
Role of Matrix Gla Protein in the Complex Network of Coronary Artery Disease: A Comprehensive Review

 ,
,  ,
, 
Abstract
:1. Introduction
2. Pathophysiology of Vascular Calcification, Arterial Stiffness and Their Interrelation
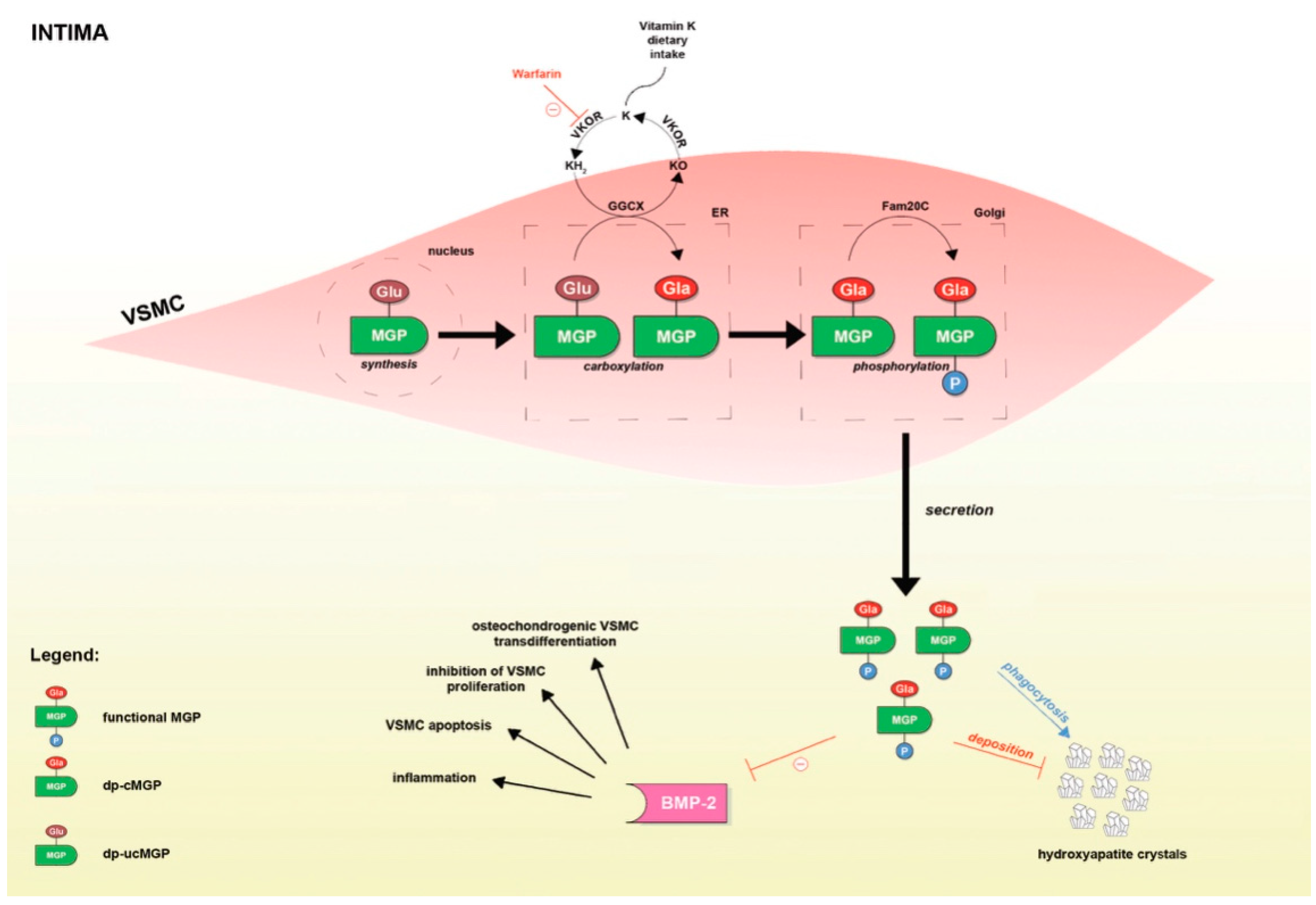
3. MGP and Its Conformations
4. MGP in Coronary Artery Disease (CAD)
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BMP | bone morphogenic protein |
| ALP | alkaline phosphatase |
| FGF-23 | fibroblast growth factor-23 |
| Runx2 | RUNX family transcription factor 2 |
| PDK4 | pyruvate dehydrogenase kinase 4 |
| MGP | matrix Gla protein |
| VSMC | vascular smooth muscle cell |
| BMP-2 | bone morphogenetic protein-2 |
| GGCX | gamma-glutamyl carboxylase |
| VKOR | vitamin K epoxide reductase |
| KH2 | vitamin KH2 |
| KO | vitamin KO |
| Fam20C | family with sequence similarity 20, member C |
| dp-cMGP | dephosphorylated-carboxylated MGP |
| dp-ucMGP | dephosphorylated-uncarboxylated MGP |
| ER | endoplasmic reticulum |
| HR | hazard ratio |
| RR | relative risk |
| CHD | coronary heart disease |
| CKD | chronic kidney disease |
| CVD | cardiovascular disease |
| CVM | cardiovascular mortality |
| DM | diabetes mellitus |
| eGFR | glomerular filtration rate estimated from serum creatinine |
| HF | heart failure |
| BMI | body mass index |
References
- Kumrić, M.; Tičinović Kurir, T.; Borovac, J.A.; Božić, J. The Role of Natural Killer (NK) Cells in Acute Coronary Syndrome: A Comprehensive Review. Biomolecules 2020, 10, 1514. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Lee, I.-K.; Jeon, J.-H. Vascular Calcification—New Insights into Its Mechanism. Int. J. Mol. Sci. 2020, 21, 2685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrett, H.; O’Keeffe, M.; Kavanagh, E.; Walsh, M.; O’Connor, E.M. Is Matrix Gla Protein Associated with Vascular Calcification? A Systematic Review. Nutrients 2018, 10, 415. [Google Scholar] [CrossRef] [Green Version]
- Criqui, M.H.; Knox, J.B.; Denenberg, J.O.; Forbang, N.I.; McClelland, R.L.; Novotny, T.E.; Sandfort, V.; Waalen, J.; Blaha, M.J.; Allison, M.A. Coronary Artery Calcium Volume and Density: Potential Interactions and Overall Predictive Value: The Multi-Ethnic Study of Atherosclerosis. JACC Cardiovasc. Imaging 2017, 10, 845–854. [Google Scholar] [CrossRef]
- Ciccone, M.M.; Bilianou, E.; Balbarini, A.; Gesualdo, M.; Ghiadoni, L.; Metra, M.; Palmiero, P.; Pedrinelli, R.; Salvetti, M.; Scicchitano, P.; et al. Task force on: Early markers of atherosclerosis: Influence of age and sex. J. Cardiovasc. Med. 2013, 14, 757–766. [Google Scholar] [CrossRef] [PubMed]
- Balietti, P.; Spannella, F.; Giulietti, F.; Rosettani, G.; Bernardi, B.; Cocci, G.; Bonfigli, A.R.; Sarzani, R. Ten-year changes in ambulatory blood pressure: The prognostic value of ambulatory pulse pressure. J. Clin. Hypertens. 2018, 20, 1230–1237. [Google Scholar] [CrossRef] [PubMed]
- Price, P.A.; Urist, M.R.; Otawara, Y. Matrix Gla protein, a new gamma-carboxyglutamic acid-containing protein which is associated with the organic matrix of bone. Biochem. Biophys. Res. Commun. 1983, 117, 765–771. [Google Scholar] [CrossRef]
- Roijers, R.B.; Debernardi, N.; Cleutjens, J.P.; Schurgers, L.J.; Mutsaers, P.H.; van der Vusse, G.J. Microcalcifications in early intimal lesions of atherosclerotic human coronary arteries. Am. J. Pathol. 2011, 178, 2879–2887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doherty, T.M.; Asotra, K.; Fitzpatrick, L.A.; Qiao, J.H.; Wilkin, D.J.; Detrano, R.C.; Dunstan, C.R.; Shah, P.K.; Rajavashisth, T.B. Calcification in atherosclerosis: Bone biology and chronic inflammation at the arterial crossroads. Proc. Natl. Acad. Sci. USA 2003, 100, 11201–11206. [Google Scholar] [CrossRef] [Green Version]
- Joshi, N.V.; Vesey, A.; Newby, D.E.; Dweck, M.R. Will 18F-sodium fluoride PET-CT imaging be the magic bullet for identifying vulnerable coronary atherosclerotic plaques? Curr. Cardiol. Rep. 2014, 16, 521. [Google Scholar] [CrossRef] [PubMed]
- Virmani, R.; Burke, A.P.; Farb, A.; Kolodgie, F.D. Pathology of the vulnerable plaque. J. Am. Coll. Cardiol. 2006, 47, C13–C18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehara, S.; Kobayashi, Y.; Yoshiyama, M.; Shimada, K.; Shimada, Y.; Fukuda, D.; Nakamura, Y.; Yamashita, H.; Yamagishi, H.; Takeuchi, K.; et al. Spotty calcification typifies the culprit plaque in patients with acute myocardial infarction: An intravascular ultrasound study. Circulation 2004, 110, 3424–3429. [Google Scholar] [CrossRef] [Green Version]
- Vengrenyuk, Y.; Carlier, S.; Xanthos, S.; Cardoso, L.; Ganatos, P.; Virmani, R.; Einav, S.; Gilchrist, L.; Weinbaum, S. A hypothesis for vulnerable plaque rupture due to stress-induced debonding around cellular microcalcifications in thin fibrous caps. Proc. Natl. Acad. Sci. USA 2006, 103, 14678–14683. [Google Scholar] [CrossRef] [Green Version]
- Raggi, P.; Shaw, L.J.; Berman, D.S.; Callister, T.Q. Prognostic value of coronary artery calcium screening in subjects with and without diabetes. J. Am. Coll. Cardiol. 2004, 43, 1663–1669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rennenberg, R.J.; Kessels, A.G.; Schurgers, L.J.; van Engelshoven, J.M.; de Leeuw, P.W.; Kroon, A.A. Vascular calcifications as a marker of increased cardiovascular risk: A meta-analysis. Vasc. Health Risk Manag. 2009, 5, 185–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grundy, S.M.; Stone, N.J. Coronary Artery Calcium: Where Do We Stand after over three Decades? Am. J. Med. 2021, S0002-9343, 291–296. [Google Scholar] [CrossRef]
- Simons, D.B.; Schwartz, R.S.; Edwards, W.D.; Sheedy, P.F.; Breen, J.F.; Rumberger, J.A. Noninvasive definition of anatomic coronary artery disease by ultrafast computed tomographic scanning: A quantitative pathologic comparison study. J. Am. Coll. Cardiol. 1992, 20, 1118–1126. [Google Scholar] [CrossRef] [Green Version]
- Rumberger, J.A.; Simons, D.B.; Fitzpatrick, L.A.; Sheedy, P.F.; Schwartz, R.S. Coronary artery calcium area by electron-beam computed tomography and coronary atherosclerotic plaque area. A histopathologic correlative study. Circulation 1995, 92, 2157–2162. [Google Scholar] [CrossRef]
- Rumberger, J.A.; Schwartz, R.S.; Simons, D.B.; Sheedy, P.F., 3rd; Edwards, W.D.; Fitzpatrick, L.A. Relation of coronary calcium determined by electron beam computed tomography and lumen narrowing determined by autopsy. Am. J. Cardiol. 1994, 73, 1169–1173. [Google Scholar] [CrossRef]
- Valenti, V.; Ó Hartaigh, B.; Heo, R.; Cho, I.; Schulman-Marcus, J.; Gransar, H.; Truong, Q.A.; Shaw, L.J.; Knapper, J.; Kelkar, A.A. A 15-Year Warranty Period for Asymptomatic Individuals without Coronary Artery Calcium: A Prospective Follow-Up of 9715 Individuals. JACC Cardiovasc. Imaging 2015, 8, 900–909. [Google Scholar] [CrossRef] [Green Version]
- Adelhoefer, S.; Uddin, S.M.I.; Osei, A.D.; Obisesan, O.H.; Blaha, M.J.; Dzaye, O. Coronary Artery Calcium Scoring: New Insights into Clinical Interpretation-Lessons from the CAC Consortium. Radiol. Cardiothorac. Imaging 2020, 2, e200281. [Google Scholar] [CrossRef]
- Sangiorgi, G.; Rumberger, J.A.; Severson, A.; Edwards, W.D.; Gregoire, J.; Fitzpatrick, L.A.; Schwartz, R.S. Arterial calcification and not lumen stenosis is highly correlated with atherosclerotic plaque burden in humans: A histologic study of 723 coronary artery segments using nondecalcifying methodology. J. Am. Coll. Cardiol. 1998, 31, 126–133. [Google Scholar] [CrossRef] [Green Version]
- Golub, I.; Lakshmanan, S.; Dahal, S.; Budoff, M.J. Utilizing coronary artery calcium to guide statin use. Atherosclerosis 2021, 326, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Soni, M.; Ambrosino, M.; Jacoby, D.S. The Use of Subclinical Atherosclerosis Imaging to Guide Preventive Cardiology Management. Curr. Cardiol. Rep. 2021, 23, 61. [Google Scholar] [CrossRef] [PubMed]
- Dube, P.; DeRiso, A.; Patel, M.; Battepati, D.; Khatib-Shahidi, B.; Sharma, H.; Gupta, R.; Malhotra, D.; Dworkin, L.; Haller, S.; et al. Vascular Calcification in Chronic Kidney Disease: Diversity in the Vessel Wall. Biomedicines 2021, 9, 404. [Google Scholar] [CrossRef]
- Giachelli, C.M. Mechanisms of vascular calcification in uremia. Semin. Nephrol. 2004, 24, 401–402. [Google Scholar] [CrossRef] [PubMed]
- Lanzer, P.; Boehm, M.; Sorribas, V.; Thiriet, M.; Janzen, J.; Zeller, T.; St Hilaire, C.; Shanahan, C. Medial vascular calcification revisited: Review and perspectives. Eur. Heart J. 2014, 35, 1515–1525. [Google Scholar] [CrossRef] [PubMed]
- Essalihi, R.; Ouellette, V.; Dao, H.H.; McKee, M.D.; Moreau, P. Phenotypic modulation of vascular smooth muscle cells during medial arterial calcification: A role for endothelin? J. Cardiovasc. Pharmacol. 2004, 44 (Suppl. 1), S147–S150. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Lu, C.; Chen, X.; Zhao, X.; Xia, D. A new method to quantify coronary calcification by intravascular ultrasound—The different patterns of calcification of acute myocardial infarction, unstable angina pectoris and stable angina pectoris. J. Invasive Cardiol. 2008, 20, 587–590. [Google Scholar] [PubMed]
- Ho, C.Y.; Shanahan, C.M. Medial Arterial Calcification: An Overlooked Player in Peripheral Arterial Disease. Arter. Thromb. Vasc. Biol. 2016, 36, 1475–1482. [Google Scholar] [CrossRef] [Green Version]
- Schurgers, L.J.; Akbulut, A.C.; Kaczor, D.M.; Halder, M.; Koenen, R.R.; Kramann, R. Initiation and Propagation of Vascular Calcification Is Regulated by a Concert of Platelet- and Smooth Muscle Cell-Derived Extracellular Vesicles. Front. Cardiovasc. Med. 2018, 5, 36. [Google Scholar] [CrossRef] [PubMed]
- Leopold, J.A. Vascular calcification: Mechanisms of vascular smooth muscle cell calcification. Trends Cardiovasc. Med. 2015, 25, 267–274. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Cao, F.; Liu, S.; Mi, Y.; Liu, J. BMP2/Smad signaling pathway is involved in the inhibition function of fibroblast growth factor 21 on vascular calcification. Biochem. Biophys. Res. Commun. 2018, 503, 930–937. [Google Scholar] [CrossRef]
- Liu, L.; Zeng, P.; Yang, X.; Duan, Y.; Zhang, W.; Ma, C.; Zhang, X.; Yang, S.; Li, X.; Yang, J.; et al. Inhibition of Vascular Calcification. Arterioscler. Thromb. Vasc. Biol. 2018, 38, 2382–2395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hruska, K.A.; Mathew, S.; Saab, G. Bone morphogenetic proteins in vascular calcification. Circ. Res. 2005, 97, 105–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demer, L.L.; Tintut, Y. Vascular calcification: Pathobiology of a multifaceted disease. Circulation 2008, 117, 2938–2948. [Google Scholar] [CrossRef] [PubMed]
- Scialla, J.J.; Lau, W.L.; Reilly, M.P.; Isakova, T.; Yang, H.Y.; Crouthamel, M.H.; Chavkin, N.W.; Rahman, M.; Wahl, P.; Amaral, A.P.; et al. Chronic Renal Insufficiency Cohort Study Investigators. Fibroblast growth factor 23 is not associated with and does not induce arterial calcification. Kidney Int. 2013, 83, 1159–1168. [Google Scholar] [CrossRef] [Green Version]
- Jimbo, R.; Kawakami-Mori, F.; Mu, S.; Hirohama, D.; Majtan, B.; Shimizu, Y.; Yatomi, Y.; Fukumoto, S.; Fujita, T.; Shimosawa, T. Fibroblast growth factor 23 accelerates phosphate-induced vascular calcification in the absence of Klotho deficiency. Kidney Int. 2014, 85, 1103–1111. [Google Scholar] [CrossRef] [Green Version]
- Tyson, K.L.; Reynolds, J.L.; McNair, R.; Zhang, Q.; Weissberg, P.L.; Shanahan, C.M. Osteo/chondrocytic transcription factors and their target genes exhibit distinct patterns of expression in human arterial calcification. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 489–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teplyuk, N.M.; Haupt, L.M.; Ling, L.; Dombrowski, C.; Mun, F.K.; Nathan, S.S.; Lian, J.B.; Stein, J.L.; Stein, G.S.; Cool, S.M.; et al. The osteogenic transcription factor Runx2 regulates components of the fibroblast growth factor/proteoglycan signaling axis in osteoblasts. J. Cell. Biochem. 2009, 107, 144–154. [Google Scholar] [CrossRef] [Green Version]
- Yang, R.; Zhu, Y.; Wang, Y.; Ma, W.; Han, X.; Wang, X.; Liu, N. HIF-1alpha/PDK4/autophagy pathway protects against advanced glycation end-products induced vascular smooth muscle cell calcification. Biochem. Biophys. Res. Commun. 2019, 517, 470–476. [Google Scholar] [CrossRef]
- Lee, S.J.; Jeong, J.Y.; Oh, C.J.; Park, S.; Kim, J.Y.; Kim, H.J.; Doo Kim, N.; Choi, Y.K.; Do, J.Y.; Go, Y.; et al. Pyruvate Dehydrogenase Kinase 4 Promotes Vascular Calcification via SMAD1/5/8 Phosphorylation. Sci. Rep. 2015, 5, 16577. [Google Scholar] [CrossRef] [Green Version]
- Sukhova, G.K.; Shi, G.P.; Simon, D.I.; Chapman, H.A.; Libby, P. Expression of the elastolytic cathepsins S and K in human atheroma and regulation of their production in smooth muscle cells. J. Clin. Investig. 1998, 102, 576–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lutgens, E.; Lutgens, S.P.; Faber, B.C.; Heeneman, S.; Gijbels, M.M.; de Winther, M.P.; Frederik, P.; van der Made, I.; Daugherty, A.; Sijbers, A.M.; et al. Disruption of the cathepsin K gene reduces atherosclerosis progression and induces plaque fibrosis but accelerates macrophage foam cell formation. Circulation 2006, 113, 98–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Campenhout, A.; Golledge, J. Osteoprotegerin, vascular calcification and atherosclerosis. Atherosclerosis 2009, 204, 321–329. [Google Scholar] [CrossRef] [Green Version]
- Kaden, J.J.; Bickelhaupt, S.; Grobholz, R.; Haase, K.K.; Sarikoc, A.; Kilic, R.; Brueckmann, M.; Lang, S.; Zahn, I.; Vahl, C.; et al. Receptor activator of nuclear factor kappaB ligand and osteoprotegerin regulate aortic valve calcification. J. Mol. Cell. Cardiol. 2004, 36, 57–66. [Google Scholar] [CrossRef]
- Scatena, M.; Liaw, L.; Giachelli, C.M. Osteopontin: A multifunctional molecule regulating chronic inflammation and vascular disease. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 2302–2309. [Google Scholar] [CrossRef] [Green Version]
- Giachelli, C.M.; Speer, M.Y.; Li, X.; Rajachar, R.M.; Yang, H. Regulation of vascular calcification: Roles of phosphate and osteopontin. Circ. Res. 2005, 96, 717–722. [Google Scholar] [CrossRef] [Green Version]
- Heiss, A.; DuChesne, A.; Denecke, B.; Grotzinger, J.; Yamamoto, K.; Renne, T.; Jahnen-Dechent, W. Structural basis of calcification inhibition by alpha 2-HS glycoprotein/fetuin-A. Formation of colloidal calciprotein particles. J. Biol. Chem. 2003, 278, 13333–13341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zazzeroni, L.; Faggioli, G.; Pasquinelli, G. Mechanisms of Arterial Calcification: The Role of Matrix Vesicles. Eur. J. Vasc. Endovasc. Surg. 2018, 55, 425–432. [Google Scholar] [CrossRef] [Green Version]
- Jara, A.; Chacón, C.; Burgos, M.E.; Droguett, A.; Valdivieso, A.; Ortiz, M.; Troncoso, P.; Mezzano, S. Expression of gremlin, a bone morphogenetic protein antagonist, is associated with vascular calcification in uraemia. Nephrol. Dial. Transplant. 2009, 24, 1121–1129. [Google Scholar] [CrossRef]
- Shanahan, C.M.; Crouthamel, M.H.; Kapustin, A.; Giachelli, C.M. Arterial calcification in chronic kidney disease: Key roles for calcium and phosphate. Circ. Res. 2011, 109, 697–711. [Google Scholar] [CrossRef] [Green Version]
- Iyemere, V.P.; Proudfoot, D.; Weissberg, P.L.; Shanahan, C.M. Vascular smooth muscle cell phenotypic plasticity and the regulation of vascular calcification. J. Intern. Med. 2006, 260, 192–210. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Fujiyoshi, A.; Willcox, B.; Choo, J.; Vishnu, A.; Hisamatsu, T.; Ahuja, V.; Takashima, N.; Barinas-Mitchell, E.; Kadota, A.; et al. ERA JUMP Study Group. Increased Aortic Calcification Is Associated With Arterial Stiffness Progression in Multiethnic Middle-Aged Men. Hypertension 2017, 69, 102–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamenskiy, A.; Poulson, W.; Sim, S.; Reilly, A.; Luo, J.; MacTaggart, J. Prevalence of Calcification in Human Femoropopliteal Arteries and its Association with Demographics, Risk Factors, and Arterial Stiffness. Arter. Thromb. Vasc. Biol. 2018, 38, e48–e57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsao, C.W.; Pencina, K.M.; Massaro, J.M.; Benjamin, E.J.; Levy, D.; Vasan, R.S.; Hoffmann, U.; O’Donnell, C.J.; Mitchell, G.F. Cross-sectional relations of arterial stiffness, pressure pulsatility, wave reflection, and arterial calcification. Arter. Thromb. Vasc. Biol. 2014, 34, 2495–2500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Safar, M.E.; Levy, B.I.; Struijker-Boudier, H. Current perspectives on arterial stiffness and pulse pressure in hypertension and cardiovascular diseases. Circulation 2003, 107, 2864–2869. [Google Scholar] [CrossRef] [Green Version]
- Van den Bergh, G.; Opdebeeck, B.; D’Haese, P.C.; Verhulst, A. The Vicious Cycle of Arterial Stiffness and Arterial Media Calcification. Trends Mol. Med. 2019, 25, 1133–1146. [Google Scholar] [CrossRef]
- Hashimoto, J.; Ito, S. Central pulse pressure and aortic stiffness determine renal hemodynamics: Pathophysiological implication for microalbuminuria in hypertension. Hypertension 2011, 58, 839–846. [Google Scholar] [CrossRef] [Green Version]
- O’Rourke, M.F.; Safar, M.E. Relationship between aortic stiffening and microvascular disease in brain and kidney: Cause and logic of therapy. Hypertension 2005, 46, 200–204. [Google Scholar] [CrossRef]
- Zieman, S.J.; Melenovsky, V.; Kass, D.A. Mechanisms, pathophysiology, and therapy of arterial stiffness. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 932–943. [Google Scholar] [CrossRef] [Green Version]
- McEniery, C.M.; McDonnell, B.J.; So, A.; Aitken, S.; Bolton, C.E.; Munnery, M.; Hickson, S.S.; Yasmin; Maki-Petaja, K.M.; Maki-Petaja, K.M.; et al. Aortic calcification is associated with aortic stiffness and isolated systolic hypertension in healthy individuals. Hypertension 2009, 53, 524–531. [Google Scholar] [CrossRef] [Green Version]
- Hinz, B.; Phan, S.H.; Thannickal, V.J.; Prunotto, M.; Desmouliere, A.; Varga, J.; De Wever, O.; Mareel, M.; Gabbiani, G. Recent developments in myofibroblast biology: Paradigms for connective tissue remodeling. Am. J. Pathol. 2012, 180, 1340–1355. [Google Scholar] [CrossRef]
- Luo, G.; Ducy, P.; McKee, M.D.; Pinero, G.J.; Loyer, E.; Behringer, R.R.; Karsenty, G. Spontaneous calcification of arteries and cartilage in mice lacking matrix GLA protein. Nature 1997, 386, 78–81. [Google Scholar] [CrossRef]
- Cranenburg, E.C.; Koos, R.; Schurgers, L.J.; Magdeleyns, E.J.; Schoonbrood, T.H.; Landewé, R.B.; Brandenburg, V.M.; Bekers, O.; Vermeer, C. Characterisation and potential diagnostic value of circulating matrix Gla protein (MGP) species. Thromb. Haemost. 2010, 104, 811–822. [Google Scholar] [CrossRef]
- Wallin, R.; Cain, D.; Hutson, S.M.; Sane, D.C.; Loeser, R. Modulation of the binding of matrix Gla protein (MGP) to bone morphogenetic protein-2 (BMP-2). Thromb. Haemost. 2000, 84, 1039–1044. [Google Scholar]
- Wolinsky, H. Response of the rat aortic media to hypertension. Morphological and chemical studies. Circ. Res. 1970, 26, 507–522. [Google Scholar] [CrossRef] [Green Version]
- Wajih, N.; Borras, T.; Xue, W.; Hutson, S.M.; Wallin, R. Processing and transport of matrix gamma-car- boxyglutamic acid protein and bone morphogenetic protein-2 in cultured human vascular smooth muscle cells: Evidence for an uptake mechanism for serum fetuin. J. Biol. Chem. 2004, 279, 43052–43060. [Google Scholar] [CrossRef] [Green Version]
- Price, P.A.; Faus, S.A.; Williamson, M.K. Warfarin causes rapid calcification of the elastic lamellae in rat arteries and heart valves. Arter. Thromb. Vasc. Biol. 1998, 18, 1400–1407. [Google Scholar] [CrossRef] [Green Version]
- Price, P.A.; Faus, S.A.; Williamson, M.K. Warfarin induced artery calcification is accelerated by growth and vitamin D. Arter. Thromb. Vasc. Biol. 2000, 20, 317–327. [Google Scholar] [CrossRef] [Green Version]
- Sweatt, A.; Sane, D.C.; Hutson, S.M.; Wallin, R. Matrix Gla protein (MGP) and bone mor- phogenetic protein-2 in aortic calcified lesions of aging rats. J. Thromb. Haemost. 2003, 1, 178–185. [Google Scholar] [CrossRef]
- Schurgers, L.J.; Teunissen, K.J.; Knapen, M.H.; Kwaijtaal, M.; van Diest, R.; Appels, A.; Reutelingsperger, A.P.; Cleutjens, J.P.M.; Vermeer, C. Novel conformation-specific anti- bodies against matrix gamma-carboxyglutamic acid (Gla) protein: Undercar-boxylated matrix Gla protein as marker for vascular calcification. Arter. Thromb. Vasc. Biol. 2005, 25, 1629–1633. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, P.; Epstein, D.L.; Borrás, T. Characterization of gene expression in human trabecular meshwork using single-pass sequencing of 1060 clones. Investig. Ophthalmol. Vis. Sci. 2000, 41, 3678–3693. [Google Scholar]
- Borrás, T.; Smith, M.H.; Buie, L.K. A novel Mgp-Cre knock-in mouse reveals an anticalcification/antistiffness candidate gene in the trabecular meshwork and peripapillary scleral region. Investig. Ophthalmol. Vis. Sci. 2015, 56, 2203–2214. [Google Scholar] [CrossRef] [Green Version]
- Wei, F.F.; Huang, Q.F.; Zhang, Z.Y.; Van Keer, K.; Thijs, L.; Trenson, S.; Yang, W.Y.; Cauwenberghs, N.; Mujaj, B.; Kuznetsova, T.; et al. Inactive matrix Gla protein is a novel circulating biomarker predicting retinal arteriolar narrowing in humans. Sci. Rep. 2018, 8, 15088. [Google Scholar] [CrossRef] [Green Version]
- Asokan, P.; Mitra, R.N.; Periasamy, R.; Han, Z.; Borrás, T. A naturally fluorescent Mgp transgenic mouse for angiogenesis and glaucoma longitudinal studies. Investig. Ophthalmol. Vis. Sci. 2018, 59, 746–756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, F.F.; Drummen, N.E.; Thijs, L.; Jacobs, L.; Herfs, M.; Van’t Hoofd, C.; Vermeer, C.; Staessen, J.A. Vitamin-K-dependent protection of the renal microvasculature: Histopathological studies in normal and diseased kidneys. Pulse 2016, 4, 85–91. [Google Scholar] [CrossRef] [Green Version]
- Wei, F.F.; Drummen, N.E.; Schutte, A.E.; Thijs, L.; Jacobs, L.; Petit, T.; Yang, W.-Y.; Smith, W.; Zhang, Z.-Y.; Gu, Y.-M.; et al. Vitamin K dependent protection of renal function in multi-ethnic population studies. EBioMedicine 2016, 4, 162–169. [Google Scholar] [CrossRef] [Green Version]
- Wei, F.F.; Trenson, S.; Thijs, L.; Huang, Q.F.; Zhang, Z.Y.; Yang, W.Y.; Moliterno, P.; Allegaert, K.; Boggia, J.; Janssens, S.; et al. Desphospho-uncarboxylated matrix Gla protein is a novel circulating biomarker predicting deterioration of renal function in the general population. Nephrol. Dial. Transplant. 2018, 33, 1122–1128. [Google Scholar] [CrossRef]
- Puzantian, H.; Akers, S.R.; Oldland, G.; Javaid, K.; Miller, R.; Ge, Y.; Ansari, B.; Lee, J.; Suri, A.; Hasmath, Z. Circulating dephospho-uncarboxylated matrix Gla-protein is associated with kidney dysfunction and arterial stiffness. Am. J. Hypertens. 2018, 31, 988–994. [Google Scholar] [CrossRef]
- Brnic, D.; Martinovic, D.; Zivkovic, P.M.; Tokic, D.; Vilovic, M.; Rusic, D.; Tadin Hadjina, I.; Libers, C.; Glumac, S.; Supe-Domic, D.; et al. Inactive matrix Gla protein is elevated in patients with inflammatory bowel disease. World J. Gastroenterol. 2020, 26, 4866–4877. [Google Scholar] [CrossRef]
- Vilovic, M.; Dogas, Z.; Ticinovic Kurir, T.; Borovac, J.A.; Supe-Domic, D.; Vilovic, T.; Ivkovic, N.; Rusic, D.; Novak, A.; Bozic, J. Bone metabolism parameters and inactive matrix Gla protein in patients with obstructive sleep apnea. Sleep 2020, 43, zsz243. [Google Scholar] [CrossRef]
- Shanahan, C.M.; Cary, N.R.; Metcalfe, J.C.; Weissberg, P.L. High expression of genes for calcification-regulating proteins in human atherosclerotic plaques. J. Clin. Investig. 1994, 93, 2393–2402. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Bennett, B.J.; Wang, X.; Rosenfeld, M.E.; Giachelli, C.; Lusis, A.J.; Boström, K.I. Inhibition of bone morphogenetic proteins protects against atherosclerosis and vascular calcification. Circ. Res. 2010, 107, 485–494. [Google Scholar] [CrossRef]
- Schurgers, L.J.; Joosen, I.A.; Laufer, E.M.; Chatrou, M.L.; Herfs, M.; Winkens, M.H.; Westenfeld, R.; Veulemans, V.; Krueger, T.; Shanahan, C.M.; et al. Vitamin K-antagonists accelerate atherosclerotic calcification and induce a vulnerable plaque phenotype. PLoS ONE 2012, 7, e43229. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, Y.; Ikeda, K.; Akakabe, Y.; Koide, M.; Uraoka, M.; Yutaka, K.; Kurimoto-Nakano, R.; Takahashi, T.; Matoba, S.; Yamada, H.; et al. Paracrine osteogenic signals via bone morphogenetic protein-2 accelerate the atherosclerotic intimal calcification in vivo. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 1908–1915. [Google Scholar] [CrossRef] [Green Version]
- van Gorp, R.H.; Dijkgraaf, I.; Bröker, V.; Bauwens, M.; Leenders, P.; Jennen, D.; Dweck, M.R.; Bucerius, J.; Briedé, J.J.; van Ryn, J.; et al. Off-target effects of oral anticoagulants—Vascular effects of vitamin K antagonist and non-vitamin K antagonist oral anticoagulant dabigatran etexilate. J. Thromb. Haemost. 2021, 19, 1348–1363. [Google Scholar] [CrossRef]
- Rattazzi, M.; Faggin, E.; Bertacco, E.; Nardin, C.; Pagliani, L.; Plebani, M.; Cinetto, F.; Guidolin, D.; Puato, M.; Pauletto, P. Warfarin, but not rivaroxaban, promotes the calcification of the aortic valve in ApoE-/- mice. Cardiovasc. Ther. 2018, 36, e12438. [Google Scholar] [CrossRef]
- Debernardi, N.; Roijers, R.B.; Krams, R.; de Crom, R.; Mutsaers, P.H.; van der Vusse, G.J. Microcalcifications in atherosclerotic lesion of apolipoprotein E-deficient mouse. Int. J. Exp. Pathol. 2010, 91, 485–494. [Google Scholar] [CrossRef]
- Dalmeijer, G.W.; van der Schouw, Y.T.; Vermeer, C.; Magdeleyns, E.J.; Schurgers, L.J.; Beulens, J.W. Circulating matrix Gla protein is associated with coronary artery calcification and vitamin K status in healthy women. J. Nutr. Biochem. 2013, 24, 624–628. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.J. Assessing the role of circulating, genetic, and imaging biomarkers in cardiovascular risk prediction. Circulation 2011, 123, 551–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vassalle, C.; Iervasi, G. New insights for matrix Gla protein, vascular calcification and cardiovascular risk and outcome. Atherosclerosis 2014, 235, 236–238. [Google Scholar] [CrossRef] [PubMed]
- Collet, J.P.; Thiele, H.; Barbato, E.; Barthélémy, O.; Bauersachs, J.; Bhatt, D.L.; Dendale, P.; Dorobantu, M.; Edvardsen, T.; Folliguet, T.; et al. ESC Scientific Document Group. 2020 ESC Guidelines for the management of acute coronary syndromes in patients presenting without persistent ST-segment elevation. Eur. Heart J. 2020, 37, ehaa575. [Google Scholar]
- Mayer, O., Jr.; Seidlerová, J.; Bruthans, J.; Filipovský, J.; Timoracká, K.; Vaněk, J.; Cerná, L.; Wohlfahrt, P.; Cífková, R.; Theuwissen, E.; et al. Desphospho-uncarboxylated matrix Gla-protein is associated with mortality risk in patients with chronic stable vascular disease. Atherosclerosis 2014, 235, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Dalmeijer, G.W.; van der Schouw, Y.T.; Magdeleyns, E.J.; Vermeer, C.; Verschuren, W.M.; Boer, J.M.; Beulens, J.W. Matrix Gla protein species and risk of cardiovascular events in type 2 diabetic patients. Diabetes Care 2013, 36, 3766–3771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van den Heuvel, E.G.; van Schoor, N.M.; Lips, P.; Magdeleyns, E.J.; Deeg, D.J.; Vermeer, C.; den Heijer, M. Circulating uncarboxylated matrix Gla protein, a marker of vitamin K status, as a risk factor of cardiovascular disease. Maturitas 2014, 77, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Dalmeijer, G.W.; van der Schouw, Y.T.; Magdeleyns, E.J.; Vermeer, C.; Verschuren, W.M.; Boer, J.M.; Beulens, J.W. Circulating desphospho-uncarboxylated matrix γ-carboxyglutamate protein and the risk of coronary heart disease and stroke. J. Thromb. Haemost. 2014, 12, 1028–1034. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.P.; Gu, Y.M.; Thijs, L.; Knapen, M.H.; Salvi, E.; Citterio, L.; Petit, T.; Carpini, S.D.; Zhang, Z.; Jacobs, L.; et al. Inactive matrix Gla protein is causally related to adverse health outcomes: A Mendelian randomization study in a Flemish population. Hypertension 2015, 65, 463–470. [Google Scholar] [CrossRef] [Green Version]
- Zwakenberg, S.R.; van der Schouw, Y.T.; Vermeer, C.; Pasterkamp, G.; den Ruijter, H.M.; Beulens, J.W.J. Matrix Gla protein, plaque stability, and cardiovascular events in patients with severe atherosclerotic disease. Cardiology 2018, 141, 32–36. [Google Scholar] [CrossRef]
- Csiszar, A.; Smith, K.E.; Koller, A.; Kaley, G.; Edwards, J.G.; Ungvari, Z. Regulation of bone morphogenetic protein-2 expression in endothelial cells: Role of nuclear factor-kappaB activation by tumor necrosis factor-alpha, H2O2, and high intravascular pressure. Circulation 2005, 111, 2364–2372. [Google Scholar] [CrossRef] [Green Version]
- Tota-Maharaj, R.; Joshi, P.H.; Budoff, M.J.; Whelton, S.; Zeb, I.; Rumberger, J.; Al-Mallah, M.; Blumenthal, R.S.; Nasir, K.; Blaha, M.J. Usefulness of regional distribution of coronary artery calcium to improve the prediction of all-cause mortality. Am. J. Cardiol. 2015, 115, 1229–1234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kizu, A.; Jono, S. Mechanism of vascular calcification. Clin. Calcium 2004, 14, 92–96. [Google Scholar]
- Kolodgie, F.D.; Narula, J.; Haider, N.; Virmani, R. Apoptosis in atherosclerosis. Does it contribute to plaque instability? Cardiol. Clin. 2001, 19, 127–139. [Google Scholar] [CrossRef]
- Tabas, I. Macrophage death and defective inflammation resolution in atherosclerosis. Nat. Rev. Immunol. 2010, 10, 36–46. [Google Scholar] [CrossRef]
- Borissoff, J.I.; Spronk, H.M.; Ten Cate, H. The Hemostatic System as a Modulator of Atherosclerosis. N. Engl. J. Med. 2011, 364, 1746–1760. [Google Scholar] [CrossRef] [PubMed]
- Bilalic, A.; Ticinovic Kurir, T.; Kumric, M.; Borovac, J.A.; Matetic, A.; Supe-Domic, D.; Bozic, J. Circulating Levels of Dephosphorylated-Uncarboxylated Matrix Gla Protein in Patients with Acute Coronary Syndrome. Molecules 2021, 26, 1108. [Google Scholar] [CrossRef]
- Keyzer, C.A.; Vermeer, C.; Joosten, M.M.; Knapen, M.H.; Drummen, N.E.; Navis, G.; Bakker, S.J.; de Borst, M.H. Vitamin K status and mortality after kidney transplantation: A cohort study. Am. J. Kidney Dis. 2015, 65, 474–483. [Google Scholar] [CrossRef] [PubMed]
- Riphagen, I.J.; Keyzer, C.A.; Drummen, N.E.A.; de Borst, M.H.; Beulens, J.W.J.; Gansevoort, R.T.; Geleijnse, J.M.; Muskiet, F.A.J.; Navis, G.; Visser, S.T.; et al. Prevalence and effects of functional vitamin K insufficiency: The PREVEND study. Nutrients 2017, 9, 1334. [Google Scholar] [CrossRef] [Green Version]
- Schurgers, L.J.; Barreto, D.V.; Barreto, F.C.; Liabeuf, S.; Renard, C.; Magdeleyns, E.J.; Vermeer, C.; Choukroun, G.; Massy, Z.A. The circulating inactive form of matrix gla protein is a surrogate marker for vascular calcification in chronic kidney disease: A preliminary report. Clin. J. Am. Soc. Nephrol. 2010, 5, 568–575. [Google Scholar] [CrossRef] [Green Version]
- Ueland, T.; Gullestad, L.; Dahl, C.P.; Aukrust, P.; Aakhus, S.; Solberg, O.G.; Vermeer, C.; Schurgers, L.J. Undercarboxylated matrix Gla protein is associated with indices of heart failure and mortality in symptomatic aortic stenosis. J. Intern. Med. 2010, 268, 483–492. [Google Scholar] [CrossRef]
- Ueland, T.; Dahl, C.P.; Gullestad, L.; Aakhus, S.; Broch, K.; Skårdal, R.; Vermeer, C.; Aukrust, P.; Schurgers, L.J. Circulating levels of non-phosphorylated undercarboxylated matrix Gla protein are associated with disease severity in patients with chronic heart failure. Clin. Sci. 2011, 121, 119–127. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Role | Molecule | Mechanism | Evidence |
|---|---|---|---|
| Vascular calcification inducers | BMP-2 | Contributes to the transdifferentiation of VSMCs into osteochondrogenic cells; induces osteoblast differentiation; enhances apoptosis, oxidative stress and inflammation in VSMCs | [33,34,35] |
| ALP | Its activity is important for hydroxyapatite formation | [36] | |
| FGF-23 | Increases osteoblastic marker expression in VSMCs * | [37,38] | |
| Runx2 | Transcription factor—increases expression of osteogenic genes | [39,40] | |
| PDK4 | Induces osteogenic differentiation of VSMCs | [41,42] | |
| Cathepsin K | Degrades organic bone matrix in osteoclasts | [43,44] | |
| Vascular calcification inhibitors | Osteoprotegerin | Interferes with RANK-RANKL interaction | [45,46] |
| Osteopontin | Strong binding affinity for hydroxyapatite | [47,48] | |
| Fetuin-A | Binds to early calcium phosphate crystals inhibits growth and deposition | [49,50] | |
| BMP-7 | Reduces transformation to VSMC osteogenic phenotypes | [51] | |
| MGP | Binding to hydroxyapatite; BMP-2 binding and inhibition | [52,53] |
| Study | Study Population | Measured Outcomes (Median Duration) | Results |
|---|---|---|---|
| Dalmeijer et al. [93] | 518 DM type II patients | HRSD of dp-ucMGP, t-ucMGP and dp-cMGP for CVD, CHD, PAD, HF and stroke adjusted for sex, age, BMI, waist-to-hip ratio, blood pressure, total cholesterol, smoking, physical activity, and education (11.2 y) | Significant HRSD 1:
|
| Dalmeijer et al. [95] | 1154 incident cases of CHD and 380 of stroke + 1406 random participants (EPIC-NL) | HRSD of dp-ucMGP for CHD risk and stroke (11.5 y) | HRSD of dp-ucMGP:
|
| Keyzer et al. [107] | 518 stable kidney transplant recipients | HR highest vs. lowest tertile of dp-ucMGP for TM and transplant failure (9.8 y) | HRs highest vs. lowest tertile of dp-ucMGP:
|
| Liu et al. [96] | 2318 FLEMENGHO participants | HR associated with dp-ucMGP doubling for TM, CVM, CVD and CHD adjusted for sex, age, body mass index, systolic blood pressure, heart rate, smoking and drinking, total cholesterol, DM, antihypertensive drug treatment, and history of CVD (14.1 y) | Significant HRs for dp-ucMGP doubling:
|
| Mayer et al. [94] | 799 patients with myocardial infarction, coronary revascularization or first ischemic stroke | HR highest vs. other quartiles of dp-ucMGP and dp-cMGP for TM and CVM (5.6 y) | HRs for highest quartile vs. Q1–Q3:
|
| Riphagen et al. [108] | 4275 PREVEND participants | HR associated with dp-ucMGP doubling for TM and CVM, adjusted for 2 (8.5 y) | HRs for dp-ucMGP doubling:
|
| Schurgers et al. [109] | 107 patients with CKD | RR of dp-ucMGP median (>921 p·mol/L) for TM adjusted for age, CKD stage or hemoglobin | RR for TM: 2.85 (1.36–5.90); significance lost in multivariable-adjusted models |
| Ueland et al. [110] | 147 patients with symptomatic severe aortic stenosis | HR high versus low dp-cMGP and dp-ucMGP concentration for TM (23 months) | HRs high vs. low:
|
| Ueland et al. [111] | 179 patients with chronic HF | HRSD of dp-ucMGP for TM, fatal HF and heart transplant (2.9 y) | HRSD of dp-ucMGP:
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumric, M.; Borovac, J.A.; Ticinovic Kurir, T.; Martinovic, D.; Frka Separovic, I.; Baric, L.; Bozic, J. Role of Matrix Gla Protein in the Complex Network of Coronary Artery Disease: A Comprehensive Review. Life 2021, 11, 737. https://doi.org/10.3390/life11080737
Kumric M, Borovac JA, Ticinovic Kurir T, Martinovic D, Frka Separovic I, Baric L, Bozic J. Role of Matrix Gla Protein in the Complex Network of Coronary Artery Disease: A Comprehensive Review. Life. 2021; 11(8):737. https://doi.org/10.3390/life11080737
Chicago/Turabian StyleKumric, Marko, Josip A. Borovac, Tina Ticinovic Kurir, Dinko Martinovic, Ivan Frka Separovic, Ljupka Baric, and Josko Bozic. 2021. "Role of Matrix Gla Protein in the Complex Network of Coronary Artery Disease: A Comprehensive Review" Life 11, no. 8: 737. https://doi.org/10.3390/life11080737