


Permian Cyanobacterial Blooms Resulted in Enrichment of Organic Matter in the Lucaogou Formation in the Junggar Basin, NW China
Abstract
:1. Introduction
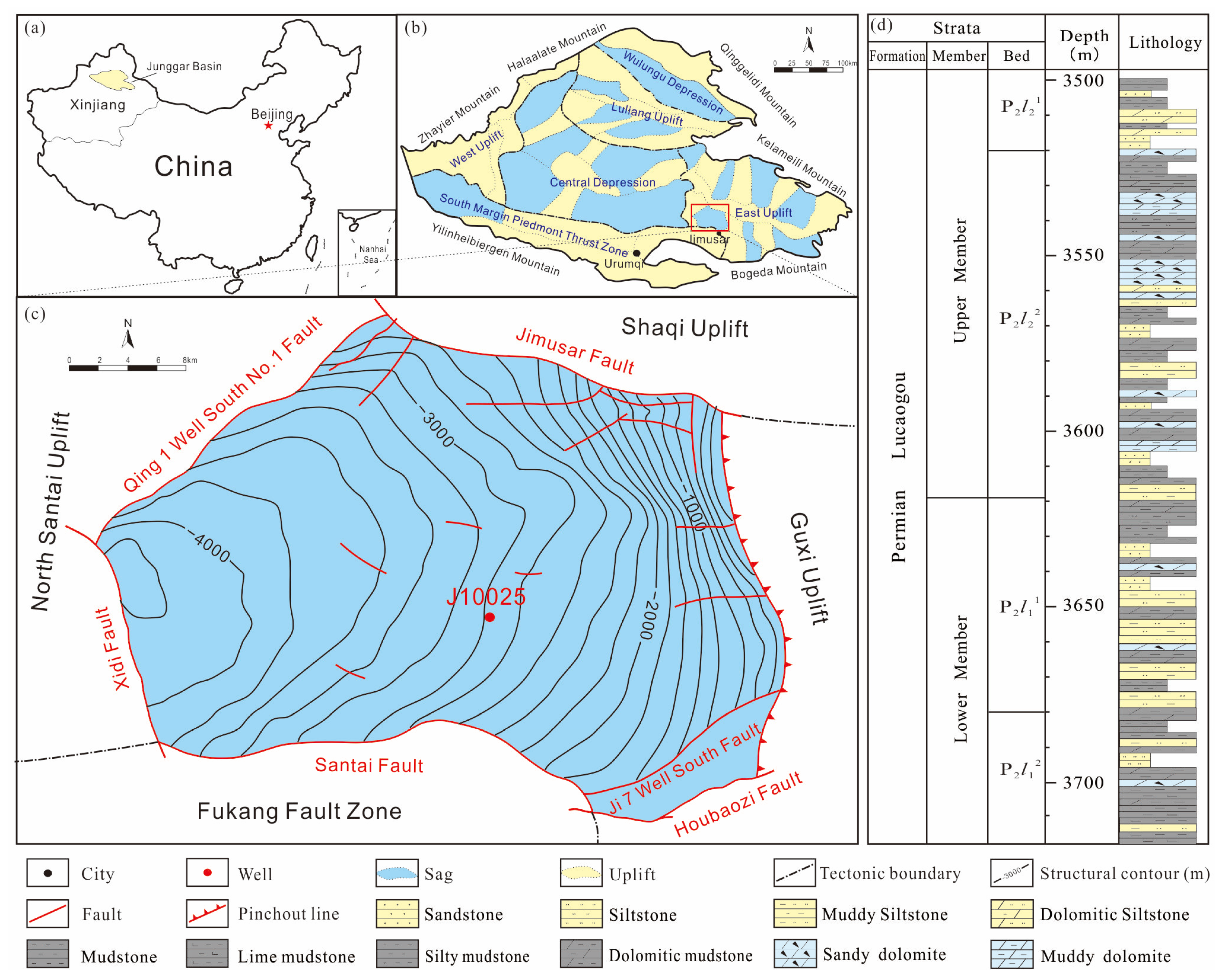
2. Geological Setting

3. Materials and Methods
3.1. Petrological and Paleontological Analysis
3.2. Biomarker Compound Analysis
3.3. TOC and Element Concentration Analysis
4. Results
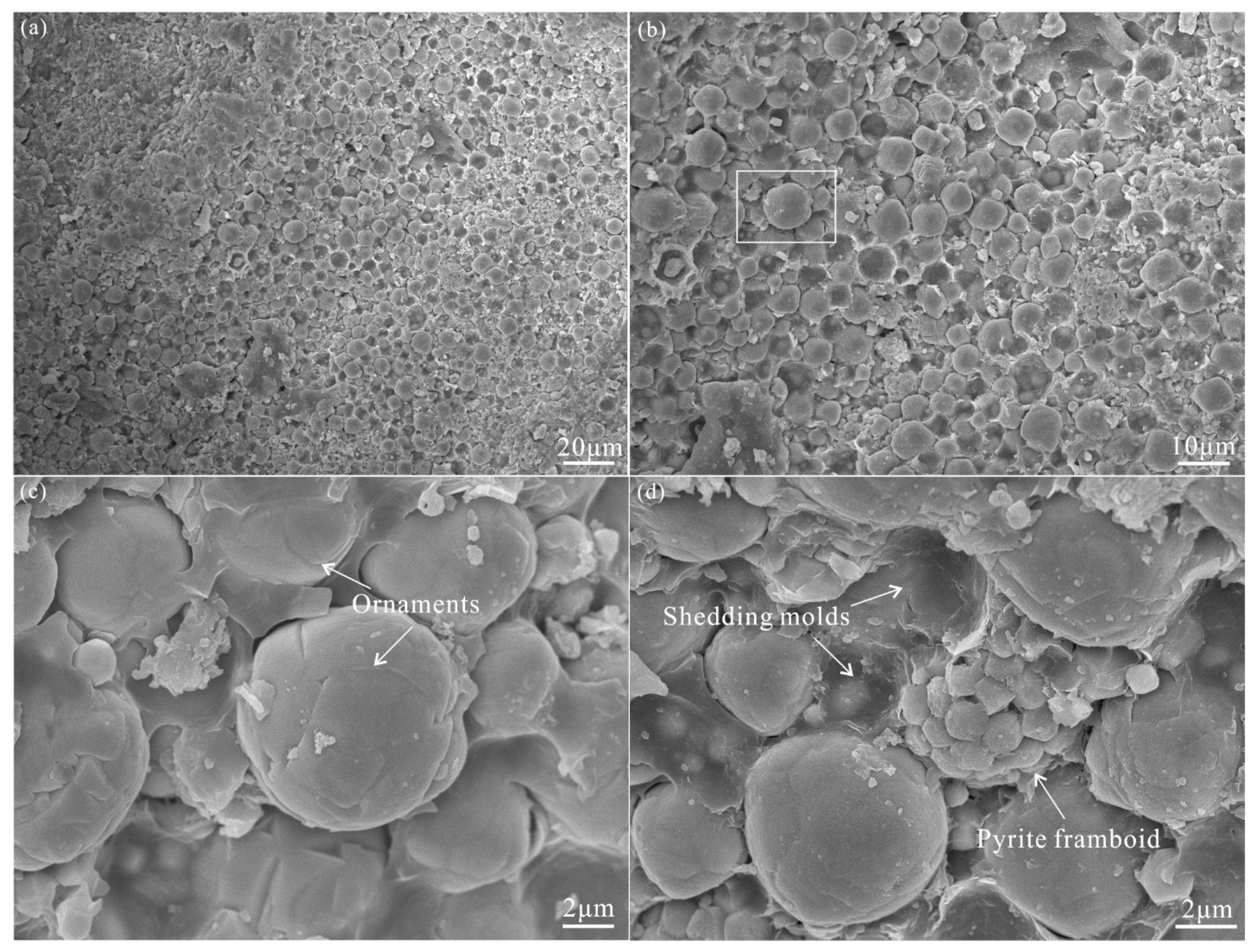
4.1. Microfossils in the Lucaogou Formation
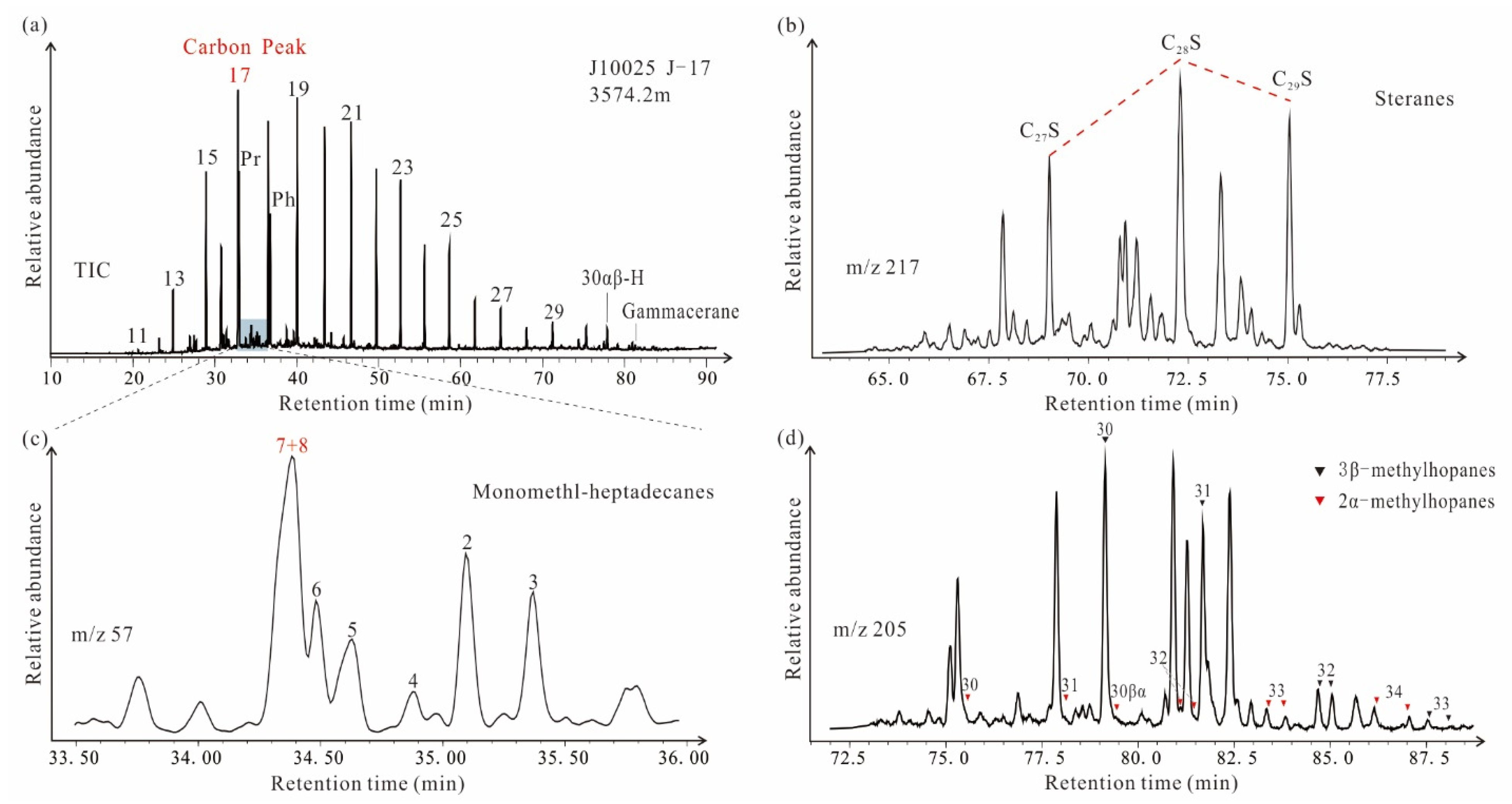
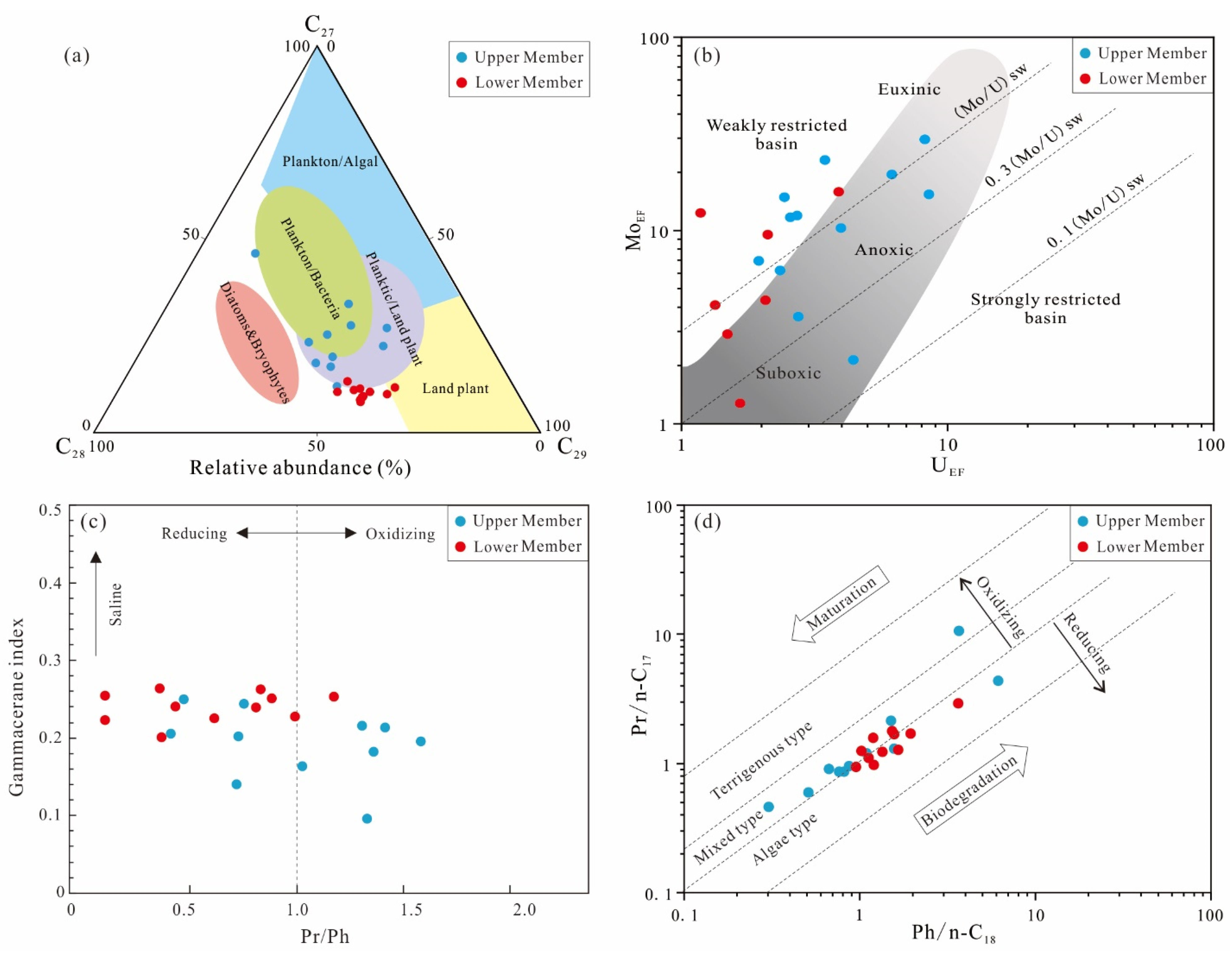
4.2. Biomarkers
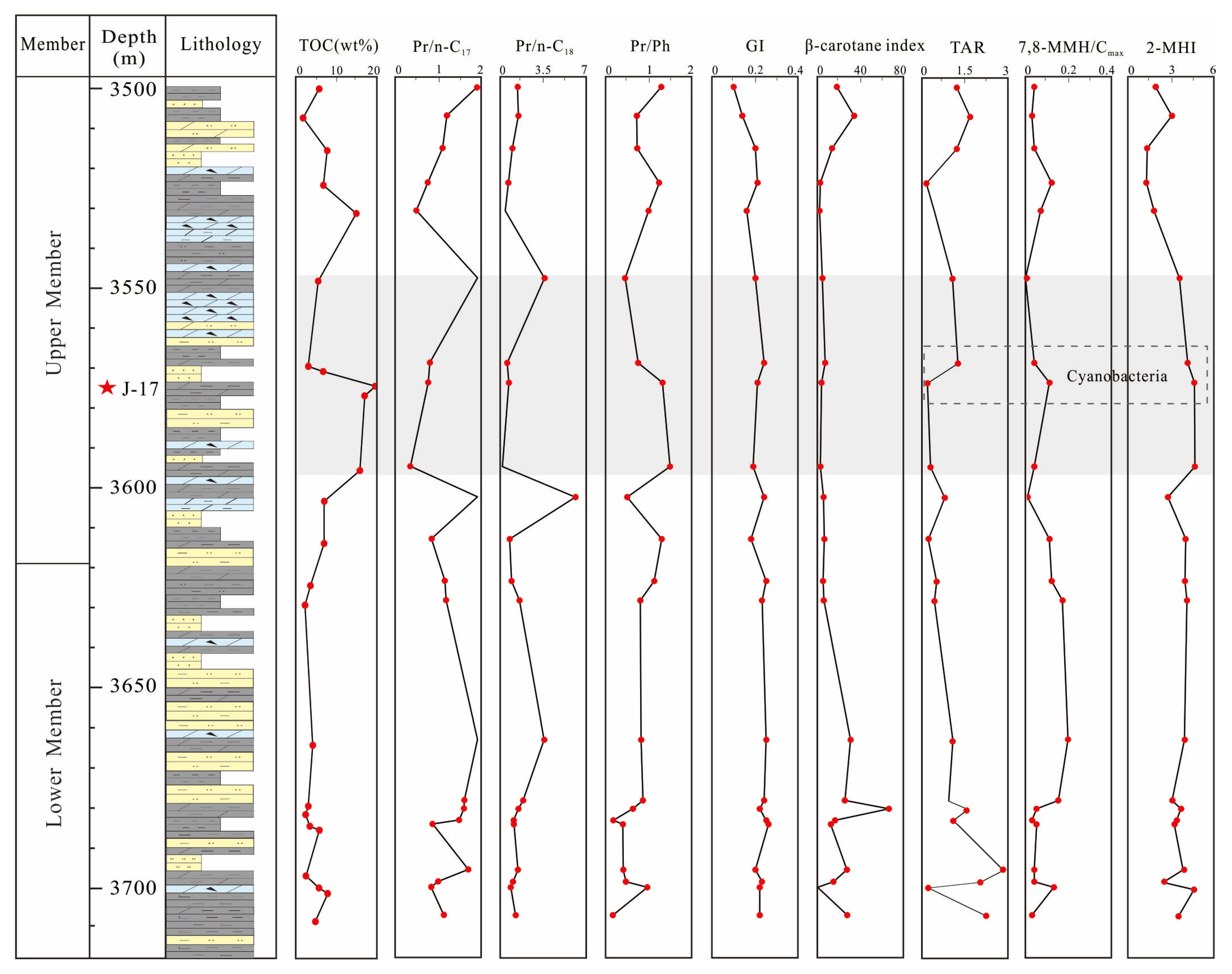
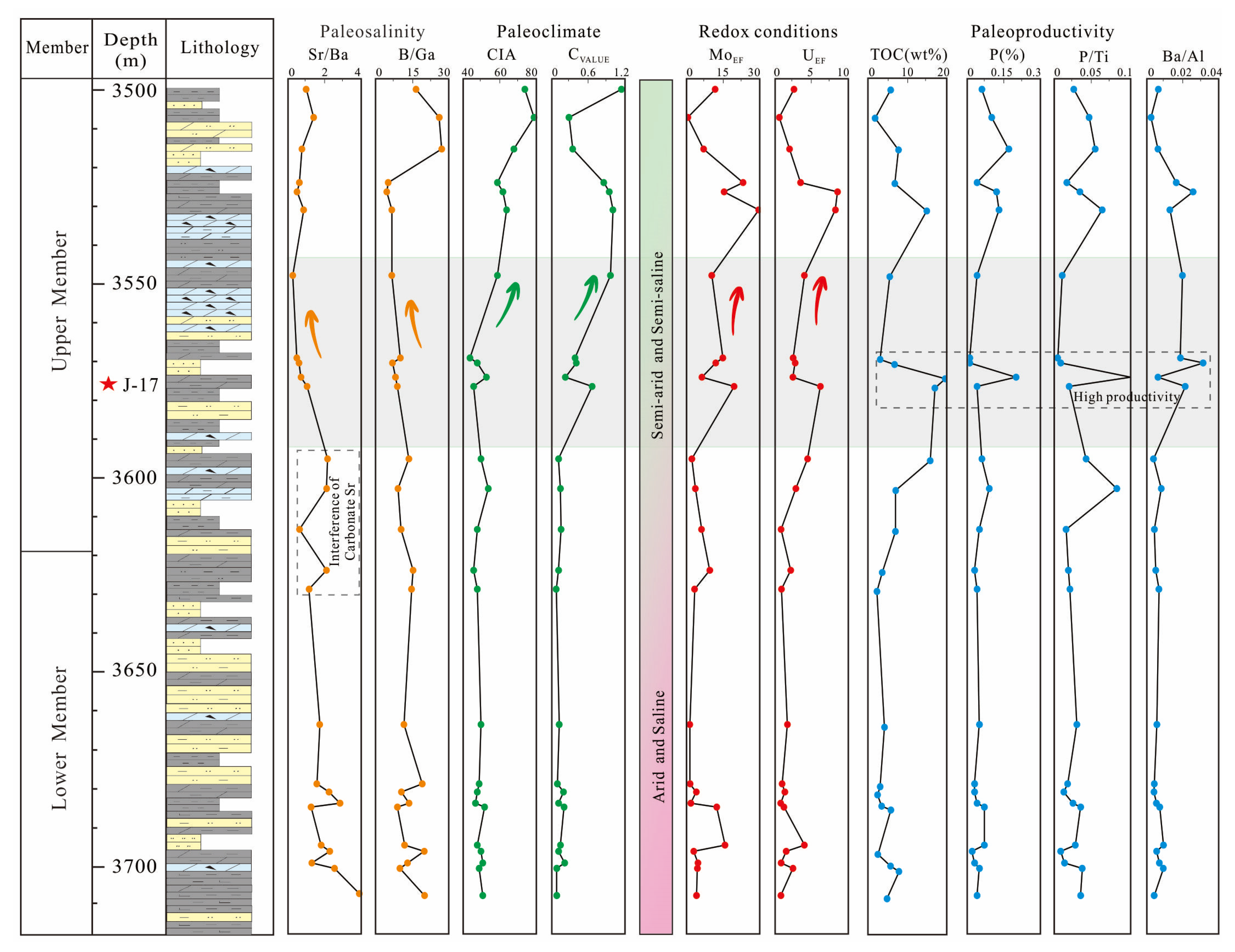
4.3. TOC and Elemental Geochemical Characteristics
5. Discussion
5.1. Biological Precursors of the Lucaogou Source Rocks

5.2. Paleoenvironmental Conditions of the Permian Microcystis Bloom
5.2.1. Environmental Factors Triggering Modern Cyanobacterial Blooms
5.2.2. Paleoclimate and Paleosalinity
5.2.3. Redox Conditions
5.2.4. Productivity
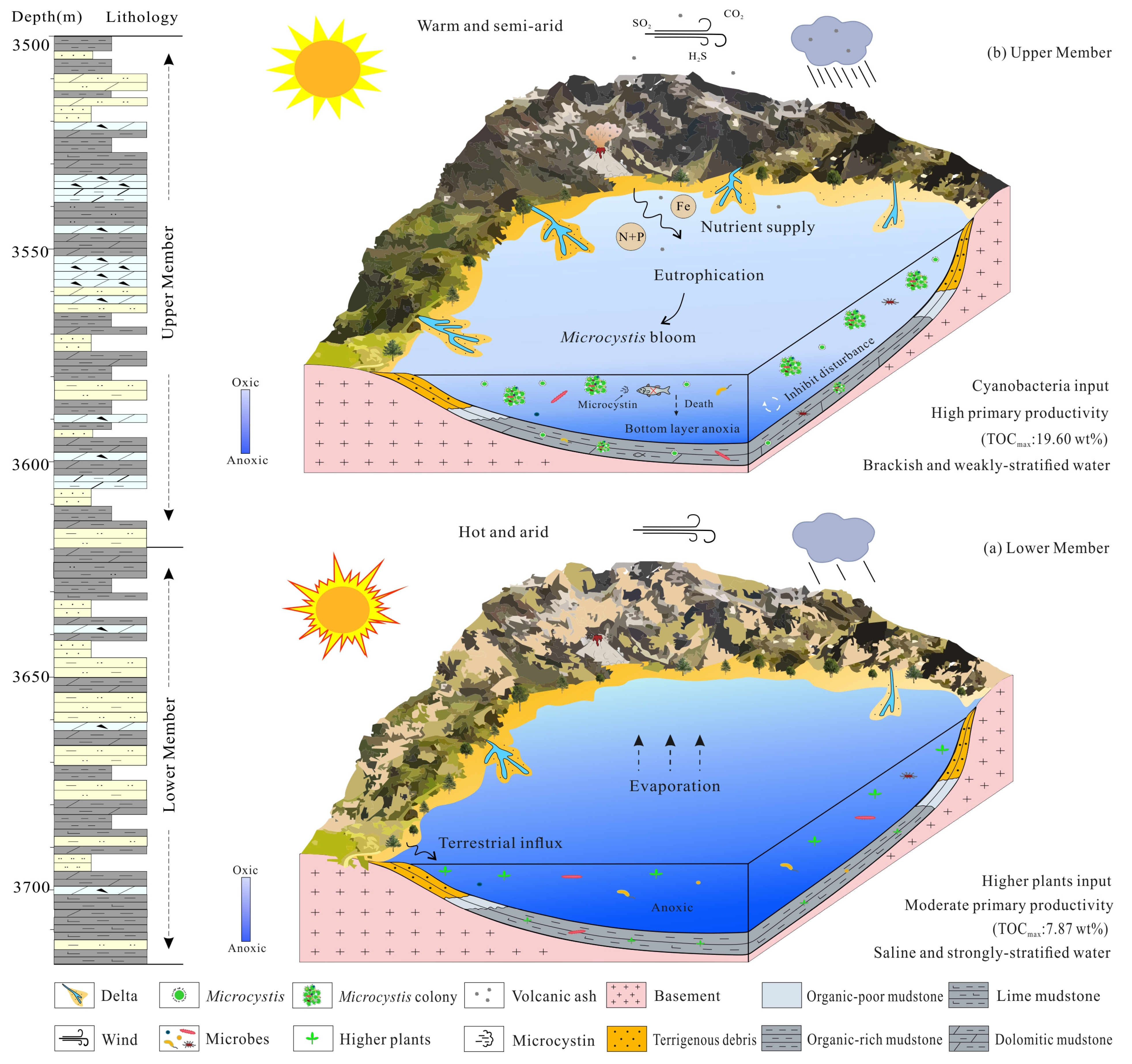
5.3. Mechanism of Organic Matter Enrichment under Cyanobacterial Blooms
6. Conclusions
- (1)
- Abundantly distributed, globular to elliptical shaped microfossils in the upper member of the Lucaogou Formation are interpreted as Microcystis, a kind of cyanobacteria. The great abundance of these fossils referred to the Chroococcaceae indicates the occurrence of Permian cyanobacterial blooms, which are further proven by cyanobacteria-derived biomarkers.
- (2)
- The upper member of the Lucaogou Formation was deposited in a brackish lacustrine environment with anoxic to suboxic water conditions, under a semiarid paleoclimate. Such a paleoenvironment together with abundant nutrients from volcanism ultimately led to cyanobacterial blooms in Permian lakes.
- (3)
- Permian Microcystis blooms contributed to organic matter enrichment in two ways. The first was by directly promoting the accumulation of algal biomass, and the second by creating an oxygen-depleted environment for better preservation of organic matter. This study thus provides new insight into shale oil exploration and development in the Junggar Basin, from the perspective of cyanobacterial blooms.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Latour, D.; Giraudet, H. Factors influencing the spatio-temporal distribution of benthic Microcystis aeruginosa colonies (Cyanobacteria) in the hypertrophic Grangent Reservoir (Loire, France). Comptes Rendus Biol. 2004, 327, 753–761. [Google Scholar] [CrossRef]
- Imai, H.; Chang, K.-H.; Kusaba, M.; Nakano, S.-I. Temperature-dependent dominance of Microcystis (Cyanophyceae) species: M. aeruginosa and, M. wesenbergii. J. Plankton Res. 2009, 31, 171–178. [Google Scholar] [CrossRef] [Green Version]
- Bullerjahn, G.S.; McKay, R.M.; Davis, T.W.; Baker, D.B.; Boyer, G.L.; Anglada, L.; Doucette, G.J. Global solutions to regional problems: Collecting global expertise to address the problem of harmful cyanobacterial blooms. A Lake Erie case study. Harmful Algae 2016, 54, 223–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schopf, J.W.; Packer, B.M. Early archean (3.3-billion to 3.5-billion-year-old) microfossils from warrawoona group. Aust. Sci. 1987, 237, 70–73. [Google Scholar] [CrossRef]
- Schopf, J.W.; Kudryavtsev, A.B.; Czaja, A.D.; Tripathi, A.B. Evidence of Archean life: Stromatolites and microfossils. Precambrian Res. 2007, 158, 141–155. [Google Scholar] [CrossRef]
- Kazmierczak, J.; Altermann, W. Paleontology-neoarchean biomineralization by benthic cyanobacteria. Science 2002, 298, 2351. [Google Scholar] [CrossRef]
- Yang, H.; Chen, Z.Q.; Papineau, D. Cyanobacterial spheroids and other biosignatures from microdigitate stromatolites of Mesoproterozoic Wumishan Formation in Jixian, North China. Precambrian Res. 2022, 368, 106496. [Google Scholar] [CrossRef]
- Whitton, B. Ecology of Cyanobacteria II: Their Diversity in Space and Time; Springer: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Riding, R. Temporal variation in calcification in marine cyanobacteria. J. Geol. Soc. Aust. 1992, 149, 979–989. [Google Scholar] [CrossRef]
- Lee, H.; Chen, I.; Chough, S.K. The middle–late Cambrian reef transition and related geological events: A review and new view. Earth-Sci. Rev. 2015, 145, 66–84. [Google Scholar] [CrossRef]
- Kremer, B.; Kamierczak, J. Cyanobacterial Mats from Silurian Black Radiolarian Cherts: Phototrophic Life at the Edge of Darkness? J. Sediment. Res. 2005, 75, 897–906. [Google Scholar] [CrossRef]
- Kremer, B. Mat-forming coccoid cyanobacteria from early Silurian marine deposits of Sudetes, Poland. Acta Palaeontol. Pol. 2009, 51, 143–154. [Google Scholar]
- Wu, Y.B.; Feng, Q.; Gong, Y.M. Blooming of bacteria and algae is a biokiller for mass-extinction of Devonian coral-stromatoporoid reef ecosystems. Sci. China-Earth Sci. 2013, 56, 1221–1232. [Google Scholar] [CrossRef]
- Kazmierczak, J.; Kremer, B.; Racki, G. Late Devonian marine anoxia challenged by benthic cyanobacterial mats. Geobiology 2012, 10, 371–383. [Google Scholar] [CrossRef]
- Wu, Y.S.; Yu, G.L.; Li, R.H.; Song, L.R.; Jiang, H.X.; Riding, R.; Liu, L.J.; Liu, D.Y.; Zhao, R. Cyanobacterial fossils from 252 Ma old microbialites and their environmental significance. Sci. Rep. 2014, 4, 3820. [Google Scholar] [CrossRef] [Green Version]
- Carroll, A.R. Upper Permian lacustrine organic facies evolution, southern Junggar Basin, NW China. Org. Geochem. 1998, 28, 649–667. [Google Scholar] [CrossRef]
- Pan, S.Q.; Horsfield, B.; Zou, C.N.; Yang, Z. Upper Permian Junggar and Upper Triassic Ordos lacustrine source rocks in Northwest and Central China: Organic geochemistry, petroleum potential and predicted organofacies. Int. J. Coal Geol. 2016, 158, 90–106. [Google Scholar] [CrossRef]
- Zhang, S.H.; Liu, C.Y.; Liang, H.; Wang, J.Q.; Yang, M.H.; Liu, G.H.; Huang, H.X.; Guan, Y.Z. Paleoenvironmental conditions, organic matter accumulation, and unconventional hydrocarbon potential for the Permian Lucaogou Formation organic-rich rocks in Santanghu Basin, NW China. Int. J. Coal Geol. 2018, 185, 44–60. [Google Scholar] [CrossRef]
- Cao, Z.; Liu, G.; Kong, Y.; Wang, C.; Niu, Z.; Zhang, J.; Geng, C.; Shan, X.; Wei, Z. Lacustrine tight oil accumulation characteristics: Permian Lucaogou Formation in Jimusaer Sag, Junggar Basin. Int. J. Coal Geol. 2016, 153, 37–51. [Google Scholar] [CrossRef]
- Gao, G.; Zhang, W.; Xiang, B.; Liu, G.; Ren, J. Geochemistry characteristics and hydrocarbon-generating potential of lacustrine source rock in Lucaogou Formation of the Jimusaer Sag, Junggar Basin. J. Pet. Sci. Eng. 2016, 145, 168–182. [Google Scholar] [CrossRef]
- Meng, Z.Y.; Liu, Y.Q.; Jiao, X.; Ma, L.T.; Zhou, D.W.; Li, H.; Cao, Q.; Zhao, M.R.; Yang, Y.Y. Petrological and organic geochemical characteristics of the Permian Lucaogou Formation in the Jimsar Sag, Junggar Basin, NW China: Implications on the relationship between hydrocarbon accumulation and volcanic-hydrothermal activities. J. Pet. Sci. Eng. 2022, 210, 110078. [Google Scholar] [CrossRef]
- Carroll, A.R.; Brassell, S.C.; Graham, S.A. Upper Permian lacustrine oil shales, southern Junggar Basin, Northwest China. AAPG Bull. 1992, 76, 1874–1902. [Google Scholar]
- Tao, S.; Wang, Y.B.; Tang, D.Z.; Wu, D.M.; Xu, H.; He, W. Organic petrology of Fukang Permian Lucaogou Formation oil shales at the northern foot of Bogda Mountain, Junggar Basin, China. Int. J. Coal Geol. 2012, 99, 27–34. [Google Scholar] [CrossRef]
- Xie, X.M.; Borjigin, T.; Zhang, Q.Z.; Zhang, Z.R.; Qin, J.Z.; Bian, L.Z.; Volkman, J.K. Intact microbial fossils in the Permian Lucaogou Formation oil shale, Junggar Basin, NW China. Int. J. Coal Geol. 2015, 146, 166–178. [Google Scholar] [CrossRef]
- Su, Y.; Zha, M.; Ding, X.J.; Qu, J.X.; Cao, C.H.; Jin, J.H.; Iglauer, S. Petrographic, palynologic and geochemical characteristics of source rocks of the Permian Lucaogou formation in Jimsar Sag, Junggar Basin, NW China: Origin of organic matter input and depositional environments. J. Pet. Sci. Eng. 2019, 183, 106364. [Google Scholar] [CrossRef]
- Liu, Y.Q.; Zhou, D.W.; Jiao, X.; Feng, Q.; Zhou, X.H. A preliminary study on the relationship between deep-sourced materials and hydrocarbon generation in lacustrine source rocks: An example from the Permian black rock series in Jimusar sag, Junggar Basin. J. Palaeogeogr. 2019, 21, 16. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Zhuang, X.G.; Tu, Q.J.; Xv, S.Q.; Zhang, Y. Depositional process and mechanism of organic matter accumulation of Lucaogou shale in Southern Junggar Basin, Northwest China. Earth Sci. 2018, 43, 538–550. [Google Scholar]
- Jiang, Y.; Hou, D.; Li, H.; Zhang, Z.; Guo, R. Impact of the Paleoclimate, Paleoenvironment, and Algae Bloom: Organic Matter Accumulation in the Lacustrine Lucaogou Formation of Jimsar Sag, Junggar Basin, NW China. Energies 2020, 13, 1488. [Google Scholar] [CrossRef] [Green Version]
- Kuang, L.C.; Tang, Y.; Lei, D.W.; Chang, Q.S.; Ou, Y.M.; Hou, L.H.; Liu, D.G. Formation conditions and exploration potential of tight oil in the Permian saline lacustrine dolomitic rock, Junggar Basin, NW China. Pet. Explor. Dev. 2012, 039, 657–667. [Google Scholar]
- Ding, X.J.; Qu, J.Q.; Imin, A.; Zha, M.; Jiang, Z.F.; Jiang, H. Organic matter origin and accumulation in tuffaceous shale of the lower Permian Lucaogou Formation, Jimsar Sag. J. Pet. Sci. Eng. 2019, 179, 696–706. [Google Scholar] [CrossRef]
- Sun, F.N.; Hu, W.X.; Wang, X.L.; Cao, J.; Fu, B.; Wu, H.G.; Yang, S.C. Methanogen microfossils and methanogenesis in Permian lake deposits. Geology 2021, 49, 13–18. [Google Scholar] [CrossRef]
- Li, Y.; Xue, Z.; Cheng, Z.; Jiang, H.; Wang, R. Progress and development directions of deep oil and gas exploration and development in China. China Pet. Explor. 2020, 25, 45–57. [Google Scholar]
- Carroll, A.R.; Graham, S.A. Late paleozoic tectonic amalgamation of northwestern China: Sedimentary record of the northern Tarim, northern Turpan, and southern Junggar Basins. Geol. Soc. Am. Bull. 1995, 107, 571–594. [Google Scholar] [CrossRef]
- Kuang, L.H.; Hu, W.X.; Wang, X.L.; Wu, H.G.; L. W.X. Research of the Tight Oil Reservoir in the Lucaogou Formation in Jimusar Sag: Analysis of Lithology and Porosity Characteristics. Geol. J. China Univ. 2013, 19, 7. [Google Scholar]
- Wang, J.; Yuan, B.; Liu, J.; Li, Y.; Li, E. Genesis and pore development characteristics of Permian Lucaogou migmatites, Jimsar Sag, Junggar Basin. Pet. Geol. Exp. 2022, 44, 413–424. [Google Scholar]
- Jiang, Y.Q.; Liu, Y.Q.; Yang, Z.; Nan, Y.; Wang, R.; Zhou, P.; Yang, Y.J.; Kou, J.Y.; Zhou, N.C. Characteristics and origin of tuff-type tight oil in Jimusar Depression, Junggar Basin, NW China. Pet. Explor. Dev. 2015, 42, 741–749. [Google Scholar] [CrossRef]
- Tao, K.Y.; Cao, J.; Chen, X.; Nueraili, Z.; Hu, W.X.; Shi, C.H. Deep hydrocarbons in the northwestern Junggar Basin (NW China): Geochemistry, origin, and implications for the oil vs. gas generation potential of post-mature saline lacustrine source rocks. Mar. Pet. Geol. 2019, 109, 623–640. [Google Scholar] [CrossRef]
- Guo, P.; Wen, H.G.; Gibert, L.; Jin, J.; Wang, J.; Lei, H.Y. Deposition and diagenesis of the Early Permian volcanic-related alkaline playa-lake dolomitic shales, NW Junggar Basin, NW China. Mar. Pet. Geol. 2021, 123, 104780. [Google Scholar] [CrossRef]
- Wang, X.; Yang, Z.; Guo, X.; Xiatian, W.; Feng, Y.; Huang, L. Practices and Prospects of Shale Oil Exploration in Jimsar Sag of Junggar Basin. Xinjiang Pet. Geol. 2019, 40, 402–413. [Google Scholar]
- Zhi, D.; Song, Y.; He, W.; Jia, X.; Yang, Z.; Huang, L. Geological Characteristics, Resource Potential and Exploration Direction of Shale Oil in Middle-Lower Permian, Junggar Basin. Xinjiang Pet. Geol. 2019, 40, 389–401. [Google Scholar]
- Kang, J.L.; Wang, J.H.; Ma, Q.; Li, C.Q.; Chen, X. Fine-grained sublacustrine fan deposits and their significance of shale oil reservoirs in the Lucaogou Formation in Jimsar Sag, Junggar Basin. Bull. Geol. Sci. Technol. 2022, 4, 1–12. [Google Scholar]
- Wu, H.; Hu, W.; Tang, Y.; Cao, J.; Wang, X.; Wang, Y.; Kang, X. The impact of organic fluids on the carbon isotopic compositions of carbonate-rich reservoirs: Case study of the Lucaogou Formation in the Jimusaer Sag, Junggar Basin, NW China. Mar. Pet. Geol. 2017, 85, 136–150. [Google Scholar]
- Tissot, B.; Welte, D. Petroleum Formation and Occurrence; Earth Science Reviews; Springer: Berlin/Heidelberg, Germany, 1984. [Google Scholar]
- Peters, K.E.; Walters, C.C.; Moldowan, J.M. The Biomarker Guide: Interpreting Molecular Fossils in Petroleum and Ancient Sediments; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Calvert, S.E.; Pedersen, T.F. Geochemistry of Recent oxic and anoxic marine sediments: Implications for the geological record. Mar. Geol. 1993, 113, 67–88. [Google Scholar] [CrossRef]
- Tribovillard, N.; Algeo, T.; Lyons, T.; Riboulleau, A. Trace metals as paleoredox and paleoproductivity proxies: An update. Chem. Geol. 2006, 232, 12–32. [Google Scholar] [CrossRef]
- Mclennan; Scott, M. Relationship between the trace element composition of sedimentary rocks and upper continental crust. Geochem. Geophys. Geosystems 2001, 2, 203–236. [Google Scholar]
- Nesbitt, H.W.; Young, G.M. Early Proterozoic climates and plate motions inferred from major element chemistry of lutites. Nature 1982, 299, 715–717. [Google Scholar] [CrossRef]
- Mclennan; Scott, M. 100th Anniversary Symposium: Evolution of the Earth’s Surface || Weathering and Global Denudation. J. Geol. 1993, 101, 295–303. [Google Scholar]
- Liu, J.; Wang, J.; Zhang, X.G.; Luo, Z.J.; Shang, L.; Wang, G.J. Scanning electron microscope study on oil generation characteristics of ostracod fossils from the Lucaogou formation in eastern Junggar Basin. J. Chin. Electron Microsc. Soc. 2021, 40, 270–278. [Google Scholar]
- Wu, H.G.; Hu, W.X.; Cao, J.; Wang, X.L.; Liao, Z.W. A unique lacustrine mixed dolomitic-clastic sequence for tight oil reservoir within the middle Permian Lucaogou Formation of the Junggar Basin, NW China: Reservoir characteristics and origin. Mar. Pet. Geol. 2016, 76, 115–132. [Google Scholar] [CrossRef]
- Wang, R.Y.; Ding, W.L.; Zhang, Y.Q.; Wang, Z.; Wang, X.; He, J.; Zeng, W.; Dai, P. Analysis of developmental characteristics and dominant factors of fractures in Lower Cambrian marine shale reservoirs: A case study of Niutitang formation in Cen’gong block, southern China. J. Pet. Sci. Eng. 2016, 138, 31–49. [Google Scholar] [CrossRef]
- Caracciolo, A.B.; Dejana, C.; Fajardo, P.; Grenni, M. A new fluorescent oligonucleotide probe for in-situ identification of Microcystis aeruginosa in freshwater. Microchem. J. 2019, 148, 503–513. [Google Scholar]
- Taylor, T.N.; Krings, M. A colony-forming microorganism with probable affinities to the Chroococcales (cyanobacteria) from the Lower Devonian Rhynie chert. Rev. Palaeobot. Palynol. 2015, 219, 147–156. [Google Scholar] [CrossRef]
- Xiao, M.; Li, M.; Reynolds, C.S. Colony formation in the cyanobacterium Microcystis. Biol. Rev. 2018, 93, 1399–1420. [Google Scholar] [CrossRef] [Green Version]
- Phelan, R.R.; Downing, T.G. A growth advantage for microcystin production by Microcystis PCC7806 under high light. J. Phycol. 2011, 47, 1241–1246. [Google Scholar] [CrossRef]
- Van Le, V.; Srivastava, A.; Ko, S.-R.; Ahn, C.-Y.; Oh, H.-M. Microcystis colony formation: Extracellular polymeric substance, associated microorganisms, and its application. Bioresour. Technol. 2022, 360, 127610. [Google Scholar]
- Wei, W.; Algeo, T.J. Elemental proxies for paleosalinity analysis of ancient shales and mudrocks. Geochim. Et Cosmochim. Acta 2020, 287, 341–366. [Google Scholar] [CrossRef]
- Gelpi, E.; Schneider, J.; Mann, J.; Oró, J. Hydrocarbons of geochemical significance in microscopic algae. Phytochemistry 1970, 9, 603–612. [Google Scholar] [CrossRef]
- Reed, J.D.; Illich, H.A.; Horsfield, B.J.O.G. Biochemical evolutionary significance of Ordovician oils and their sources. Org. Geochem. 1986, 10, 347–358. [Google Scholar] [CrossRef]
- Cranwell, P.A.; Eglinton, G.; Robinson, N. Lipids of aquatic organisms as potential contributors to lacustrine sediments-II. Org. Geochem. 1987, 11, 513–527. [Google Scholar] [CrossRef]
- Huang, W.Y.; Meinschein, W.G. Sterols as ecological indicators. Geochim. Et Cosmochim. Acta 1979, 43, 739–745. [Google Scholar] [CrossRef]
- Volkman, J. Sterols in microorganisms. Appl. Microbiol. Biotechnol. 2003, 60, 495–506. [Google Scholar] [CrossRef] [PubMed]
- Grantham, P.J.; Wakefield, L.L. Variations in the sterane carbon number distributions of marine source rock derived crude oils through geological time. Org. Geochem. 1988, 12, 61–73. [Google Scholar] [CrossRef]
- Strobl, S.; Sachsenhofer, R.F.; Bechtel, A.; Gratzer, R.; Gross, D.; Bokhari, S.; Liu, R.; Liu, Z.; Meng, Q.; Sun, P. Depositional environment of oil shale within the Eocene Jijuntun Formation in the Fushun Basin (NE China). Mar. Pet. Geol. 2014, 56, 166–183. [Google Scholar] [CrossRef]
- Summons, R.E.; Jahnke, L.L.; Hope, J.M.; Logan, G.A. 2-Methylhopanoids as biomarkers for cyanobacterial oxygenic photosynthesis. Nature 1999, 400, 554–557. [Google Scholar] [CrossRef] [PubMed]
- Hou, M.; Qu, J.; Zha, M.; Swennen, R.; Ding, X.; Imin, A.; Liu, H.; Bian, B. Significant contribution of haloalkaliphilic cyanobacteria to organic matter in an ancient alkaline lacustrine source rock: A case study from the Permian Fengcheng Formation, Junggar Basin, China. Mar. Pet. Geol. 2022, 138, 105546. [Google Scholar] [CrossRef]
- Doughty, D.M.; Hunter, R.C.; Summons, R.E.; Newman, D.K. 2-Methylhopanoids are maximally produced in akinetes of Nostoc punctiforme: Geobiological implications. Geobiology 2009, 7, 524–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boudou, J.P.; Trichet, J.; Robinson, N.; Brassell, S.C. Profile of aliphatic hydrocarbons in a recent polynesian microbial mat. Int. J. Environ. Anal. Chem. 1986, 26, 137–155. [Google Scholar] [CrossRef]
- Ding, W.J.; Hou, D.J.; Jiang, L.; Jiang, Y.H.; Wu, P. High abundance of carotanes in the brackish-saline lacustrine sediments: A possible cyanobacteria source? Int. J. Coal Geol. 2020, 219, 103373. [Google Scholar] [CrossRef]
- Dobson, G.; Ward, D.M.; Robinson, N.; Eglinton, G. Biogeochemistry of hot spring environments: Extractable lipids of a cyanobacterial mat. Chem. Geol. 1988, 68, 155–179. [Google Scholar] [CrossRef]
- Luo, G.; Hallmann, C.; Xie, S.; Ruan, X.; Summons, R.E. Comparative microbial diversity and redox environments of black shale and stromatolite facies in the Mesoproterozoic Xiamaling Formation. Geochim. Et Cosmochim. Acta 2015, 151, 150–167. [Google Scholar] [CrossRef]
- Ding, X.J.; Gao, C.H.; Zha, M.; Chen, H.; Su, Y. Depositional environment and factors controlling β-carotane accumulation: A case study from the Jimsar Sag, Junggar Basin, northwestern China. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2017, 485, 833–842. [Google Scholar] [CrossRef]
- Tribovillard, N.; Algeo, T.J.; Baudin, F.; Riboulleau, A.J.C.G. Analysis of marine environmental conditions based onmolybdenum–uranium covariation—Applications to Mesozoic paleoceanography. Chem. Geol. 2012, 324–325, 46–58. [Google Scholar] [CrossRef]
- Shanmugam, G. Significance of coniferous rain forests and related organic matter in generating commercial quantities of oil, Gippsland Basin, Australia. AAPG (Am. Assoc. Pet. Geol.) Bull. 1985, 69, 1241–1254. [Google Scholar] [CrossRef]
- Burford, M.A.; Gobler, C.J.; Hamilton, D.P.; Visser, P.M.; Lurling, M.; Codd, G.A. Solutions for Managing Cyanobacterial Blooms: A Scientific Summary for Policy Makers; IOC/UNESCO: Paris, France, 2019. [Google Scholar]
- Ji, X.; Verspagen, J.M.H.; Van de Waal, D.B.; Rost, B.; Huisman, J. Phenotypic plasticity of carbon fixation stimulates cyanobacterial blooms at elevated CO2. Sci. Adv. 2020, 6, 2926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verspagen, J.M.H.; Snelder, E.; Visser, P.M.; Huisman, J.; Ibelings, B.W. Recruitment of benthic Microcystis (Cyanophyceae) to the water column: Internal buoyancy changes or resuspension? J. Phycol. 2004, 40, 260–270. [Google Scholar] [CrossRef]
- Preston, T.; Stewart, W.D.P.; Reynolds, C.S. Bloomforming cyanobacterium Microcystis aeruginosa overwinters on the sediment. Nature 1980, 288, 365–367. [Google Scholar] [CrossRef]
- Schindler, D.W. Evolution of phosphorus limitation in lakes. Science 1977, 195, 260–262. [Google Scholar] [CrossRef] [Green Version]
- Paerl, H.W. Mitigating harmful cyanobacterial blooms in a human-and climatically-impacted world. Life 2014, 4, 988–1012. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Du, Y.; Yu, W.; Algeo, T.J.; Pan, W. The chemical index of alteration (CIA) as a proxy for climate change during glacial-interglacial transitions in Earth history. Earth-Sci. Rev. 2019, 201, 103032. [Google Scholar] [CrossRef]
- Moradi, A.V.; Sari, A.; Akkaya, P. Geochemistry of the Miocene oil shale (Hançili Formation) in the Çankırı-Çorum Basin, Central Turkey: Implications for Paleoclimate conditions, source–area weathering, provenance and tectonic setting. Sediment. Geol. 2016, 341, 289–303. [Google Scholar] [CrossRef]
- Xie, W.Q.; Tan, J.Q.; Wang, W.H.; Schulz, H.; Liu, Z.J.; Kang, X.; Wang, Y.; Shahzad, A.; Jan, I.U. Middle Jurassic lacustrine source rocks controlled by an aridification event: A case study in the northern Qaidam Basin (NW China). Int. J. Coal Geol. 2021, 242, 103779. [Google Scholar] [CrossRef]
- Rohling, E.J. Paleosalinity: Confidence limits and future applications. Mar. Geol. 2000, 163, 1–11. [Google Scholar] [CrossRef]
- Jiang, Z.; Ding, X.; Wang, Z.; Zhao, X. Sedimentary paleoenvironment of source rocks of Permian Lucaogou Formation in Jimsar Sag. Lithologic Reservoirs. Lithol. Reserv. 2020, 32, 11. [Google Scholar]
- Salminen, R.; Batista, M.J.; Bidovec, M.; Demetriades, A.; Tarvainen, T. FOREGS Geochemical Atlas of Europe, Part 1: Background Information, Methodology and Maps; Electric publication; Geochemical Survey of Finland: Espoo, Finland, 2005. [Google Scholar]
- Summons, R.E.; Hope, J.M.; Swart, R.; Walter, M.R. Origin of Nama Basin bitumen seeps: Petroleum derived from a Permian lacustrine source rock traversing southwestern Gondwana. Org. Geochem. 2008, 39, 589–607. [Google Scholar] [CrossRef]
- Algeo, T.J.; Tribovillard, N. Environmental Analysis of Paleoceanographic Systems Based on Molybdenum–Uranium Covariation. Chem. Geol. 2009, 268, 211–225. [Google Scholar] [CrossRef]
- Liu, D.D.; Fan, Q.Q.; Zhang, C.; Gao, Y.; Du, W.; Song, Y. Paleoenvironment evolution of the Permian Lucaogou Formation in the southern Junggar Basin, NW China. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2022, 603, 111198. [Google Scholar] [CrossRef]
- Walters, C. The Biomarker Guide; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Peters, K.E.; Fraser, T.H.; Amris, W.; Rustanto, B. Geochemistry of crude oils from eastern Indonesia. AAPG Bull. 1999, 83, 1927–1942. [Google Scholar]
- Jiang, Z.; Fowler, M.G. Carotenoid-derived alkanes in oils from northwestern China. Org. Geochem. 1986, 10, 831–839. [Google Scholar] [CrossRef]
- Wang, R.Y.; Gu, Y.; Ding, W.L.; Gong, D.; Yin, S.; Wang, X.; Zhou, X.; Li, A.; Xiao, Z.; Cui, Z. Characteristics and dominant controlling factors of organic-rich marine shales with high thermal maturity: A case study of the Lower Cambrian Niutitang Formation in the Cen’gong block, southern China. J. Nat. Gas Sci. Eng. 2016, 33, 81–96. [Google Scholar] [CrossRef]
- Calvert, S.; Pedersen, T. Elemental proxies for paleoclimatic and palaeoceanographic variability in marine sediments: Interpretation and application. Dev. Mar. Geol. 2007, 1, 568–644. [Google Scholar]
- Cheng, D.W.; Zhou, C.M.; Zhang, Z.J.; Yuan, X.J.; Liu, Y.H.; Chen, X.Y. Paleo-Environment Reconstruction of the Middle Permian Lucaogou Formation, Southeastern Junggar Basin, NW China: Implications for the Mechanism of Organic Matter Enrichment in Ancient Lake. J. Earth Sci. 2022, 33, 963–976. [Google Scholar] [CrossRef]
- Algeo, T.J.; Kuwahara, K.; Sano, H.; Bates, S.; Lyons, T.; Elswick, E.; Hinnov, L.; Ellwood, B.; Moser, J.; Maynard, J.B. Spatial variation in sediment fluxes, redox conditions, and productivity in the Permian–Triassic Panthalassic Ocean. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2011, 308, 65–83. [Google Scholar] [CrossRef]
- Shen, J.; Schoepfer, S.D.; Feng, Q.; Zhou, L.; Yu, J.; Song, H.; Wei, H.; Algeo, T.J. Marine productivity changes during the end-Permian crisis and Early Triassic recovery. Earth Sci. Rev. 2015, 149, 136–162. [Google Scholar] [CrossRef]
- Schoepfer, S.D.; Shen, J.; Wei, H.; Tyson, R.V.; Ingall, E.; Algeo, T.J.J.E.-S.R. Total organic carbon, organic phosphorus, and biogenic barium fluxes as proxies for paleomarine productivity. Earth Sci. Rev. 2015, 149, 23–52. [Google Scholar] [CrossRef]
- Pedersen, T.E.; Calvert, S.E. Anoxia vs. productivity: What controls the formation of organic-carbon-rich sediments and sedimentary rocks? Am. Assoc. Pet. Geol. Bull. 1990, 74, 454–466. [Google Scholar]
- Guan, M.; Wu, S.; Hou, L.; Jiang, X.; Ba, D.; Hua, G. Paleoenvironment and chemostratigraphy heterogenity of the Cretaceous organic-rich shales. Adv. Geo-Energy Res. 2021, 5, 444–445. [Google Scholar] [CrossRef]
- Goldhammer, T.; Brüchert, V.; Ferdelman, T.G.; Zabel, M. Microbial sequestration of phosphorus in anoxic upwelling sediments. Nat. Geosci. 2010, 3, 557–561. [Google Scholar] [CrossRef]
- Langmann, B. On the Role of Climate Forcing by Volcanic Sulphate and Volcanic Ash. Adv. Meteorol. 2014, 2014, 340123. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.S.; Huang, Z.L.; Li, T.J.; Guo, X.; Chen, X. Environmental response to volcanic activity and its effect on organic matter enrichment in the Permian Lucaogou Formation of the Malang Sag, Santanghu Basin, Northwest China. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2020, 560, 110024. [Google Scholar] [CrossRef]
- Liao, Z.; Hu, W.; Cao, J.; Wang, X.; Hu, Z.J.M.; Geology, P. Petrologic and geochemical evidence for the formation of organic-rich siliceous rocks of the Late Permian Dalong Formation, Lower Yangtze region, southern China. Mar. Pet. Geol. 2019, 103, 41–54. [Google Scholar] [CrossRef]
- Anderson, L.D.; Delaney, M.L. Use of multiproxy records on the Agulhas Ridge, Southern Ocean (Ocean Drilling Project Leg 177, Site 1090) to investigate sub-Antarctic hydrography from the Oligocene to the early Miocene. Paleoceanogr. Palaeoclimatol. 2005, 20. [Google Scholar] [CrossRef]
- Rimmer, S.M.; Thompson, J.A.; Goodnight, S.A.; Robl, T.L. Multiple controls on the preservation of organic matter in Devonian–Mississippian marine black shales: Geochemical and petrographic evidence. Palaeogeogr. Palaeoclim. Palaeoecol. 2004, 215, 125–154. [Google Scholar] [CrossRef]
- Duggen, S.; Croot, P.; Schacht, U.; Hoffmann, L. Subduction zone volcanic ash can fertilize the surface ocean and stimulate phytoplankton growth: Evidence from biogeochemical experiments and satellite data. Geophys. Res. Lett. 2007, 34, 95–119. [Google Scholar] [CrossRef] [Green Version]
- Wignall, P.B. Large igneous provinces and mass extinctions. Earth-Sci. Rev. 2001, 53, 1–33. [Google Scholar] [CrossRef]
- Qu, C.S.; Qiu, L.W.; Yang, Y.Q.; Yu, K.H.; Tang, L.L.; Wan, M. Environmental response of the Permian volcanism in Lucaogou formation in Jimsar sag, Junggar Basin, Northwest China. Seismol. Geol. 2019, 560, 110024. [Google Scholar]
- Li, Y.F.; Zhang, T.W.; Ellis, G.S. Depositional environment and organic matter accumulation of Upper Ordovician-Lower Silurian marine shale in the Upper Yangtze Platform, South China. Palaeogeogr. Palaeoclimatol. Palaeoecol 2017, 466, 252–264. [Google Scholar] [CrossRef]
- Wang, R.Y.; Hu, Z.Q.; Zhou, T.; Bao, H.Y.; Wu, J.; Du, W.; He, J.H. Characteristics of fractures and their significance for reservoirs in Wufeng-Longmaxi shale, Sichuan Basin and its periphery. Oil Gas Geol. 2021, 42, 1295–1306. [Google Scholar]
- Zhao, D.F.; Guo, Y.H.; Yin, S.; Ren, C.Y.; Wang, Y.J. Prediction of geomechanical sweet spots in a tight gas sandstone reservoir: A case study of lower Permian strata in the southern Qinshui Basin, China. Interpretation 2019, 7, 207–219. [Google Scholar] [CrossRef]
- Zhao, D.F.; Guo, Y.H.; Wang, G.X.; Zhou, X.Q.; Zhou, Y.Y.; Zhang, J.M.; Ren, G.Y. Organic matter enrichment mechanism of Youganwo Formation oil shale in the Maoming Basin. Heliyon 2023, 9, 13173. [Google Scholar] [CrossRef]
- Hedges, J.I.; Keil, R.G. Sedimentary organic matter preservation: An assessment and speculative synthesis. Mar. Chem. 1995, 49, 81–115. [Google Scholar] [CrossRef]
- Lash, G.G.; Blood, D.R. Organic matter accumulation, redox, and diagenetic history of the Marcellus Formation, southwestern Pennsylvania, Appalachian basin. Mar. Pet. Geol. 2014, 57, 244–263. [Google Scholar] [CrossRef]
- Zhao, D.F.; Guo, Y.H.; Zhu, Y.M.; Zhao, S.X.; Chen, Z.H.; Jiao, W.W. Comments on the evaluation system of accurate evaluation and selection of deep marine shale reservoirs. Unconv. Oil Gas 2022, 9, 1–7. [Google Scholar]
- Paerl, H.W.; Huisman, J. Climate change: A catalyst for global expansion of harmful cyanobacterial blooms. Environ. Microbiol. Rep. 2009, 1, 27–37. [Google Scholar] [CrossRef]
- O’Neil, J.M.; Davis, T.W.; Burford, M.A.; Gobler, C.J. The rise of harmful cyanobacteria blooms: The potential roles of eutrophication and climate change. Harmful Algae 2012, 14, 313–334. [Google Scholar]
- Verspagen, J.M.H.; Van, D.; Finke, J.F.; Visser, P.M.; Donk, E.V.; Huisman, J.; Dam, H.G. Rising CO2 Levels Will Intensify Phytoplankton Blooms in Eutrophic and Hypertrophic Lakes. PLoS ONE 2014, 9, 104325. [Google Scholar]
- Huisman, J.; Codd, G.A.; Paerl, H.; Ibelings, B.W.; Verspagen, J.M.H.; Visser, P.M. Cyanobacterial blooms. Nat. Rev. Microbiol. 2018, 16, 471–483. [Google Scholar] [CrossRef]
- Hackley, P.C.; Fishman, N.; Wu, T.; Baugher, G. Organic petrology and geochemistry of mudrocks from the lacustrine Lucaogou Formation, Santanghu Basin, northwest China: Application to lake basin evolution. Int. J. Coal Geol. 2016, 168, 20–34. [Google Scholar] [CrossRef] [Green Version]
- Zheng, R.; Zeng, W.; Li, Z.; Chen, X.; Man, K.; Zhang, Z.; Wang, G.; Shi, S. Differential enrichment mechanisms of organic matter in the Chang 7 Member mudstone and shale in Ordos Basin, China: Constraints from organic geochemistry and element geochemistry. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2022, 601, 111126. [Google Scholar] [CrossRef]
- Bohacs, K.M.; Grabowski, J.G.; Carroll, A.R.; Mankiewicz, P.J.; Simo, T.J. Production, Destruction, and Dilution—The Many Paths to Source-Rock Development. In The Deposition of Organic-Carbon-Rich Sediments: Models, Mechanisms, and Consequences; Society for Sedimentary Geology Special Publication; GeoScienceWorld: McLean, VA, USA, 2005; Volume 82, pp. 61–101. [Google Scholar]
- Paerl, H.W.; Tucker, C.S. Ecology of Blue-Green Algae in Aquaculture Ponds. J. World Aquac. Soc. 1995, 26, 109–131. [Google Scholar] [CrossRef]
- Elliott, J.A. The seasonal sensitivity of Cyanobacteria and other phytoplankton to changes in flushing rate and water temperature. Glob. Change Biol. 2010, 16, 864–876. [Google Scholar] [CrossRef]
- Qin, J.Z.; Pan, A.; Shen, B.J. Insoluble organic matter in source rocks: Derived from organic macromolecules in the skeleton, cell wall and shell of organisms. Pet. Geol. Exp. 2020, 42, 6. [Google Scholar]
- Yang, Y.; Zheng, L.; Wu, Y.L.; Yang, F.Y.; Liu, F.; Han, R. Research progress on hydrodynamic mechanisms and simulation of Microcystis blooms. J. China Inst. Water Resour. Hydropower Res. 2022, 20, 449–463. [Google Scholar]
- Chorus, I. Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management; E&FN Spon: London, UK, 1999. [Google Scholar]
- Song, H.; Xu, J.; Lavoie, M.; Fan, X.; Liu, G.; Sun, L.; Fu, Z.; Qian, H.J.A.M.; Biotechnology. Biological and chemical factors driving the temporal distribution of cyanobacteria and heterotrophic bacteria in a eutrophic lake (West Lake, China). Appl. Microbiol. Biotechnol. 2017, 101, 1685–1696. [Google Scholar] [CrossRef]
- Wu, Y.S.; Yu, G.L.; Jiang, H.X.; Liu, L.J.; Zhao, R.; Wu, Y.S.; Yu, G.L.; Jiang, H.X.; Liu, L.J.; Zhao, R. Role and lifestyle of calcified cyanobacteria (Stanieria) in Permian-Triassic boundary microbialites. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2015, 448, 9. [Google Scholar] [CrossRef]
- Mort, H.; Jacquat, O.; Adatte, T.; Steinmann, P.; Föllmi, K.; Matera, V.; Berner, Z.; Stüben, D. The Cenomanian/Turonian anoxic event at the Bonarelli Level in Italy and Spain: Enhanced productivity and/or better preservation? Cretac. Res. 2007, 28, 597–612. [Google Scholar] [CrossRef] [Green Version]
- Xie, S.; Pancost, R.D.; Huang, J.H.; Wignall, P.B.; Yu, J.; Tang, X.; Chen, L.; Huang, X.; Lai, X. Changes in the global carbon cycle occurred as two episodes during the Permian-Triassic crisis. Geology 2007, 35, 1083–1086. [Google Scholar] [CrossRef]
- Xie, S.; Pancost, R.D.; Yin, H.; Wang, H.; Yang, H.; Wignall, P.B.; Luo, G.; Jia, C.; Chen, L. Cyanobacterial blooms tied to volcanism during the 5 m.y. Permo-Triassic biotic crisis. Geology 2010, 38, 447–450. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Depth (m) | TOC (%) | Sr/Ba | B/Ga | CVALUE | CIA | MoEF | UEF | P (%) | P/Ti | Ba/Al |
|---|---|---|---|---|---|---|---|---|---|---|---|
| J-26 | 3499.5 | 5.77 | 0.99 | 16.33 | 1.14 | 74 | 11.67 | 2.56 | 0.06 | 0.26 | 7.02 × 10−3 |
| J-25 | 3506.8 | 1.81 | 1.40 | 25.86 | 0.28 | 79 | 0.51 | 0.56 | 0.10 | 0.46 | 3.06 × 10−3 |
| J-24 | 3515.0 | 7.76 | 0.76 | 26.89 | 0.34 | 68 | 6.95 | 1.95 | 0.17 | 0.54 | 6.78 × 10−3 |
| J-23 | 3523.7 | 6.81 | 0.62 | 4.93 | 0.85 | 59 | 23.13 | 3.46 | 0.04 | 0.17 | 1.68 × 10−2 |
| J-22 | 3526.1 | 5.74 | 0.49 | 4.35 | 0.94 | 62 | 15.35 | 8.51 | 0.12 | 0.34 | 2.60 × 10−2 |
| J-21 | 3530.8 | 14.93 | 0.85 | 6.41 | 1.00 | 64 | 29.54 | 8.22 | 0.13 | 0.63 | 1.33 × 10−2 |
| J-20 | 3547.8 | 5.53 | 0.26 | 6.40 | 0.96 | 59 | 10.28 | 3.98 | 0.04 | 0.11 | 2.02 × 10−2 |
| J-19 | 3569.2 | 3.10 | 0.48 | 9.92 | 0.38 | 44 | 14.83 | 2.44 | 0.01 | 0.05 | 1.91 × 10−2 |
| J-18 | 3570.5 | 6.75 | 0.60 | 6.68 | 0.40 | 48 | 11.91 | 2.72 | 0.01 | 0.09 | 3.15 × 10−2 |
| J-17 | 3574.2 | 19.60 | 0.71 | 7.93 | 0.22 | 53 | 6.25 | 2.41 | 0.20 | 1.05 | 6.79 × 10−3 |
| J-16 | 3576.6 | 16.97 | 1.04 | 8.70 | 0.66 | 46 | 19.44 | 6.17 | 0.04 | 0.20 | 2.17 × 10−2 |
| J-15 | 3595.4 | 15.80 | 2.16 | 13.42 | 0.11 | 50 | 2.13 | 4.42 | 0.06 | 0.42 | 4.36 × 10−3 |
| J-14 | 3603.1 | 7.04 | 2.11 | 8.93 | 0.14 | 54 | 3.57 | 2.81 | 0.09 | 0.82 | 8.60 × 10−3 |
| J-13 | 3613.7 | 6.98 | 0.63 | 10.24 | 0.15 | 48 | 6.08 | 0.79 | 0.05 | 0.16 | 4.84 × 10−3 |
| J-12 | 3624.3 | 3.62 | 2.10 | 15.14 | 0.11 | 46 | 9.49 | 2.11 | 0.03 | 0.19 | 5.59 × 10−3 |
| J-11 | 3629.2 | 2.29 | 1.15 | 14.50 | 0.07 | 48 | 3.23 | 0.85 | 0.04 | 0.21 | 7.32 × 10−3 |
| J-10 | 3664.3 | 4.20 | 1.73 | 11.37 | 0.12 | 50 | 1.28 | 1.66 | 0.05 | 0.30 | 6.21 × 10−3 |
| J-09 | 3679.5 | 3.11 | 1.57 | 18.89 | 0.09 | 49 | 1.41 | 0.93 | 0.03 | 0.18 | 4.82 × 10−3 |
| J-08 | 3681.6 | 2.44 | 2.24 | 10.35 | 0.19 | 48 | 3.97 | 1.31 | 0.03 | 0.13 | 4.66 × 10−3 |
| J-07 | 3684.5 | 3.48 | 2.84 | 13.48 | 0.11 | 47 | 1.72 | 0.74 | 0.04 | 0.25 | 5.93 × 10−3 |
| J-06 | 3685.5 | 5.82 | 1.26 | 8.72 | 0.20 | 52 | 12.32 | 1.18 | 0.07 | 0.35 | 7.76 × 10−3 |
| J-05 | 3695.3 | 1.95 | 1.81 | 11.65 | 0.14 | 48 | 15.70 | 3.98 | 0.07 | 0.28 | 9.83 × 10−3 |
| J-04 | 3696.9 | 2.50 | 2.28 | 19.72 | 0.11 | 50 | 2.90 | 1.49 | 0.02 | 0.09 | 6.09 × 10−3 |
| J-03 | 3699.9 | 5.73 | 1.30 | 12.86 | 0.21 | 51 | 4.64 | 0.80 | 0.03 | 0.14 | 7.56 × 10−3 |
| J-02 | 3701.3 | 7.87 | 2.54 | 9.72 | 0.08 | 49 | 63.49 | 2.42 | 0.05 | 0.37 | 9.71 × 10−3 |
| J-01 | 3708.3 | 4.85 | 4.38 | 19.85 | 0.08 | 51 | 4.04 | 0.77 | 0.04 | 0.35 | 4.72 × 10−3 |
| Sample | Depth (m) | Max-Peak | CPI | OEP | TAR | Pr/n-C17 | Ph/n-C18 | Pr/Ph | GI | 2-MHI | β-Carotane Index | 7,8-MMH/Cmax | Regular Steranes (%) | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| C27 | C28 | C29 | |||||||||||||
| J-26 | 3499.5 | C23 | 1.08 | 1.40 | 0.91 | 2.14 | 1.51 | 1.30 | 0.10 | 2.00 | 18.74 | 0.04 | 22.4 | 24.0 | 53.7 |
| J-25 | 3506.8 | C23 | 1.09 | 1.34 | 1.26 | 1.30 | 1.57 | 0.74 | 0.14 | 3.12 | 34.69 | 0.03 | 42.8 | 41.6 | 15.6 |
| J-24 | 3515.0 | C23 | 1.06 | 1.23 | 0.91 | 1.20 | 1.09 | 0.75 | 0.20 | 1.41 | 14.38 | 0.04 | 27.7 | 28.6 | 43.7 |
| J-23 | 3523.7 | C17 | 1.08 | 1.29 | 0.09 | 0.86 | 0.76 | 1.25 | 0.21 | 1.35 | 3.46 | 0.12 | 33.3 | 26.3 | 40.4 |
| J-21 | 3530.8 | C19 | 1.07 | 1.01 | 0.31 | 0.60 | 0.51 | 1.01 | 0.16 | 1.88 | 2.86 | 0.07 | 25.3 | 35.0 | 39.7 |
| J-20 | 3547.8 | C25 | 1.05 | 1.06 | 0.62 | 10.59 | 3.67 | 0.47 | 0.20 | 3.64 | 5.56 | 0.00 | 19.6 | 36.7 | 43.7 |
| J-19 | 3569.2 | C23 | 1.05 | 1.18 | 0.92 | 0.91 | 0.67 | 0.77 | 0.24 | 4.20 | 8.03 | 0.04 | 11.9 | 39.5 | 48.5 |
| J-17 | 3574.2 | C17 | 1.09 | 1.24 | 0.11 | 0.87 | 0.81 | 1.33 | 0.21 | 4.65 | 4.69 | 0.11 | 23.4 | 40.2 | 36.5 |
| J-15 | 3595.4 | C19 | 1.09 | 1.14 | 0.20 | 0.46 | 0.30 | 1.50 | 0.19 | 4.69 | 3.69 | 0.04 | 18.0 | 41.2 | 40.8 |
| J-14 | 3603.1 | C21 | 1.08 | 1.07 | 0.58 | 4.37 | 6.15 | 0.52 | 0.24 | 2.84 | 6.65 | 0.01 | 27.1 | 20.8 | 52.1 |
| J-13 | 3613.7 | C17 | 1.07 | 1.35 | 0.13 | 0.95 | 0.87 | 1.31 | 0.18 | 4.05 | 7.34 | 0.11 | 17.1 | 38.4 | 44.6 |
| J-12 | 3624.3 | C21 | 1.07 | 1.05 | 0.36 | 1.25 | 1.02 | 1.14 | 0.25 | 4.01 | 6.01 | 0.12 | 11.0 | 36.2 | 52.8 |
| J-11 | 3664.3 | C23 | 1.05 | 1.17 | 0.80 | 2.93 | 3.64 | 0.84 | 0.25 | 4.18 | 31.42 | 0.24 | 11.4 | 34.7 | 53.9 |
| J-10 | 3629.2 | C17 | 1.06 | 1.38 | 0.29 | 1.28 | 1.66 | 0.82 | 0.23 | 4.15 | 6.77 | 0.17 | 10.6 | 40.2 | 49.3 |
| J-09 | 3679.5 | C23 | 1.05 | 1.14 | 0.68 | 1.70 | 1.95 | 0.88 | 0.24 | 3.33 | 26.04 | 0.15 | 8.3 | 36.1 | 55.6 |
| J-08 | 3681.6 | C23 | 1.05 | 1.13 | 1.21 | 1.69 | 1.57 | 0.65 | 0.22 | 3.93 | 66.47 | 0.05 | 8.5 | 36.0 | 55.5 |
| J-07 | 3684.5 | C20 | 1.03 | 1.10 | 0.79 | 1.58 | 1.19 | 0.20 | 0.25 | 3.64 | 17.15 | 0.03 | 9.9 | 29.3 | 60.7 |
| J-06 | 3685.5 | C22 | 1.05 | 1.08 | 0.92 | 0.97 | 1.20 | 0.42 | 0.26 | 3.48 | 13.25 | 0.05 | 11.7 | 26.7 | 61.6 |
| J-04 | 3696.9 | C23 | 1.06 | 1.11 | 2.16 | 1.79 | 1.53 | 0.43 | 0.20 | 4.37 | 28.02 | 0.04 | 9.2 | 35.4 | 55.5 |
| J-03 | 3699.9 | C23 | 1.06 | 1.11 | 1.52 | 1.10 | 1.13 | 0.49 | 0.23 | 2.91 | 15.66 | 0.04 | 9.2 | 35.2 | 55.6 |
| J-02 | 3701.3 | C17 | 1.10 | 1.43 | 0.13 | 0.94 | 0.95 | 0.98 | 0.22 | 5.26 | 1.19 | 0.13 | 13.3 | 36.5 | 50.2 |
| J-01 | 3708.3 | C23 | 1.05 | 1.12 | 1.70 | 1.23 | 1.35 | 0.19 | 0.22 | 4.21 | 28.30 | 0.03 | 10.5 | 32.9 | 56.6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, W.; Cui, H.; Tan, J.; Liu, J.; Song, X.; Wang, J.; Chen, L. Permian Cyanobacterial Blooms Resulted in Enrichment of Organic Matter in the Lucaogou Formation in the Junggar Basin, NW China. Minerals 2023, 13, 537. https://doi.org/10.3390/min13040537
Wang W, Cui H, Tan J, Liu J, Song X, Wang J, Chen L. Permian Cyanobacterial Blooms Resulted in Enrichment of Organic Matter in the Lucaogou Formation in the Junggar Basin, NW China. Minerals. 2023; 13(4):537. https://doi.org/10.3390/min13040537
Chicago/Turabian StyleWang, Wenhui, Haisu Cui, Jingqiang Tan, Jin Liu, Xueqi Song, Jian Wang, and Lichang Chen. 2023. "Permian Cyanobacterial Blooms Resulted in Enrichment of Organic Matter in the Lucaogou Formation in the Junggar Basin, NW China" Minerals 13, no. 4: 537. https://doi.org/10.3390/min13040537