
‘Whole Organism’, Systems Biology, and Top-Down Criteria for Evaluating Scenarios for the Origin of Life


Abstract
:1. Introduction
1.1. The Value of Scenarios
1.2. The Range and Scope of Proposed Initial Venues and Accompanying Scenarios
1.3. Criteria for Evaluating Scenarios
1.4. Some Terminology
2. ‘Whole Organism’ Criteria
Probability of Life
3. Systems Biology Criteria
3.1. Integration Began from the Outset
3.2. Spatial Heterogeneity—Multi-Pot Rather Than Single-Pot Prebiotic Chemistry
4. Top-Down Criteria and the Principle of Continuity
4.1. The Principle of Continuity Connects Bottom-Up with Top-Down Data
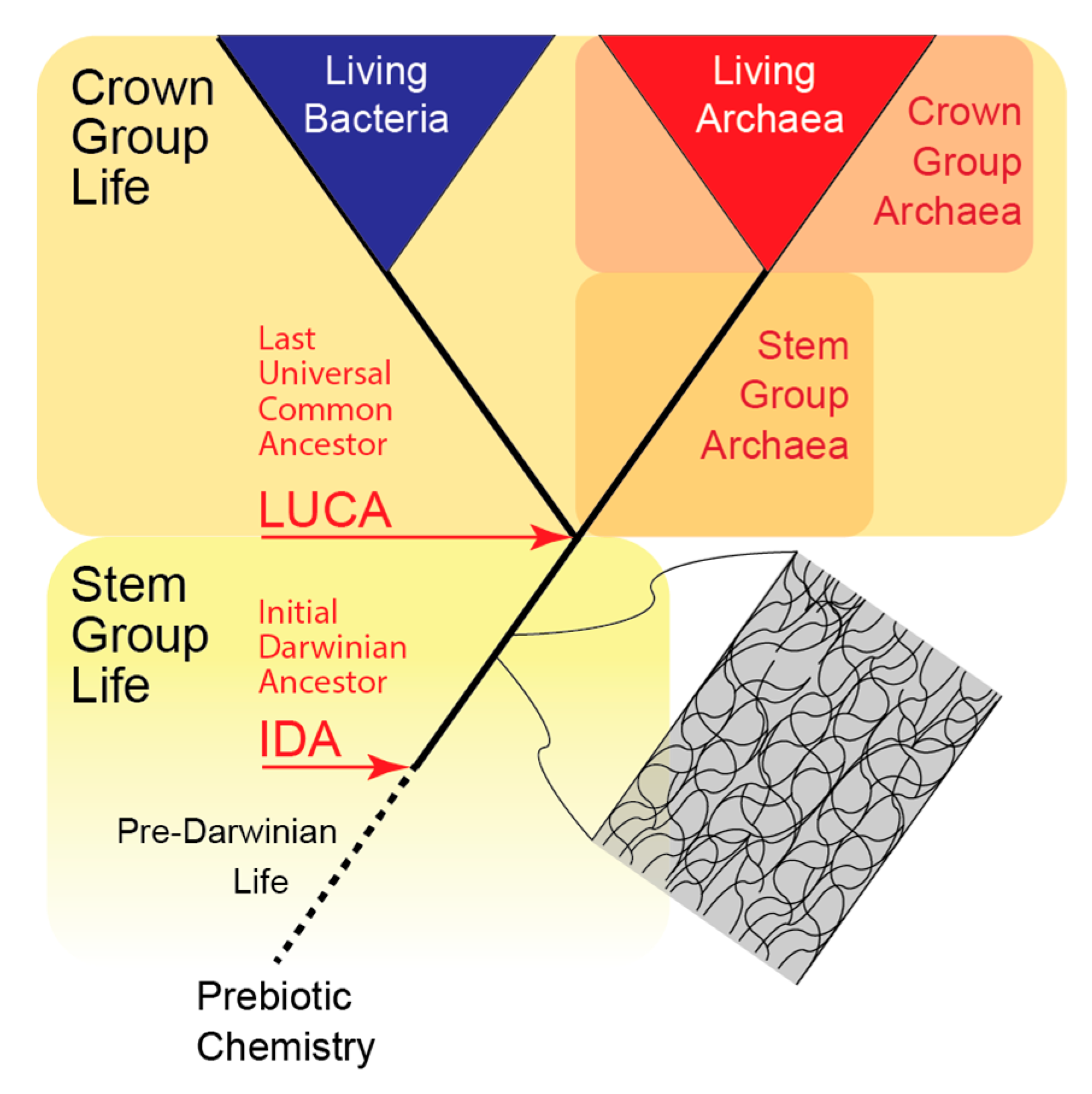
4.2. Early Versus Late Cellularization
4.3. Additional Evidence for Late Cellularization—Different Locomotory Structures
4.4. Analog Versus Non-Analog Properties and the Principle of Continuity
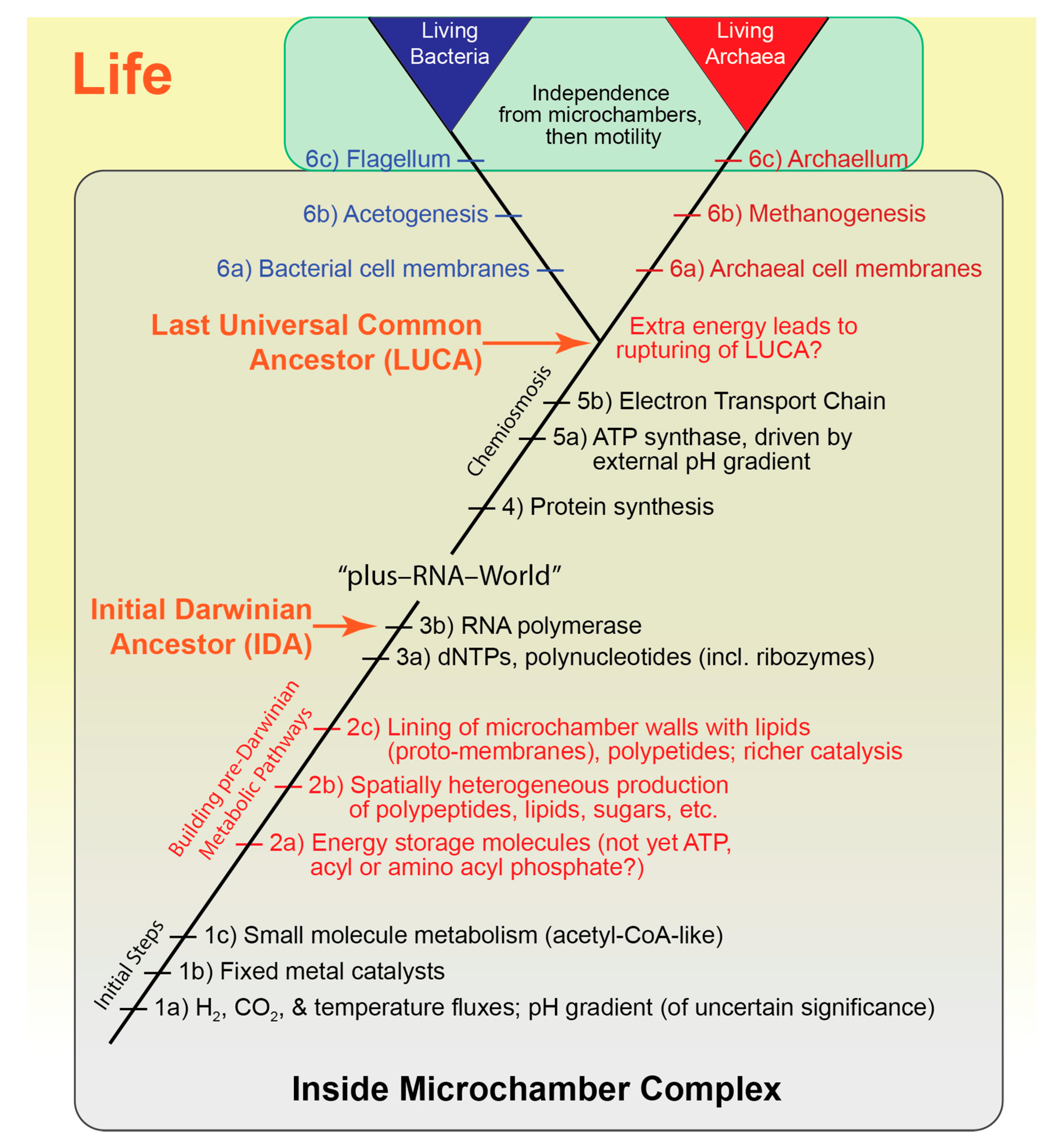
4.5. Pre-Darwinian Complexity—Implications from Top-Down Systems Biology
4.6. Implications for the “RNA-World”—The “Plus-RNA-World”
5. The Role of Chemical and Physical Principles
6. Value-Added Criterion
7. Evaluation of Scenarios
7.1. Primary Weakness of the Marine Microchamber Scenario—Dehydration Reactions in the Deep
7.2. Water
7.3. A Hybrid Scenario?
8. The Microchamber Complex Scenario
8.1. A Little History
8.2. The Alkaline Vent Microchamber Complexes—Megacities
8.3. Suitable Boundary Conditions
8.4. The Origin of Life in the Microchamber Complexes as a Massively Parallel Process
Multiple Origins of Life
8.5. The Microchamber Complex Scenario—Initial Steps
8.6. Building Biochemical Pathways
8.6.1. Linking Exergonic and Endergonic Steps—Energy Storage Molecules
8.6.2. Spatial Heterogeneity
8.6.3. Likely Complexity of the Prebiotic Molecular Community
8.6.4. Multiple Microchambers May Be the Homolog of the Cell
8.6.5. Transition from Fixed Inorganic Catalysts to Mobile Organic Catalysts
8.7. The Transition to Darwinian Life—From Nucleotides to Ribozymes to RNA Polymerase
8.8. From Ribozymes to Enzymes
8.8.1. Protein Synthesis Greatly Accelerated the Rate of Evolution
8.8.2. ATP Synthase
8.8.3. The Electron Transport Chain
8.8.4. LUCA
8.9. The Increase in Available Energy Ruptured the Microchamber Molecular Community, Thereby Creating LUCA
8.9.1. Independent Emergence of Bacteria and Archaea
8.9.2. A Note on the Role of DNA
9. Quo Vadis?
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Preiner, M.; Asche, S.; Becker, S.; Betts, H.C.; Boniface, A.; Camprubi, E.; Chandru, K.; Erastova, V.; Garg, S.G.; Khawaja, N.; et al. The future of origin of life research: Bridging decades-old divisions. Life 2020, 10, 20. [Google Scholar] [CrossRef] [Green Version]
- Russell, M.J. The “Water Problem”(sic), the Illusory Pond and Life’s Submarine Emergence—A Review. Life 2021, 11, 429. [Google Scholar] [CrossRef]
- Miller, S.L. A production of amino acids under possible primitive earth conditions. Science 1953, 117, 528–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, S.L.; Urey, H.C. Organic compound synthesis on the primitive Earth. Science 1959, 130, 245–251. [Google Scholar] [CrossRef]
- Zaug, A.J.; Cech, T.R. In vitro splicing of the ribosomal RNA precursor in nuclei of Tetrahymena. Cell 1980, 19, 331–338. [Google Scholar] [CrossRef]
- Guerrier-Takada, C.; Gardiner, K.; Marsh, T.; Pace, N.; Altman, S. The RNA moiety of ribonuclease P is the catalytic subunit of the enzyme. Cell 1983, 35, 849–857. [Google Scholar] [CrossRef]
- Kitadai, N.; Maruyama, S. Origins of building blocks of life: A review. Geosci. Front. 2018, 9, 1117–1153. [Google Scholar] [CrossRef]
- Becker, S.; Schneider, C.; Crisp, A.; Carell, T. Non-canonical nucleosides and chemistry of the emergence of life. Nat. Commun. 2018, 9, 5174. [Google Scholar] [CrossRef]
- Chandru, K.; Jia, T.Z.; Mamajanov, I.; Bapat, N.; Cleaves, H.J. Prebiotic oligomerization and self-assembly of structurally diverse xenobiological monomers. Sci. Rep. 2020, 10, 17560. [Google Scholar] [CrossRef]
- Benner, S.A.; Bell, E.A.; Biondi, E.; Brasser, R.; Carell, T.; Kim, H.-J.; Mojzsis, S.J.; Omran, A.; Pasek, M.A.; Trail, D. When Did Life Likely Emerge on Earth in an RNA-First Process? ChemSystemsChem 2020, 2, e2000010. [Google Scholar] [CrossRef]
- Martin, W.; Baross, J.; Kelley, D.; Russell, M.J. Hydrothermal vents and the origin of life. Nat. Rev. Microbiol. 2008, 6, 805–814. [Google Scholar] [CrossRef]
- Martin, W.; Russell, M.J. On the origin of biochemistry at an alkaline hydrothermal vent. Philos. Trans. R. Soc. B Biol. Sci. 2007, 362, 1887–1925. [Google Scholar] [CrossRef]
- Russell, M.J.; Hall, A.J.; Cairns-Smith, A.G.; Braterman, P.S. Submarine hot springs and the origin of life. Nature 1988, 336, 117. [Google Scholar] [CrossRef]
- Russell, M.J.; Hall, A.J.; Martin, W. Serpentinization as a source of energy at the origin of life. Geobiology 2010, 8, 355–371. [Google Scholar] [CrossRef]
- Russell, M.J.; Barge, L.M.; Bhartia, R.; Bocanegra, D.; Bracher, P.J.; Branscomb, E.; Kidd, R.; McGlynn, S.; Meier, D.H.; Nitschke, W.; et al. The drive to life on wet and Icy Worlds. Astrobiology 2014, 14, 308–343. [Google Scholar] [CrossRef]
- Sousa, F.L.; Thiergart, T.; Landan, G.; Nelson-Sathi, S.; Pereira, I.A.C.; Allen, J.F.; Lane, N.; Martin, W.F. Early bioenergetic evolution. Philos. Trans. R. Soc. B Biol. Sci. 2013, 368. [Google Scholar] [CrossRef] [Green Version]
- Russell, M.J. Green rust: The simple organizing ‘seed’ of all life? Life 2018, 8, 35. [Google Scholar] [CrossRef] [PubMed]
- Westall, F.; Hickman-Lewis, K.; Hinman, N.; Gautret, P.; Campbell, K.A.; Bréhéret, J.G.; Foucher, F.; Hubert, A.; Sorieul, S.; Dass, A.V.; et al. A Hydrothermal-Sedimentary Context for the Origin of Life. Astrobiology 2018, 18, 259–293. [Google Scholar] [CrossRef] [PubMed]
- Wächtershäuser, G. On the chemistry and evolution of the pioneer organism. Chem. Biodivers. 2007, 4, 584–602. [Google Scholar] [CrossRef]
- Mulkidjanian, A.Y.; Bychkov, A.Y.; Dibrova, D.V.; Galperin, M.Y.; Koonin, E.V. Origin of first cells at terrestrial, anoxic geothermal fields. Proc. Natl. Acad. Sci. USA 2012, 109. [Google Scholar] [CrossRef] [Green Version]
- Damer, B.; Deamer, D. The hot spring hypothesis for an origin of life. Astrobiology 2020, 20, 429–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasselov, D.D.; Grotzinger, J.P.; Sutherland, J.D. The origin of life as a planetary phenomenon. Sci. Adv. 2020, 6, eaax3419. [Google Scholar] [CrossRef] [Green Version]
- Brasier, M.D.; Matthewman, R.; McMahon, S.; Wacey, D. Pumice as a remarkable substrate for the origin of life. Astrobiology 2011, 11, 725–735. [Google Scholar] [CrossRef] [Green Version]
- Brasier, M.D.; Matthewman, R.; McMahon, S.; Kilburn, M.R.; Wacey, D. Pumice from the ∼3460Ma Apex Basalt, Western Australia: A natural laboratory for the early biosphere. Precambrian Res. 2013, 224, 1–10. [Google Scholar] [CrossRef]
- Hansma, H.G. Better than membranes at the origin of life? Life 2017, 7, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernhardt, H.S. Making molecules with clay: Layered double hydroxides, pentopyranose nucleic acids and the origin of life. Life 2019, 9, 19. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, B.; Muhling, J.R.; Fischer, W.W. Greenalite Nanoparticles in Alkaline Vent Plumes as Templates for the Origin of Life. Astrobiology 2021, 21, 246–259. [Google Scholar] [CrossRef] [PubMed]
- Camprubí, E.; de Leeuw, J.W.; House, C.H.; Raulin, F.; Russell, M.J.; Spang, A.; Tirumalai, M.R.; Westall, F. The Emergence of Life. Space Sci. Rev. 2019, 215, 1–53. [Google Scholar] [CrossRef] [Green Version]
- Darwin Correspondence Project, “Letter no. 7471”. Available online: https://www.darwinproject.ac.uk/letter/DCP-LETT-7471.xml (accessed on 13 May 2021).
- Peretó, J.; Bada, J.L.; Lazcano, A. Charles Darwin and the origin of life. Orig. Life Evol. Biosph. 2009, 39, 395–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wächtershäuser, G. Chemoautotrophic origin of life: The iron-sulfur world hypothesis. In Geomicrobiology: Molecular and Environmental Perspective; Springer: Dordrecht/Heidelberg, Germany, 2010; pp. 1–35. [Google Scholar]
- Sojo, V.; Herschy, B.; Whicher, A.; Camprubí, E.; Lane, N. The Origin of Life in Alkaline Hydrothermal Vents. Astrobiology 2016, 16, 181–197. [Google Scholar] [CrossRef]
- Blackmond, D.G. Autocatalytic Models for the Origin of Biological Homochirality. Chem. Rev. 2020, 120, 4831–4847. [Google Scholar] [CrossRef]
- Gould, S.B. Membranes and evolution. Curr. Biol. 2018, 28, R381–R385. [Google Scholar] [CrossRef] [Green Version]
- Marshall, C.R.; Valentine, J.W. The importance of preadapted genomes in the origin of the animal bodyplans and the Cambrian explosion. Evolution 2010, 64, 1189–1201. [Google Scholar] [CrossRef]
- Donoghue, P.C.J. Saving the stem group—a contradiction in terms? Paleobiology 2005, 31, 553. [Google Scholar] [CrossRef]
- Yarus, M. Getting past the RNA world: The initial Darwinian ancestor. Cold Spring Harb. Perspect. Biol. 2011, 3, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Morowitz, H.J. Energy Flow in Biology; Biological Organization as a Problem in Thermal Physics; Academic Press: Waltham, MA, USA, 1968. [Google Scholar]
- Darwin, C. On the Origin of Species; Dover Publications: Mineola, NY, USA, 1859. [Google Scholar]
- Mayr, E. One Long Argument: Charles Darwin and the Genesis of Modern Evolutionary Thought; Harvard University Press: Cambridge, MA, USA, 1991; Volume 2. [Google Scholar]
- Trifonov, E.N. Vocabulary of definitions of life suggests a definition. J. Biomol. Struct. Dyn. 2011, 29, 259–266. [Google Scholar] [CrossRef] [Green Version]
- Bartlett, S.; Wong, M.L. Defining lyfe in the universe: From three privileged functions to four pillars. Life 2020, 10, 42. [Google Scholar] [CrossRef] [Green Version]
- Astrobiology at NASA—About Life Detection. Available online: https://astrobiology.nasa.gov/research/life-detection/about/ (accessed on 13 July 2021).
- Thornhill, R.H.; Ussery, D.W. A classification of possible routes of Darwinian evolution. J. Theor. Biol. 2000, 203, 111–116. [Google Scholar] [CrossRef] [Green Version]
- Lyon, A.S.; Peeples, W.B.; Rosen, M.K. A framework for understanding the functions of biomolecular condensates across scales. Nat. Rev. Mol. Cell Biol. 2021, 22, 215–235. [Google Scholar] [CrossRef]
- Cohan, M.C.; Pappu, R.V. Making the Case for Disordered Proteins and Biomolecular Condensates in Bacteria. Trends Biochem. Sci. 2020, 45, 668–680. [Google Scholar] [CrossRef]
- Patel, B.H.; Percivalle, C.; Ritson, D.J.; Duffy, C.D.; Sutherland, J.D. Common origins of RNA, protein and lipid precursors in a cyanosulfidic protometabolism. Nat. Chem. 2015, 7, 301–307. [Google Scholar] [CrossRef] [Green Version]
- Clark, B.C.; Kolb, V.M. Macrobiont: Cradle for the origin of life and creation of a biosphere. Life 2020, 10, 278. [Google Scholar] [CrossRef]
- do Vieira, A.N.; Kleinermanns, K.; Martin, W.F.; Preiner, M. The ambivalent role of water at the origins of life. FEBS Lett. 2020, 594, 2717–2733. [Google Scholar] [CrossRef]
- Koonin, E.V.; Martin, W. On the origin of genomes and cells within inorganic compartments. Trends Genet. 2005, 21, 647–654. [Google Scholar] [CrossRef]
- Weiss, M.C.; Sousa, F.L.; Mrnjavac, N.; Neukirchen, S.; Roettger, M.; Nelson-Sathi, S.; Martin, W.F. The physiology and habitat of the last universal common ancestor. Nat. Microbiol. 2016, 1, 16116. [Google Scholar] [CrossRef]
- Weiss, M.C.; Preiner, M.; Xavier, J.C.; Zimorski, V.; Martin, W.F. The last universal common ancestor between ancient Earth chemistry and the onset of genetics. PLoS Genet. 2018, 14, e1007518. [Google Scholar] [CrossRef] [Green Version]
- Gogarten, J.P.; Deamer, D. Is LUCA a thermophilic progenote? Nat. Microbiol. 2016, 1. [Google Scholar] [CrossRef]
- Berkemer, S.J.; McGlynn, S.E.; Berkemer, S.J.; McGlynn, S.E. A new analysis of archaea-bacteria domain separation: Variable phylogenetic distance and the tempo of early evolution. Mol. Biol. Evol. 2021, 37, 2332–2340. [Google Scholar] [CrossRef]
- Orgel, L.E. Evolution of the genetic apparatus. J. Mol. Biol. 1968, 38, 381–393. [Google Scholar] [CrossRef]
- Morowitz, H.J. Beginnings of Cellular Life: Metabolism Recapitulates Biogenesis; Yale University Press: London, UK, 1993. [Google Scholar]
- Leduc, S. The Mechanism of Life; The British Institute of Radiology: London, UK, 1911. [Google Scholar]
- Bleichert, F.; Botchan, M.R.; Berger, J.M. Mechanisms for initiating cellular DNA replication. Science 2017, 355, eaah6317. [Google Scholar] [CrossRef] [Green Version]
- Sojo, V.; Pomiankowski, A.; Lane, N. A Bioenergetic Basis for Membrane Divergence in Archaea and Bacteria. PLoS Biol. 2014, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koga, Y.; Kyuragi, T.; Nishihara, M.; Sone, N. Did archaeal and bacterial cells arise independently from noncellular precursors? A hypothesis stating that the advent of membrane phospholipid with enantiomeric glycerophosphate backbones caused the separation of the two lines of descent. J. Mol. Evol. 1998, 46, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Lombard, J.; López-García, P.; Moreira, D. The early evolution of lipid membranes and the three domains of life. Nat. Rev. Microbiol. 2012, 10, 507–515. [Google Scholar] [CrossRef]
- Vollmer, W.; Bertsche, U. Murein (peptidoglycan) structure, architecture and biosynthesis in Escherichia coli. Biochim. Biophys. Acta Biomembr. 2008, 1778, 1714–1734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albers, S.V.; Meyer, B.H. The archaeal cell envelope. Nat. Rev. Microbiol. 2011, 9, 414–426. [Google Scholar] [CrossRef] [PubMed]
- Martin, W.; Russell, M.J. On the origins of cells: A hypothesis for the evolutionary transitions from abiotic geochemistry to chemoautotrophic prokaryotes, and from prokaryotes to nucleated cells. Philos. Trans. R. Soc. B Biol. Sci. 2003, 358, 59–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, I.A.; Walde, P. From self-assembled vesicles to protocells. Cold Spring Harb. Perspect. Biol. 2010, 2, a002170. [Google Scholar] [CrossRef] [PubMed]
- Beeby, M.; Ferreira, J.L.; Tripp, P.; Albers, S.V.; Mitchell, D.R. Propulsive nanomachines: The convergent evolution of archaella, flagella and cilia. FEMS Microbiol. Rev. 2020, 44, 253–304. [Google Scholar] [CrossRef]
- Harrison, S.A.; Lane, N. Life as a guide to prebiotic nucleotide synthesis. Nat. Commun. 2018, 9, 18–21. [Google Scholar] [CrossRef]
- Yadav, M.; Kumar, R.; Krishnamurthy, R. Chemistry of Abiotic Nucleotide Synthesis. Chem. Rev. 2020, 120, 4766–4805. [Google Scholar] [CrossRef]
- Muchowska, K.B.; Varma, S.J.; Moran, J. Nonenzymatic Metabolic Reactions and Life’s Origins. Chem. Rev. 2020, 120, 7708–7744. [Google Scholar] [CrossRef]
- Hordijk, W.; Steel, M. Detecting autocatalytic, self-sustaining sets in chemical reaction systems. J. Theor. Biol. 2004, 227, 451–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steel, M.; Hordijk, W.; Xavier, J.C. Autocatalytic networks in biology: Structural theory and algorithms. J. R. Soc. Interface 2019, 16. [Google Scholar] [CrossRef] [Green Version]
- Hordijk, W.; Kauffman, S.A.; Steel, M. Required levels of catalysis for emergence of autocatalytic sets in models of chemical reaction systems. Int. J. Mol. Sci. 2011, 12, 3085–3101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kauffman, S.A. Autocatalytic sets of proteins. J. Theor. Biol. 1986, 119, 1–24. [Google Scholar] [CrossRef]
- Xavier, J.C.; Hordijk, W.; Kauffman, S.; Steel, M.; Martin, W.F. Autocatalytic chemical networks at the origin of metabolism. Proc. R. Soc. B Biol. Sci. 2020, 287. [Google Scholar] [CrossRef]
- Bowman, J.C.; Hud, N.V.; Williams, L.D. The Ribosome Challenge to the RNA World. J. Mol. Evol. 2015, 80, 143–161. [Google Scholar] [CrossRef]
- Frenkel-Pinter, M.; Haynes, J.W.; Mohyeldin, A.M.; Martin, C.; Sargon, A.B.; Petrov, A.S.; Krishnamurthy, R.; Hud, N.V.; Williams, L.D.; Leman, L.J. Mutually stabilizing interactions between proto-peptides and RNA. Nat. Commun. 2020, 11, 3137. [Google Scholar] [CrossRef]
- Wheat, C.G.; McManus, J.; Mottl, M.J.; Giambalvo, E. Oceanic phosphorus imbalance: Magnitude of the mid-ocean ridge flank hydrothermal sink. Geophys. Res. Lett. 2003, 30, 17–20. [Google Scholar] [CrossRef]
- Albarède, F.; Blichert-Toft, J. The terrestrial cradle of life. Orig. Life Self-Organ. Biol. Evol. 2009, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Goldford, J.E.; Hartman, H.; Smith, T.F.; Segrè, D. Remnants of an Ancient Metabolism without Phosphate. Cell 2017, 168, 1126–1134.e9. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, B.; Muhling, J.R.; Suvorova, A.; Fischer, W.W. Apatite nanoparticles in 3.46–2.46 Ga iron formations: Evidence for phosphorus-rich hydrothermal plumes on early Earth. Geology 2021, 49, 647–651. [Google Scholar] [CrossRef]
- Deamer, D. The role of lipid membranes in life’s origin. Life 2017, 7, 5. [Google Scholar] [CrossRef]
- Szostak, J.W.; Bartel, D.P.; Luisi, P.L. Synthesizing life. Nature 2001, 409, 387–390. [Google Scholar] [CrossRef] [PubMed]
- Jordan, S.F.; Rammu, H.; Zheludev, I.N.; Hartley, A.M.; Maréchal, A.; Lane, N. Promotion of protocell self-assembly from mixed amphiphiles at the origin of life. Nat. Ecol. Evol. 2019, 3, 1705–1714. [Google Scholar] [CrossRef] [PubMed]
- Price, R.; Boyd, E.S.; Hoehler, T.M.; Wehrmann, L.M.; Bogason, E.; Valtýsson, H.P.; Örlygsson, J.; Gautason, B.; Amend, J.P. Alkaline vents and steep Na+ gradients from ridge-flank basalts-Implications for the origin and evolution of life. Geology 2017, 45, 1135–1138. [Google Scholar] [CrossRef]
- Dibrova, D.V.; Galperin, M.Y.; Koonin, E.V.; Mulkidjanian, A.Y. Ancient systems of sodium/potassium homeostasis as predecessors of membrane bioenergetics. Biochemistry 2015, 80, 495–516. [Google Scholar] [CrossRef]
- Mulkidjanian, A.Y.; Galperin, M.Y.; Koonin, E.V. Co-evolution of primordial membranes and membrane proteins. Trends Biochem. Sci. 2009, 34, 206–215. [Google Scholar] [CrossRef] [Green Version]
- Mulkidjanian, A.Y.; Galperin, M.Y.; Makarova, K.S.; Wolf, Y.I.; Koonin, E.V. Evolutionary primacy of sodium bioenergetics. Biol. Direct 2008, 3, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lane, N.; Allen, J.F.; Martin, W. How did LUCA make a living? Chemiosmosis in the origin of life. BioEssays 2010, 32, 271–280. [Google Scholar] [CrossRef]
- Lakatos, I. Falsification and the Methodology of Scientific Research Programmes. Criticism and the Growth of Knowledge; Lakatos, I., Musgrave, A., Eds.; Cambridge University Press: Cambridge, UK, 1970. [Google Scholar]
- Sutherland, J.D. Opinion: Studies on the origin of life-The end of the beginning. Nat. Rev. Chem. 2017, 1, 12. [Google Scholar] [CrossRef]
- Jackson, J.B. Natural pH Gradients in Hydrothermal Alkali Vents Were Unlikely to Have Played a Role in the Origin of Life. J. Mol. Evol. 2016, 83, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, J.B. Ancient living organisms escaping from, or imprisoned in, the vents? Life 2017, 7, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ross, D.S. It is Neither Frankenstein Nor a Submarine Alkaline Vent, It is Just the Second Law. BioEssays 2018, 40, 1800149. [Google Scholar] [CrossRef] [PubMed]
- Lane, N. Proton gradients at the origin of life. BioEssays 2017, 39. [Google Scholar] [CrossRef] [Green Version]
- Hudson, R.; de Graaf, R.; Rodin, M.S.; Ohno, A.; Lane, N.; McGlynn, S.E.; Yamada, Y.M.A.; Nakamura, R.; Barge, L.M.; Braun, D.; et al. CO2 reduction driven by a pH gradient. Proc. Natl. Acad. Sci. USA 2020, 117, 22873–22879. [Google Scholar] [CrossRef]
- Nam, I.; Lee, J.K.; Nam, H.G.; Zare, R.N. Abiotic production of sugar phosphates and uridine ribonucleoside in aqueous microdroplets. Proc. Natl. Acad. Sci. USA 2017, 114, 12396–12400. [Google Scholar] [CrossRef] [Green Version]
- Sutherland, J.D. The Origin of Life—Out of the Blue. Angew. Chem. Int. Ed. 2016, 55, 104–121. [Google Scholar] [CrossRef]
- Hawkesworth, C.J.; Cawood, P.A.; Dhuime, B. The Evolution of the Continental Crust and the Onset of Plate Tectonics. Front. Earth Sci. 2020, 8. [Google Scholar] [CrossRef]
- Harrison, T.M. Hadean Earth; Springer: Cham, Switzerland, 2020; ISBN 9783030466879. [Google Scholar]
- Flament, N.; Coltice, N.; Rey, P.F. The evolution of the 87sr/86sr of marine carbonates does not constrain continental growth. Precambrian Res. 2013, 229, 177–188. [Google Scholar] [CrossRef] [Green Version]
- Keller, C.B.; Harrison, T.M. Constraining crustal silica on ancient Earth. Proc. Natl. Acad. Sci. USA 2020, 117, 21101–21107. [Google Scholar] [CrossRef]
- Johnson, B.W.; Wing, B.A. Limited Archaean continental emergence reflected in an early Archaean 18O-enriched ocean. Nat. Geosci. 2020, 13, 243–248. [Google Scholar] [CrossRef]
- Harrison, T.M. The hadean crust: Evidence from >4 Ga zircons. Annu. Rev. Earth Planet. Sci. 2009, 37, 479–505. [Google Scholar] [CrossRef] [Green Version]
- Arndt, N.T.; Nisbet, E.G. Processes on the young earth and the habitats of early life. Annu. Rev. Earth Planet. Sci. 2012, 40, 521–549. [Google Scholar] [CrossRef] [Green Version]
- Westall, F.; Brack, A. The Importance of Water for Life. Space Sci. Rev. 2018, 214, 50. [Google Scholar] [CrossRef]
- Ball, P. Water as an active constituent in cell biology. Chem. Rev. 2008, 108, 74–108. [Google Scholar] [CrossRef] [PubMed]
- Frenkel-Pinter, M.; Rajaei, V.; Glass, J.B.; Hud, N.V.; Williams, L.D. Water and Life: The Medium is the Message. J. Mol. Evol. 2021, 89, 2–11. [Google Scholar] [CrossRef]
- De Duve, C. Vital Dust: Life as a Cosmic Imperative; Basic Books: New York, NY, USA, 1995. [Google Scholar]
- Branscomb, E.; Russell, M.J. On the beneficent thickness of water. Interface Focus 2019, 9. [Google Scholar] [CrossRef] [PubMed]
- Ball, P. Water: Water—An enduring mystery. Nature 2008, 452, 291–292. [Google Scholar] [CrossRef]
- Saha, R.; Pohorille, A.; Chen, I.A. Molecular Crowding and Early Evolution. Orig. Life Evol. Biosph. 2014, 44, 319–324. [Google Scholar] [CrossRef]
- Ellis, R.J. Macromolecular crowding: An important but neglected aspect of the intracellular environment. Curr. Opin. Struct. Biol. 2001, 11, 114–119. [Google Scholar] [CrossRef]
- Griffith, E.C.; Vaida, V. In situ observation of peptide bond formation at the water-air interface. Proc. Natl. Acad. Sci. USA 2012, 109, 15697–15701. [Google Scholar] [CrossRef] [Green Version]
- Morasch, M.; Liu, J.; Dirscherl, C.F.; Ianeselli, A.; Kühnlein, A.; Le Vay, K.; Schwintek, P.; Islam, S.; Corpinot, M.K.; Scheu, B.; et al. Heated gas bubbles enrich, crystallize, dry, phosphorylate and encapsulate prebiotic molecules. Nat. Chem. 2019, 11, 779–788. [Google Scholar] [CrossRef] [PubMed]
- White, L.M.; Shibuya, T.; Vance, S.D.; Christensen, L.E.; Bhartia, R.; Kidd, R.; Hoffmann, A.; Stucky, G.D.; Kanik, I.; Russell, M.J. Simulating serpentinization as it could apply to the emergence of life using the JPL hydrothermal reactor. Astrobiology 2020, 20, 307–326. [Google Scholar] [CrossRef]
- Barge, L.M.; Flores, E.; VanderVelde, D.G.; Weber, J.M.; Baum, M.M.; Castonguay, A. Effects of Geochemical and Environmental Parameters on Abiotic Organic Chemistry Driven by Iron Hydroxide Minerals. J. Geophys. Res. Planets 2020, 125, e2020JE006423. [Google Scholar] [CrossRef]
- Flores, E.; Martinez, E.; Rodriguez, L.E.; Weber, J.M.; Khodayari, A.; Vandervelde, D.G.; Barge, L.M. Effects of Amino Acids on Phosphate Adsorption onto Iron (Oxy)hydroxide Minerals under Early Earth Conditions. ACS Earth Space Chem. 2021. [Google Scholar] [CrossRef]
- Lu, G.S.; LaRowe, D.E.; Fike, D.A.; Druschel, G.K.; Gilhooly, W.P.; Price, R.E.; Amend, J.P. Bioenergetic characterization of a shallow-sea hydrothermal vent system: Milos Island, Greece. PLoS ONE 2020, 15, e0234175. [Google Scholar] [CrossRef] [PubMed]
- Edmond, J.M.; Von Damm, K.L.; McDuff, R.E.; Measures, C.I. Chemistry of hot springs on the East Pacific Rise and their effluent dispersal. Nature 1982, 297, 187–191. [Google Scholar] [CrossRef]
- Jannasch, H.W. Review Lecture-The chemosynthetic support of life and the microbial diversity at deep-sea hydrothermal vents. Proc. R. Soc. London Ser. B. Biol. Sci. 1985, 225, 277–297. [Google Scholar]
- Baross, J.A.; Hoffman, S. Submarine hydrothermal vents and associated gradient environments as sites for the origin and evolution of life. Orig. Life 1985, 15, 327–345. [Google Scholar] [CrossRef]
- Miller, S.L.; Bada, J.L. Submarine hot springs and the origin of life. Nature 1988, 334, 609–611. [Google Scholar] [CrossRef]
- Bada, J.L.; Lazcano, A. Some like it hot, but not the first biomolecules. Science 2002, 296, 1982–1983. [Google Scholar] [CrossRef]
- Boyce, A.J.; Coleman, M.L.; Russell, M.J. Formation of fossil hydrothermal chimneys and mounds from Silvermines, Ireland. Nature 1983, 306, 545–550. [Google Scholar] [CrossRef]
- Russell, M. Fist Life. Am. Sci. 2006, 94, 32–39. [Google Scholar] [CrossRef]
- Kelley, D.S.; Karson, J.A.; Blackman, D.K.; Früh-Green, G.L.; Butterfield, D.A.; Lilley, M.D.; Olson, E.J.; Schrenk, M.O.; Roe, K.K.; Lebon, G.T.; et al. An off-axis hydrothermal vent field near the mid-Atlantic ridge at 30° n. Nature 2001, 412, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Whitney, D.L.; Teyssier, C.; Rey, P.; Roger Buck, W. Continental and oceanic core complexes. Bull. Geol. Soc. Am. 2013, 125, 273–298. [Google Scholar] [CrossRef]
- Ildefonse, B.; Blackman, D.K.; John, B.E.; Ohara, Y.; Miller, D.J.; MacLeod, C.J.; Abe, N.; Abratis, M.; Andal, E.S.; Andréani, M.; et al. Oceanic core complexes and crustal accretion at slow-spreading ridges. Geology 2007, 35, 623–626. [Google Scholar] [CrossRef] [Green Version]
- Blackmann, D.K.; Karson, J.A.; Kelley, D.S.; Cann, J.R.; Früh-Green, G.L.; Gee, J.S.; Hurst, S.D.; John, B.E.; Morgan, J.; Nooner, S.L.; et al. Geology of the Atlantis Massif (Mid-Atlantic Ridge, 30°N): Implications for the evolution of an ultramafic oceanic core complex. Mar. Geophys. Res. 2002, 23, 443–469. [Google Scholar] [CrossRef]
- Cardoso, S.S.S.; Cartwright, J.H.E. On the differing growth mechanisms of black-smoker and Lost City-type hydrothermal vents. Proc. R. Soc. A Math. Phys. Eng. Sci. 2017, 473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ludwig, K.A.; Kelley, D.S.; Butterfield, D.A.; Nelson, B.K.; Früh-Green, G. Formation and evolution of carbonate chimneys at the Lost City Hydrothermal Field. Geochim. Cosmochim. Acta 2006, 70, 3625–3645. [Google Scholar] [CrossRef]
- Denny, A.R.; Kelley, D.S.; Früh-Green, G.L. Geologic evolution of the Lost City Hydrothermal Field. Geochem. Geophys. Geosyst. 2015, 17, 375–394. [Google Scholar] [CrossRef] [Green Version]
- Kelley, D.S.; Karson, J.A.; Früh-Green, G.L.; Yoerger, D.R.; Shank, T.M.; Butterfield, D.A.; Hayes, J.M.; Schrenk, M.O.; Olson, E.J.; Proskurowski, G.; et al. A serpentinite-hosted ecosystem: The Lost City hydrothermal field. Science 2005, 307, 1428–1434. [Google Scholar] [CrossRef]
- Budin, I.; Szostak, J.W. Expanding roles for diverse physical phenomena during the origin of life. Annu. Rev. Biophys. 2010, 39, 245–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, W.F. Older Than Genes: The Acetyl CoA Pathway and Origins. Front. Microbiol. 2020, 11, 817. [Google Scholar] [CrossRef] [PubMed]
- Varma, S.J.; Muchowska, K.B.; Chatelain, P.; Moran, J. Native iron reduces CO2 to intermediates and end-products of the acetyl-CoA pathway. Nat. Ecol. Evol. 2018, 2, 1019–1024. [Google Scholar] [CrossRef]
- Preiner, M.; Igarashi, K.; Muchowska, K.B.; Yu, M.; Varma, S.J.; Kleinermanns, K.; Nobu, M.K.; Kamagata, Y.; Tüysüz, H.; Moran, J.; et al. A hydrogen-dependent geochemical analogue of primordial carbon and energy metabolism. Nat. Ecol. Evol. 2020, 4, 534–542. [Google Scholar] [CrossRef] [PubMed]
- Baaske, P.; Weinert, F.M.; Duhr, S.; Lemke, K.H.; Russell, M.J.; Braun, D. Extreme accumulation of nucleotides in simulated hydrothermal pore systems. Proc. Natl. Acad. Sci. USA 2007, 104, 9346–9351. [Google Scholar] [CrossRef] [Green Version]
- Salditt, A.; Keil, L.M.R.; Horning, D.P.; Mast, C.B.; Joyce, G.F.; Braun, D. Thermal Habitat for RNA Amplification and Accumulation. Phys. Rev. Lett. 2020, 125, 48104. [Google Scholar] [CrossRef] [PubMed]
- Duhr, S.; Braun, D. Thermophoretic depletion follows Boltzmann distribution. Phys. Rev. Lett. 2006, 96, 168301. [Google Scholar] [CrossRef] [Green Version]
- Joyce, G.F.; Szostak, J.W. Protocells and RNA self-replication. Cold Spring Harb. Perspect. Biol. 2018, 10. [Google Scholar] [CrossRef] [Green Version]
- Ludwig, K.A.; Shen, C.C.; Kelley, D.S.; Cheng, H.; Edwards, R.L. U-Th systematics and 230Th ages of carbonate chimneys at the Lost City Hydrothermal Field. Geochim. Cosmochim. Acta 2011, 75, 1869–1888. [Google Scholar] [CrossRef]
- Schrenk, M.O.; Brazelton, W.J.; Lang, S.Q. Serpentinization, carbon, and deep life. Rev. Mineral. Geochem. 2013, 75, 575–606. [Google Scholar] [CrossRef] [Green Version]
- Mullineaux, L.S.; Metaxas, A.; Beaulieu, S.E.; Bright, M.; Gollner, S.; Grupe, B.M.; Herrera, S.; Kellner, J.B.; Levin, L.A.; Mitarai, S.; et al. Exploring the ecology of deep-sea hydrothermal vents in a metacommunity framework. Front. Mar. Sci. 2018, 4. [Google Scholar] [CrossRef] [Green Version]
- Sleep, N.H. Plate Tectonics through Time. Treatise Geophys. 2007, 9, 145–169. [Google Scholar]
- Sleep, N.H. The Hadean-Archaean Environment. Cold Spring Harb. Perspect. Biol. 2010, 2, a002527. [Google Scholar] [CrossRef] [Green Version]
- Davies, G.F. Dynamics of the Hadean and Archaean Mantle. Dev. Precambrian Geol. 2007, 15, 61–73. [Google Scholar] [CrossRef]
- Shibuya, T.; Yoshizaki, M.; Sato, M.; Shimizu, K.; Nakamura, K.; Omori, S.; Suzuki, K.; Takai, K.; Tsunakawa, H.; Maruyama, S. Hydrogen-rich hydrothermal environments in the Hadean ocean inferred from serpentinization of komatiites at 300 °C and 500 bar. Prog. Earth Planet. Sci. 2015, 2. [Google Scholar] [CrossRef] [Green Version]
- Marshall, C.R. Five palaeobiological laws needed to understand the evolution of the living biota. Nat. Ecol. Evol. 2017, 1, 165. [Google Scholar] [CrossRef] [PubMed]
- Raup, D.M.; Valentine, J.W. Multiple origins of life. Proc. Natl. Acad. Sci. USA 1983, 80, 2981–2984. [Google Scholar] [CrossRef] [Green Version]
- Wachtershauser, G. Evolution of the first metabolic cycles. Proc. Natl. Acad. Sci. USA 1990, 87, 200–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camprubi, E.; Jordan, S.F.; Vasiliadou, R.; Lane, N. Iron catalysis at the origin of life. IUBMB Life 2017, 69, 373–381. [Google Scholar] [CrossRef]
- Bonfio, C.; Valer, L.; Scintilla, S.; Shah, S.; Evans, D.J.; Jin, L.; Szostak, J.W.; Sasselov, D.D.; Sutherland, J.D.; Mansy, S.S. UV-light-driven prebiotic synthesis of iron-sulfur clusters. Nat. Chem. 2017, 9, 1229–1234. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Bartlett, S.; Yung, Y.L. Quantifying mineral-ligand structural similarities: Bridging the geological world of minerals with the biological world of enzymes. Life 2020, 10, 338. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Kitadai, N.; Nakamura, R. Chemical diversity of metal sulfide minerals and its implications for the origin of life. Life 2018, 8, 46. [Google Scholar] [CrossRef] [Green Version]
- Schäfer, G.; Engelhard, M.; Müller, V. Bioenergetics of the Archaea. Microbiol. Mol. Biol. Rev. 1999, 63, 570–620. [Google Scholar] [CrossRef] [Green Version]
- Wächtershäuser, G. Before enzymes and templates: Theory of surface metabolism. Microbiol. Rev. 1988, 52, 452–484. [Google Scholar] [CrossRef] [PubMed]
- Knowles, J.R. Enzyme-catalyzed phosphoryl transfer reactions. Annu. Rev. Biochem. 1980, 49, 877–919. [Google Scholar] [CrossRef]
- Semenov, S.N.; Kraft, L.J.; Ainla, A.; Zhao, M.; Baghbanzadeh, M.; Campbell, V.E.; Kang, K.; Fox, J.M.; Whitesides, G.M. Autocatalytic, bistable, oscillatory networks of biologically relevant organic reactions. Nature 2016, 537, 656–660. [Google Scholar] [CrossRef] [Green Version]
- Müller, V.; Chowdhury, N.P.; Basen, M. Electron bifurcation: A long-hidden energy-coupling mechanism. Annu. Rev. Microbiol. 2018, 72, 331–353. [Google Scholar] [CrossRef]
- Herrmann, G.; Jayamani, E.; Mai, G.; Buckel, W. Energy conservation via electron-transferring flavoprotein in anaerobic bacteria. J. Bacteriol. 2008, 190, 784–791. [Google Scholar] [CrossRef] [Green Version]
- Whicher, A.; Camprubi, E.; Pinna, S.; Herschy, B.; Lane, N. Acetyl Phosphate as a Primordial Energy Currency at the Origin of Life. Orig. Life Evol. Biosph. 2018, 48, 159–179. [Google Scholar] [CrossRef] [Green Version]
- Lane, N.; Martin, W.F. The origin of membrane bioenergetics. Cell 2012, 151, 1406–1416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segré, D.; Ben-Eli, D.; Deamer, D.W.; Lancet, D. The Lipid World. Orig. Life Evol. Biosph. 2001, 31, 119–145. [Google Scholar] [CrossRef]
- Rajamani, S.; Vlassov, A.; Benner, S.; Coombs, A.; Olasagasti, F.; Deamer, D. Lipid-assisted synthesis of RNA-like polymers from mononucleotides. Orig. Life Evol. Biosph. 2008, 38, 57–74. [Google Scholar] [CrossRef] [PubMed]
- Frenkel-Pinter, M.; Samanta, M.; Ashkenasy, G.; Leman, L.J.; Örlygsson, J. Prebiotic Peptides: Molecular Hubs in the Origin of Life. Chem. Rev. 2020, 120, 4707–4765. [Google Scholar] [CrossRef]
- Tran, Q.P.; Adam, Z.R.; Fahrenbach, A.C. Prebiotic reaction networks in water. Life 2020, 10, 352. [Google Scholar] [CrossRef]
- Gilbert, W. The RNA world. Nature 1986, 319, 618. [Google Scholar] [CrossRef]
- Pressman, A.; Blanco, C.; Chen, I.A. The RNA world as a model system to study the origin of life. Curr. Biol. 2015, 25, R953–R963. [Google Scholar] [CrossRef] [Green Version]
- Tjhung, K.F.; Shokhirev, M.N.; Horning, D.P.; Joyce, G.F. An RNA polymerase ribozyme that synthesizes its own ancestor. Proc. Natl. Acad. Sci. USA 2020, 117, 2906–2913. [Google Scholar] [CrossRef]
- Eigen, M. Selforganization of matter and the evolution of biological macromolecules. Naturwissenschaften 1971, 58, 465–523. [Google Scholar] [CrossRef]
- Robertson, M.P.; Joyce, G.F. The origins of the RNA world. Cold Spring Harb. Perspect. Biol. 2012, 4, a003608. [Google Scholar] [CrossRef] [PubMed]
- Ban, N.; Nissen, P.; Hansen, J.; Moore, P.B.; Steitz, T.A. The complete atomic structure of the large ribosomal subunit at 2.4 Åresolution. Science 2000, 289, 905–920. [Google Scholar] [CrossRef]
- Fedor, M.J.; Williamson, J.R. The catalytic diversity of RNAs. Nat. Rev. Mol. Cell Biol. 2005, 6, 399–412. [Google Scholar] [CrossRef]
- Koonin, E.V.; Novozhilov, A.S. Origin and Evolution of the Universal Genetic Code. Annu. Rev. Genet. 2017, 51, 45–62. [Google Scholar] [CrossRef] [PubMed]
- Koonin, E.V. Frozen accident pushing 50: Stereochemistry, expansion, and chance in the evolution of the genetic code. Life 2017, 7, 22. [Google Scholar] [CrossRef] [PubMed]
- Wolf, Y.I.; Koonin, E.V. On the origin of the translation system and the genetic code in the RNA world by means of natural selection, exaptation, and subfunctionalization. Biol. Direct 2007, 2. [Google Scholar] [CrossRef] [Green Version]
- Anfinsen, C.B. Principles that govern protein folding. Science 1973, 181, 223–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perutz, M.F. Introductory lecture: What are enzyme structures telling us? Faraday Discuss. 1992, 93, 1–11. [Google Scholar] [CrossRef]
- Perutz, M.F. Co-chairman’s remarks: Before the double helix. Gene 1993, 135, 9–13. [Google Scholar] [CrossRef]
- Petrov, A.S.; Gulen, B.; Norris, A.M.; Kovacs, N.A.; Bernier, C.R.; Lanier, K.A.; Fox, G.E.; Harvey, S.C.; Wartell, R.M.; Hud, N.V.; et al. History of the ribosome and the origin of translation. Proc. Natl. Acad. Sci. USA 2015, 112, 15396–15401. [Google Scholar] [CrossRef] [Green Version]
- Boyer, P.D. A research journey with ATP synthase. J. Biol. Chem. 2002, 277, 39045–39061. [Google Scholar] [CrossRef] [Green Version]
- Grüber, G.; Manimekalai, M.S.S.; Mayer, F.; Müller, V. ATP synthases from archaea: The beauty of a molecular motor. Biochim. Biophys. Acta Bioenerg. 2014, 1837, 940–952. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, P. Coupling of phosphorylation to electron and hydrogen transfer by a chemi-osmotic type of mechanism. Nature 1961, 191, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Krissansen-Totton, J.; Arney, G.N.; Catling, D.C. Constraining the climate and ocean pH of the early Earth with a geological carbon cycle model. Proc. Natl. Acad. Sci. USA 2018, 115, 4105–4110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lane, N. Why are cells powered by proton gradients? Nat. Educ. 2010, 3, 18. [Google Scholar]
- Bowman, J.C.; Petrov, A.S.; Frenkel-Pinter, M.; Penev, P.I.; Williams, L.D. Root of the Tree: The Significance, Evolution, and Origins of the Ribosome. Chem. Rev. 2020, 120, 4848–4878. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Wang, J.; Wang, J.; Wang, J.; Li, Y. Estimate of the sequenced proportion of the global prokaryotic genome. Microbiome 2020, 8, 134. [Google Scholar] [CrossRef]
- Koonin, E.V.; Makarova, K.S.; Wolf, Y.I. Evolution of Microbial Genomics: Conceptual Shifts over a Quarter Century. Trends Microbiol. 2021, 29, 582–592. [Google Scholar] [CrossRef]
- Mende, D.R.; Letunic, I.; Maistrenko, O.M.; Schmidt, T.S.B.; Milanese, A.; Paoli, L.; Hernández-Plaza, A.; Orakov, A.N.; Forslund, S.K.; Sunagawa, S.; et al. ProGenomes2: An improved database for accurate and consistent habitat, taxonomic and functional annotations of prokaryotic genomes. Nucleic Acids Res. 2020, 48, D621–D625. [Google Scholar] [CrossRef]
- Tonkin-Hill, G.; MacAlasdair, N.; Ruis, C.; Weimann, A.; Horesh, G.; Lees, J.A.; Gladstone, R.A.; Lo, S.; Beaudoin, C.; Floto, R.A.; et al. Producing polished prokaryotic pangenomes with the Panaroo pipeline. Genome Biol. 2020, 21, 180. [Google Scholar] [CrossRef]
- Howe, K.L.; Contreras-Moreira, B.; De Silva, N.; Maslen, G.; Akanni, W.; Allen, J.; Alvarez-Jarreta, J.; Barba, M.; Bolser, D.M.; Cambell, L.; et al. Ensembl Genomes 2020-enabling non-vertebrate genomic research. Nucleic Acids Res. 2020, 48, D689–D695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fialho, D.M.; Roche, T.P.; Hud, N.V. Prebiotic Syntheses of Noncanonical Nucleosides and Nucleotides. Chem. Rev. 2020, 120, 4806–4830. [Google Scholar] [CrossRef]
- Ponce-Salvatierra, A.; Wawrzyniak-Turek, K.; Steuerwald, U.; Höbartner, C.; Pena, V. Crystal structure of a DNA catalyst. Nature 2016, 529, 231–234. [Google Scholar] [CrossRef] [PubMed]
- Leu, K.; Obermayer, B.; Rajamani, S.; Gerland, U.; Chen, I.A. The prebiotic evolutionary advantage of transferring genetic information from RNA to DNA. Nucleic Acids Res. 2011, 39, 8135–8147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polanyi, M. Personal Knowledge: Towards a Post-Critical Philosophy; University of Chicago Press: Chicago, IL, USA, 1958. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Criterion |
|---|
| Bottom-up criteria—initial conditions |
| 1. OoLoE initiated with permeable containment in an environment with pervasive and steady energy and material flows. |
| 2. Probability of life—cradle of life was long-lived and/or geographically extensive. |
| 3. Integration of developing metabolic pathways began from the outset. |
| 4. Spatial heterogeneity was present from the outset. |
| Accommodation of top-down data |
| 5. Full cellular autonomy and motility occurred post-LUCA. |
| 6. Minimization of the number of inferred non-analog to analog transitions (see text). |
| 7. Origin of complex integrated metabolism before ribozymes and enzymes, made possible by mineral and metal ion catalysts and energy storage molecules. |
| Additional Criterion |
| 8. Scenario offers unexpected insights and generates a wide range of predictions. |
| Property | Attractiveness of the Microchamber Complexes |
|---|---|
| Cell-Like Boundary Conditions | |
| 1. Delineation of inside and outside | Microchamber walls provided protection from the external environment, while being permeable to energy and material flows. |
| 2. Energy flow | The chemical energy flowing through the microchambers, H2, is used by the most primitive carbon fixation pathway, the acetyl-CoA pathway. |
| 3. Material flow | Abundant single-carbon molecules, including CO2 which is used in the acetyl-CoA pathway, are available. |
| 4. Ease of concentration | The tiny chambers in conjunction with the thermal gradient concentrates molecules (thermophoresis). |
| 5. Little precursor material needed | The tiny spaces mean that relatively little prebiotic material would be needed to produce high concentrations of prebiotic molecules. |
| Probability of Life Originating | |
| 6. Temporally long-lived | Hydrothermal fields are estimated to have a lifetime of ~105 years (1012 s). On the Hadean seafloor they may have been even longer lived. |
| 7. Numerous microchambers in a complex | The sub-millimeter size of the chambers translates to some 1011 interconnected microchambers/m3 per vent. |
| 8. Many microchamber complexes | Hadean serpentinization may have occurred over much of the seafloor, thus over the (perhaps) 10s to 100s of millions of years between the formation of the sea floor and the origin of life, the number of potential cradles of life may have been enormous. |
| Establishing Biochemical Pathways | |
| 9. Ample prebiotic catalysts | Abundant Fe(Ni)S minerals present, some of which would have served as excellent prebiotic catalysts. |
| 10. Varying degrees of separation/connectedness/spatial heterogeneity | The 1011 variably interconnected microchambers/m3 translates into a vast range of subtly different geochemical conditions and spatial heterogeneity, facilitating complex prebiotic and early biotic synthesis. |
| Accounts for Top-Down Observations | |
| 11. Accompanying pH gradient offers explanation for the origin of chemiosmosis | Chemiosmotic generation of ATP is central to most organisms’ energy production but is hard to explain. The presence of an extremal pH (and possibly Na+) gradient across the microchamber complex means that the flow of protons was integral to the proposed cradle of life and means that ATP synthase could have functioned without the electron transport chain. |
| 12. Explains Bacterial and Archaeal differences | Includes the evidence that the two lineages became fully cellularized and mobile indpendently (see Section 8.9). |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brunk, C.F.; Marshall, C.R. ‘Whole Organism’, Systems Biology, and Top-Down Criteria for Evaluating Scenarios for the Origin of Life. Life 2021, 11, 690. https://doi.org/10.3390/life11070690
Brunk CF, Marshall CR. ‘Whole Organism’, Systems Biology, and Top-Down Criteria for Evaluating Scenarios for the Origin of Life. Life. 2021; 11(7):690. https://doi.org/10.3390/life11070690
Chicago/Turabian StyleBrunk, Clifford F., and Charles R. Marshall. 2021. "‘Whole Organism’, Systems Biology, and Top-Down Criteria for Evaluating Scenarios for the Origin of Life" Life 11, no. 7: 690. https://doi.org/10.3390/life11070690