
Microbial Mat Stratification in Travertine Depositions of Greek Hot Springs and Biomineralization Processes
 ,
,  ,
,  , ,
, ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Microbial Identification
2.2. Mineralogical Study
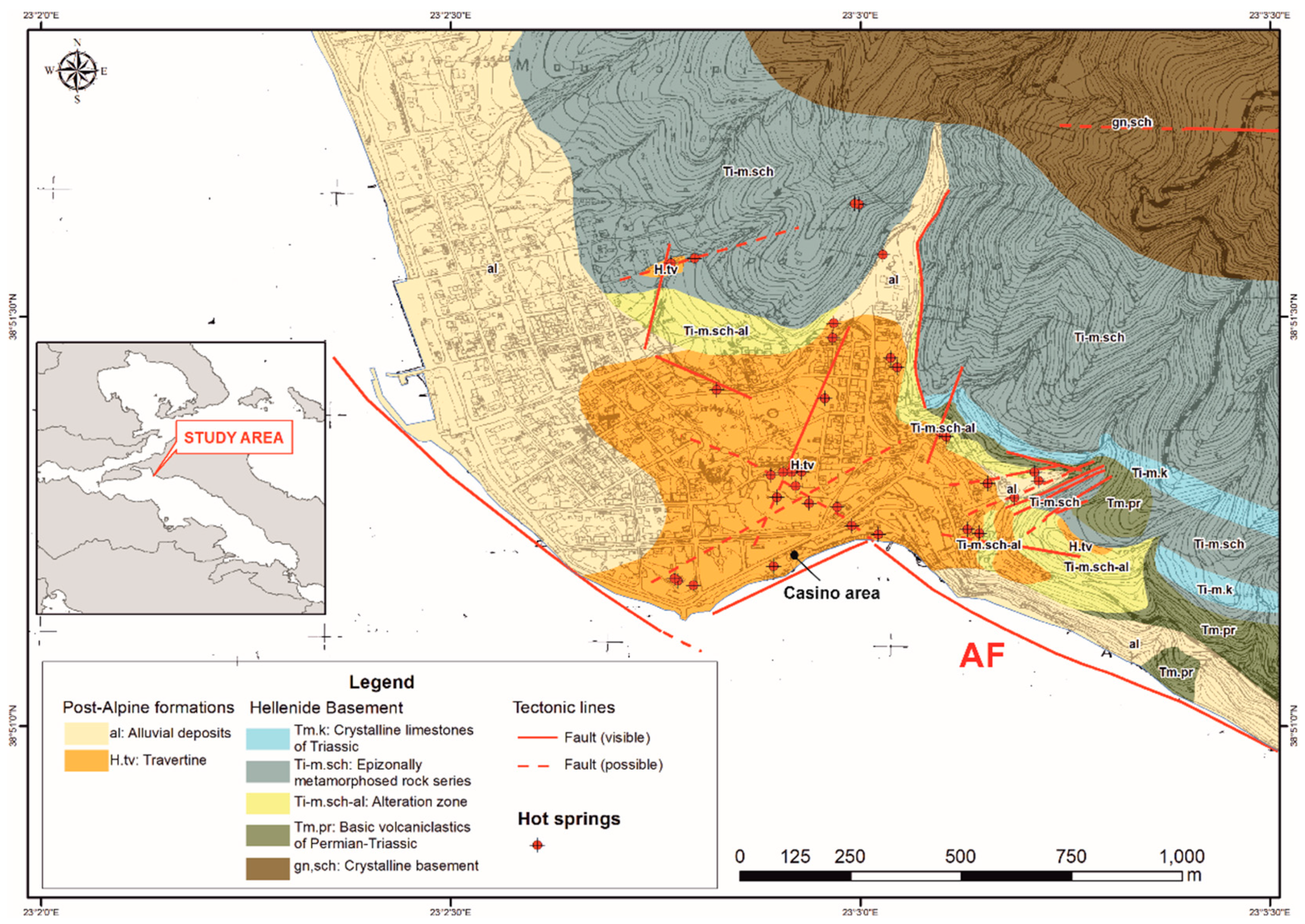
3. Geological Setting
4. Results and Discussion
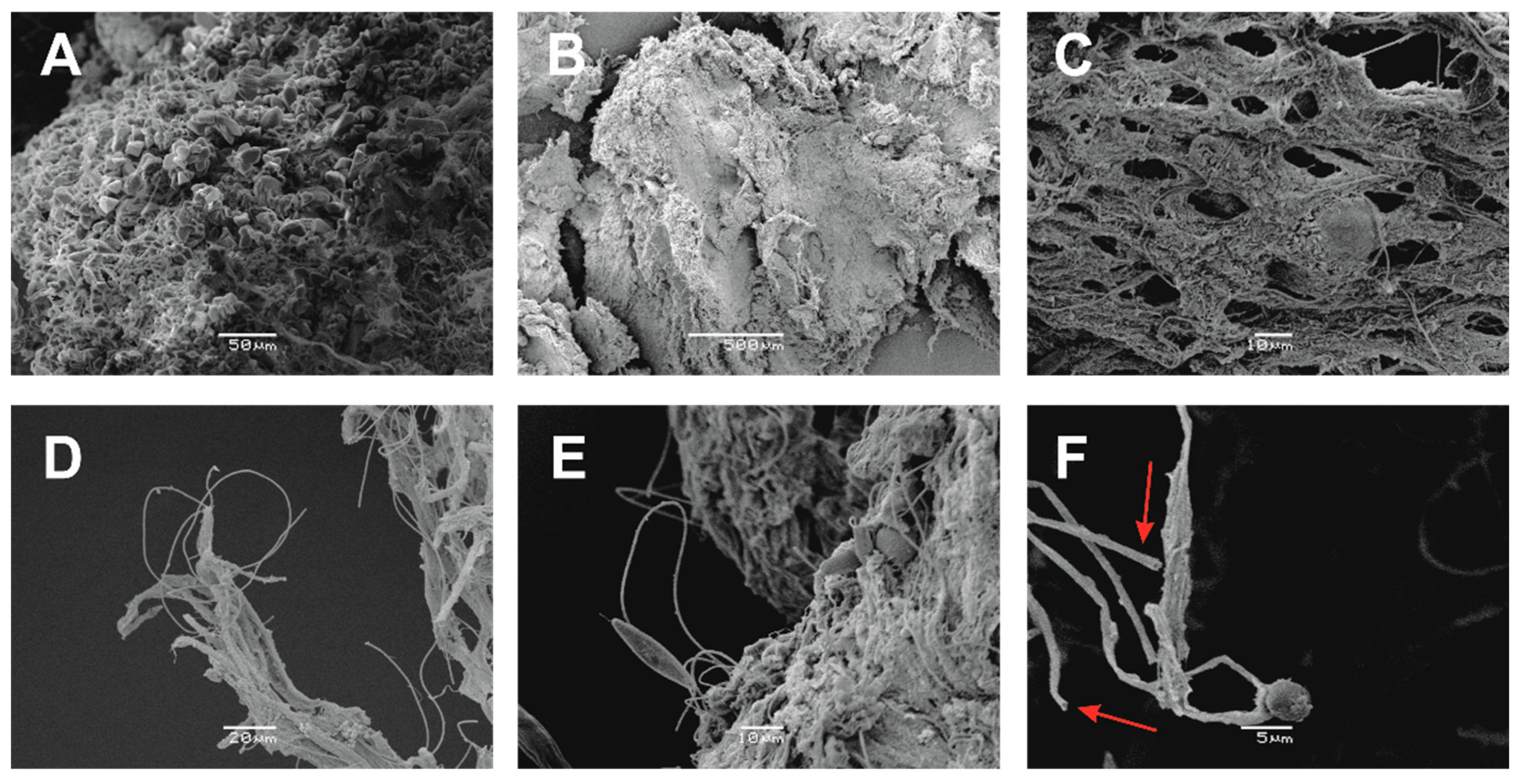
4.1. Sample Description and In-Situ Analysis
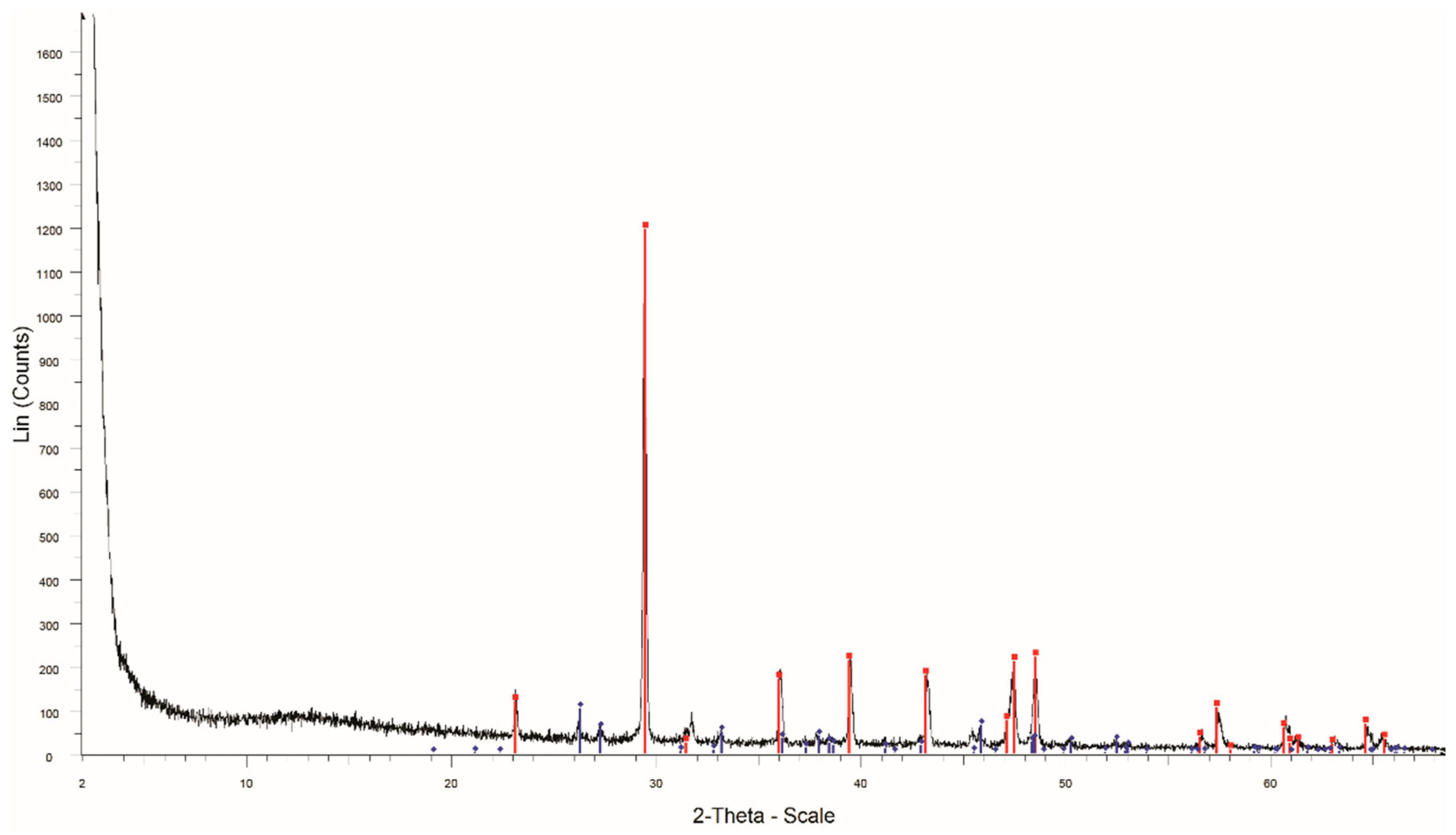
4.2. Mineralogical Characterization and Facies
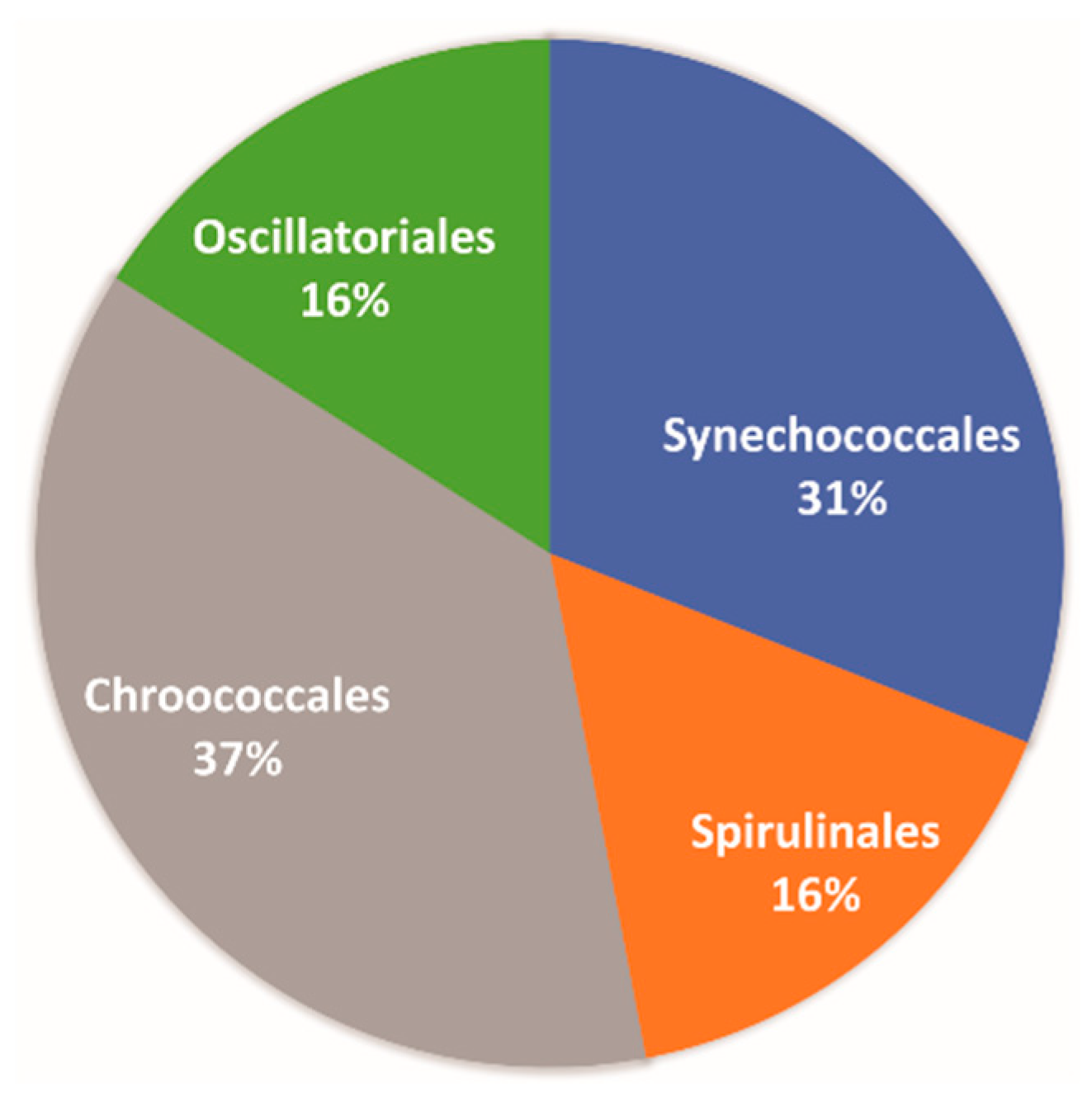
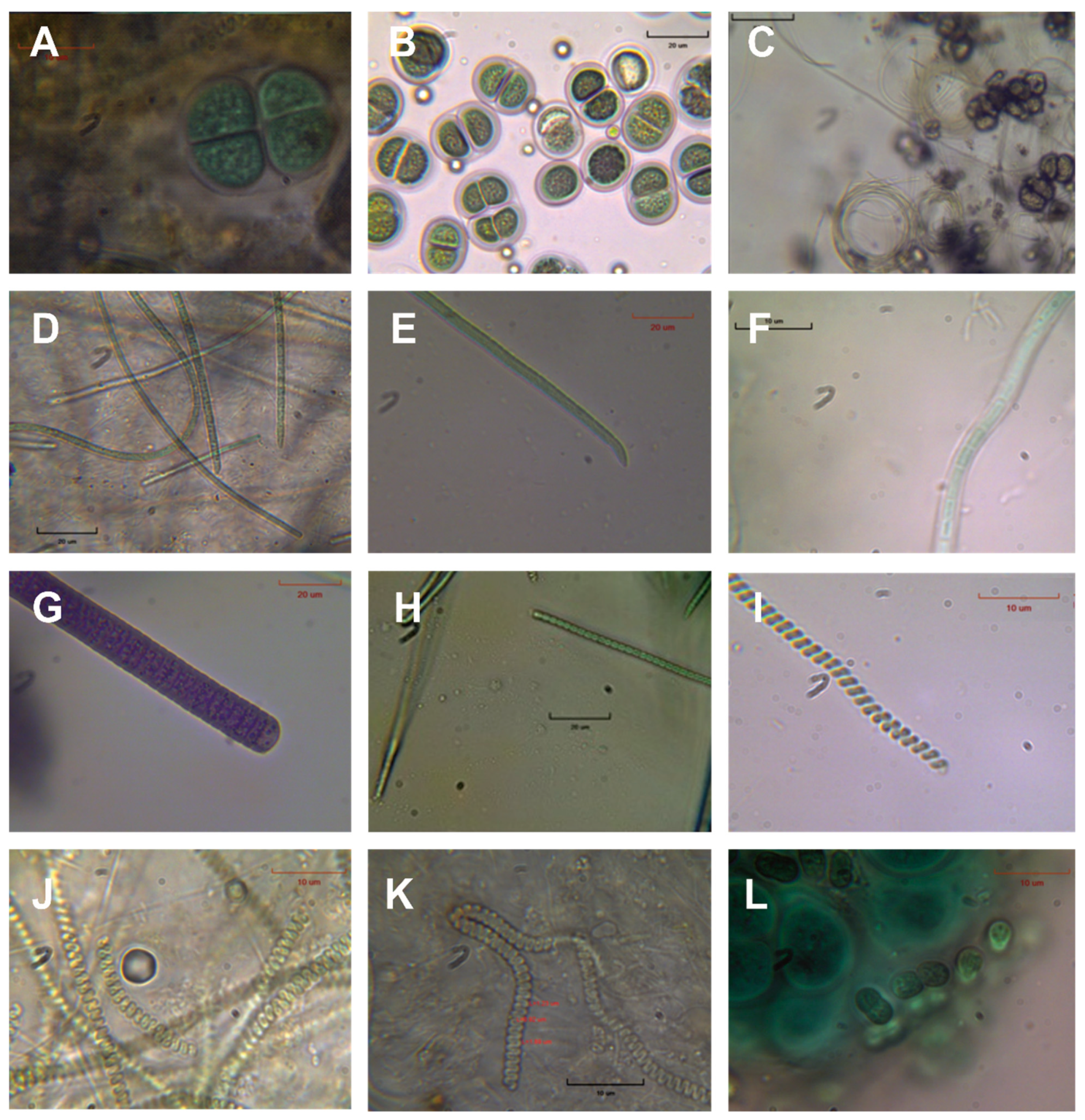
4.3. Cyanobacteria Diversity and Possible Applications
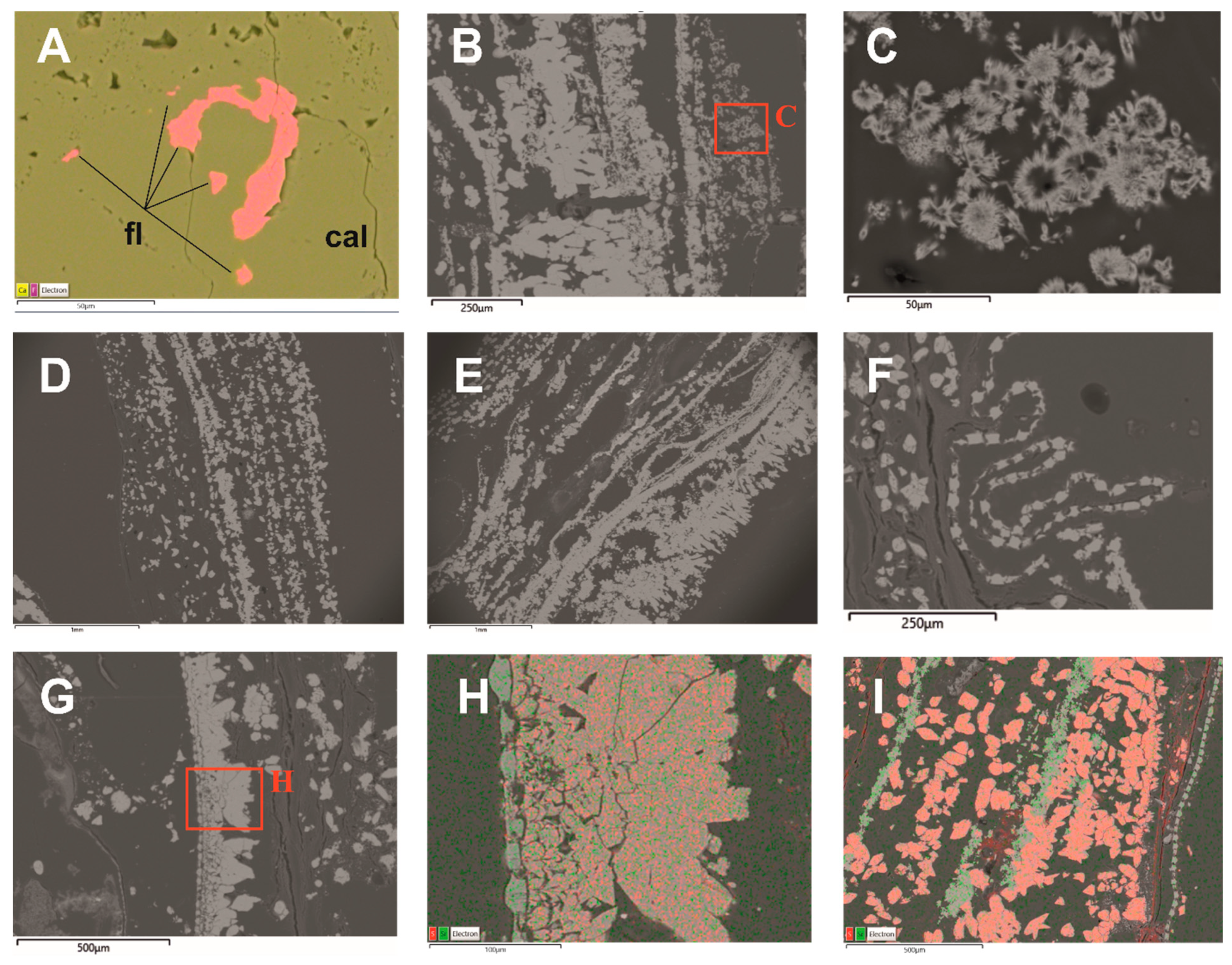
4.4. Biomineralisation
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reitner, J.; Thiel, V. Encyclopedia of Geobiology; Springer Science and Business Media: Berlin/Heidelberg, Germany, 2011; ISBN 978-1-4020-9211-4. [Google Scholar]
- Allwood, A.C.; Walter, M.R.; Kamber, B.S.; Marshal, C.P.; Burch, I.W. Stromatolite reef from the Early Archaean era of Australia. Nature 2006, 441, 714–717. [Google Scholar] [CrossRef]
- Van Kranendonk, M.J. A review of the evidence for putative Paleoarchean life in the Pilbara Craton. In Earth’s Oldest Rocks—Developments in Precambrian Geology; Van Kranendonk, M.J., Smithies, R.H., Bennet, V., Eds.; Elsevier: Amsterdam, The Netherlands, 2007; pp. 855–896. [Google Scholar]
- Homann, M. Earliest life on earth: Evidence from the Barberton Greenstone Belt, South Africa. Earth-Sci. Rev. 2019, 196, 102888. [Google Scholar] [CrossRef]
- Westall, F.; de Vries, S.T.; Nijman, W.; Rouchon, V.; Orberger, B.; Pearson, V.; Watson, J.; Verchovski, A.; Wright, I.; Rouzaud, J.N.; et al. The 3.466 Ga “Kitty’s gap Chert” an early Archean microbial ecosystem. In Processes on the Early Earth; Reimod, W.U., Gibson, R.L., Eds.; Geological Society of America: Boulder, CO, USA, 2006; Volume 405, pp. 105–131. [Google Scholar] [CrossRef]
- Van Dover, C.L. The Ecology of Deep-Sea Hydrothermal Vents; Princeton University Press: Princeton, NJ, USA, 2000; 424p. [Google Scholar]
- Riding, R.; Awramik, S.M. (Eds.) Microbial Sediments; Springer: Berlin/Heidelberg, Germany, 2000; 331p. [Google Scholar]
- Arp, G.; Reimer, A.; Reitner, J. Microbialite formation in seawater of increased alkalinity, Satonda Crater Lake, Indonesia. J. Sediment. Res. 2003, 73, 105–127. [Google Scholar] [CrossRef]
- Pedersen, K. Microbial life in deep granitic rock. FEMS Microbiol. Rev. 2006, 20, 399–414. [Google Scholar] [CrossRef]
- Characklis, W.G.; Wilderer, P.A. Structure and Function of Biofilms; Wiley: Hoboken, NJ, USA, 1989; 387p. [Google Scholar]
- Fouke, B.W.; Bonheyo, G.T.; Sanzenbacher, B.; Frias-Lopez, J. Partitioning of bacterial communities between travertine depositional facies at Mammoth Hot Springs, Yellowstone National Park, USA. Can. J. Earth Sci. 2003, 40, 1531–1548. [Google Scholar] [CrossRef] [Green Version]
- Meyer-Dombard, D.R.; Shock, E.L.; Amend, J.P. Archaeal and bacterial communities in geochemically diverse hot springs of Yellowstone National Park, USA. Geobiology 2005, 3, 211–227. [Google Scholar] [CrossRef]
- Fouke, B.W. Hot-spring systems geobiology: Abiotic and biotic influences on travertine formation at mammoth hot springs, Yellowstone National Park, USA. Sedimentology 2011, 58, 170–199. [Google Scholar] [CrossRef]
- Thomas, C. Investigating the Subsurface Biosphere of a Hypersaline Environment—The Dead Sea (Levant). Ph.D. Thesis, Université de Genève, Geneva, Switzerland, 2015. [Google Scholar]
- Thomas, C.; Ariztegui, D. Fluid inclusions from the deep Dead Sea sediment provide new insights on Holocene extreme microbial life. Quat. Sci. Rev. 2019, 212, 18–27. [Google Scholar] [CrossRef] [Green Version]
- Merino, N.; Aronson, H.S.; Bojanova, D.P.; Feyhl-Buska, J.; Wong, M.L.; Zhang, S.; Giovannelli, D. Living at the Extremes: Extremophiles and the Limits of Life in a Planetary Context. Front. Microbiol. 2019, 10, 780. [Google Scholar] [CrossRef] [PubMed]
- Bech, J. Les Terres Medicinales. Discurs per Reial Academia de Farmàcia de Barcelona; Reial Acadèmia de Farmàcia de Barcelona-CIRIT (Generalitat de Calalunya): Barcelona, Spain, 1987; p. 105. [Google Scholar]
- Canfield, D.E.; Des Marais, D.J. Biogeochemical cycles of carbon, sulfur, and free oxygen in a microbial mat. GCA 1993, 57, 3971–3984. [Google Scholar] [CrossRef]
- Allison, D.G.; Gilbert, P.; Lappin-Scott, H.M.; Wilson, M. Community Structure and Co-Operation in Biofilms; Cambridge University Press: Cambridge, UK, 2000; 349p. [Google Scholar]
- Wingender, J.; Neu, T.R.; Flemming, H.-C. Microbial Extracellular Polymeric Substances; Characterization, Structure, and Function; Springer: Berlin/Heidelberg, Germany, 1999; 258p. [Google Scholar]
- Decho, A.W.; Visscher, P.T.; Reid, P. Production and cycling of natural microbial exopolymers (EPS) within a marine stromatolite. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2005, 219, 71–86. [Google Scholar] [CrossRef]
- Mourelle, M.L.; Gómez, C.P.; Legido, J.L. Microalgal Peloids for Cosmetic and Wellness Uses. Mar. Drugs 2021, 19, 666. [Google Scholar] [CrossRef] [PubMed]
- Gomes, C.; Carretero, M.I.; Pozo, M.; Maraver, F.; Cantista, P.; Armijo, F.; Legido, J.L.; Teixeira, F.; Rautureau, M.; Delgado, R. Peloids and pelotherapy: Historical evolution, classification and glossary. Appl. Clay Sci. 2013, 75–76, 28–38. [Google Scholar] [CrossRef]
- Yilmaz, B.; Goktepe, A.S.; Alaca, R.; Mohur, H.; Kayar, A.H. Comparison of a generic and a disease specific quality of life scale to assess a comprehensive spa therapy program for knee osteoarthritis. Jt. Bone Spine 2004, 71, 563–566. [Google Scholar] [CrossRef]
- Abedin, R.M.A.; Taha, H.M. Antibacterial and antifungal activity of cyanobacteria and green microalgae. Evaluation of medium components by Plackett-Burman design for antimicrobial activity of Spirulina platensis. GJBBR 2008, 3, 22–31. [Google Scholar]
- Gademann, K.; Portmann, C. Secondary metabolites from cyanobacteria: Complex structures and powerful bioactivities. Curr. Org. Chem. 2008, 12, 326–341. [Google Scholar] [CrossRef]
- Martins, R.F.; Ramos, M.F.; Herfindal, L.; Sousa, J.A.; Skærven, K.; Vasconcelos, V.T. Antimicrobial and cytotoxic assessment of marine cyanobacteria Synechocystis & Synechococcus. Mar. Drugs 2008, 6, 1–11. [Google Scholar] [PubMed]
- Kanellopoulos, C.; Mitropoulos, P.; Valsami-Jones, E.; Voudouris, P. A new terrestrial active mineralizing hydrothermal system associated with ore-bearing travertines in Greece (northern Euboea Island and Sperchios area). J. Geochem. Explor. 2017, 179, 9–24. [Google Scholar] [CrossRef]
- Kanellopoulos, C. Geochemical Research on the Distribution of Metallic and Other Elements in the Cold and Thermal Groundwater, Soils and Plants in Fthiotida Prefecture and N. Euboea. Environmental Impact. Ph.D. Thesis, National and Kapodistrian University of Athens, Athens, Greece, 2011. (In Greek with English abstract). [Google Scholar]
- Kanellopoulos, C. Distribution, lithotypes and mineralogical study of newly formed thermogenic travertines in Northern Euboea and Eastern Central Greece. Open Geosci. (former Cent. Eur. Geol.) 2012, 4, 545–560. [Google Scholar] [CrossRef]
- Kanellopoulos, C. Various morphological types of thermogenic travertines in northern Euboea and Eastern Central Greece. Bull. Geol. Soc. Greece 2013, 47, 1929–1938. [Google Scholar] [CrossRef]
- Kanellopoulos, C.; Lamprinou, V.; Politi, A.; Voudouris, P.; Economou-Amilli, A. Insights on the biomineralization processes and related diversity of cyanobacterial microflora in thermogenic travertine deposits in Greek hot springs (North-West Euboea Island). Depos. Rec. 2022, 8, 1055–1078. [Google Scholar] [CrossRef]
- Kanellopoulos, C.; Lamprinou, V.; Politi, A.; Voudouris, P.; Economou-Amilli, A. Pioneer species of Cyanobacteria in hot springs and their role to travertine formation: The case of Aedipsos hot springs, Euboea (Evia), Greece. Depos. Rec. 2022, 8, 1079–1092. [Google Scholar] [CrossRef]
- Kanellopoulos, C.; Thomas, C.; Xirokostas, N.; Ariztegui, D. Banded Iron Travertines at the Ilia Hot Spring (Greece): An interplay of biotic and abiotic factors leading to a modern BIF analog? Depos. Rec. 2019, 5, 109–130. [Google Scholar] [CrossRef]
- Stanier, R.Y.; Kunisawa, R.; Mandel, M.; Cohen-Bazire, G. Purification and properties of unicellular blue-green algae (order Chroococcales). Bacteriol. Rev. 1971, 35, 171. [Google Scholar] [CrossRef]
- Komárek, J.; Anagnostidis, K. Modern approach to the classification system of Cyanophytes 4-Nostocales. Arch. Hydrobiol. Suppl. Bd. Algol. Stud. 1989, 56, 247–345. [Google Scholar]
- Komárek, J.; Anagnostidis, K. Cyanoprokaryota, Part 1: Chroococcales. In Süßwasserflora von Mitteleuropa, Bd. 19 (1); Büdel, B., Gärtner, G., Krienitz, L., Schagerl, M., Eds.; Elsevier GmbH: München, Germany, 1999; pp. 1–548. [Google Scholar]
- Komárek, J.; Anagnostidis, K. Cyanoprokaryota, Part 2: Oscillatoriales. In Süßwasserflora von Mitteleuropa, Bd. 19 (2); Büdel, B., Gärtner, G., Krienitz, L., Schagerl, M., Eds.; Elsevier GmbH: München, Germany, 2005; pp. 1–759. [Google Scholar]
- Komárek, J.; Kastovsky, J.; Mares, J.; Johansen, J.R. Taxonomic classification of cyanoprokaryotes (cyanobacterial genera), using a polyphasic approach. Preslia 2014, 86, 295–335. [Google Scholar]
- Kanellopoulos, C.; Xenakis, M.; Vakalopoulos, P.; Kranis, H.; Christopoulou, M.; Vougioukalakis, G. Seawater-dominated, tectonically controlled and volcanic related geothermal systems: The case of the geothermal area in the northwest of the Island of Euboea (Evia), Greece. Int. J. Earth Sci. 2020, 109, 2081–2112. [Google Scholar] [CrossRef]
- Aubouin, J. Contribution a l’ étude géologique de la Grèce septentrionale: Les confins de l’Epire et de la Thessalie. Ann. Geol. Pays Hellen. 1959, 10, 1–483. [Google Scholar]
- Mountrakis, D. The Pelagonian zone in Greece: A polyphase-deformed fragment of the Cimmerian continent and its role in the geotectonic evolution of the eastern Mediterranean. J. Geol. 1986, 94, 335–347. [Google Scholar] [CrossRef]
- Vavassis, I. Geology of the Pelagonian Zone in Northern Evia Island (Greece): Implications for the Geodynamic Evolution of the Hellenides. Ph.D. Thesis, Universite de Lausanne, Lausanne, Switzerland, 2001. [Google Scholar]
- Jolivet, L.; Faccenna, C.; Huet, B.; Labrousse, L.; Le Pourhiet, L.; Lacombe, O.; Lecomte, E.; Burov, E.; Denèle, Y.; Brun, J.-P.; et al. Aegean tectonics: Strain localization, slab tearing and trench retreat. Tectonophysics 2013, 597–598, 1–33. [Google Scholar] [CrossRef] [Green Version]
- Katsikatsos, G.; Mettos, A.; Vidakis, M.; Bavay, P.; Panagopoulos, A.; Basilaki, A.; Papazeti, E. Geological study of Aedipos Area—Euboea; Geothemal Studies (P.E.C.); IGME: Athens, Greece, 1982. (In Greek) [Google Scholar]
- Scherreiks, R. Platform margin and oceanic sedimentation in a divergent and convergent plate setting (Jurassic, Pelagonian Zone, NE Evvoia, Greece). Int. J. Earth Sci. 2000, 89, 90–107. [Google Scholar] [CrossRef]
- Kanellopoulos, C.; Christopoulou, M.; Xenakis, M.; Vakalopoulos, P. Hydrochemical characteristics and geothermometry applications of hot groundwater in Edipsos area, NW Euboea (Evia), Greece. Bull. Geol. Soc. Greece 2016, 50, 720–729. [Google Scholar] [CrossRef] [Green Version]
- Fytikas, M.; Giuliani, O.; Innocenti, F.; Marinelli, G.; Mazzuoli, R. Geochronological data on recent magmatism of the Aegean Sea. Tectonophysics 1976, 31, T29–T34. [Google Scholar] [CrossRef]
- De Boever, E.; Brasier, A.T.; Foubert, A.; Kele, S. What do we really know about early diagenesis of non-marine carbonates? Sediment. Geol. 2017, 361, 25–51. [Google Scholar] [CrossRef] [Green Version]
- Jorgensen, B.B.; Revsbech, N.P.; Cohen, Y. Photosynthesis and structure of benthic microbial mats: Microelectrode and SEM studies of four cyanobacterial communities. Limnol. Oceanogr. 1983, 28, 1075–1093. [Google Scholar] [CrossRef]
- Stal, L.J. Cyanobacterial mats and stromatolites. In The Ecology of Cyanobacteria; Whitton, B.A., Potts, M., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2000; pp. 61–120. [Google Scholar]
- Pentecost, A. Travertine; Springer: Berlin/Heidelberg, Germany, 2005. [Google Scholar]
- Ward, D.M.; Ferris, M.J.; Nold, S.C.; Bateson, M.M.; Kopczynski, E.D.; Ruff-Roberts, A.L. Species diversity in hot spring microbial mats as revealed by both molecular and enrichment culture approaches—Relationship between biodiversity and community structure. In Microbial Mats; Stal, J.L., Caumette, P., Eds.; Springer: Berlin/Heidelberg, Germany, 1994; pp. 33–44. [Google Scholar]
- Takashima, C.; Kano, A. Microbial processes forming daily lamination in a stromatolitic travertine. Sediment. Geol. 2008, 208, 114–119. [Google Scholar] [CrossRef]
- Mucci, A.; Mucci, J.W. The incorporation of Mg2+ and Sr2+ into calcite overgrowths: Influences of growth rate and solution composition. Geochim. Cosmochim. Acta 1983, 47, 217–233. [Google Scholar] [CrossRef]
- Ishikawa, M.; Ichikuni, M. Uptake of sodium and potassium by calcite. Chem. Geol. 1984, 42, 137–146. [Google Scholar] [CrossRef]
- Long, X.; Ma, Y.; Qi, L. Biogenic and synthetic high magnesium calcite—A review. J. Struct. Biol. 2014, 185, 1–14. [Google Scholar] [CrossRef]
- Yoshimura, T.; Tamenori, Y.; Suzuki, A.; Kawahata, H.; Iwasaki, N.; Hasegawa, H.; Nguyen, L.T.; Kuroyanagi, A.; Yamazaki, T.; Kuroda, J.; et al. Altervalent substitution of sodium for calcium in biogenic calcite and aragonite. Geochim. Cosmochim. Acta 2017, 202, 21–38. [Google Scholar] [CrossRef]
- Okumura, T.; Kim, H.-J.; Kim, J.-W.; Kogure, T. Sulfate-containing calcite: Crystallographic characterization of natural and synthetic materials. Eur. J. Mineral. 2018, 30, 929–937. [Google Scholar] [CrossRef]
- Kampschulte, A.; Strauss, H. The sulfur isotopic evolution of Phanerozoic seawater based on the analysis of structurally substituted sulfate in carbonates. Chem. Geol. 2004, 204, 255–286. [Google Scholar] [CrossRef]
- Balan, E.; Aufort, J.; Pouillé, S.; Dabos, M.; Blanchard, M.; Lazzeri, M.; Rollion-Bard, C.; Blamart, D. Infrared spectroscopic study of sulfate-bearing calcite from deep-sea bamboo coral. Eur. J. Mineral. Copernic. 2017, 29, 397–408. [Google Scholar] [CrossRef]
- Reitner, J. Modern cryptic microbialite/metazoan facies from Lizard Island (Great Barrier Reef, Australia): Formation and Concept. Facies 1993, 29, 3–40. [Google Scholar] [CrossRef] [Green Version]
- Görgen, S.; Benzerara, K.; Skouri-Panet, F.; Gugger, M.; Chauvat, F.; Cassier-Chauvat, C. The diversity of molecular mechanisms of carbonate biomineralization by bacteria. Discov. Mater. 2021, 1, 2. [Google Scholar] [CrossRef]
- Della Porta, G.; Hoppert, M.; Hallmann, C.; Schneider, D.; Reitner, J. The influence of microbial mats on travertine precipitation in active hydrothermal systems (Central Italy). Depos. Rec. 2021, 8, 165–209. [Google Scholar] [CrossRef]
- Della Porta, G. Carbonate build-ups in lacustrine, hydrothermal and fluvial settings: Comparing depositional geometry, fabric types and geochemical signature. Geol. Soc. Lond. Spec. Publ. 2015, 418, 17–68. [Google Scholar] [CrossRef]
- Hauer, T.; Komárek, J. CyanoDB 2.0—On-line Database of Cyanobacterial Genera; World-Wide Electronic Publication; University of South Bohemia & Institute of Botany AS CR: České Budějovice, Czechia, 2021; Available online: https://www.cyanodb.cz (accessed on 19 December 2021).
- Pesciaroli, C.; Viseras, C.; Aguzzi, C.; Rodelas, B.; Gonzalez-Lopez, J. Study of bacterial community structure and diversity during the maturation process of a therapeutic peloid. Appl. Clay Sci. 2016, 132, 59–67. [Google Scholar] [CrossRef]
- Vadlja, D.; Bujak, M.; Čož-Rakovac, R.; Roje, M.; Čižmek, L.; Horvatić, A.; Svetličić, E.; Diminić, J.; Rakovac, S.; Oros, D.; et al. Bioprospecting for Microorganisms in Peloids—Extreme Environment Known for Its Healing Properties. Front. Mar. Sci. 2022, 9, 822139. [Google Scholar] [CrossRef]
- Sompong, U.; Hawkins, P.R.; Besley, C.; Peerapornpisal, Y. The distribution of cyanobacteria across physical and chemical gradients in hot springs in Northern Thailand. FEMS 2005, 52, 365–376. [Google Scholar] [CrossRef] [Green Version]
- Flemming, H.-C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Riding, R. Cyanobacterial calcification, carbon dioxide concentrating mechanisms, and Proterozoic-Cambrian changes in atmospheric composition. Geobiology 2006, 4, 299–316. [Google Scholar] [CrossRef]
- Jansson, C.; Northen, T. Calcifying cyanobacteria—The potential of biomineralization for carbon capture and storage. Curr. Opin. Biotechnol. 2010, 21, 365–371. [Google Scholar] [CrossRef] [PubMed]
- Merz-Preiss, M.; Riding, R. Cyanobacterial tufa calcification in two freshwater streams: Ambient environment, chemical thresholds and biological processes. Sediment. Geol. 1999, 126, 103–124. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| I | II | III | IV | V | VI | VII | VIII | |
|---|---|---|---|---|---|---|---|---|
| Na2O | - | 0.16 | - | 0.41 | 0.23 | 0.19 | - | - |
| MgO | 0.56 | 0.59 | 0.11 | 0.87 | 0.52 | 0.45 | 0.7 | 0.96 |
| SO3 | - | - | - | 3.26 | 3.76 | 3.99 | - | - |
| K2O | - | - | - | - | - | - | - | 0.14 |
| CaO | 53.58 | 51.43 | 54.71 | 52.32 | 51.62 | 53.81 | 52.51 | 51.69 |
| SrO | - | - | - | - | - | - | 0.28 | 0.25 |
| Total | 54.13 | 52.37 | 54.82 | 56.86 | 56.13 | 58.44 | 53.49 | 53.64 |
| Aphanothece cf. minutissima (West) J.Komárková-Legnerová and G.Cronberg 1994 |
| Aphanothece sp.A |
| Chondrocystis dermochroa (Nägeli) Komárek and Anagnostidis, 1995 |
| Chroococcus minutus (Kützing) Nägeli 1849 |
| Chroococcus minutus var. thermalis J.J.Copeland |
| Chroococcus thermalis (Meneghini) Nägeli 1849 |
| Chroococcus turgidus (Kützing) Nägeli 1849 |
| Leptolyngbya cf. angusta (Skuja) Anagnostidis 2001 |
| Johannesbaptistia pellucida (Dickie) W.R.Taylor and Drouet in Drouet 1938 |
| Kamptonema chlorinum (Kützing ex Gomont) Strunecký, Komárek and J.Smarda 2014 |
| Kamptonemaformosum (Bory ex Gomont) Strunecký, Komárek and J.Smarda 2014 |
| Leptolyngbya perforans (Geitler) Anagnostidis and Komárek 1988 |
| Limnothrix redekei (Goor) Meffert 1988 |
| Oscillatoria crassa (C.B.Rao) Anagnostidis 2001 |
| Pseudanabaena minima (G.S.An) Anagnostidis 2001 |
| Spirulina cf. labyrinthiformis Gomont 1892 |
| Spirulina subtilissima Kützing ex Gomont 1892 |
| Spirulina tenuior (Lagerheim) Kirchner 1900 |
| Synechocystis salina Wislouch 1924 |
| Diatoms |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kanellopoulos, C.; Lamprinou, V.; Politi, A.; Voudouris, P.; Iliopoulos, I.; Kokkaliari, M.; Moforis, L.; Economou-Amilli, A. Microbial Mat Stratification in Travertine Depositions of Greek Hot Springs and Biomineralization Processes. Minerals 2022, 12, 1408. https://doi.org/10.3390/min12111408
Kanellopoulos C, Lamprinou V, Politi A, Voudouris P, Iliopoulos I, Kokkaliari M, Moforis L, Economou-Amilli A. Microbial Mat Stratification in Travertine Depositions of Greek Hot Springs and Biomineralization Processes. Minerals. 2022; 12(11):1408. https://doi.org/10.3390/min12111408
Chicago/Turabian StyleKanellopoulos, Christos, Vasiliki Lamprinou, Artemis Politi, Panagiotis Voudouris, Ioannis Iliopoulos, Maria Kokkaliari, Leonidas Moforis, and Athena Economou-Amilli. 2022. "Microbial Mat Stratification in Travertine Depositions of Greek Hot Springs and Biomineralization Processes" Minerals 12, no. 11: 1408. https://doi.org/10.3390/min12111408