
A Novel Dipeptidyl Peptidase-4 Inhibitor DA-1229 Ameliorates Tubulointerstitial Fibrosis in Cyclosporine Nephrotoxicity in Mice

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Studies
2.2. DPP-4 Activity Assay
2.3. Histological and Immunohistochemical Analysis
2.4. Analysis of Gene Expression by Real-Time Quantitative PCR
2.5. Protein Extraction and Western Blot Analysis
2.6. Statistical Analysis
3. Results
3.1. Physical and Biochemical Parameters of Experimental Animals
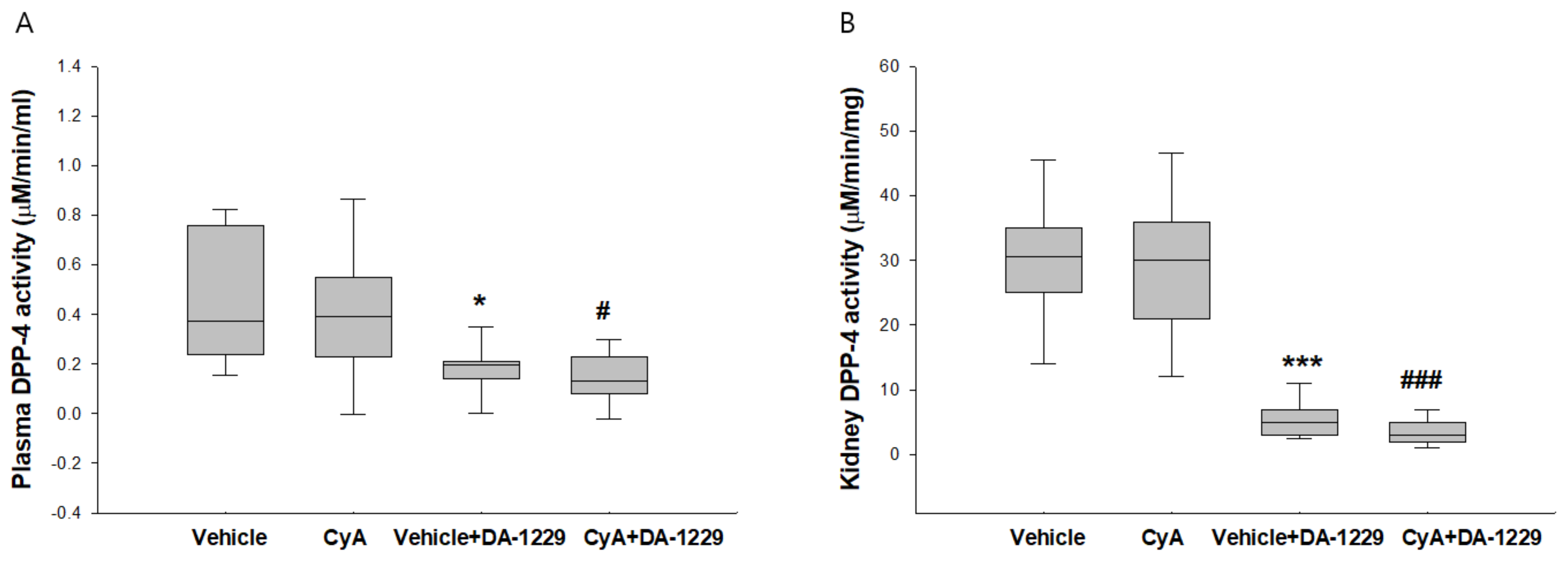
3.2. Effects of DA-1229 on DPP-4 Activity in the Plasma and Kidney Tissue
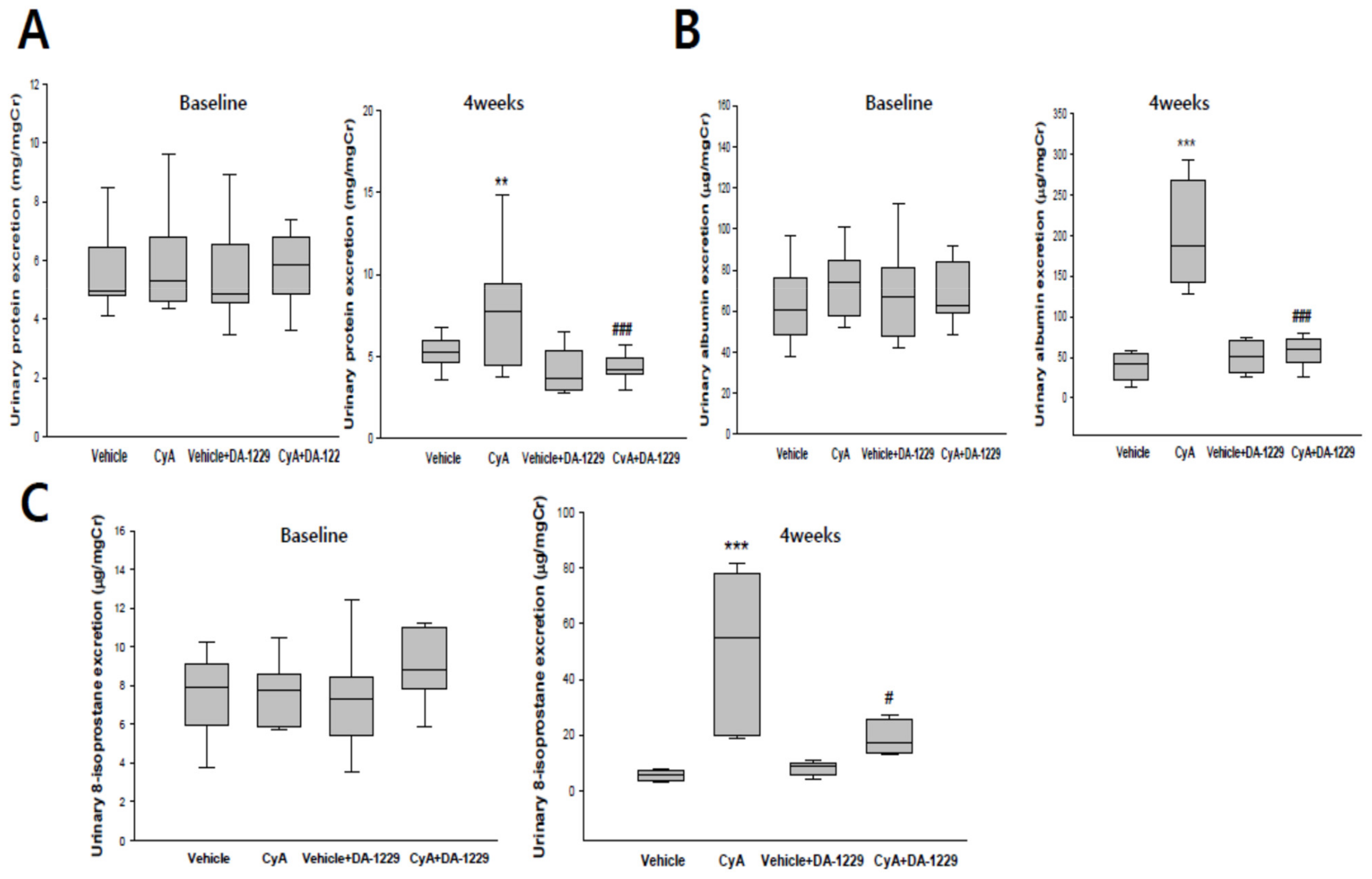
3.3. Effects of DA-1229 on Urinary Excretion of Protein, Albumin, and 8-Isoprostane
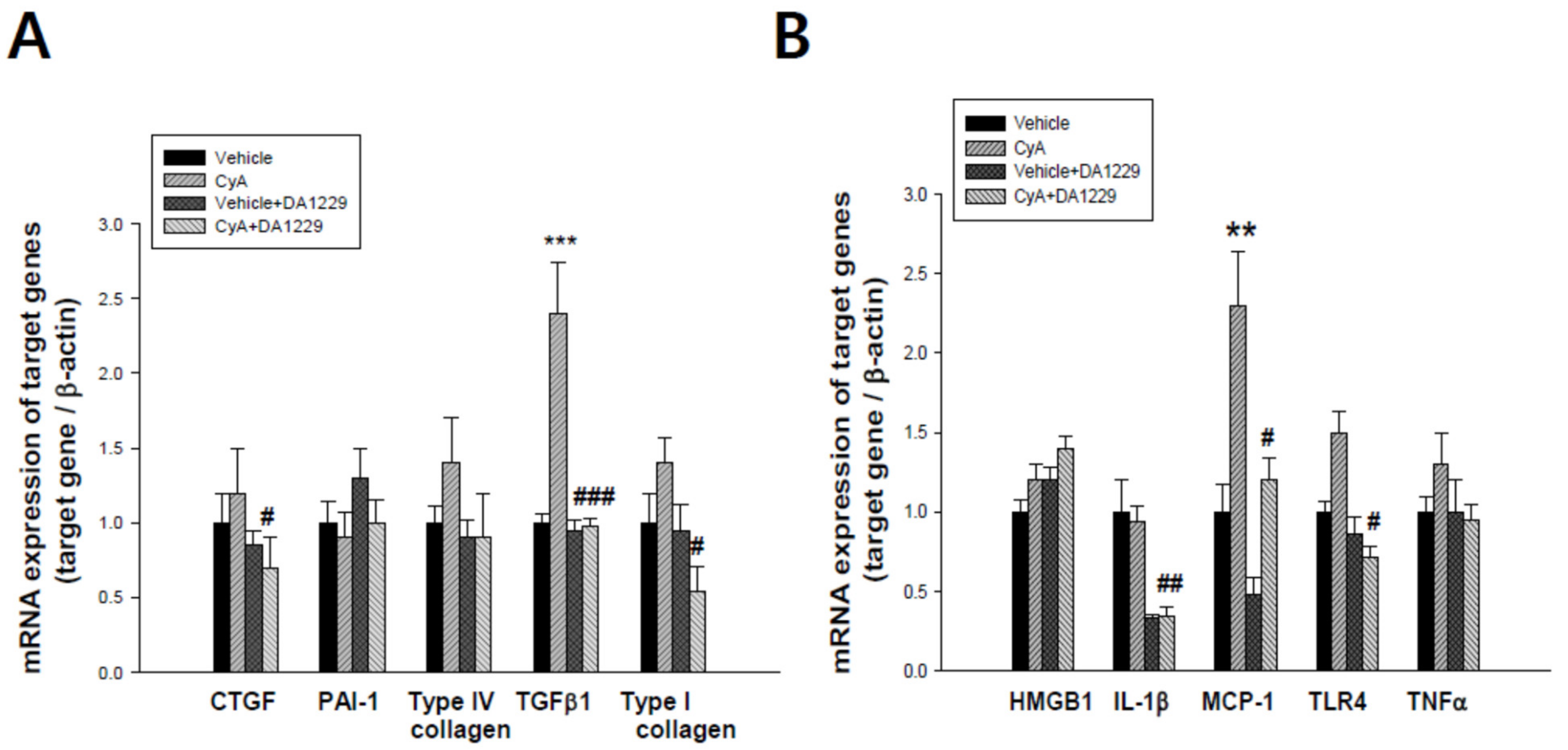
3.4. Effects of DA-1229 on Gene Expression Related to Inflammation and Fibrosis
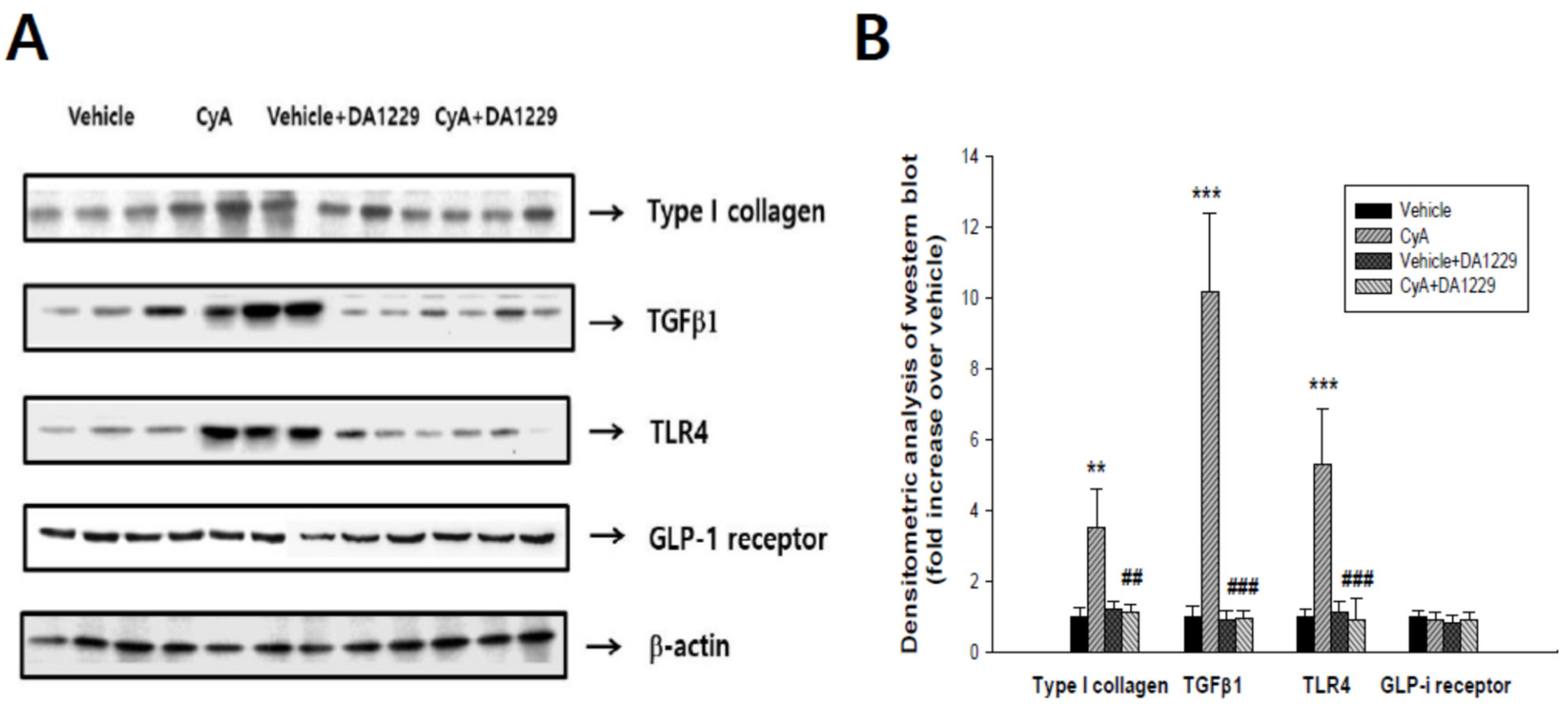
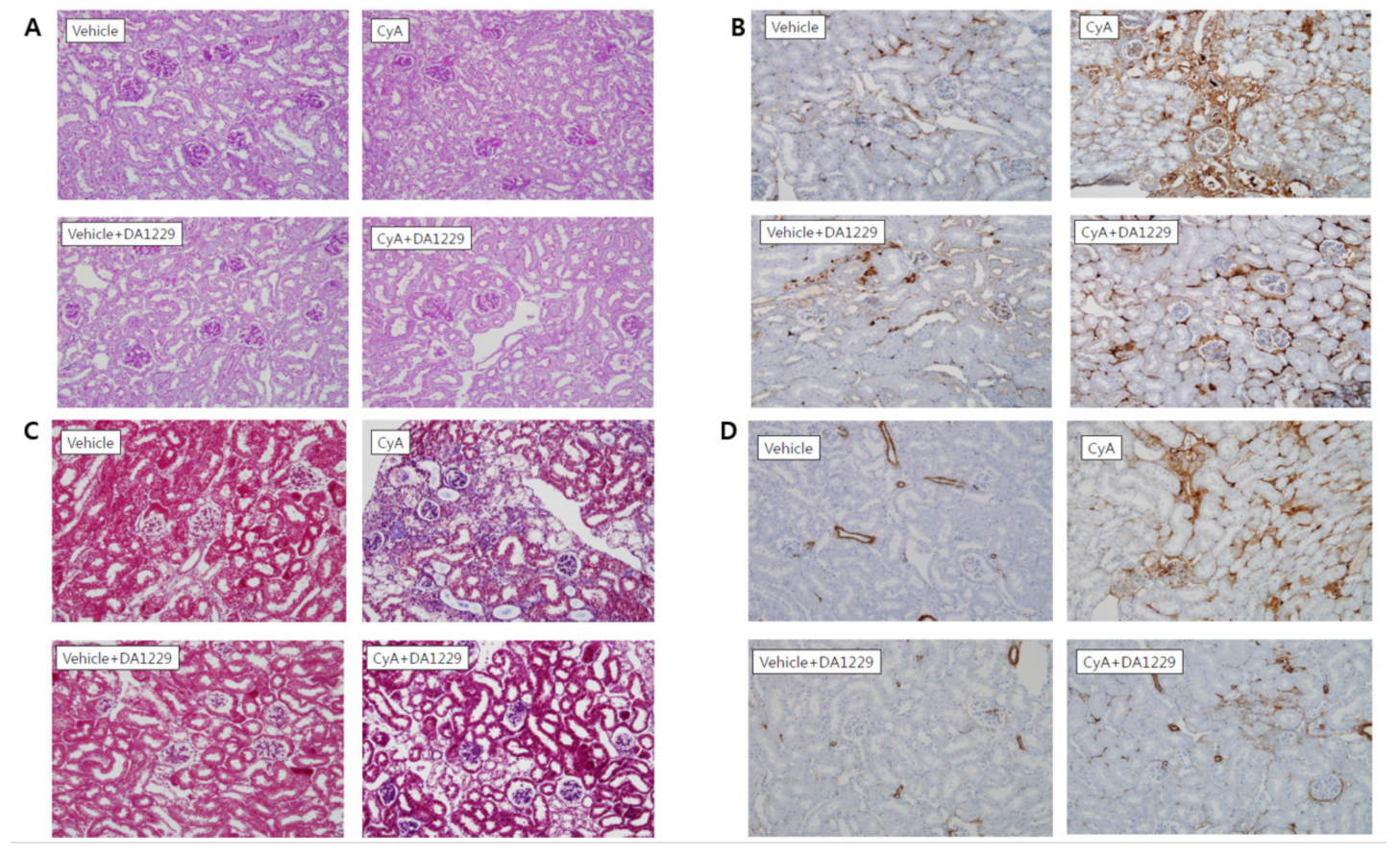
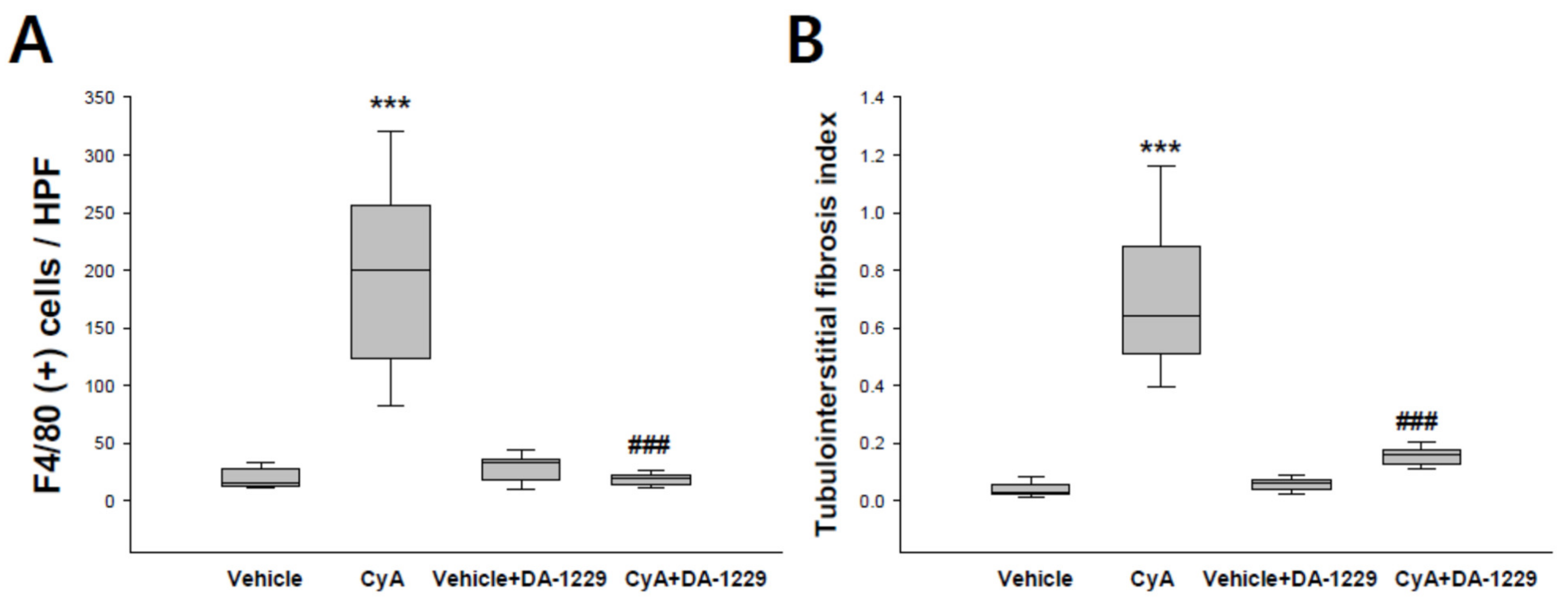
3.5. Effects of DA1229 on Renal Structural Change
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mulvihill, E.E.; Drucker, D.J. Pharmacology, physiology, and mechanisms of action of dipeptidyl peptidase-4 inhibitors. Endocr. Rev. 2014, 35, 992–1019. [Google Scholar] [CrossRef] [Green Version]
- Haluzík, M.; Frolík, J.; Rychlík, I. Renal effects of DPP-4 inhibitors: A focus on microalbuminuria. Int. J. Endocrinol. 2013, 2013, 895102. [Google Scholar] [CrossRef] [Green Version]
- Mentlein, R. Dipeptidyl-peptidase IV (CD26)-role in the inactivation of regulatory peptides. Regul. Pept. 1999, 85, 9–24. [Google Scholar] [CrossRef]
- Zhong, J.; Rao, X.; Rajagopalan, S. An emerging role of dipeptidyl peptidase 4 (DPP4) beyond glucose control: Potential implications in cardiovascular disease. Atherosclerosis 2013, 226, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Ohnuma, K.; Hosono, O.; Dang, N.H.; Morimoto, C. Dipeptidyl peptidase in autoimmune pathophysiology. Adv. Clin. Chem. 2011, 53, 51–84. [Google Scholar] [PubMed]
- Shingu, K.; Helfritz, A.; Kuhlmann, S.; Skowronek, M.Z.; Jacobs, R.; Schmidt, R.E.; Pabst, R.; Hörsten, S.V. Kinetics of the early recruitment of leukocyte subsets at the sites of tumor cells in the lungs: Natural killer (NK) cells rapidly attract monocytes but not lymphocytes in the surveillance of micrometastasis. Int. J. Cancer. 2002, 99, 74–81. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.J.; Xie, S.H.; Liu, Y.N.; Kim, W.; Jin, H.Y.; Park, S.K.; Shao, Y.M.; Park, T.S. Dipeptidyl peptidase IV inhibitor attenuates kidney injury in streptozotocin-induced diabetic rats. J. Pharmacol. Exp. Ther. 2012, 340, 248–255. [Google Scholar] [CrossRef]
- Alter, M.L.; Ott, I.M.; Von Websky, K.; Tsuprykov, O.; Sharkovskam, Y.; Krause-Relle, K.; Raila, J.; Henze, A.; Klein, T.; Hocher, B. DPP-4 inhibition on top of angiotensin receptor blockade offers a new therapeutic approach for diabetic nephropathy. Kidney Blood Press. Res. 2012, 36, 119–130. [Google Scholar] [CrossRef]
- Glorie, L.L.; Verhulst, A.; Matheeussen, V.; Baerts, L.; Magielse, J.; Hermans, N.; Haese, P.C.; Meester, I.D.; Beuf, A.D. DPP4 inhibition improves functional outcome after renal ischemia-reperfusion injury. Am. J. Physiol. Renal. Physiol. 2012, 303, F681–F688. [Google Scholar] [CrossRef]
- Min, H.S.; Kim, J.E.; Lee, M.H.; Song, H.K.; Kang, Y.S.; Lee, M.J.; Lee, J.E.; Kim, H.W.; Cha, J.J.; Chung, Y.Y.; et al. Dipeptidyl peptidase IV inhibitor protects against renal interstitial fibrosis in a mouse model of ureteral obstruction. Lab. Invest. 2014, 94, 598–607. [Google Scholar] [CrossRef] [Green Version]
- Nistala, R.; Habibi, J.; Aroor, A.; Sowers, J.R.; Hayden, M.R.; Meuth, A.; Knight, W.; Hancock, T.; Klein, T.; DeMarco, V.G.; et al. DPP4 Inhibition attenuates filtration barrier injury and oxidant stress in the zucker obese rat. Obesity 2014, 22, 2172–2179. [Google Scholar] [CrossRef] [Green Version]
- Jin, X.; Jin, C.; Nakamura, K.; Jin, T.; Xin, M.; Wan, Y.; Yue, X.; Jin, S.; Wang, H.; Inoue, A.; et al. Increased dipeptidyl peptidase-4 accelerates chronic stress-related thrombosis in a mouse carotid artery model. J. Hypertens. 2020, 38, 1504–1513. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, B.P.; Crajoinas, R.O.; Couto, G.K.; Davel, A.P.; Lessa, L.M.; Rossoni, L.V.; Girardi, A.C. Dipeptidyl peptidase IV inhibition attenuates blood pressure rising in young spontaneously hypertensive rats. J. Hypertens. 2011, 29, 520–528. [Google Scholar] [CrossRef] [PubMed]
- Adithan, A.; John Peter, J.S.; Hossain, M.A.; Kang, C.W.; Kim, B.; Kim, N.S.; Hwang, K.C.; Kim, J.H. Biological effects of cyclosporin A on CD3—CD161 + and CD3 + CD161 + lymphocytes. Mol. Cell. Biochem. 2019, 458, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Balakumar, P.; Chakkarwar, V.A.; Kumar, V.; Jain, A.; Reddy, J.; Singh, M. Experimental models for nephropathy. J. Renin. Angiotensin. Aldosterone. Syst. 2008, 9, 189–195. [Google Scholar] [CrossRef] [Green Version]
- Naesens, M.; Kuypers, D.R.; Sarwal, M. Calcineurin inhibitor nephrotoxicity. Clin. J. Am. Soc. Nephrol. 2009, 4, 481–508. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.K.; Chae, Y.N.; Kim, H.D.; Yang, E.K.; Cho, E.J.; Choi, S.H.; Cheong, Y.H.; Kim, H.S.; Kim, H.J.; Jo, Y.W.; et al. DA-1229, a novel and potent DPP4 inhibitor, improves insulin resistance and delays the onset of diabetes. Life Sci. 2012, 90, 21–29. [Google Scholar] [CrossRef]
- Yang, C.W.; Faulkner, G.R.; Wahba, I.M.; Christianson, T.A.; Bagby, G.C.; Jin, D.C.; Abboud, H.E.; Andoh, T.F.; Bennett, W.M. Expression of apoptosis-related genes in chronic cyclosporine nephrotoxicity in mice. Am. J. Transplant. 2002, 2, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Damiano, S.; Ciarcia, R.; Montagnaro, S.; Pagnini, U.; Garofano, T.; Capasso, G.; Florio, S.; Giordano, A. Prevention of Nephrotoxicity Induced by Cyclosporine-A: Role of Antioxidants. J. Cell. Biochem. 2015, 116, 364–369. [Google Scholar] [CrossRef]
- Bobadilla, N.A.; Gamba, G. New insights into the pathophysiology of cyclosporine nephrotoxicity: A role of aldosterone. Am. J. Physiol. Renal. Physiol. 2007, 293, F2–F9. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, T.; Watanabe, A.; Tanaka, M.; Shiota, M.; Osada-Oka, M.; Sano, S.; Yoshiyama, M.; Miura, K.; Kitajima, S.; Matsunaga, S.; et al. A dipeptidyl peptidase-4 (DPP-4) inhibitor, linagliptin, attenuates cardiac dysfunction after myocardial infarction independently of DPP-4. J. Pharmacol. Sci. 2019, 139, 112–119. [Google Scholar] [CrossRef]
- Kimura, S.; Inoguchi, T.; Yamasaki, T.; Yamato, M.; Ide, M.; Sonoda, N.; Yamada, K.; Takayanagi, R. A novel DPP-4 inhibitor teneligliptin scavenges hydroxyl radicals: In vitro study evaluated by electron spin resonance spectroscopy and in vivo study using DPP-4 deficient rats. Metabolism 2016, 65, 138–145. [Google Scholar] [CrossRef]
- Tsuprykov, O.; Ando, R.; Reichetzeder, C.; Von Websky, K.; Antonenko, V.; Sharkovska, Y.; Chaykovska, L.; Rahnenführer, J.; Hasan, A.A.; Tammen, H.; et al. The dipeptidyl peptidase inhibitor linagliptin and the angiotensin II receptor blocker telmisartan show renal benefit by different pathways in rats with 5/6 nephrectomy. Kidney. Int. 2016, 89, 1049–1061. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Guerrero, C.; Cannata-Ortiz, P.; Guerri, C.; Egido, J.; Ortiz, A.; Ramos, A.M. TLR4-mediated inflammation is a key pathogenic event leading to kidney damage and fibrosis in cyclosporine nephrotoxicity. Arch. Toxicol. 2017, 91, 1925–1939. [Google Scholar] [CrossRef] [PubMed]
- Dange, R.B.; Agarwal, D.; Masson, G.S.; Villa, J.; Wilson, B.; Nair, A.; Francis, J. Central blockade of TLR4 improves cardiac function and attenuates myocardial inflammation in angiotensin II-induced hypertension. Cardiovasc. Res. 2014, 103, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Calò, L.A.; Davis, P.A.; Giacon, B.; Pagnin, E.; Sartori, M.; Riegler, P.; Antonello, A.; Huber, W.; Semplicini, A. Oxidative Stress in Kidney Transplant Patients with Calcineurin Inhibitor–Induced Hypertension: Effect of Ramipril. J. Cardiovasc. Pharmacol. 2002, 40, 625–631. [Google Scholar] [CrossRef]
- Mega, C.; De Lemos, E.T.; Vala, H.; Fernandes, R.; Oliveira, J.; Mascarenhas-Melo, F.; Teixeira, F.; Reis, F. Diabetic nephropathy amelioration by a low-dose sitagliptin in an animal model of type 2 diabetes (Zucker diabetic fatty rat). Exp. Diabetes. Res. 2011, 2011, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizzo, M.R.; Barbieri, M.; Marfella, R.; Paolisso, G. Reduction of oxidative stress and inflammation by blunting daily acute glucose fluctuations in patients with type 2 diabetes: Role of dipeptidyl peptidase-IV inhibition. Diabetes. Care. 2012, 35, 2076–2082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ateyya, H. Amelioration of cyclosporine induced nephrotoxicity by dipeptidyl peptidase inhibitor vildagliptin. Int. Immunopharmacol. 2015, 28, 571–577. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.E.; Kim, J.E.; Lee, M.H.; Song, H.K.; Ghee, J.Y.; Kang, Y.S.; Min, H.S.; Kim, H.W.; Cha, J.J.; Han, J.Y.; et al. DA-1229, a dipeptidyl peptidase IV inhibitor, protects against renal injury by preventing podocyte damage in an animal model of progressive renal injury. Lab. Invest. 2016, 96, 547–560. [Google Scholar]
- Cha, H.G.; Lee, S.J.; Kim, H.J.; Kong, G.; Kang, G.W.; Kang, J.M. The Effects of Cyclosporine on the Generation of ROS and Extracellular Matrix Accumulation in Cultured Human Mesangial Cells. Kidney. Res. Clin. Pract. 2000, 19, 1024–1032. [Google Scholar]
- Lee, S.Y.; Kim, S.I.; Choi, M.E. Therapeutic targets for treating fibrotic kidney diseases. Transl. Res. 2015, 165, 512–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Week | Vehicle | CyA | Vehicle + DA1229 | CyA + DA1229 |
|---|---|---|---|---|---|
| Number | 4 | 10 | 10 | 10 | 10 |
| Body weight (g) | 0 | 33 (32–35) | 33 (31–34) | 33 (32–35) | 33 (32–38) |
| 4 | 37.5 (33–39) | 36 (33–38) | 38 (37–43) | 38.5 (35–44) | |
| Food intake (g/day) | 0 | 5.5 (5.1–6) | 5.0 (4.7–5.3) | 4.4 (4.2–5.1) | 5.1 (4.6–5.6) |
| 4 | 4.6 (4.1–5.7) | 3.0 (2.3–3.2) | 5.2 (4.4–5.9) | 4.7 (4.1–5.6) | |
| Water intake (g/day) | 0 | 5.4 (5.1–5.7) | 4.5 (4.4–5.1) | 4.0 (3.2–4.2) | 5.8 (4.8–7.1) |
| 4 | 5.2 (3.1–6.5) | 5.3 (2.5–7.7) | 6.9 (5.5–8.1) | 8.5 (6.6–11.1) | |
| Urine volume (mL/day) | 0 | 3.2 (2.6–4.1) | 3.3 (2.3–4.7) | 4.1 (2.9–4.7) | 3.2 (2.3–4.1) |
| 4 | 3.1 (2.6–4.1) | 3.1 (1.9–3.6) | 3.1 (1.9–5.1) | 3.5 (2.3–4.2) | |
| Serum Cr (μmol/L) | 4 | 46 (35–53) | 76 (57–82) *** | 48 (41–59) | 47 (39–52) ### |
| Active GLP-1 (pM) | 4 | 2.25 (1.56–3.14) | 2.12 (0.98–4.21) | 2.02 (1.11–4.12) | 2.22 (0.76–5.12) |
| SBP (mmHg) | 4 | 95 (79–111) | 98 (89–116) | 92 (81–110) | 102 (98–110) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Min, H.S.; Lee, J.E.; Ghee, J.Y.; Kang, Y.S.; Cha, J.J.; Han, J.Y.; Han, S.Y.; Cha, D.R. A Novel Dipeptidyl Peptidase-4 Inhibitor DA-1229 Ameliorates Tubulointerstitial Fibrosis in Cyclosporine Nephrotoxicity in Mice. Life 2021, 11, 251. https://doi.org/10.3390/life11030251
Min HS, Lee JE, Ghee JY, Kang YS, Cha JJ, Han JY, Han SY, Cha DR. A Novel Dipeptidyl Peptidase-4 Inhibitor DA-1229 Ameliorates Tubulointerstitial Fibrosis in Cyclosporine Nephrotoxicity in Mice. Life. 2021; 11(3):251. https://doi.org/10.3390/life11030251
Chicago/Turabian StyleMin, Hye Sook, Ji Eun Lee, Jung Yeon Ghee, Young Sun Kang, Jin Joo Cha, Jee Young Han, Sang Youb Han, and Dae Ryong Cha. 2021. "A Novel Dipeptidyl Peptidase-4 Inhibitor DA-1229 Ameliorates Tubulointerstitial Fibrosis in Cyclosporine Nephrotoxicity in Mice" Life 11, no. 3: 251. https://doi.org/10.3390/life11030251