
In Vitro and In Vivo Assessments of Anti-Hyperglycemic Properties of Soybean Residue Fermented with Rhizopus oligosporus and Lactiplantibacillus plantarum

 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Soybean Residue Fermentation
2.1.1. Sources of Soybean Residue and Microbes
2.1.2. Solid-State Fermentation of Soybean Residues
2.2. Content Analysis
2.2.1. Extraction of Isoflavones and GABA
2.2.2. Analysis of Isoflavones
2.2.3. Analysis of GABA
2.3. In Tube Assessment
2.3.1. DPPH Assay
2.3.2. ABTS Assay
2.3.3. Fe(II) Ion Chelating Activity
2.4. In Vitro Assessment
2.4.1. Cell Line and Cell Culture
2.4.2. Cell Viability
2.4.3. Intracellular Reactive Oxygen Species Level
2.5. In Vivo Assessment
2.5.1. Animals and Treatment Preparations
2.5.2. Blood Biochemical Measurements
2.5.3. Blood Glucose Level Monitoring and Glucose Tolerance Test
2.5.4. Kidney Histopathological Examination
2.6. Statistics
3. Results
3.1. Characteristics of Fermented Soybean Residue and In Vitro Assessments
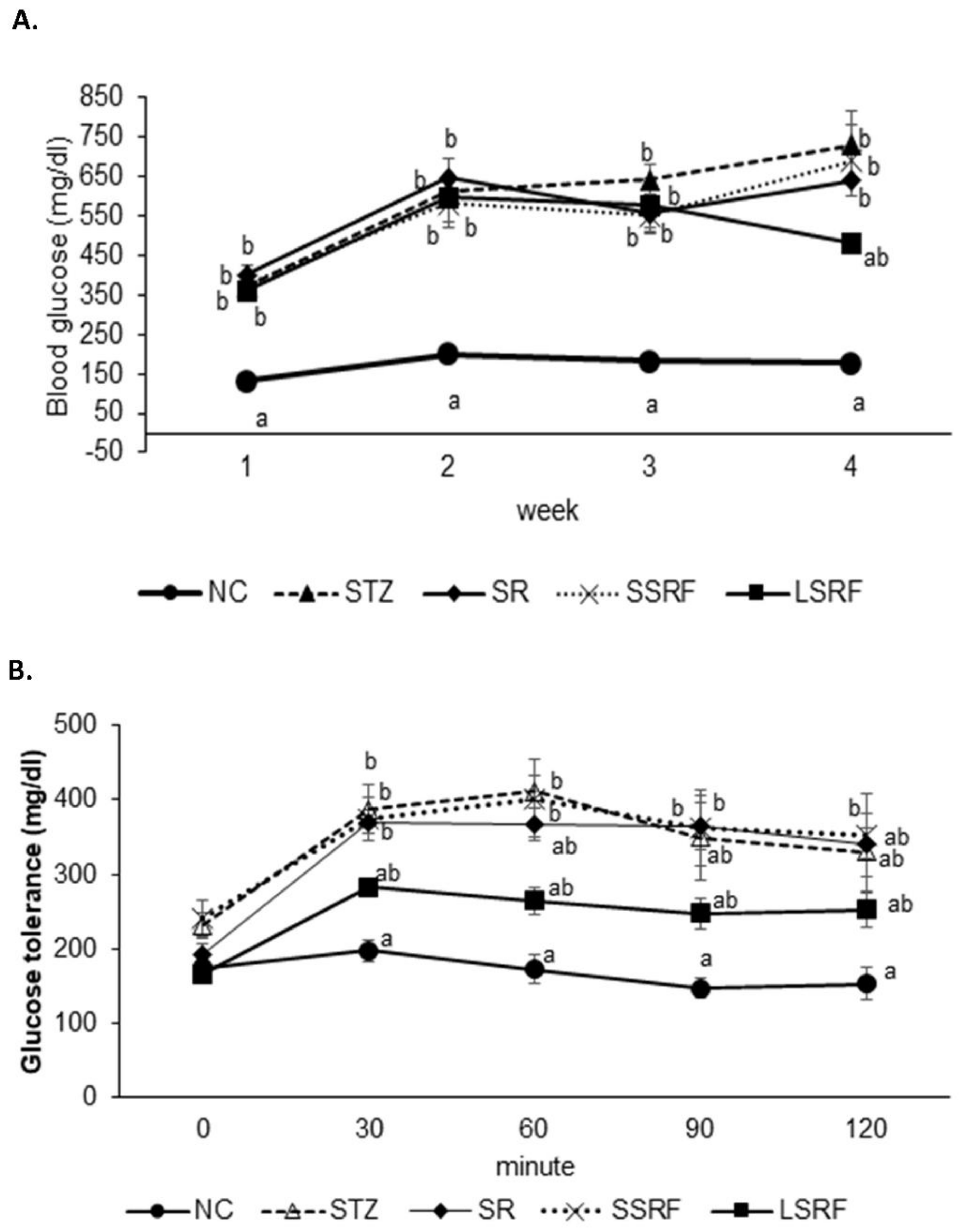
3.2. In Vivo Assessments of Fermented Soybean Residue Supplementation in STZ-Induced Diabetic Mice
3.2.1. Improved Blood Biochemical Profile
3.2.2. Ameliorated Blood Glucose Homeostasis
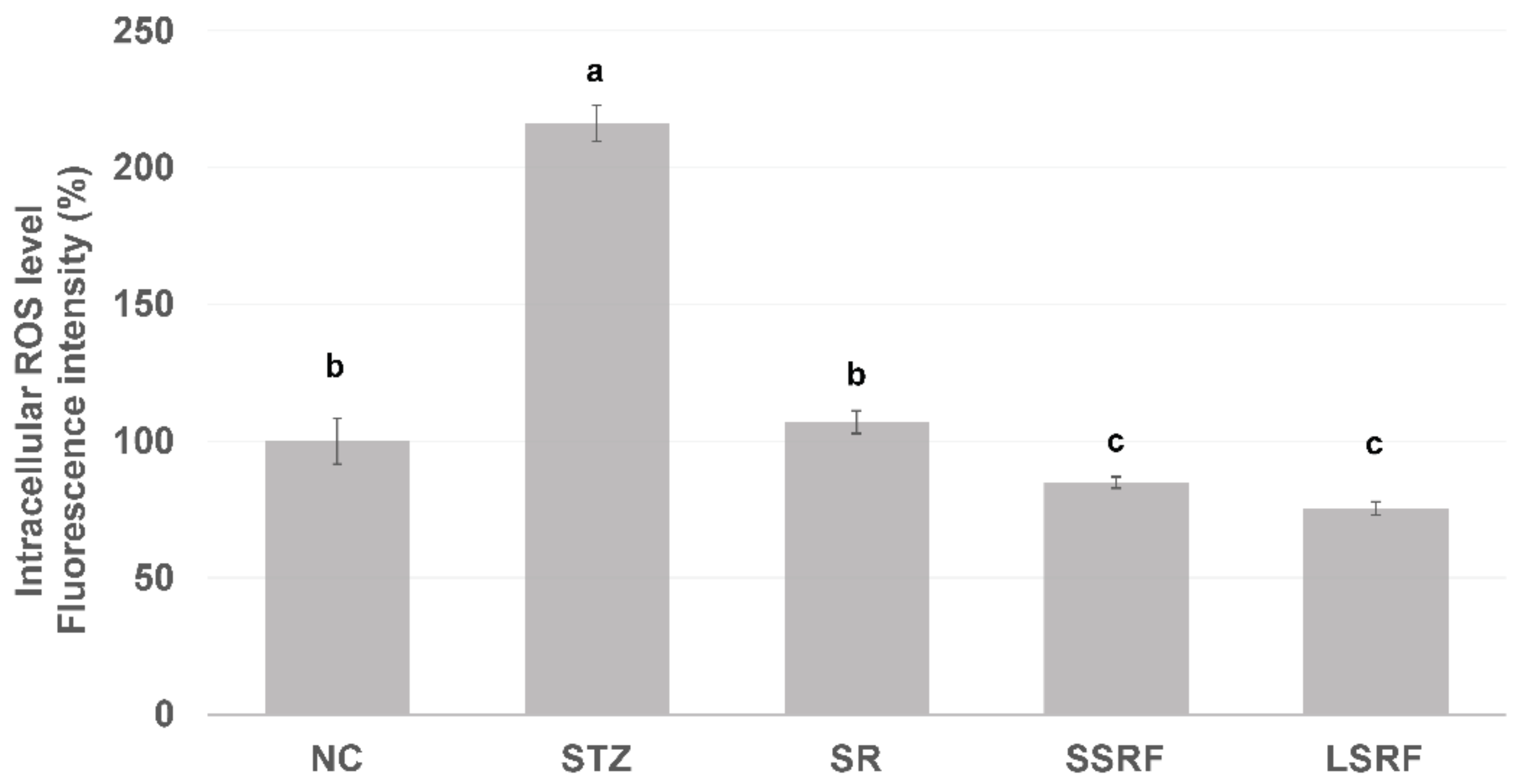
3.2.3. Normalized Intracellular ROS Level in Leukocytes
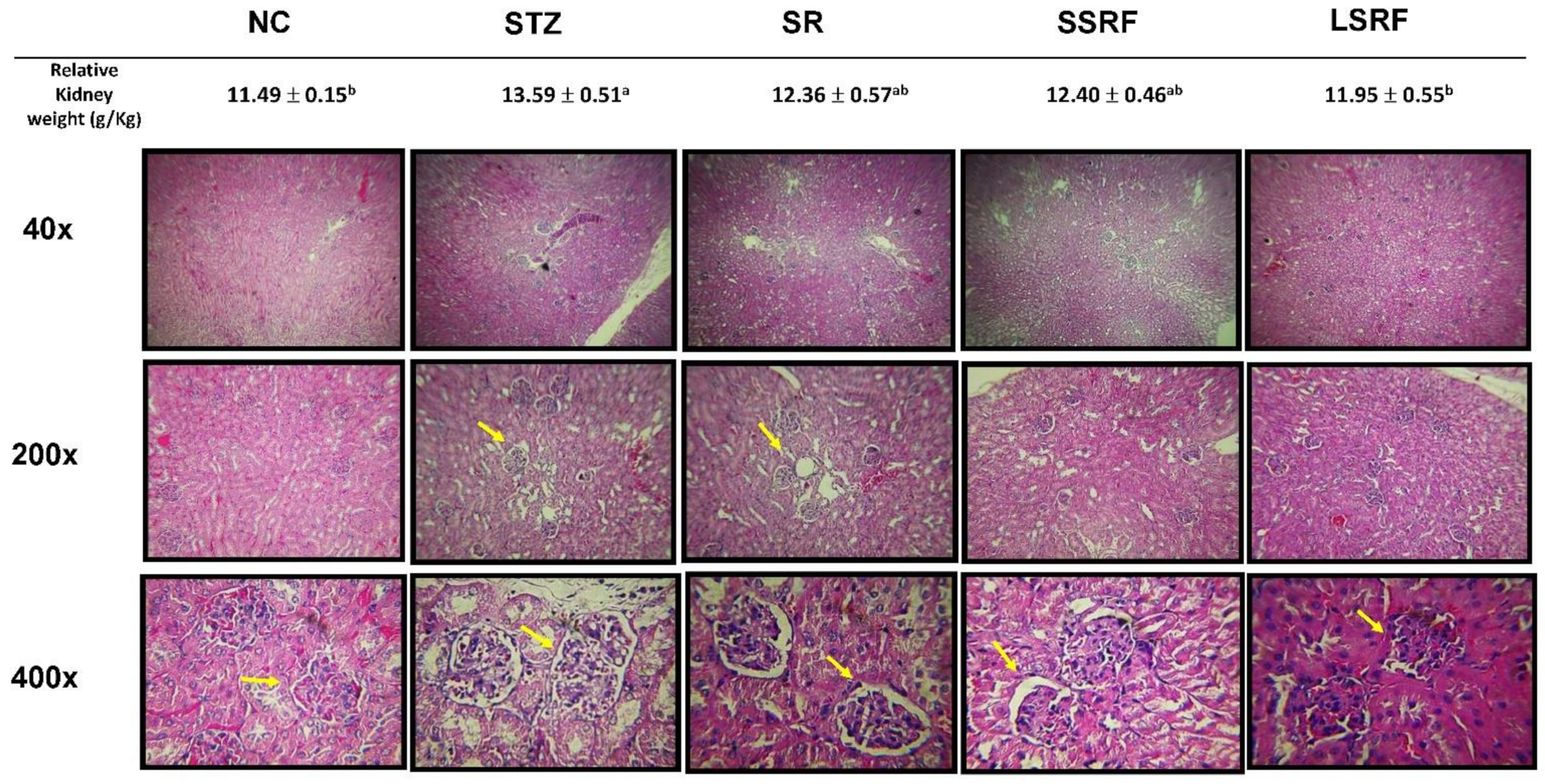
3.3. Protection of Hyperglycemic Mice as Evidenced by Gross Anatomy and Histopathological Evaluations in Kidney
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gilbert, R.E.; Liu, D. Anti-Diabetic Functions of Soy Isoflavone Genistein: Mechanisms Underlying Its Effects on Pancreatic β-Cell Function. Food Funct. 2013, 4, 200–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Yang, S. Chapter 18-Solid State Fermentation and Its Applications. In Bioprocessing for Value-Added Products from Renewable Resources; Yang, S.-T., Ed.; Elsevier: Amsterdam, The Netherlands, 2007; pp. 465–489. [Google Scholar] [CrossRef]
- Hachmeister, K.A.; Fung, D.Y.C. Tempeh: A Mold-Modified Indigenous Fermented Food Made from Soybeans and/or Cereal Grains. Critical Reviews in Microbiology (USA). 1993. Available online: https://scholar.google.com/scholar_lookup?title=Tempeh%3A+a+mold-modified+indigenous+fermented+food+made+from+soybeans+and%2For+cereal+grains&author=Hachmeister%2C+K.A.&publication_year=1993 (accessed on 1 October 2021).
- Małgorzata, W.; Honke, J.; Konrad, M.P. Effect of Solid-State Fermentation with Rhizopus Oligosporus on Bioactive Compounds and Antioxidant Capacity of Raw and Roasted Buckwheat Groats. Ital. J. Food Sci. 2015, 27, 424–431. [Google Scholar] [CrossRef]
- Achirawit, N.; Kaewkod, T.; Konkit, M.; Tragoolpua, Y.; Bovonsombut, S.; Chitov, T. Characterisation of Lactobacillus Plantarum of Dairy-Product Origin for Probiotic Chèvre Cheese Production. Foods 2022, 11, 934. [Google Scholar] [CrossRef]
- Liu, Y.-W.; Liong, M.T.; Tsai, Y.-T. New Perspectives of Lactobacillus Plantarum as a Probiotic: The Gut-Heart-Brain Axis. J. Microbiol. 2018, 56, 601–613. [Google Scholar] [CrossRef]
- Sahab, N.R.M.; Subroto, E.; Roostita, L.B.; Gemilang, L.U. γ-Aminobutyric Acid Found in Fermented Foods and Beverages: Current Trends. Heliyon 2020, 6, e05526. [Google Scholar] [CrossRef]
- Soltani, N.; Qiu, H.; Aleksic, M.; Glinka, Y.; Zhao, F.; Liu, R.; Li, Y. GABA Exerts Protective and Regenerative Effects on Islet Beta Cells and Reverses Diabetes. Proc. Natl. Acad. Sci. USA 2011, 108, 11692–11697. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.-C.; Hsieh, S.-L.; Hu, C.-Y. Effects of Red-Bean Tempeh with Various Strains of Rhizopus on GABA Content and Cortisol Level in Zebrafish. Microorganisms 2020, 8, 1330. [Google Scholar] [CrossRef]
- Chen, B.-H.; Hsieh, C.-W.; Chen, L.-G.; Chu, C.-S.; Wang, S.-H. The changes of isoflavones and GABA in soymilk fermented with an acid-dormant mutant of Lactobacillus plantarum. J. Agric. For. 2016, 13, 15–27. [Google Scholar]
- World Health Organization. Global Report on Diabetes; WHO Library: Geneva, Switzerland, 2016; ISBN 9789241565257. [Google Scholar]
- Groop, P.-H.; Thomas, M.C.; Moran, J.L.; Wadèn, J.; Thorn, L.M.; Ville-Petteri, M.; Rosengård-Bärlund, M.; Saraheimo, M.; Hietala, K.; Heikkila, O.; et al. The Presence and Severity of Chronic Kidney Disease Predicts All-Cause Mortality in Type 1 Diabetes. Diabetes 2009, 58, 1651–1658. [Google Scholar] [CrossRef] [Green Version]
- Alshehri, M.M.; Sharifi-Rad, J.; Bravo, J.; Jara, E.L.; Salazar, L.A.; Kregiel, D.; Uprety, Y.; Akram, M.; Iqbal, M.; Martorell, M.; et al. Therapeutic Potential of Isoflavones with an Emphasis on Daidzein. Oxid. Med. Cell. Longev. 2021, 2021, 6331630. [Google Scholar] [CrossRef]
- Sharif, M.S.; Fan, H.; Wang, S.; Yu, H.; Piao, C. Prebiotic Impacts of Soybean Residue (Okara) on Eubiosis/Dysbiosis Condition of the Gut and the Possible Effects on Liver and Kidney Functions. Molecules 2021, 26, 326. [Google Scholar] [CrossRef]
- Eizirik, D.L.; Gurzov, E.N. Can GABA Turn Pancreatic α-Cells into β-Cells? Nature Reviews. Endocrinology 2018, 14, 629–630. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.-J.; Kim, H.S.; Lim, S.T.; Chagam, K.R. Enhanced Accumulation of Gamma-Aminobutyric Acid in Rice Bran Using Anaerobic Incubation with Various Additives. Food Chem. 2019, 271, 187–192. [Google Scholar] [CrossRef]
- Chang, C.-T.; Hsu, C.-K.; Chou, S.-T.; Chen, Y.-C.; Huang, F.-S.; Chung, Y.-C. Effect of Fermentation Time on the Antioxidant Activities of Tempeh Prepared from Fermented Soybean Using Rhizopus Oligosporus. Int. J. Food Sci. Technol. 2009, 44, 799–806. [Google Scholar] [CrossRef]
- Santos, Q.; Vidiany, A.; Nascimento, C.G.; Schmidt, C.A.P.; Mantovani, D.; Dekker, R.F.H.; da Cunha, M.A.A. Solid-State Fermentation of Soybean Okara: Isoflavones Biotransformation, Antioxidant Activity and Enhancement of Nutritional Quality. LWT 2018, 92, 509–515. [Google Scholar] [CrossRef]
- Chen, L.-G.; Chang, C.; Tsay, J.; Weng, B.B. Hepatoprotective effects of litchi (Litchi chinensis) procyanidin A2 on carbon tetrachloride-induced liver injury in ICR mice. Exp. Ther. Med. 2017, 13, 2839–2847. [Google Scholar] [CrossRef] [Green Version]
- Jankowiak, L. Separation of Isoflavones Form Okara: Process Mechanisms & Synthesis. Ph.D. Thesis, Wageningen University, Wageningen, The Netherlands, 2014. [Google Scholar]
- Vong, W.C.; Liu, S.-Q. Biovalorisation of Okara (Soybean Residue) for Food and Nutrition. Trends Food Sci. Technol. 2016, 52, 139–147. [Google Scholar] [CrossRef]
- Wang, H.; Murphy, P.A. Isoflavone Content in Commercial Soybean Foods. J. Agric. Food Chem. 1994, 42, 1666–1673. [Google Scholar] [CrossRef]
- Song, T.; Barua, K.; Buseman, G.; Murphy, P.A. Soy Isoflavone Analysis: Quality Control and a New Internal Standard. Am. J. Clin. Nutr. 1998, 68 (Suppl. 6), 1474S–1479S. [Google Scholar] [CrossRef] [Green Version]
- Kamble, D.B.; Rani, S. Bioactive Components, in Vitro Digestibility, Microstructure and Application of Soybean Residue (Okara): A Review. Legume Sci. 2020, 2, e32. [Google Scholar] [CrossRef]
- McCue, P.; Akiyo, H.; Kalidas, S. Solid-State Bioconversion of Phenolic Antioxidants from Defatted Soybean Powders by Rhizopus Oligosporus: Role of Carbohydrate-Cleaving Enzymes. J. Food Biochem. 2003, 27, 501–514. [Google Scholar] [CrossRef]
- Feng, X.M.; Anders, R.B.; Eriksson, J.; Schnürer, J. Growth of Lactic Acid Bacteria and Rhizopus Oligosporus during Barley Tempeh Fermentation. Int. J. Food Microbiol. 2005, 104, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Arena, M.P.; Silvain, A.; Normanno, G.; Grieco, F.; Drider, D.; Spano, G.; Fiocco, D. Use of Lactobacillus plantarum Strains as a Bio-Control Strategy against Food-Borne Pathogenic Microorganisms. Front. Microbiol. 2016, 7, 464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasquale, R.; Fares, C.; Longo, A.; Spano, G.; Capozzi, V. Lactobacillus plantarum with Broad Antifungal Activity as a Protective Starter Culture for Bread Production. Foods 2017, 6, 110. [Google Scholar] [CrossRef] [Green Version]
- Chang, H.-Y.; Ho, Y.; Sheu, M.; Lin, Y.; Tseng, M.; Wu, S.; Huang, G.; Chang, Y. Antioxidant and Free Radical Scavenging Activities of Phellinus Merrillii Extracts. Bot. Stud. 2007, 48, 11. [Google Scholar]
- Egbune, E.; Ezedom, T.; Anigboro, A.; Aganbi, E.; Amata, A.; Tonukari, N. Antioxidants and antigenotoxic properties of Rhizopus oligosporus fermented cassava (Manihot esculenta Crantz). Afr. J. Biochem. Res. 2022, 16, 39–46. [Google Scholar] [CrossRef]
- Schaefer, M.J. The Phytic Acid Trace Mineral Content of Soybeans and Their Effects on Tofu Texture. Ph.D. Thesis, Iowa State University, Ames, IA, USA, 1986. Available online: https://core.ac.uk/download/pdf/38899165.pdf (accessed on 1 October 2021).
- Buckle, K.A. Phytic Acid Changes in Soybeans Fermented by Traditional Inoculum and Six Strains of Rhizopus Oligosporus. J. Appl. Bacteriol. 1985, 58, 539–543. [Google Scholar] [CrossRef]
- Adeyemo, S.M.; Onilude, A.A. Enzymatic Reduction of Anti-Nutritional Factors in Fermenting Soybeans by Lactobacillus Plantarum Isolates from Fermenting Cereals. Niger. Food J. 2013, 31, 84–90. [Google Scholar] [CrossRef] [Green Version]
- Borges, C.; Chong, W.; Carrão-Panizzi, M.C.; Mandarino, J.M.G.; da Silva, J.B.; Benedetti, S.; Ida, E.I. Contents and Bioconversion of β-Glycoside Isoflavones to Aglycones in the Processing Conditions of Soybean Tempeh. Pesq. Agropecuária Bras. 2016, 51, 271–279. [Google Scholar] [CrossRef] [Green Version]
- Kolb, H. Mouse Models of Insulin Dependent Diabetes: Low-Dose Streptozocin-Induced Diabetes and Nonobese Diabetic (NOD) Mice. Diabetes Metab. Rev. 1987, 3, 751–778. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Mi, S.; Du, L.; Li, X.; Li, P.; Jia, K.; Zhao, J.; Zhang, H.; Zhao, W.; Gao, Y. The Associations between Plasma Phytoestrogens Concentration and Metabolic Syndrome Risks in Chinese Population. PLoS ONE 2018, 13, e0194639. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
 |  |  |  |  | |
|---|---|---|---|---|---|
| Fermentation Condition & Antioxidative Capacity | SR Soy Residues | RSF R. oligosporus 24 h | RLF R. oligosporus 48 h | RLSF R. oligosporus + L. plantarum Short Fermentation 24 h | RLLF R. oligosporus + L. plantarum Long Fermentation 48 h |
| Moisture | 60.8 a | 50.4 c | 55.6 ab | 44.6 c | 56.5 ab |
| Temperature | 26.3 d | 35.7 c | 43.3 b | 35.2 c | 45.7 a |
| pH | 5 | 6 | 6 | 6 | 5 |
| DPPH% | 13.8 c | 51.9 a | 53.9 a | 47.9 ab | 44.3 b |
| ABTS% | 3.62 c | 43.1 a | 46.7 a | 14.0 b | 10.9 b |
| Fe(II) Chelating% | 5.86 d | 42.7 a | 44.6 a | 39.1 ab | 19.2 c |
| Contents of Functional Chemicals (μg/g w/w) | |||||
|---|---|---|---|---|---|
| Item (mg/g) | SR Soy Residues | RSF R. oligosporus 24 h | RLF R. oligosporus 48 h | RLSF R. oligosporus + L. plantarum 24 h | RLLF R. oligosporus + L. plantarum 48 h |
| Genistin | ND | ND | ND | ND | ND |
| glycitin | ND | ND | ND | ND | ND |
| daidzin | ND | ND | ND | ND | ND |
| Genistein (aglycone) | 17.0 | 20.1 | 20.7 | 22.2 | 19.4 |
| Glycitein (aglycone) | ND | ND | ND | ND | ND |
| Daidzein (aglycone) | ND | 21.4 | 24.5 | 24.6 | 26.8 |
| GABA | ND | 0.3154 | 0.6387 | 0.2676 | 0.6762 |
| In vitro assessments on HG-MDCK cells | |||||
| Cell viability (%) | 118 b | 121 b | 103 b | 109 b | 129 a |
| Intracellular ROS (%) | 104 a | 88.9 c | 87.5 c | 91.9 b | 93.2 b |
| Items | Treatments | ||||
|---|---|---|---|---|---|
| NC | STZ | SR | SSRF | LSRF | |
| HbA1c (mg/dL) | 2.9 ± 0.12 a | 3.8 ± 0.19 b | 3.9 ± 0.25 b | 4.2 ± 0.73 b | 3.8 ± 0.18 b |
| GOT (IU/L) | 41 ± 5 a | 177 ± 14 b | 96.6 ± 6 c | 53.4 ± 5 c | 61.6 ± 10 c |
| GPT (IU/L) | 15.4 ± 3.9 a | 89 ± 12 b | 33.6 ± 3.7 ab | 26.6 ± 3.9 ac | 38 ± 7.3 c |
| T-Protein (IU/L) | 1.7 ± 0.3 a | 1.3 ± 0.1 b | 1.5 ± 0.08 ab | 1.5 ± 0.17 ab | 1.6 ± 0.11 ac |
| T-Bilirubin (IU/L) | 0.208 ± 0.01 a | 0.244 ± 0.01 b | 0.226 ± 0.01 ab | 0.218 ± 0.01 ac | 0.214 ± 0.01 ac |
| BUN (IU/L) | 6.3 ± 0.60 a | 8.1 ± 0.70 b | 6.6 ± 0.60 ac | 6.5 ± 0.17 ac | 6.4 ± 0.30 ac |
| T-Cholesterol (IU/L) | 22.4 ± 5 a | 30.2 ± 2.3 b | 24.8 ± 2.5 ab | 24.4 ± 0.5 a | 25 ± 0.7 ab |
| T-Glyceride (IU/L) | 8.4 ± 1.8 a | 20 ± 4.8 b | 16.2 ± 3.1 bc | 13.8 ± 2.6 ac | 10.4 ± 1.3 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hariyanto, I.; Hsieh, C.-W.; Hsu, Y.-H.; Chen, L.-G.; Chu, C.; Weng, B.B.-C. In Vitro and In Vivo Assessments of Anti-Hyperglycemic Properties of Soybean Residue Fermented with Rhizopus oligosporus and Lactiplantibacillus plantarum. Life 2022, 12, 1716. https://doi.org/10.3390/life12111716
Hariyanto I, Hsieh C-W, Hsu Y-H, Chen L-G, Chu C, Weng BB-C. In Vitro and In Vivo Assessments of Anti-Hyperglycemic Properties of Soybean Residue Fermented with Rhizopus oligosporus and Lactiplantibacillus plantarum. Life. 2022; 12(11):1716. https://doi.org/10.3390/life12111716
Chicago/Turabian StyleHariyanto, Istiqomah, Chia-Wen Hsieh, Yueh-Han Hsu, Lih-Geng Chen, ChiShih Chu, and Brian Bor-Chun Weng. 2022. "In Vitro and In Vivo Assessments of Anti-Hyperglycemic Properties of Soybean Residue Fermented with Rhizopus oligosporus and Lactiplantibacillus plantarum" Life 12, no. 11: 1716. https://doi.org/10.3390/life12111716