitions on NO productions. It is worth mentioning that while W5 and W15 have reduced NO inhibition capacity, they exhibit better maintaining intracellular ROS levels than wogonin in LPS-activated macrophages, as seen in
Figure 4. Moreover, coincidently parallel to the effect on reducing cell livability of YAC-1 cells (
Figure 1c), the reduction was leveled at higher concentrations of terpenylated wogonin, particularly of geranyl wogonin. Structure-functional activities of flavonoids with isoprenoid chains have been reviewed pharmacologically in favor of increasing affinity to biological membranes [
25]. Indeed, flavonoids such as wogonin are chemically amphipathic molecules of both hydrophobic and hydrophilic moieties. Substitution of the hydrophilic hydroxyl (OH) group may reduce the hydrophilic property, alternatively increasing hydrophobic interaction with the cell membrane. The number of isoprene units alkylated to wogonin may add a lipophilic property or micelle-forming property, which may differentially affect the membrane integration or retention in the lipid bilayer of the prenyl, geranyl, or farnesyl wogonin at specific niches. Furthermore, it is important to mention that although they are both biological membranes, cell membrane and nucleus membrane fundamentally evolved differently in the eukaryotes. The former is a single lipid bilayer, and the later nucleus membrane is doubled. As the result of
iNOS gene expression demonstrated (
Figure 6a), it is reasonable to suspect that the different number of isoprene units may serve differently in integration with the two distinct biological membranes. Certainly, the diverse natural occurrences of terpenylated flavonoids with
C-prenylated or
O-prenylated conjugates [
18] having various combinations of terpenyl length, conjugated position, number of terpenyl conjugates plus the diversified terpenoids, which possess independent biological roles to add more interesting questions await for further elucidations.
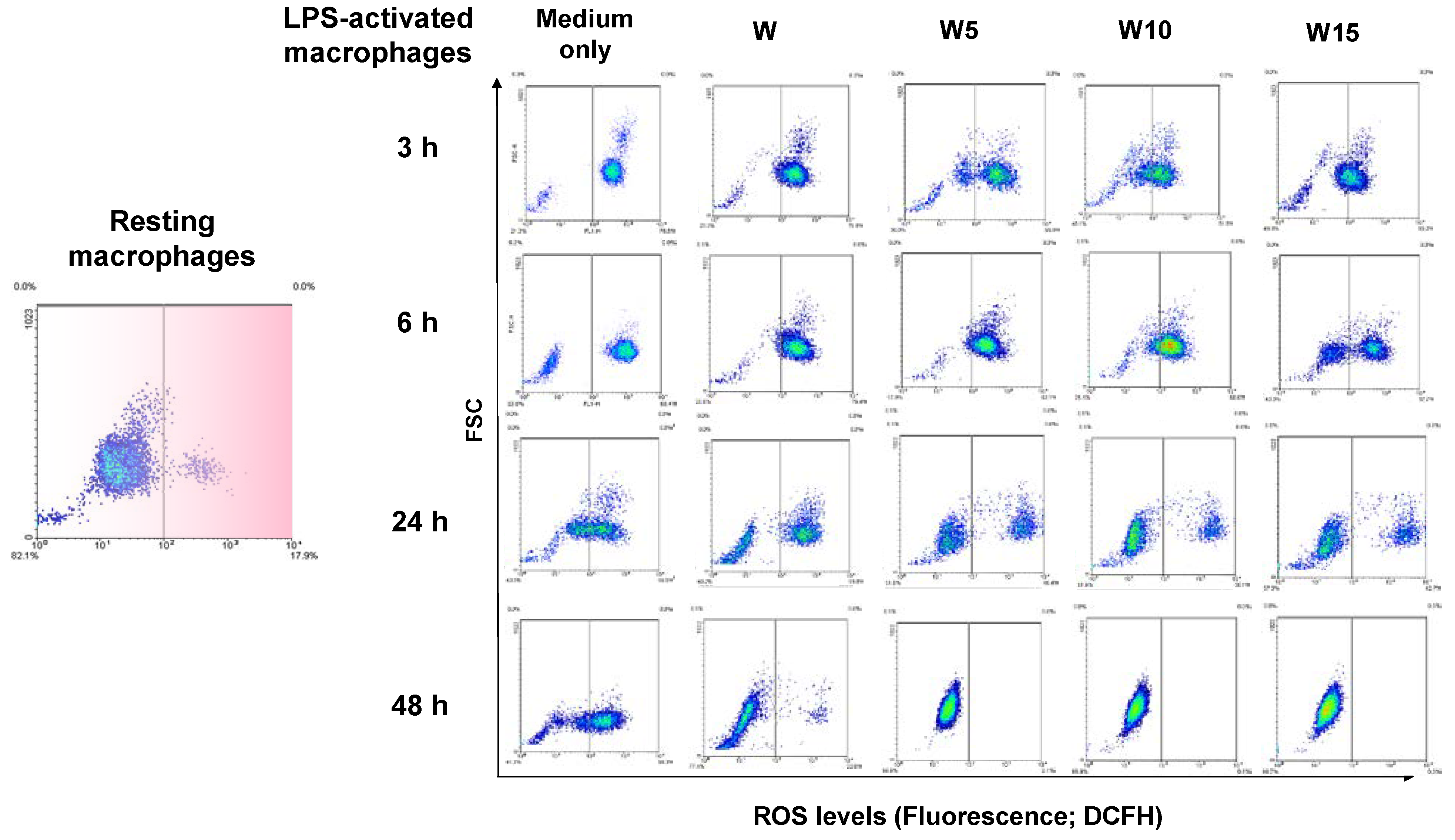
Furthermore, the antioxidative activities of wogonin and its terpenylated derivatives as assessed by flow cytometric analysis of intracellular ROS level of LPS-activated macrophages at 3, 6, 24, and 48 h (
Figure 4) have revealed the merits of terpenylated wogonin in harnessing elevating ROS over the wogonin. Prenylated flavonoids naturally isolated from Heartwood of
Artocarpus communis [
26] or bee propolis [
20] show potent antioxidative and anti-inflammatory activities. Currently, LPS-activated macrophages have almost immediately increased intracellular ROS level since 3 h, maximized at 6 h, and slowly back to resting ROS level after 48 h. As the cell density plot demonstrated, a time-dependent and isoprene unit number-dependent manner on shifting the cell density from right to left is evidenced. As discussed earlier, the higher clog
P values of the 7-
O-substituted wogonin derivatives implied higher lipophilicity. By improving membrane affinity, integration, and possible permeability, the structure-function-related intracellular ROS scavenging is attained in terpenylated wogonin. Moreover, the geranyl wogonin (W10) exhibits the best controlling elevating intracellular ROS level of LPS agitated macrophages as shown by a fast recovery to basal ROS level over time when compared with the other treatments. Nevertheless, wogonin exhibits anti-cancer effects by up-regulating intracellular ROS levels to achieve cytotoxicity and apoptosis [
17,
27,
28,
29]. The disparity of wogonin in inducing ROS generation in cancer cells or suppression of ROS levels in LPS-activated macrophages implies its complicated role in regulating intracellular ROS levels. The 7-
O-terpenylated wogonin with different number units of isoprene seems to differentially exhibit functions in the current immune characterizations. Furthermore, although the prenol, geraniol, and farnesol have been tested with no toxicity to current tested cell lines, these terpenes may independently exert potent anti-tumor and anti-inflammation activities [
30,
31]. Since the intracellular dynamic of terpenylated wogonin has not been tracked, there is no reason to not believe synergistic effects may occur sometime during the extended culture. Although it is still controversial, the terpenes’ antioxidative roles in cells have been reviewed, and isoprene may exhibit multiple regulatory functions in plants other than scavenging free radicals [
32]. The current study demonstrated the immunological advantages of chemically synthesized 7-
O-terpenylated wogonin, particularly geranyl wogonin may act with a lower effective dosage, prolonged action, and better preserves livability and functions of immune cells of lymphoid and myeloid cells origins. In addition, the current data may also support the biological importance of naturally diverse terpenylated flavonoids, which may pose new indications in future drug discovery and applications.












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}