
Proxy Archives Based on Marine Calcifying Organisms and the Role of Process-Based Biomineralization Concepts
1
Marine Biogeosciences, Alfred Wegener Institut—Helmholtz-Zentrum für Polar-und Meeresforschung, 27570 Bremerhaven, Germany
2
Institute of Environmental Science and Technology (ICTA), Universitat Autonoma de Barcelona, 08193 Barcelona, Spain
*
Author to whom correspondence should be addressed.
Minerals 2023, 13(4), 561; https://doi.org/10.3390/min13040561
Submission received: 13 February 2023
/
Revised: 10 April 2023
/
Accepted: 12 April 2023
/
Published: 17 April 2023
(This article belongs to the Special Issue Biomineralization of Organisms Used as Environmental Proxy Archives)
{kind=link}
Abstract
:Proxy archives based on marine calcifying organisms and their element and isotope signatures represent valuable tools in the reconstruction of past climates. Despite the fact that the underlying biomineralization processes behind the measured signatures are poorly understood, these tools work remarkably well. However, even though they work well many researchers felt the need to decipher the “black box” and understand the processes leading to the measured signatures. In this paper we assess how far we have come in understanding the biomineralization processes underpinning proxy signatures derived from marine calcifying organisms and how this understanding improved the way we use these proxy archives today. Biomineralization in the context of proxy research is an interdisciplinary field and cross-discipline communication can be challenging due to a lack of background in foreign disciplines. This often leads to misunderstanding and over- (or under-) estimation of certain concepts/methods/data. We, therefore, present a concise introduction to the topic, clarifying key concepts and their applicability to proxy interpretation.
1. Introduction
In 1951 Urey and co-workers [1] demonstrated for the first time that the temperature of past environments can be reconstructed from a chemical signature (oxygen isotopes) measured in calcareous structures formed by an organism (a belemnite, the hard part formed by an extinct species which is related to the modern squid). This pioneering work laid the foundation for a field we know today as proxy research. The advances in analytical instrumentation and the growing interest in understanding how our climate changes through time fostered the exploration of several other proxy archives (the material that is analyzed) and proxy signatures (the elemental or isotopic signature which is measured and related to an environmental parameter). In this paper we will focus on those proxy archives that are formed by calcifying marine organisms such as foraminifera, coccolithophores, corals, and mollusks.
Early on, researchers tried to understand the processes which determine the measured geochemical signatures of the measured proxy archives. Since it was clear from the beginning that not only inorganic processes but also physiological ones are involved in the formation of a biogenic mineral, Urey and co-workers coined the term “vital effects” [1]. For a long time, these vital effects have been used as a black box that has been held responsible for the difference in chemical signature between a mineral formed under purely inorganic conditions and the ones produced by an organism. Nevertheless, for a long period measured values have been explained mainly by the thermodynamic and kinetic processes known from crystal growth experiments. This slowly changed when it was realized that biogenic minerals are generally not pure crystals but complex composite materials containing organic and inorganic components [2]. From here on we will use the term biomineral for these calcified structures formed by an organism. Process-based biomineralization models developed in the context of proxy research started to combine physiological and inorganic mechanisms to understand the formation of biominerals and the measured proxy signatures.
Biomineralization research is a good example of the challenges interdisciplinary research is confronted with. On the one hand, it is obvious that processes from two completely different fields of research are involved (in this case biology and mineralogy/geochemistry); on the other hand, the view of a process is often dominated by the background of the researcher involved, oversimplifying aspects of the other fields. Put bluntly, biologists tend to see biomineralization as mainly biology with some geochemistry and geoscientists see it as mainly geochemistry with some biology. In the view of the authors, one being a mineralogist, the other a cell biologist, working together on this topic for many years, much misunderstanding in the field often originates from such unbalanced understanding of fundamental concepts. For example, the fundamental terms intra- and extracellular, absolutely central to understanding any process-based biomineralization model, are often fuzzy or meaningless to geochemists. Discussions with many colleagues working in the field showed that everybody has a more or less good “feeling” of the functioning of the processes applied but often crucial aspects are missing.
The aim of this paper is, therefore, not to present a review on biomineralization models or to judge them but to clarify fundamental concepts of cell biology and crystal growth that are often misunderstood when one’s own background is in a different field of research. In addition, we will analyze if and how process-based biomineralization models changed how proxy archives are used.
2. Background
The aim of this chapter is to explain briefly the fundamental definitions and concepts of cell biology needed in the context of process-based biomineralization models which are used to explain proxy signatures of calcifying organisms. These are basic concepts every biology student learns early on but are often enigmatic to readers from other fields of research. In the second part, the same will be carried out with general concepts of the field of crystal growth and element incorporation. In the next section, we will present the central feature of process-based biomineralization models that result from the combination of fundamental cell biological mechanisms with the ones known from crystal growth. Our aim is not to explain the biomineralization process for a certain species but rather to explain the absolutely fundamental processes that are valid for various calcifiers ranging from unicellular organisms such as foraminifera and coccolithophores to multicellular organisms such as mollusks and corals.
2.1. Fundamentals of Cell Biology
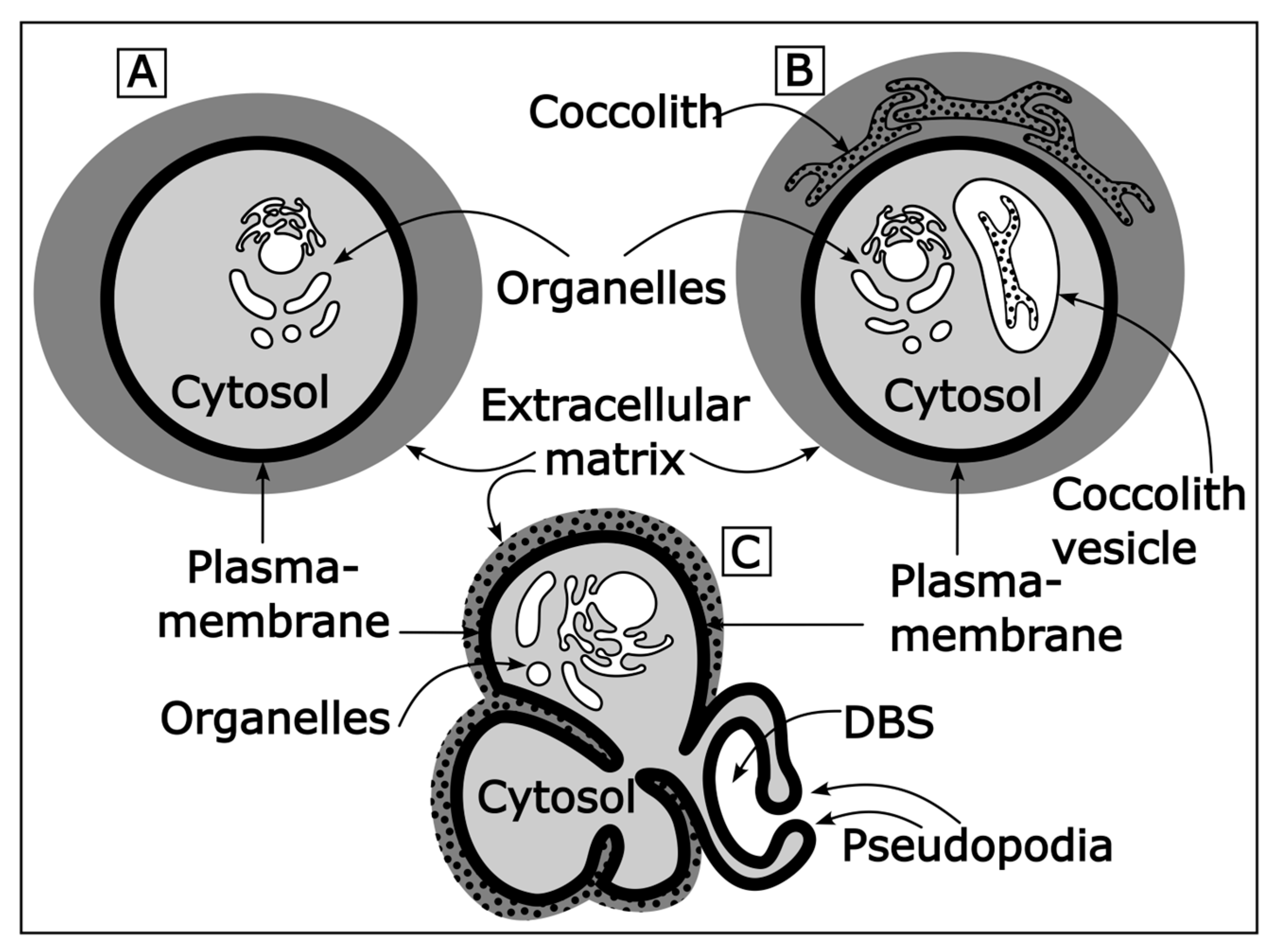
Every cell’s interior is separated from the environment by the plasma membrane. Everything inside the plasma membrane is called “intracellular”, everything outside “extracellular”. The “protoplast” refers to all intracellular material plus the plasma membrane. The protoplast is often equated with the cell. However, in technical terms the “cell” includes both the protoplast and the so-called extracellular matrix. It is easiest to understand this by example. Some unicellular organisms such as coccolithophores and most foraminifera surround themselves with a calcitic shell. This shell (for foraminifera called test and coccosphere in the case of coccolithophores) is part of the cell. Precisely, it is the extracellular part of the cell, the extracellular matrix (see also Figure 1A).
The protoplast of eukaryotic cells (cells with a nucleus) features a basic aqueous solution which performs many biochemical and structural tasks, the so-called cytosol (or cytoplasm). In addition to this central compartment there are numerous other compartments located inside the cytosol, each of which are surrounded by a membrane (collectively these membranes are termed endomembranes). Different membranes are similar in that they consist of a lipid bilayer with proteins. Both the particular lipids and the proteins vary and membranes of different compartments are characterized by their specific lipid/protein composition. The membranes function as both a barrier and a gateway for substances that travel into/out of the cell and between compartments within the cell. Membranes allow selective transport, which is central to any form of life. Different cellular tasks require different sub-cellular chemical environments created by selective transport. The precipitation of a biomineral is a highly specialized task requiring its own compartment to guarantee chemical and morphogenetic control. In shell-building calcifiers, this compartment can be extracellular or intracellular. In both cases, the necessary building blocks, i.e., calcium and carbonate ions, have to be transported. This transport can take three routes: (1) Trans-membrane transport mediated by either ion channels or transporters. (2) Vesicle transport. (3) Direct seawater transport; either by diffusion or pressure-driven mass flow. Note that direct seawater transport is only possible in extracellular calcifiers because in intracellular calcifiers at least one membrane has to be crossed.
2.2. Fundamentals of Element Incorporation during Mineral Formation
Numerous studies (mostly using precipitation experiments) have been performed to quantify the incorporation of various elements into calcium carbonates. In general, this incorporation is expressed by the so-called distribution (or partitioning) coefficient KD, which is defined as the amount of a given element in solution divided by its concentration within the mineral [3].
In the following paragraphs we will concentrate on processes related to element and isotope fractionation during mineral formation used to describe the observed proxy signature of biominerals. In addition, we will discuss how a biomineral differs from an inorganically formed mineral. If we apply this definition very strictly, a crystal originating from a precipitation experiment would not be a mineral, e.g., calcite but a crystal having the crystal structure of calcite. However, for practical reasons often the term mineral and its mineral name are used for artificially precipitated crystals too, as we will do in this discussion. For the sake of completeness, it should be mentioned that the process of crystal formation has a nucleation and a growth part, which both affect element incorporation (the interested reader is referred to, e.g., [4,5]. To not get lost in too many details, we do not separate these parts in this discussion and use the terms crystal (=mineral) and growth (=precipitation).
The main cation of calcium carbonate (CaCO3) is calcium (Ca). However, small amounts of other elements can be incorporated. Depending on their concentration, they are divided into minor and trace elements. The exact range of concentration that relates to these two terms is not clearly defined across disciplines and in the field of biomineralization not used uniformly. The described processes will focus on divalent cations such as magnesium (Mg), strontium (Sr), and barium (Ba) which substitute for Ca; nevertheless, many of these processes will also apply to elements not sitting in a Ca position, e.g., boron (B) and sodium (Na). For the sake of simplification, we will use the term foreign elements for all minor and trace elements.
Calcium carbonate occurs as different polymorphs (crystals having the same chemical composition but different crystal structures and solubility products), of which at ambient conditions calcite is the most stable. The difference in crystal structure between polymorphs directly affects the incorporation of foreign elements (an example for biominerals is found in, e.g., [6]). For example, under the same physico-chemical conditions higher amounts of Sr are incorporated into aragonite than into calcite (the opposite trend holds for Mg). This polymorph specific incorporation is related to the size difference of the foreign ions.
This polymorph specific incorporation is altered by three different factors: concentration, growth rate, and temperature. In general, an increase in the concentration of the foreign ions present in the parent solution (from which the mineral is precipitated) increases the amount available for incorporation. This incorporation is strongly affected by the growth rate of the mineral [7]. Depending on the size of the ion involved, the amount incorporated with increasing growth rate is increasing or decreasing compared to the equilibrium value (the value not altered by the growth rate, experimentally determined at extremely low growth rates). However, it is important to realize that the amount of a foreign element incorporated into a mineral, e.g., Sr into calcite can only be altered by the growth rate within a limited range [7].
As the velocity of chemical reactions is related to temperature, the growth rate of minerals will be affected by temperature, too. At high temperatures and pressures, temperature will directly influence the incorporation of foreign elements into a crystal lattice, as the interatomic distances within a crystal lattice are altered. However, for the temperature and pressure range in which biominerals are generally formed, a temperature effect on the incorporation of foreign elements, other than its impact on the growth rate, can be neglected.
From the six existing polymorphs of CaCO3 (amorphous calcium carbonate, calcium carbonate monohydrate, ikaite, vaterite, aragonite, and calcite), aragonite and calcite are the two polymorphs that form most of the biominerals made of calcium carbonate. If aragonite or calcite are precipitated from a solution saturated for amorphous calcium carbonate (ACC) too, it is often observed that aragonite or calcite do not precipitate directly from the solution but that ACC is the first polymorph precipitated followed by rapid transformation into aragonite or calcite. In this case, ACC is termed a “precursor” or “transient” phase. The process in which the final polymorph is formed via transient phases is known as “Ostwald Rule of Stages”. Due to the polymorph specific incorporation of foreign elements, a transient phase could influence the elemental composition of the polymorph this is finally present. In other words, the concentration of foreign ions in a calcite (or aragonite) could differ even if they are precipitated from the same solution depending on the precipitation pathway: direct precipitation or formation via a transient phase. However, since ACC often occurs as very short-lived transient phase measurements, especially in biogenic materials are challenging (e.g., [8]).
A crystal that looks homogeneous on the macroscopic level can be made up of several small crystals (crystallites or grains) of which each shows a different orientation (such a material is called polycrystalline); alternatively, it can represent one “single crystal” (uniform throughout the entire crystal). Already mentioned in the beginning, biominerals are complex composite materials containing organic and inorganic components. Strictly speaking, if a crystal is interrupted by organic material it should be polycrystalline. However, it is often observed that large biominerals show a uniform orientation of the crystal, even if layers of organic material are present. In this case, often the term mesocrystal is used [9]. Crystallographers have shown that often unit cell parameters of biominerals (defining the smallest building block of a crystal structure) can be measurably different when compared to their purely inorganic counterparts, e.g., [10].
It should be mentioned that most studies on element incorporation into CaCO3 put their focus on a single element. Studies using precipitations experiments in which the concentration of two or even more elements are changed simultaneously are very rare. However, the ones that have been performed show that the concentration of a certain element is affected by concentration changes of other elements. For example, the Mg and Sr concentrations in calcite are linked to the concentration of both elements in solution [11].
3. Discussion
Early biomineralization models analyzed if the measured KD values can be explained by purely inorganic processes. As the seawater concentration for most elements is in general constant, the only process that could alter KD significantly is the growth rate of the biomineral. However, it turned out that measured KD values could in most cases not be explained to be a result of the growth rate [12]. It has become commonly accepted in the last two decades that proxy signals are the result of the inorganic and cell biological processes, as presented in the previous chapter. The relationship between the two sets of processes is easiest visualized when picturing cell biological processes as putting obstacles in the way of “simple” precipitation from seawater. The source of raw material (calcium and carbonate ions) is seawater, the end product is the biomineral, and in between is the cell (or cells) transporting the raw material to the site of calcification. Usually, the element/isotope signature of the biomineral is normalized to the one of seawater. It is exactly this simple and straight forward normalization that causes so much “trouble” in understanding proxy signals since it would only work if the biomineral was precipitated directly from seawater. However, this simple prediction of the values measured is not possible and a large number of studies have been performed to figure out why. We will now start our discussion chronologically with one of the first attempts to explain foreign element fractionation including cell biological processes.
3.1. Seawater Transport versus Transmembrane Transport
A quarter century ago, it was proposed that foraminiferal minor element fractionation could be understood in terms of calcite precipitation from a closed reservoir containing initially seawater [13]. In the course of mineral growth, the initial composition of the calcifying fluid was thought to be altered by Rayleigh fractionation due to the finite reservoir, which serves the purpose of chamber formation but is too small to leave the chemical composition unchanged if ions are taken out in different proportions. This idea is in many ways a good starting point. Firstly, it assumes the simplest way of transporting the raw material, i.e., just taking seawater. Secondly, precipitation from seawater can be initiated by changing pH, keeping foreign element to calcium ratios constant. Thirdly, Rayleigh fractionation is a calculable process with only one unknown, i.e., the volume of the reservoir (which was thought to be inferable from microscopic evidence of the day). Please note that similar ideas of biomineral formation from pH manipulated seawater were put forth for molluscs [14,15] and corals [2].
In general, and in foraminifera in particular, more recent advances in the cell biology of calcification have led to the notion that things are not that simple. We will use foraminifera as a case in point here. Some twenty years ago, scientists in the group of Jonathan Erez began to use fluorescent dyes to track vesicle transport in benthic foraminifera. A series of papers published mainly in the following decade have detailed a new view of foraminiferal calcification. According to this concept, seawater is still the starting point, as it was originally conceived of in the seminal study discussed above. However, instead of being merely pH manipulated, it was now proposed that more modifications are performed [16,17]. At this point it is helpful to describe the general cell biology of foraminiferal chamber formation (see also Figure 1).
Foraminifera build a new chamber in an extracellular compartment termed delimited biomineralization space (DBS). Since the DBS is extracellular it is, in principle, open to seawater. The term “delimited” was chosen because the general consensus is that the DBS is partly closed to seawater and represents a spatially defined volume in which chamber growth can be controlled [18,19]. The DBS is formed by the pseudopodial network of foraminifera [20]. Pseudopodia are dynamic cell protrusions used for various purposes such as feeding, locomotion, and chamber formation. In contrast to the CV in coccolithophores, it is unclear whether the DBS volume is sufficient to contain all raw material required for chamber formation. It is generally assumed that this is not the case and that vesicles provide a pool of Ca and inorganic carbon. Whether this pool is sufficient for chamber formation is also debated [21,22,23].
The physiological steps of seawater modification in foraminifera can be summarized as follows. The first step is endocytosis (invagination of the plasma membrane and formation of a vesicle) of seawater, followed by vesicle transport through the cell towards the DBS. En route, the fluid inside these vesicles is modified by means of transmembrane ion transport, and the modified seawater is exocytosed (inverse endocytosis) into the DBS and used for chamber formation. This concept is thought to, inter alia, solve the “Mg problem”. Different foraminifera species have different Mg/Ca which are mostly too low to be the result of calcite precipitation from seawater. A transmembrane export of Mg from the vesicle into the cytosol could account for these low Mg/Ca values.
An alternative view of foraminiferal ion transport is that vesicle transport and direct seawater transport play minor roles and the bulk of Ca and inorganic carbon enters the DBS via transmembrane transport through the pseudopodial network [22]. On this account there is no need for Mg removal because Mg is kept out in the first step, which is thought to be a plasma membrane Ca-channel.
The important point in the context of this discussion is that, although direct seawater transport can never be fully excluded in extracellular calcifiers, the development of biomineralization concepts for all marine calcifiers in the last two decades shows that transmembrane transport plays key roles in calcification substrate supply and foreign element fractionation. Transmembrane transport also plays a key role in “waste removal” as opposed to substrate supply.
Foraminifera:
Since foraminifera, and- also coccolithophores, are faced with the “proton load problem” due to calcium carbonate precipitation, there is a need to export protons from the DBS into the surrounding seawater. Acidification adjacent to a newly forming chamber has been shown experimentally, as well as the concomitant high pH inside the cell [19,21,24]. It is unknown how protons are exported but a transmembrane transport across the pseudopodial network has been argued for [19].
Coccolithophores:
Since calcite precipitation would rapidly acidify the CV fluid, protons are transported across CV and plasma membrane. The latter step is ion channel mediated and does not function properly at seawater pH 7.6 (normal pH is 8.2), resulting in cytosol acidification and coccolith malformations [25]. This illustrates the importance of transmembrane transport for virtually all aspects of coccolith formation, from Ca supply and proton removal to morphogenesis.
The importance of transmembrane transport in marine calcifiers and the difficulty of estimating the percentages of transmembrane transport and direct seawater transport in extracellular calcifiers makes it particularly interesting to consider intracellular calcifiers. Coccolithophores are the prime example here because they are thought to deliver the Ca ions for calcification by means of transmembrane transport. A qualification should be mentioned here, i.e., a vesicle Ca transport inside the cell is being considered [26,27,28]. The distinguishing feature is the transmembrane transport of Ca ions across the plasmamembrane; the consequence being that coccolithophores do not calcify from modified seawater, neither do they mix seawater into their calcifying fluid.
Coccolithophores form coccoliths inside a Golgi derived vesicle, the coccolith vesicle (CV, [29]). The CV fluid is separated from seawater by two membranes (the CV membrane and the plasma membrane). Ca and dissolved inorganic carbon (mainly bicarbonate) are thought to be transported across the plasma membrane and possibly across the CV membrane too [30,31]. The small volume of the CV makes it necessary to transport ions into the CV while they are removed by precipitation [30]. In other words, the CV fluid contains, at any given time, only a minute fraction of the ions necessary for coccolith formation. This insufficient reservoir rules out Rayleigh fractionation as an explanation for minor element fractionation during coccolith formation.
By contrast, it was recently proposed that Rayleigh fractionation is sufficient to explain minor element incorporation in some foraminifera [23]. This shows that, despite considerable progress in the development of mechanistic biomineralization concepts, the seminal idea of Elderfield et al. [13] is not obsolete. More broadly, it illustrates general features of the relationship between mechanistic biomineralization concepts and minor element fractionation patterns. This relationship is truly bidirectional, i.e., a biomineralization concept does not determine a certain fractionation pattern, neither is it the other way round. The hope is that the two will go well together. If so, the biomineralization concept helps to understand the fractionation pattern and the fractionation pattern supports the biomineralization concept. Here, we focus on the former aspect, but since the latter aspect is important, it is helpful to give an example. In Patella caerulea, the fractionation patterns of several elements taken together help to decide between two rival models of shell formation [32].
In the particular case of proxy application, we may ask, as Urey et al. (1951) did, “is there a vital effect?”. Today, the answer arguably is yes, always. That does not mean that there is always a difference between the foreign element fractionation behavior of a calcifying organism and inorganic precipitation because vital effects (cell biological fractionation steps) can be invisible [33,34,35,36].
If vital effects are ubiquitous, it might seem that mechanistic biomineralization concepts are essential to successful proxy application. We will say that these concepts are certainly essential to a deeper understanding of proxy signals and they have the potential to improve proxy application. However, have they conducted this yet? Furthermore, are they essential?
3.2. Adding Complexity
Today, conceptual biomineralization models designed to explain foreign element fractionation are based on both fractionation during crystal precipitation and fractionation during cellular ion transport [37]. As outlined above, both “crystal fractionation” and “cell fractionation” contain several fractionation steps. Crystal fractionation includes thermodynamic, kinetic, and polymorph effects [7]. Cell fractionation includes ion channels and active transporters, polysaccharides, and proteins [27]. Some of these fractionation steps are quantified, especially, in the inorganic system. Cellular fractionation steps are, by contrast, mostly not quantified. This lack of knowledge is the reason why there is no conceptual biomineralization model that can predict foreign element fractionation. Current models are being “wise after the event”, i.e., they use measured fractionation patterns to fit fractionation factors that are not constrained by independent measurements. Hence, the task is to conduct these independent measurements and provide the required constraints for the models. The following example might illustrate why this is a monumental task. Note that different foreign elements might require different models because of their differential transport pathway and biochemistry [38].
The fractionation of Sr into coccoliths and foraminiferal shells is considered a relatively easy case because it is highly likely that Sr simply follows Ca on its route through the cell [30,39]. Since Ca is a central ion for virtually all organisms, its cellular transport has been extensively studied and a lot is known about its intracellular distribution [28,40,41]. However, measurements of Sr/Ca fractionation factors are rare and, importantly, not available for the organisms considered as proxy archives here. The first step of cellular Sr fractionation in coccolithophores, and potentially foraminifera too, is a plasma membrane Ca channel [22,27]. The coccolithophore Coccolithus braarudii has three different types of Ca channels. None of these is characterized in terms of its Sr fractionation [27]. If Ca channels of other organisms are taken as substitutes one faces the difficulty that some of these channels fractionate against Sr, some for Sr, and some do not fractionate at all [42,43,44]. Therefore, in order to characterize the first step of Sr fractionation in one single coccolithophore, a dedicated project determining Sr and Ca permeability of three Ca channels is required. Furthermore, even if successful, this project is merely the first step. Following steps include fractionation during transmembrane transport across endomembranes, intracellular polysaccharides, proteins, etc. [27,45]. Some of these steps have already been included in models, but all quantification is based on measurements conducted on animals, seed plants, brown macro-algae, etc., potentially providing misleading values (e.g., [46]). To constrain models in such a way that they can predict fractionation patterns is possible, given a concerted effort including mineralogists, cell biologists, chemists, and physicists. Such an effort has been made, but the progress over the last three decades shows that it will take some time before models can be used to predict proxy signals.
3.3. The Role of Analytical Advances
Our modern picture of biomineralization is strongly linked to the huge advances in analytical methods during the last three decades. These analytical methods made it possible to investigate the distribution of elements and molecules in biominerals down to the nm scale and to observe transport processes within calcifying organisms in vivo.
These micro-focusing methods do not only reveal previously hidden features they also represent new challenges as they add additional complexity that has to be considered if trying to understand the biomineralization process of marine calcifying organisms. For example, dyes used in confocal laser scanning microscopy allow the visualization of processes that occur when an organism is calcifying, e.g., the transport of vesicles containing seawater. These observations alone neither reveal the purpose of this transport nor prove that it is directly linked to the biomineralization process and the proxy signature measured.
However, these observations are valuable because they stimulate further research that will, given the sometimes necessary technical developments, test the initial hypotheses. Intracellular Ca transport in Emiliania huxleyi is an example. Although intracellular Ca transport via vesicles had been suggested for coccolithophores in general, there was no evidence for it in E. huxleyi and the conservative assumption was that it does not exist [27,30]. Analytical advances allowed the detection of Ca-rich vesicles and pulse chase experiments showed that these vesicles are involved in Ca transport for coccolith formation [39,47].
During the last decades it has been shown that foreign elements (and organic molecules) in biominerals are not homogeneously distributed. Many biominerals show a banding pattern that is somehow related to their growth phases such as the banding pattern in the biominerals of foraminifera and corals. Again, this is important information but adds complexity to the biomineralization model, since a model that aims to explain the elemental/isotopic signatures measured in the biomineral would not only have to account for the bulk composition measured but explain the observed distribution pattern too, e.g., [48].
4. Conclusions
The key question raised in the beginning was how a better understanding of the biomineralization process has improved the way we use proxy archives formed by marine calcifiers today.
Recent biomineralization models applicable to marine calcifying organisms are not comparable to the early models discussed a few decades ago. Many physiological processes hypothesized by early models are now detected and we have quite a good understanding of the general biomineralization process (e.g., [39]). As the early models had been developed mostly in the context of proxy research, recent models concentrate on the biomineralization process itself. Some of these models even try to find common features of calcifying organisms that can be linked on an evolutionary level [49]. However, as explained in Section 3.3, the quantification of proxy signatures would require quantifying the element/isotope fractionation of all physiological processes involved. Now that the complexity of involved processes is not only hypothesized but demonstrated it has become clear how difficult this will be. To the best of our knowledge there is no biomineralization model for marine calcifiers that changed or improved the way the proxy signature measured is actually used. We better understand which processes are involved but proxy signatures are still used in the same “classical way”, i.e., a calibration (using cultured or field samples) is performed and this calibration is then applied to reconstruct environmental parameters, e.g., temperature or pH.
However, in the process of developing biomineralization models, observations were made that did indeed improve how we use proxy archives or read proxy signals. Two classes of observations stand out: Firstly, microanalytical techniques revealed inhomogeneities of foreign elements in biominerals and suggested ways in which the proxy archive can be used to the greatest advantage (e.g., [50,51]. Secondly, experiments revealed correlations between proxies and target variables that were misinterpreted in core top calibrations and down-core applications [12].
Although indirect, these two examples show that the development of biomineralization models is useful for proxy application, even if the models are not capable of predicting proxy signals solely based on independent constraints.
Author Contributions
G.N. and G.L. contributed equally to this work. Conceptualization, G.N. and G.L.; methodology, G.N. and G.L.; software, G.N. and G.L.; validation, G.N. and G.L.; formal analysis, G.N. and G.L.; investigation, G.N. and G.L.; resources, G.N. and G.L.; data curation, G.N. and G.L.; writing—original draft preparation, G.N. and G.L.; writing—review and editing, G.N. and G.L.; visualization, G.N. and G.L.; supervision, G.N. and G.L.; project administration, G.N. and G.L.; funding acquisition, G.L. All authors have read and agreed to the published version of the manuscript.
Funding
GL acknowledges funding from the Spanish Ministry of Education through a Maria Zambrano grant.
Data Availability Statement
This paper does not contain original data.
Acknowledgments
Special thanks go to Karina Kaczmarek who produced the figures.
Conflicts of Interest
The authors declare no conflict of interest.
Glossary
| Amoeboid | a unicellular shape-shifting organism possessing pseudopodia |
| Cell | protoplast plus extracellular matrix |
| Coccolith vesicle (CV) | a specialised Golgi derived organelle producing the coccoliths |
| Delimited biomineralization space (DBS) | the extracellular compartment of foraminifera containing the so-called calcifying fluid in which a new chamber is produced |
| Endomembrane | blanket term for all membranes of intracellular organelles |
| Extracellular | everything outside the plasmamembrane |
| Extracellular matrix | all cellular components outside the plasmamembrane |
| Intracellular | everything inside the plasmamembrane |
| Membrane | lipid bilayer with proteins |
| Organelle | an intracellular compartment surrounded by an endomembrane |
| Plasmamembrane | membrane forming the boundary of the protoplast |
| Protoplast | all cellular components inside the plasmamembrane plus the plasmamembrane |
| Pseudopodia | highly dynamic protrusions of the plasmamembrane used for e.g., moving, feeding, chamber formation (foraminifera). |
References
- Urey, H.C.; Lowenstam, H.A.; Epstein, S.; McKinney, C.R. Measurement of Paleotemperatures and Temperatures of the Upper Cretaceous of England, Denmark, and the Southeastern United States. Geol. Soc. Am. Bull. 1951, 62, 399–416. [Google Scholar] [CrossRef]
- Cuif, J.-P.; Dauphin, Y.; Sorauf, J.E. Biominerals and Fossils through Time; Cambridge University Press: Cambridge, UK, 2011. [Google Scholar]
- McIntire, W.L. Trace element partition coefficients—A review of theory and applications to geology. Geochim. Cosmochim. Acta 1963, 27, 1209–1264. [Google Scholar] [CrossRef]
- Nielsen, A.E. Kinetics of Precipitation; Pergamon Press: Oxford, UK, 1964. [Google Scholar]
- Kashchiev, D. Nucleation; Butterworth-Heinemann: Oxford, UK, 2000; p. 529. [Google Scholar]
- Nehrke, G.; Poigner, H.; Wilhelms-Dick, D.; Brey, T.; Abele, D. Coexistence of three calcium carbonate polymorphs in the shell of the Antarctic clam Laternula elliptica. Geochem. Geophys. Geosyst. 2012, 13, Q05014. [Google Scholar] [CrossRef]
- Rimstidt, J.D.; Bolag, A.; Webb, J. Distribution of trace elements between carbonate minerals and aqueous solutions. Geochim. Cosmochim. Acta 1998, 62, 1851–1863. [Google Scholar] [CrossRef]
- Ramesh, K.; Melzner, F.; Griffith, A.W.; Gobler, C.J.; Rouger, C.; Tasdemir, D.; Nehrke, G. In vivo characterization of bivalve larval shells: A confocal Raman microscopy study. J. R. Soc. Interface 2018, 15, 20170723. [Google Scholar] [CrossRef]
- Bergström, L.; Sturm, E.V.; Salazar-Alvarez, G.; Cölfen, H. Mesocrystals in Biominerals and Colloidal Arrays. Acc. Chem. Res. 2015, 48, 1391–1402. [Google Scholar] [CrossRef] [PubMed]
- Farfan, G.A.; Apprill, A.; Cohen, A.; DeCarlo, T.M.; Post, J.E.; Waller, R.G.; Hansel, C.M. Crystallographic and chemical signatures in coral skeletal aragonite. Coral Reefs 2022, 41, 19–34. [Google Scholar] [CrossRef]
- Mucci, A.; Morse, J.W. The incorporation of Mg2+ and Sr2+ into calcite overgrowths: Influences of growth rate and solution composition. Geochim. Cosmochim. Acta 1983, 47, 217–233. [Google Scholar] [CrossRef]
- Stoll, H.M.; Ziveri, P. Coccolithophorid-Based Geochemical Paleoproxies. In Coccolithophores: From Molecular Processes to Global Impact; Thierstein, H.R., Young, J.R., Eds.; Springer Berlin Heidelberg: Berlin, Heidelberg, Germany, 2004; pp. 529–562. [Google Scholar]
- Elderfield, H.; Bertram, C.J.; Erez, J. A biomineralization model for the incorporation of trace elements into foraminiferal calcium carbonate. Earth Planet. Sci. Lett. 1996, 142, 409–423. [Google Scholar] [CrossRef]
- Crenshaw, M.A. The Inorganic Composition of Molluscan Extrapallial Fluid. Biol. Bull. 1972, 143, 506–512. [Google Scholar] [CrossRef]
- Misogianes, M.J.; Chasteen, N.D. A chemical and spectral characterization of the extrapallial fluid of Mytilus edulis. Anal. Biochem. 1979, 100, 324–334. [Google Scholar] [CrossRef] [PubMed]
- Erez, J. The Source of Ions for Biomineralization in Foraminifera and Their Implications for Paleoceanographic Proxies. Rev. Mineral. Geochem. 2003, 54, 115–149. [Google Scholar] [CrossRef]
- Bentov, S.; Brownlee, C.; Erez, J. The role of seawater endocytosis in the biomineralization process in calcareous foraminifera. Proc. Natl. Acad. Sci. USA 2009, 106, 21500–21504. [Google Scholar] [CrossRef] [PubMed]
- Angell, R.W. The Process of Chamber Formation in the Foraminifer Rosalina floridana (Cushman). J. Protozool. 1967, 14, 566–574. [Google Scholar] [CrossRef]
- Glas, M.S.; Langer, G.; Keul, N. Calcification acidifies the microenvironment of a benthic foraminifer (Ammonia sp.). J. Exp. Mar. Biol. Ecol. 2012, 424–425, 53–58. [Google Scholar] [CrossRef]
- Tyszka, J.; Bickmeyer, U.; Raitzsch, M.; Bijma, J.; Kaczmarek, K.; Mewes, A.; Topa, P.; Janse, M. Form and function of F-actin during biomineralization revealed from live experiments on foraminifera. Proc. Natl. Acad. Sci. USA 2019, 116, 4111–4116. [Google Scholar] [CrossRef]
- de Nooijer, L.J.; Langer, G.; Nehrke, G.; Bijma, J. Physiological controls on seawater uptake and calcification in the benthic foraminifer Ammonia tepida. Biogeosciences 2009, 6, 2669–2675. [Google Scholar] [CrossRef]
- Nehrke, G.; Keul, N.; Langer, G.; de Nooijer, L.J.; Bijma, J.; Meibom, A. A new model for biomineralization and trace-element signatures of Foraminifera tests. Biogeosciences 2013, 10, 6759–6767. [Google Scholar] [CrossRef]
- Evans, D.; Müller, W.; Erez, J. Assessing foraminifera biomineralisation models through trace element data of cultures under variable seawater chemistry. Geochim. Cosmochim. Acta 2018, 236, 198–217. [Google Scholar] [CrossRef]
- Toyofuku, T.; Matsuo, M.Y.; de Nooijer, L.J.; Nagai, Y.; Kawada, S.; Fujita, K.; Reichart, G.-J.; Nomaki, H.; Tsuchiya, M.; Sakaguchi, H.; et al. Proton pumping accompanies calcification in foraminifera. Nat. Commun. 2017, 8, 14145. [Google Scholar] [CrossRef]
- Kottmeier, D.M.; Chrachri, A.; Langer, G.; Helliwell, K.E.; Wheeler, G.L.; Brownlee, C. Reduced H+ channel activity disrupts pH homeostasis and calcification in coccolithophores at low ocean pH. Proc. Natl. Acad. Sci. USA 2022, 119, e2118009119. [Google Scholar] [CrossRef] [PubMed]
- Marsh, M.E. Regulation of CaCO3 formation in coccolithophores. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2003, 136, 743–754. [Google Scholar] [CrossRef]
- Brownlee, C.; Taylor, A. Calcification in Coccolithophores: A Cellular Perspective. In Coccolithophores: From Molecular Processes to Global Impact; Thierstein, H.R., Young, J.R., Eds.; Springer: Berlin/Heidelberg, Germany, 2004; pp. 31–49. [Google Scholar]
- Kadan, Y.; Aram, L.; Shimoni, E.; Levin-Zaidman, S.; Rosenwasser, S.; Gal, A. In situ electron microscopy characterization of intracellular ion pools in mineral forming microalgae. J. Struct. Biol. 2020, 210, 107465. [Google Scholar] [CrossRef] [PubMed]
- Wilbur, K.M.; Watabe, N. Experimental studies on calcification in molluscs and the alga coccolithus huxleyi. Ann. N. Y. Acad. Sci. 1963, 109, 82–112. [Google Scholar] [CrossRef]
- Langer, G.; Gussone, N.; Nehrke, G.; Riebesell, U.; Eisenhauer, A.; Kuhnert, H.; Rost, B.; Trimborn, S.; Thoms, S. Coccolith strontium to calcium ratios in Emiliania huxleyi: The dependence on seawater strontium and calcium concentrations. Limnol. Oceanogr. 2006, 51, 310–320. [Google Scholar] [CrossRef]
- Brownlee, C.; Langer, G.; Wheeler, G.L. Coccolithophore calcification: Changing paradigms in changing oceans. Acta Biomater. 2021, 120, 4–11. [Google Scholar] [CrossRef]
- Langer, G.; Sadekov, A.; Nehrke, G.; Baggini, C.; Rodolfo-Metalpa, R.; Hall-Spencer, J.M.; Cuoco, E.; Bijma, J.; Elderfield, H. Relationship between mineralogy and minor element partitioning in limpets from an Ischia CO2 vent site provides new insights into their biomineralization pathway. Geochim. Cosmochim. Acta 2018, 236, 218–229. [Google Scholar] [CrossRef]
- Taubner, I.; Böhm, F.; Eisenhauer, A.; Garbe-Schönberg, D.; Erez, J. Uptake of alkaline earth metals in Alcyonarian spicules (Octocorallia). Geochim. Cosmochim. Acta 2012, 84, 239–255. [Google Scholar] [CrossRef]
- Keul, N.; Langer, G.; de Nooijer, J.L.; Nehrke, G.; Reichart, G.J.; Bijma, J. Incorporation of uranium in benthic foraminiferal calcite reflects seawater carbonate ion concentration. Geochem. Geophys. Geosyst. 2013, 14, 102–111. [Google Scholar] [CrossRef]
- Gussone, N.; Filipsson, H.L.; Kuhnert, H. Mg/Ca, Sr/Ca and Ca isotope ratios in benthonic foraminifers related to test structure, mineralogy and environmental controls. Geochim. Cosmochim. Acta 2016, 173, 142–159. [Google Scholar] [CrossRef]
- Langer, G.; Sadekov, A.; Greaves, M.; Nehrke, G.; Probert, I.; Misra, S.; Thoms, S. Li Partitioning Into Coccoliths of Emiliania huxleyi: Evaluating the General Role of “Vital Effects” in Explaining Element Partitioning in Biogenic Carbonates. Geochem. Geophys. Geosyst. 2020, 21, e2020GC009129. [Google Scholar] [CrossRef]
- Langer, G.; Sadekov, A.; Thoms, S.; Keul, N.; Nehrke, G.; Mewes, A.; Greaves, M.; Misra, S.; Reichart, G.-J.; de Nooijer, L.J.; et al. Sr partitioning in the benthic foraminifera Ammonia aomoriensis and Amphistegina lessonii. Chem. Geol. 2016, 440, 306–312. [Google Scholar] [CrossRef]
- Langer, G.; Sadekov, A.; Thoms, S.; Mewes, A.; Nehrke, G.; Greaves, M.; Misra, S.; Bijma, J.; Elderfield, H. Li partitioning in the benthic foraminifera Amphistegina lessonii. Geochem. Geophys. Geosyst. 2015, 16, 4275–4279. [Google Scholar] [CrossRef]
- Gal, A.; Sviben, S.; Wirth, R.; Schreiber, A.; Lassalle-Kaiser, B.; Faivre, D.; Scheffel, A. Trace-Element Incorporation into Intracellular Pools Uncovers Calcium-Pathways in a Coccolithophore. Adv. Sci. 2017, 4, 1700088. [Google Scholar] [CrossRef]
- Berridge, M.J.; Lipp, P.; Bootman, M.D. The versatility and universality of calcium signalling. Nat. Rev. Mol. Cell Biol. 2000, 1, 11–21. [Google Scholar] [CrossRef]
- Marchadier, E.; Oates, M.E.; Fang, H.; Donoghue, P.C.J.; Hetherington, A.M.; Gough, J. Evolution of the Calcium-Based Intracellular Signaling System. Genome Biol. Evol. 2016, 8, 2118–2132. [Google Scholar] [CrossRef]
- Allen, G.J.; Sanders, D. Two Voltage-Gated, Calcium Release Channels Coreside in the Vacuolar Membrane of Broad Bean Guard Cells. Plant Cell 1994, 6, 685–694. [Google Scholar] [CrossRef]
- Bourinet, E.; Zamponi, G.W.; Stea, A.; Soong, T.W.; Lewis, B.A.; Jones, L.P.; Yue, D.T.; Snutch, T.P. The α1ECalcium Channel Exhibits Permeation Properties Similar to Low-Voltage-Activated Calcium Channels. J. Neurosci. 1996, 16, 4983–4993. [Google Scholar] [CrossRef]
- Lin, Y.C.J.; Spencer, A.N. Calcium currents from jellyfish striated muscle cells: Preservation of phenotype, characterisation of currents and channel localisation. J. Exp. Biol. 2001, 204, 3717–3726. [Google Scholar] [CrossRef] [PubMed]
- Meyer, E.M.; Langer, G.; Brownlee, C.; Wheeler, G.L.; Taylor, A.R. Sr in coccoliths of Scyphosphaera apsteinii: Partitioning behavior and role in coccolith morphogenesis. Geochim. Cosmochim. Acta 2020, 285, 41–54. [Google Scholar] [CrossRef]
- Langer, G.; Nehrke, G.; Thoms, S.; Stoll, H. Barium partitioning in coccoliths of Emiliania huxleyi. Geochim. Cosmochim. Acta 2009, 73, 2899–2906. [Google Scholar] [CrossRef]
- Sviben, S.; Gal, A.; Hood, M.A.; Bertinetti, L.; Politi, Y.; Bennet, M.; Krishnamoorthy, P.; Schertel, A.; Wirth, R.; Sorrentino, A.; et al. A vacuole-like compartment concentrates a disordered calcium phase in a key coccolithophorid alga. Nat. Commun. 2016, 7, 11228. [Google Scholar] [CrossRef]
- Thompson, D.M. Environmental records from coral skeletons: A decade of novel insights and innovation. WIREs Clim. Chang. 2022, 13, e745. [Google Scholar] [CrossRef]
- Gilbert, P.U.P.A.; Bergmann, K.D.; Boekelheide, N.; Tambutté, S.; Mass, T.; Marin, F.; Adkins, J.F.; Erez, J.; Gilbert, B.; Knutson, V.; et al. Biomineralization: Integrating mechanism and evolutionary history. Sci. Adv. 2022, 8, eabl9653. [Google Scholar] [CrossRef] [PubMed]
- Rollion-Bard, C.; Erez, J. Intra-shell boron isotope ratios in the symbiont-bearing benthic foraminiferan Amphistegina lobifera: Implications for δ11B vital effects and paleo-pH reconstructions. Geochim. Cosmochim. Acta 2010, 74, 1530–1536. [Google Scholar] [CrossRef]
- Fietzke, J.; Wall, M. Distinct fine-scale variations in calcification control revealed by high-resolution 2D boron laser images in the cold-water coral Lophelia pertusa. Sci. Adv. 2022, 8, eabj4172. [Google Scholar] [CrossRef]
Figure 1.
(A) Sketch of a cell and its basic compartments. Note that the extracellular matrix is part of the cell. However, the term “intracellular” only refers to parts of the cell that are located inside the plasma membrane. (B) Sketch of a coccolithophore cell. The coccolith vesicle is a specialized intracellular compartment producing the coccoliths. The extracellular matrix of coccolithophores is called coccosphere. (C) Sketch of a foraminifera cell. The delimited biomineralization space (DBS) is a specialized extracellular compartment that produces the test chambers. The extracellular matrix of foraminifera is called test (or shell). It should be noted that extracellular and intracellular calcification both occur within the cell, but intracellular calcification takes place in a compartment inside the plasma membrane and extracellular calcification takes place in a compartment outside the plasma membrane.
Figure 1.
(A) Sketch of a cell and its basic compartments. Note that the extracellular matrix is part of the cell. However, the term “intracellular” only refers to parts of the cell that are located inside the plasma membrane. (B) Sketch of a coccolithophore cell. The coccolith vesicle is a specialized intracellular compartment producing the coccoliths. The extracellular matrix of coccolithophores is called coccosphere. (C) Sketch of a foraminifera cell. The delimited biomineralization space (DBS) is a specialized extracellular compartment that produces the test chambers. The extracellular matrix of foraminifera is called test (or shell). It should be noted that extracellular and intracellular calcification both occur within the cell, but intracellular calcification takes place in a compartment inside the plasma membrane and extracellular calcification takes place in a compartment outside the plasma membrane.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Nehrke, G.; Langer, G. Proxy Archives Based on Marine Calcifying Organisms and the Role of Process-Based Biomineralization Concepts. Minerals 2023, 13, 561. https://doi.org/10.3390/min13040561
AMA Style
Nehrke G, Langer G. Proxy Archives Based on Marine Calcifying Organisms and the Role of Process-Based Biomineralization Concepts. Minerals. 2023; 13(4):561. https://doi.org/10.3390/min13040561
Chicago/Turabian StyleNehrke, Gernot, and Gerald Langer. 2023. "Proxy Archives Based on Marine Calcifying Organisms and the Role of Process-Based Biomineralization Concepts" Minerals 13, no. 4: 561. https://doi.org/10.3390/min13040561
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.