
Fluctuating Asymmetry of Fallow Deer Antlers Is Associated with Weapon Damage but Not Tactical Behaviour during Fights

1
School of Biological Sciences, Queen’s University Belfast, Belfast BT9 7BL, UK
2
School of Science and Computing, Atlantic Technological University, H91 T8NW Galway, Ireland
*
Author to whom correspondence should be addressed.
Symmetry 2023, 15(4), 829; https://doi.org/10.3390/sym15040829
Submission received: 19 January 2023
/
Revised: 10 March 2023
/
Accepted: 21 March 2023
/
Published: 29 March 2023
(This article belongs to the Special Issue Fluctuating Asymmetry in Ungulates)
Abstract
:The horns and antlers borne by the males of many species of ungulate are considered to be both badges of quality, and armaments for use during intraspecific combat. Underpinning arguments concerning their dual utility is the idea that these structures should be costly to produce in order that the signal value of the structure is maintained. In agreement with such theorising is the belief that fluctuating asymmetry (FA), small deviations from symmetry around a mean of zero, measures individual quality as it represents the ability of the individual to withstand stress. We investigated whether the antlers of fallow deer indicated something of the quality of the bearer by assessing whether the degree of antler FA was associated with breakage (i.e., badge of quality) or with tactical investment in fighting (i.e., armament). We show the anticipated relationship between FA and antler damage, however, there was no relationship between FA and contest tactics. The present study, therefore, shows partial support for the idea that the magnitude of fluctuating asymmetry expressed by weaponry is related to individual quality.
1. Introduction
Selection argues that a variety of showy adornments have evolved, principally in males, that serve different functions: on the one hand, they serve as display ornaments to attract females, while on the other, these structures can act as weapons of display and attack against rivals during fights [1]. In the latter instance, the idea that weapons act as visual signals of individual quality, i.e., a badge of quality, whilst also serving as an effective means of physically overpowering opponents has been incorporated into models of contest behaviour (e.g., [2]). Such theorising argues that rivals actively assess the quality of potential opponents by estimating differences in quality through mutual displays [3], with the caveat that such signals are expensive to produce and maintain to ensure signal honesty [4,5]. There is some evidence that traits used in such displays are honest [1,6], and a number of studies have shown how the display and offensive features of weapons are associated with forms of opponent assessment during agonistic encounters (e.g., [7,8,9,10]).
However, the question as to what features of a trait, or traits, communicate the quality of an individual is not straightforward. There is evidence, at least in terms of weaponry, of a positive association between size and individual quality (e.g., [11,12,13,14]). Nevertheless, where weaponry is composed of bilateral structures there is the possibility that structural features other than size could be important, because the development of both sides is dependent on the same genome. Fluctuating asymmetry (FA) is one of several different forms of bilateral symmetry, and is characterised by a normal distribution of the size of left and right structures around a mean of zero and may represent such a feature [15]. In order to link FA to individual quality, it has been suggested that the ability of individuals to withstand environmental, or genetic stress, will be evident in the degree of symmetry expressed by the structures [16,17,18,19]. To evaluate this proposition with respect to ungulate weaponry, research has focused on the extent to which antlers and horns deviate from perfect symmetry; for example, it has been shown that antler symmetry is associated with parasite load [20], while increases in population density have been shown to increase antler asymmetry ([21], see Lens et al., 2002 [22] for a review). However, several studies have challenged this argument, showing that symmetry does not necessarily correlate with quality (e.g., [23,24,25,26]). Thus, with respect to the weapons borne by ungulates, the evidence relating symmetry to quality is inconsistent.
In common with most other members of the cervid family, male fallow deer cast and regrow their antlers annually; these structures are notable by their distinct form, usually consisting of two frontal tines on the antler beam with an upper palmate section that ends with a series of spellers ([27], see Figure 1). A key feature of antler production is the cost they exert on the individual during growth [28], with estimates that production requires from a 16% increase in energy requirements in caribou [29] up to 33% in the (extinct) Irish deer [30]. The production cost, in terms of mineral requirements, is such that dietary intake alone is not sufficient to enable growth, and additional nutrients need to be sequestered from the skeleton [31]. Consequently, the chemical composition of the antler is not uniform along its length, indicating that individuals can experience physiological exhaustion during the growth phase [32]. In behavioural terms, dominance status is associated with antler growth (e.g., [33]), and there is evidence that growth rate is correlated with changes in dominance rank during different stages of production [34]. Given that the degree of FA expressed by bilateral structures is linked to individual quality, it is possible that weaker individuals either reach their investment limit earlier in production, or they have a lower overall investment capacity than stronger individuals do. If this is the case, these differences could be manifested by differences in antler strength; therefore, we test the hypothesis that quality, as expressed by antler FA, is associated with probability that an individual will experience antler damage.
Fallow deer primarily use their antlers as weapons during intraspecific combat (e.g., [14,35,36,37,38]), and one consequence of repeatedly clashing antlers is the heightened potential of experiencing breakage [39,40]. A key feature of antler damage suggests that it determines whether individuals will aggressively engage each other [41,42]. Moreover, acquired damage also affects tactical behaviour during fights where it is associated with a reduction in attacking actions [43]. The question as to whether symmetry confers advantages during fights is unclear; for example, in the maritime earwig those individuals expressing greater directional asymmetry of the forceps are more likely to win fights [44]. Conversely, symmetry may provide a mechanical advantage during fights against asymmetrical opponents [45], while other studies show no evidence that symmetry impacts contest behaviour (e.g., [46]). In terms of theory, models of contest behaviour assume that an individual’s ability to invest in fighting is indexed to its quality (e.g., [47,48]). Nevertheless, our understanding of whether developmental asymmetry, as opposed to those asymmetries acquired through damage is associated with tactical behaviour, is limited to the question of whether an individual will yield. Indeed, as far as we are aware, with one exception, there have been no studies that have investigated the relationship between symmetry and tactical behaviour during ungulate contests [36]. In the fallow deer, winners tend to use a higher proportion of attacking actions than losers (i.e., jump clashes and backward pushes), whilst losers tend to retreat more frequently [49,50]; thus, we expect that symmetry will be associated with individual investment in attacking and retreating actions during fights.
2. Methods
2.1. Study Site and Population
This study was conducted on the fallow deer population resident at Phoenix Park, Dublin (Ireland). The park is an enclosed 707 ha site of mixed grass and woodlands with most of this area available to the deer. During the annual fawning period in June, new-born fawns are located and marked with ear tags that bore a unique colour and number, measured, weighed, and sexed before being released. Using a combination of ear tag, coat colour and the unique conformation of the antlers, all mature males in the population could be identified.
2.2. Data Collection
We used all-event sampling to collect data on fighting between males during September and October [51], and used this to estimate the total number of fights each male engaged in. We also recorded fights on video and extracted information about fight structure and tactics using the Observer video analysis system (Noldus Information Technology, Wageningen, The Netherlands). From these fights, we extracted information concerning fight duration, the number of attacking actions (backward pushes and jump clashes) and number of defensive actions (retreats, [35]) made during fights for the focal individual (N = 31, [49,50]). Because fight duration is highly variable, we estimated the numbers of contest actions per minute [49]. Males could participate in multiple fights; therefore, we averaged each contest measure and then calculated the number of tactical actions per minute prior to data analysis [43].
2.3. Antler Collection and Measurement
Antlers that were cast in April of each year were collected and kept in dry storage for a year prior to measurement (N = 54 antler pairs from different males, 17 males provided antlers for two or more years). Before casting, we ensured that there was accurate attribution of the antlers to individual males by making a photographic record of the antlers and bearers’ identity. Antlers are produced by apical growth and the rate and investment in antler growth is likely indexed to individual quality during production (e.g., [32,33]); therefore, when calculating the degree of symmetry expressed by the structure, measurements should be taken at different locations along the antler. Consequently, we took four measurements from the antler pairs (N = 75, Figure 1). We initially screened the data to test for measurement error (me%) by selecting ten pairs of antlers at random, and measuring each of the traits four times on each antler over a two month period [37]. We corrected for small sample size to assess the percentage measurement error expressed by the paired antlers [52]. Following Palmer (1994 [53]), a two-way ANOVA showed that between sides variation was significantly greater than the measurement error for all traits (coronet circumference: me% = 4.73; F (9,60) = 39.8, p < 0.001; brow tine length: me% = 12.9; F (9,60) = 493.8, p < 0.001; beam circumference: me% = 6.71; F (9,60) = 40.14, p < 0.001; antler length: me% = 5.17; F (9,60) = 503.8, p < 0.001). There was an average measurement error across the four traits of 7.4%, broadly comparable with studies employing advanced photogrammetric methods to measure antler traits (e.g., [54,55]). We then tested for directional asymmetry using paired t-tests and results indicated that three measurements taken showed FA (brow tine length: t = −0.78, p = 0.44; beam circumference: t = −1.27, p = 0.44; curved antler length: t = 1.38, p = 0.17), and one measurement, coronet circumference, showed directional asymmetry (t = 2.48, p = 0.02; mean right/left = 184.8 mm/183.2 mm). From the traits that indicated FA, we estimated symmetry as the absolute difference between the two antlers (brow tine length: mean FA = 18.23 mm, s.d. = 18.88; beam width: mean FA = 3.28 mm, s.d. = 3.33; antler length: mean FA = 30.08 mm, s.d. = 28.52, [15]), and calculated a composite score for antler FA by standardising the scores (see Statistical Analyses section below for further details), and then calculating the average the standardised score [56]. Finally, we estimated the amount of DA (R-L) expressed for the fourth antler trait, coronet circumference (mean DA = 3.83 mm, s.d. = 3.63).
2.4. Statistical Analyses
We analysed all data using the R statistical platform (v 4.2.1) using a Bayesian framework with the posterior generated by MCMC sampled through the brms package (v 2.34). Variables relating to antler symmetry, and to fight parameters were calculated as described above; in addition, we included the total number of fights and individual age as covariates, with individual identity as a random intercept [57]. We ran an initial GLMM to investigate the relationship between age and with the two measures of antler symmetry entered as predictor variables. We then ran a multiple logistic regression model to assess the relationship between antler breakage and two measures of symmetry: a composite measure of three antler traits for FA, and a single trait for DA (see above). Because antlers are costly structures to produce, several other variables could be associated with damage; therefore, we also included the number of fights each individual was recorded as participating in, and the age of the individual. We ran two additional multivariate GLMM to assess the relationship between antler symmetry (FA and DA) as the dependent variables and four different contest parameters: the duration contestants maintained antler contact, and three variables relating to the focal male’s tactics (offensive: backward pushes and jump clashes; defensive: retreats, [43,49]. We initially included contestant age as an additional covariate; however, there was no association between age and symmetry in either model; therefore, we removed age to reduce overparameterisation of the models.
Continuous variables were log transformed and mean centred prior to analyses [58], and each model was run over three independent chains with dispersed initial values. The initial model burnin was run over 5000 iterations with a posterior length of each chain of 400,000 iterations and a thinning interval set at 50 to reduce autocorrelation. We visually inspected the posterior distributions to ensure adequate mixing, and used the Gelman-Rubin statistic to confirm that convergence was satisfactory [59]. We accepted that a covariate had a meaningful relationship with a predictor variable if less than 5% of the posterior chain crossed zero (Px, [60]), and if the mean of the regressor co-efficient was not very low (i.e., less than 0.02, [61]).
3. Results
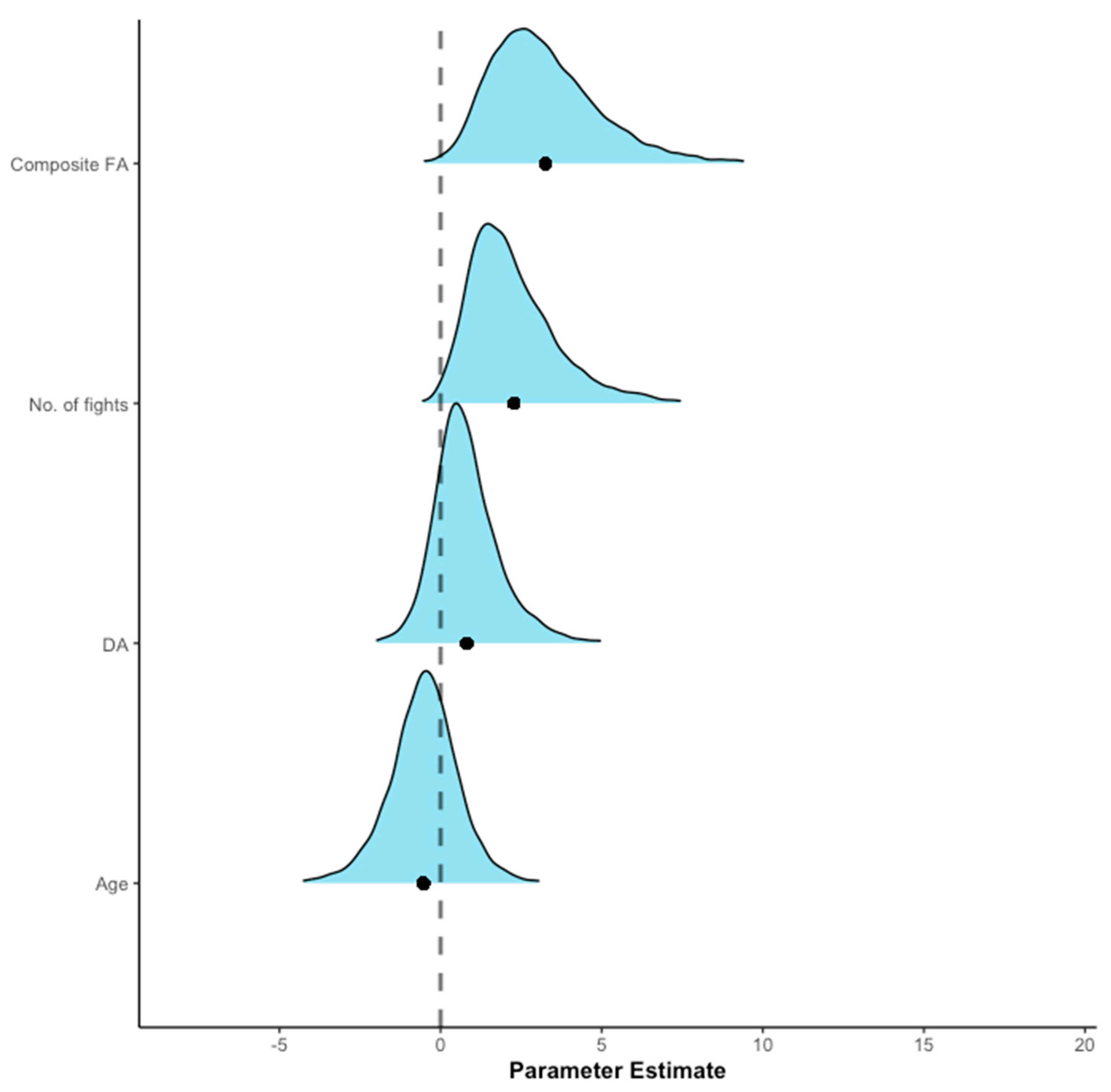
Relationship between antler damage and antler symmetry. A preliminary GLMM investigated the relationship between age and the two measures of antler symmetry (FA and DA (raw data: mean = 1.59 mm, s.d. = 5.55)); there was a very weak negative relationship between the posterior distributions for the two symmetry measures (r = −0.10), and the analysis indicated that age was positively associated with FA (mean = 0.36, s.d. = 0.16, 95% CI = 0.05, 0.67, Px = 1.03%) but not with DA (mean = 0.02, s.d. = 0.11, 95% CI = −0.19, 0.23, Px = 43.26%, R2 = 0.63). A logistic regression investigated the relationship between antler damage and the two symmetry predictors, number of fights each individual engaged in and their age. The relationship between the posterior distribution of the fixed effects was either weak or very weak: the strongest correlation was between FA and number of fights, r = 0.46, and weakest correlation was between DA and age, r = 0.01). The model indicated that there was a positive association between antler damage and a composite measure of FA (mean = 3.25, s.d. = 1.77; 95% CI = 0.62, 7.48; Px = 0.72%) and the number of fights engaged in (mean = 2.28, s.d. = 1.49, 95% CI = 0.26, 6.01; Px = 1.08%, Figure 2). There was no meaningful effect of DA (mean = 0.82, s.d. = 1.07; 95% CI = −0.94, 3.33; Px = 20.08%), or age (mean = −0.53, s.d. = 1.12, 95% CI = −2.88, 1.68; Px = 70.05%). Therefore, fluctuating asymmetry and fighting were associated with higher probability of structural damage than either directional asymmetry or individual age.
Relationship between contest tactics and symmetry. We investigated the relationship between the contest actions of individuals in relation to the degree of antler symmetry; thus, we ran two multivariate models to assess the association between FA and DA in relation to fighting tactics. Our results showed no association between either measure of antler symmetry and four contest parameters (see Table 1 and Table 2). Therefore, the level of symmetry expressed by the weapons of fallow deer has no influence on the tactical investment in fighting by the bearer.
4. Discussion
Perhaps due to the pervasive nature of symmetric structures in the natural world, the study of the extent to which individuals depart from perfect symmetry has generated considerable interest. One form of symmetry in particular, fluctuating asymmetry, has received considerable attention by students of animal behaviour as it potentially yields insights into developmental instability (e.g., [18,62,63]). Critically, this instability is thought to mirror the quality of the individual [64] and is indexed to the ability of the organism to withstand genetic and environmental stress [16]. Thus, high-quality individuals are predicted to produce large, highly symmetric structures that confer greater competitive success during intraspecific encounters, whilst also providing females with a reliable cue of the bearer’s genetic quality [65]. Our results show that as males aged there was a corresponding increase in the amount of absolute FA expressed—a finding that is consistent with some studies (e.g., [25,66]), but not others (e.g., [26,54]). We note that other studies that have employed a relative measure of asymmetry have shown a negative relationship between FA and age (e.g., [67,68]). Moreover, the antlers of fallow deer also show directional asymmetry with age [26], and DA has been reported to have considerable influence over how antlers are used during fights [26,36]. The purpose of this study was to investigate whether FA was associated with individual quality; we initially tested whether there was an association between symmetry and antler damage. We then examined whether there was an association between symmetry and tactical actions during fights. Overall, we show partial support for the idea that FA represents an index of individual quality.
Sexually selected traits are expected to represent something of the quality of the individual bearing them [1]. Whilst there are several proposed functions for horns and antlers, these structures are predominantly viewed as traits that can be used by females when selecting a mate, as signals of fighting ability, and as weapons during intrasexual contests [7,69]. These latter two ideas have been formally expressed by models of contest behaviour that stress information gathering as a strategic requirement [2,3], and several studies have shown that this is the case. For example, difference in antler size predicts whether roe deer will escalate to fighting [10], while third-party males use discrepancy in antler size between fighting opponents to decide whether to intervene in contests [70]. A key feature of the signal value of these structures involves the cost of production with individuals experiencing physiological exhaustion during the growth phase [28,32]. Moreover, because growth rate is dependent on individual quality [34], we reasoned that individuals that can afford greater investment costs should produce more symmetric, better quality, weapons (i.e., less likely to suffer damage). Thus, we hypothesised that quality is a measure of both the durability and symmetry of the armaments borne by males. Our results show that this was the case; as FA increased, there was an increase in the probability that the bearer would experience a break to one antler. However, we note that the relationship between individual quality and antler damage is complex. For example, we show that antler damage is positively associated with fighting [40,71]—a costly behaviour underpinned by individual quality [72]; moreover, there is a positive association between antler damage and social dominance [14]. Taken together, these results suggest that FA should negatively correlate with antler damage, the opposite to what we predicted and found. Further work on understanding weapon damage in relation to various correlates of individual quality is clearly needed in order to disentangle this complex relationship. Neither DA, nor the age of the bearer, showed a meaningful association with antler damage which might have been expected given how individuals use their antler during fights [36], and the greater investment by older individuals in antler growth (e.g., [73,74]).
The behaviour of individuals during fights is, at least theoretically, linked both to the ability of the individual to invest in fighting, and to the value of the resource under dispute ([75]; see also [76] and chapters within]). Models of contest behaviour allow that contestants can differ in their tactical action rates because they are expected to exert costs on the signalling individual (e.g., [2,47]); therefore, high quality individuals are expected to repeat contest actions at higher rates than lower-quality individuals. In agreement with these theoretical assumptions, numerous studies have shown that contest winners signal at higher rates than losers do [3,49,50,77,78,79]. Moreover, limited evidence from deer contests on antler use suggests that there is a tendency to use the more developed right antler during fights [36]. Contest outcome has also been associated with symmetry although studies have tended to vary in the level of support they show [45,80]. Therefore, rather than focus on outcome, we hypothesised that symmetry would be correlated with contest action rates; specifically, we predicted that the magnitude of symmetry expressed would show a negative relationship with attacking actions and a positive relationship with retreating actions [49,50]. This was not the case, and our results showed no association between attacking or retreating actions and FA or DA.
There are a number of potential explanations as to why this might be the case. It is possible that the lack of a meaningful effect is a consequence of low statistical power due to measurement error [15]; however, we note that measurement error for these antlers was smaller than between sides variation which would suggest that this is not the case (also [37]). Conversely, directional symmetry rather than FA of the antlers could play an important role in how males display their quality or engage their antlers during fights [26,36]; however, we found no evidence that DA was associated with antler damage or contest action rates. Nevertheless, FA is associated with antler breakage, and damage does have a significant impact on fight actions [43]. So it may be that the effects of FA on contest action rates are indirectly associated with fighting. In addition, we note that resource value critically influences tactical behaviour [78]; most fights occur during the annual rut where the presence and abundance of oestrus females has significant effects on how males invest in contests [38], see also [8,35,81]. Because the interaction between behavioural actions during contests and structural symmetry have been relatively neglected topics, we suggest that further research will be required to disentangle the potential explanatory effects concerning the importance of FA in relation to contest behaviour.
In conclusion, this study investigated the relationship between FA and individual quality by focusing on antlers as badges of status and as weapons [6]: thus, we examined both weapon durability and tactical behaviours used during contests to address this question. So far as we are aware, these assumed correlates of individual quality have not previously been studied in relation to weapon symmetry (but see [36]), and we show that antler damage but not tactical actions are associated with FA in the predicted direction. Moreover, there was no evidence that DA was associated with either damage or tactical actions [36]. The usefulness of FA as an explanatory factor in sexual selection and fitness (e.g., [82,83]), has been criticised on grounds of the generality of the effect (see [84] for a review, also [85]). Nevertheless, beyond the search for associations between FA and trait size for example, there are questions within the field of animal behaviour that require further investigation. We addressed two such possibilities here and suggest that investigation of the role of FA in animal behaviour remains a promising avenue of research.
Author Contributions
D.J.J.: Conceptualization, Data curation, Formal analysis, Methodology, Writing—original draft, Writing—review and editing; M.P.G.: Data curation, Writing—original draft, Writing—review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The current study was conducted in collaboration with the Office of Public Works (OPW)—the management body responsible for the management of the deer herd at Phoenix Park. The park is located close to the centre of Dublin and is open to members of the public; therefore, the deer have become habituated to human presence, and are generally tolerant of people. Observers used spotting scopes from static locations to observe and record the behaviour of free-ranging fallow deer. We generally did not approach to within 30 m of the deer, a distance that we estimated did not interfere with the behaviour of the deer.
Data Availability Statement
Not applicable.
Acknowledgments
We would like to extend thanks to the Office of Public Works and the Superintendent of Phoenix Park for facilitating the study of the deer population. We also thank Caitriona Carlin who provided invaluable help in the field with data collection.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Andersson, M. Sexual Selection; Princeton University Press: Princeton, NJ, USA, 1994. [Google Scholar]
- Enquist, M.; Leimar, O. Evolution of fighting behaviour: Decision rules and assessment of relative strength. J. Theor. Biol. 1983, 102, 387–410. [Google Scholar] [CrossRef]
- Enquist, M.; Leimar, O.; Ljungberg, T.; Mallner, Y.; Segerdahl, N. A test of the sequential assessment game: Fighting in the cichlid fish Nanncara anomala. Anim. Behav. 1990, 40, 1–14. [Google Scholar] [CrossRef]
- Rower, S. The evolution of reliable and unreliable badges of fighting ability. Am. Zool. 1982, 22, 531–546. [Google Scholar] [CrossRef] [Green Version]
- Johnstone, R.A.; Norris, K. Badges of status and the cost of aggression. Behav. Ecol. Sociobiol. 1993, 32, 127–134. [Google Scholar] [CrossRef]
- Berglund, A.; Bisazza, A.; Pilastro, A. Armaments and ornaments: An evolutionary explanation of traits of dual utility. Biol. J. Linn. Soc. 1996, 58, 385–399. [Google Scholar] [CrossRef]
- Geist, V. Mountain Sheep: A Study in Behaviour and Evolution; University of Chicago Press: Chicago, IL, USA, 1971. [Google Scholar]
- Clutton-Brock, T.; Albon, S.D.; Gibson, R.M.; Guinness, F.E. The logical stag: Adaptive aspects of fighting in red deer (Cervus elaphus L.). Anim. Behav. 1979, 27, 211–225. [Google Scholar] [CrossRef]
- Barrette, C.; Vandal, D. Sparring, relative antler size, and assessment in male caribou. Behav. Ecol. Sociobiol. 1990, 26, 383–387. [Google Scholar] [CrossRef]
- Hoem, S.A.; Melis, C.; Linnell, J.D.C.; Andersen, R. Fighting behaviour in territorial male roe deer Capreolus capreolus: The effects of antler size and residence. Eur. J. Wildl. Res. 2007, 53, 1–8. [Google Scholar] [CrossRef]
- Sneddon, L.U.; Huntingford, F.A.; Taylor, A.C. Weapon size versus body size as a predictor of winning in fights between shore crabs, Carcinus maenas (L.). Behav. Ecol. Sociobiol. 1997, 41, 237–242. [Google Scholar] [CrossRef] [Green Version]
- Kelly, C.D. Fighting for harems: Assessment strategies during male–male contests in the sexually dimorphic Wellington tree weta. Anim. Behav. 2006, 72, 727–736. [Google Scholar] [CrossRef]
- Lailvaux, S.P.; Hathway, J.; Pomfret, J.; Knell, R.J. Horn size predicts physical performance in the beetle Euoniticellus intermedius (Coleoptera: Scarabaeidae). Funct. Ecol. 2005, 19, 632–639. [Google Scholar] [CrossRef]
- Jennings, D.J.; Gammell, M.P.; Carlin, C.M.; Hayden, T.J. Is Difference in Body Weight, Antler Length, Age or Dominance Rank related to the Number of Fights between Fallow Deer (Dama dama)? Ethology 2006, 112, 258–269. [Google Scholar] [CrossRef]
- Palmer, A.R.; Strobeck, C. Fluctuating asymmetry: Measurement, analysis, patterns. Annu. Rev. Ecol. Syst. 1986, 17, 391–421. [Google Scholar] [CrossRef]
- Parsons, P.A. Fluctuating Asymmetry: An Epigenetic Measure of Stress. Biol. Rev. 1990, 65, 131–145. [Google Scholar] [CrossRef]
- Solberg, E.J.; Sæther, B.E. Fluctuating asymmetry in the antlers of moose (Alces alces): Does it signal male quality? Proc. R. Soc. B 1993, 254, 251–255. [Google Scholar]
- Møller, A.P.; Swaddle, J.P. Asymmetry, Developmental Stability, and Evolution; Oxford University Press: Oxford, UK, 1997. [Google Scholar]
- Chirichella, R.; Rocca, M.; Brugnoli, A.; Mustoni, A.; Apollonio, M. Fluctuating asymmetry in Alpine chamois horns: An indicator of environmental stress. Evol. Ecol. 2020, 34, 573–587. [Google Scholar] [CrossRef]
- Folstad, I.; Arneberg, P.; Karter, A.J. Antlers and parasites. Oecologia 1996, 105, 556–558. [Google Scholar] [CrossRef]
- Pélabon, C.; van Breukelen, L. Asymmetry in antler size in roe deer (Capreolus capreolus): An index of individual and population conditions. Oecologia 1998, 116, 1–8. [Google Scholar] [CrossRef]
- Lens, L.; van Dongen, S.; Kark, S.; Matthysen, E. Fluctuating asymmetry as an indicator of fitness: Can we bridge the gap between studies? Biol. Rev. 2002, 77, 27–38. [Google Scholar] [CrossRef] [Green Version]
- Bartoš, L.; Bahbouh, R. Antler size and fluctuating asymmetry in red deer (Cervus elaphus) stags and probability of becoming a harem holder in rut. Biol. J. Linn. Soc. 2006, 87, 59–68. [Google Scholar] [CrossRef] [Green Version]
- Ciuti, S.; Apollonio, M. Do antlers honestly advertise the phenotypic quality of fallow buck (Dama dama) in a lekking population? Ethology 2011, 117, 133–144. [Google Scholar] [CrossRef]
- Kruuk, L.E.B.; Slate, J.; Pemberton, J.M.; Clutton-Brock, T.H. Fluctuating asymmetry in a secondary sexual trait: No associations with individual fitness, environmental stress or inbreeding, and no heritability. J. Evol. Biol. 2003, 16, 101–113. [Google Scholar] [CrossRef]
- Pélabon, C.; Joly, P. What, if anything, does visual asymmetry in fallow deer antlers reveal? Anim. Behav. 2000, 59, 193–199. [Google Scholar] [CrossRef] [Green Version]
- Chapman, D.; Chapman, N. Fallow Deer: Their History, Distribution, and Biology; Coch-y-Bonddu Books: Machynlleth, UK, 1997. [Google Scholar]
- Emlen, D.J. The Evolution of Animal Weapons. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 387–413. [Google Scholar] [CrossRef] [Green Version]
- Moen, R.; Pastor, J. Simulating antler growth and energy, nitrogen, calcium and phosphorus metabolism in caribou. Rangifer 1998, 18, 85–97. [Google Scholar] [CrossRef] [Green Version]
- Moen, R.A.; Pastor, J.; Cohen, J. Antler growth and extinction of Irish elk. Evol. Ecol. Res. 1999, 1, 235–249. [Google Scholar]
- Landete-Castillejos, T.; Estevez, J.A.; Ceacero, F.; Garcia, A.J.; Gallego, L. A review of factors effecting antler composition and mechanics. Front. Biosci. 2012, E4, 2328–2339. [Google Scholar] [CrossRef]
- Landete-Castillejos, T.; Estevez, J.; Martínez, A.; Ceacero, F.; Garcia, A.; Gallego, L. Does chemical composition of antler bone reflect the physiological effort made to grow it? Bone 2007, 40, 1095–1102. [Google Scholar] [CrossRef]
- Bartoš, L.; Perner, V.; Losos, S. Red deer stags rank position, body weight and antler growth. Acta Thériol. 1988, 33, 209–217. [Google Scholar] [CrossRef] [Green Version]
- Bartoš, L.; Losos, S. Response of antler growth to changing rank of fallow deer buck during the velvet period. Can. J. Zoöl. 1997, 75, 1934–1939. [Google Scholar] [CrossRef]
- Alvarez, F. Risks of fighting in relation to age and territory holding in fallow deer. Can. J. Zool. 1993, 71, 376–383. [Google Scholar] [CrossRef]
- Alvarez, F. Functional directional asymmetry in fallow deer (Dama dama) antlers. J. Zool. 1995, 236, 563–569. [Google Scholar] [CrossRef]
- Jennings, D.J.; Gammell, M.P.; Carlin, C.M.; Hayden, T.J. Does Lateral Presentation of the Palmate Antlers During Fights by Fallow Deer (Dama dama L.) Signify Dominance or Submission? Ethology 2002, 108, 389–401. [Google Scholar] [CrossRef]
- Jennings, D.J. Contest behaviour varies in relation to reproductive opportunities and reproductive success in the fallow deer. Anim. Behav. 2020, 163, 95–103. [Google Scholar] [CrossRef]
- Bartoš, L. Relationships between Behaviour and Antler Cycle Timing in Red Deer. Ethology 1986, 71, 305–314. [Google Scholar] [CrossRef]
- Johnson, H.E.; Bleich, V.C.; Krausman, P.R.; Koprowski, J.L. Effects of antler breakage on mating behavior (Cervus elephus nannodes). Eur. J. Wildl. Res. 2007, 53, 9–15. [Google Scholar] [CrossRef]
- Lincoln, G.A. The role of antlers in the behaviour of red deer. J. Exp. Zoöl. 1972, 182, 233–249. [Google Scholar] [CrossRef]
- Lincoln, G.A.; Youngson, R.W.; Short, R.V. The social and sexual behaviour of the red deer stag. J. Reprod. Fertil. Suppl. 1970, 11, 71–103. [Google Scholar]
- Jennings, D.J.; Boys, R.J.; Gammell, M.P. Weapon damage is associated with contest dynamics but not mating success in fallow deer (Dama dama). Biol. Lett. 2017, 13, 20170565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munoz, N.E.; Zink, A.G. Asymmetric Forceps Increase Fighting Success among Males of Similar size in the Maritime Earwig. Ethology 2012, 118, 943–954. [Google Scholar] [CrossRef] [PubMed]
- Sneddon, L.U.; Swaddle, J.P. Asymmetry and fighting performance in the shore crab Carcinus maenas. Anim. Behav. 1999, 58, 431–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jablonski, P.; Matyjasiak, P. Chaffinch (Fringilla Coelebs) Epaulette Display Depends on the Degree of Exposure But Not Symmetry of Intruder’s Epaulettes. Behaviour 1997, 134, 1115–1121. [Google Scholar] [CrossRef]
- Payne, R.J. Gradually escalating fights and displays: The cumulative assessment model. Anim. Behav. 1998, 56, 651–662. [Google Scholar] [CrossRef] [PubMed]
- Payne, R.J.; Pagel, M. Escalation and Time Costs in Displays of Endurance. J. Theor. Biol. 1996, 183, 185–193. [Google Scholar] [CrossRef]
- Jennings, D.J.; Gammell, M.P.; Carlin, C.M.; Hayden, T.J. Win, lose or draw: A comparison of fight structure based on fight conclusion in the fallow deer. Behaviour 2005, 142, 423–439. [Google Scholar] [CrossRef] [Green Version]
- Jennings, D.J.; Gammell, M.P.; Payne, R.J.H.; Hayden, T.J. An Investigation of Assessment Games During Fallow Deer Fights. Ethology 2005, 111, 511–525. [Google Scholar] [CrossRef]
- Altmann, J. Observational Study of Behavior: Sampling Methods. Behaviour 1974, 49, 227–267. [Google Scholar] [CrossRef] [Green Version]
- Haldane, J.B.S. The measurement of variation. Evolution 1955, 9, 484. [Google Scholar] [CrossRef]
- Palmer, A.R. Fluctuating asymmetry analyses: A primer. In Developmental Instability: Its Origins and Evolutionary Implications; Markow, T.A., Ed.; Kluwer: Dordrecht, The Netherlands, 1994; pp. 335–364. [Google Scholar]
- Ditchkoff, S.S.; de Freese, R.L. Assessing fluctuating asymmetry of white-tailed deer antlers in a three-dimensional context. J. Mammal. 2010, 91, 27–37. [Google Scholar] [CrossRef]
- Tsuboi, M.M.; Kopperud, B.T.; Syrowatka, C.; Grabowski, M.; Voje, K.; Pélabon, C.; Hansen, T.F. Measuring complex morphological traits with 3D photogrammetry: A case study with deer antlers. Evol. Biol. 2020, 47, 175–186. [Google Scholar] [CrossRef] [Green Version]
- Leung, B.; Forbes, M.R.; Houle, D. Fluctuating asymmetry as a bioindicator of stress: Comparing efficacy of analyses involving multiple traits. Am. Nat. 2000, 155, 101–115. [Google Scholar] [CrossRef] [PubMed]
- Gelman, A.; Jakulin, A.; Pittau, M.G.; Su, Y.-S. A weakly informative default prior distribution for logistic and other regression models. Ann. Appl. Stat. 2008, 2, 1360–1383. [Google Scholar] [CrossRef]
- Kruschke, J.K. Doing Bayesian Data Analysis; Elsevier: Amsterdam, The Netherlands, 2015. [Google Scholar]
- Gelman, A.; Carlin, J.; Stern, H.; Rubin, D. Bayesian Data Analysis; CRC Press: Boca Raton, FL, USA, 2013. [Google Scholar]
- Jennings, D.J.; Boys, R.J.; Gammell, M.P. Suffering third-party intervention during fighting is associated with reduced mating success in the fallow deer. Anim. Behav. 2018, 139, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Bridger, D.; Bonner, S.J.; Briffa, M. Individual quality and personality: Bolder males are less fecund in the hermit crab Pagurus bernhardus. Proc. R. Soc. B Biol. Sci. 2015, 282, 20142492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leamy, L.J.; Klingenberg, C.P. The Genetics and Evolution of Fluctuating Asymmetry. Annu. Rev. Ecol. Evol. Syst. 2005, 36, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Polak, M. (Ed.) Developmental Instability: Causes and Consequences; Oxford University Press: Oxford, UK, 2003. [Google Scholar]
- Møller, A.P.; Pomiankowski, A. Fluctuating asymmetry and sexual selection. Genetica 1993, 89, 267–279. [Google Scholar] [CrossRef]
- Møller, A.P. Fluctuating asymmetry in male sexual ornaments may reliably reveal male quality. Anim. Behav. 1990, 40, 1185–1187. [Google Scholar] [CrossRef]
- Mateos, C.; Alarcos, S.; Carranza, J.; Sánchez-Prieto, C.B.; Valencia, J. Fluctuating asymmetry of read deer antlers negatively relates to individual condition and proximity to prime age. Anim. Behav. 2008, 75, 1629–1640. [Google Scholar] [CrossRef]
- Putman, R.J.; Sullivan, M.S. Fluctuating asymmetry in antlers of fallow deer (Dama dama): The relative roles of environmental stress and sexual selection. Biol. J. Linn. Soc. 2000, 70, 27–36. [Google Scholar] [CrossRef]
- Ditchkoff, S.S.; Lochmiller, R.L.; Masters, R.E.; Starry, W.R.; Leslie, D.M., Jr. Does fluctuating asymmetry of antlers in white-tailed deer (Odocoileus virginianus) follow patters predicted for sexually selected traits? Proc. R. Soc. Biol. Sci. B 2001, 268, 891–898. [Google Scholar] [CrossRef] [Green Version]
- Clutton-Brock, T.H. The Functions of Antlers. Behaviour 1982, 79, 108–124. [Google Scholar] [CrossRef]
- Jennings, D.J.; Gammell, M.P. Bystander fallow deer engage in third-party behaviour based on similarities in contestant resource-holding potential. Anim. Behav. 2022, 191, 33–41. [Google Scholar] [CrossRef]
- Hayden, T.J.; Lynch, J.M.; O’Corry-Crowe, G. Antler growth and morphology in a feral sika deer (Cervus nippon) population in Killarney, Ireland. J. Zoöl. 1994, 232, 21–35. [Google Scholar] [CrossRef]
- Briffa, M.; Sneddon, L.U. Physiological constraints on contest behaviour. Funct. Ecol. 2007, 21, 627–637. [Google Scholar] [CrossRef] [Green Version]
- Stewart, K.M.; Bowyer, R.T.; Kie, J.G.; Gawaway, W.C. Antler size relative to body mass in moose: Tradeoffs associated with reproduction. Alces 2000, 36, 77–83. [Google Scholar]
- Vanpé, C.; Gaillard, J.-M.; Kjellander, P.; Mysterud, A.; Magnien, P.; Delorme, D.; van Laere, G.; Klein, F.; Liberg, O.; Hewison, A.J.M. Antler size provides an honest signal of male phenotypic quality in roe deer. Am. Nat. 2007, 169, 481–493. [Google Scholar] [CrossRef]
- Arnott, G.; Elwood, R.W. Information gathering and decision making about resource value in animal contests. Anim. Behav. 2008, 76, 529–542. [Google Scholar] [CrossRef]
- Hardy, I.C.W.; Briffa, M. Animal Contests; Cambridge University Press: Cambridge, UK, 2013. [Google Scholar]
- Clutton-Brock, T.; Albon, S.D. The Roaring of Red Deer and the Evolution of Honest Advertisement. Behaviour 1979, 69, 145–170. [Google Scholar] [CrossRef] [Green Version]
- Briffa, M.; Elwood, R.W.; Dick, J.T.A. Analysis of repeated signals during shell fights in the hermit crab Pagurus bernhardus. Proc. R. Soc. Biol. Sci. B 1998, 265, 1467–1474. [Google Scholar] [CrossRef] [Green Version]
- Green, P.A.; Patek, S.N. Mutual assessment during ritualized fighting in mantis shrimp (Stomatopoda). Proc. Biol. Sci. 2018, 285, 20172542. [Google Scholar] [CrossRef] [Green Version]
- Thornhill, R. Fluctuating asymmetry and the mating system of the Japanese scorpionfly, Panorpa japonica. Anim. Behav. 1992, 44, 867–879. [Google Scholar] [CrossRef]
- Jennings, D.J.; Gammell, M.P.; Carlin, C.M.; Hayden, T.J. Effect of body weight, antler length, resource value and experience on fight duration and intensity in fallow deer. Anim. Behav. 2004, 68, 213–221. [Google Scholar] [CrossRef]
- Møller, A.P.; Thornhill, R. A meta-analysis of the heritability of developmental stability. Am. Nat. 1997, 151, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Møller, A.P. Patterns of fluctuating asymmetry in weapons: Evidence for reliable signalling of quality in beetle horns and bird spurs. Proc. R. Soc. B Biol. Sci. 1992, 248, 199–206. [Google Scholar] [CrossRef]
- Tomkins, J.L.; Simmons, L. Fluctuating asymmetry and sexual selection: Paradigm shifts, publication bias, and observer expectation. In Developmental Instability: Causes and Consequences; Polak, M., Ed.; Oxford University Press: New York, NY, USA, 2003; pp. 231–261. [Google Scholar]
- Simmons, L.W.; Tomkins, J.L.; Kotiaho, J.S.; Hunt, J. Fluctuating paradigm. Proc. R. Soc. B 1999, 266, 593–595. [Google Scholar] [CrossRef]
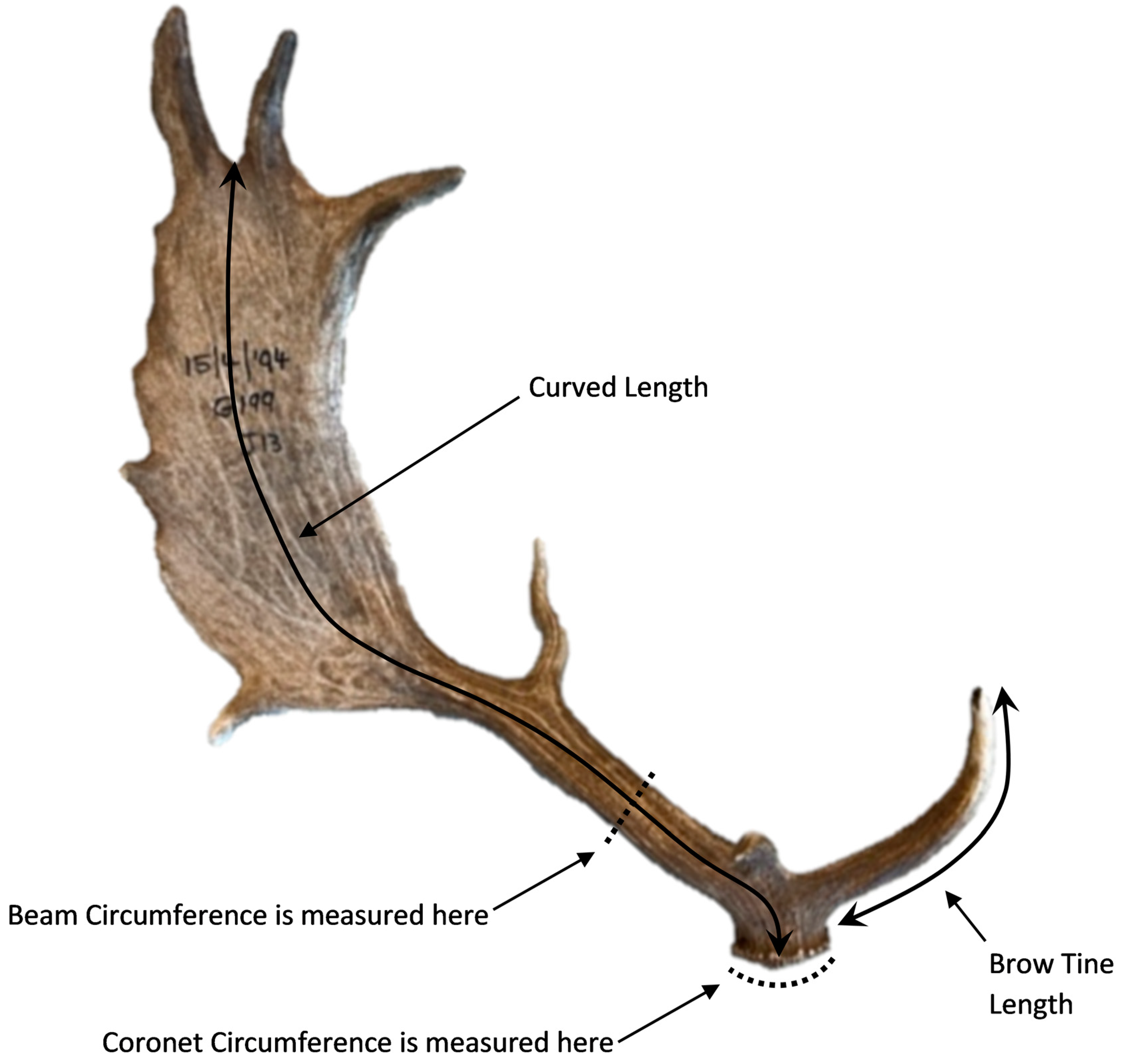
Figure 1.
A fallow deer antler showing the four measurements taken (mm) for the composite scores for antler fluctuating asymmetry and size.
Figure 1.
A fallow deer antler showing the four measurements taken (mm) for the composite scores for antler fluctuating asymmetry and size.

Figure 2.
The mean distribution and 95% credible intervals (CI) showing the association between antler damage and two measures of antler symmetry, number of fights and age (model R2 = 0.69). The black circle represents the mean value of the posterior distribution for the parameters. The intercept is excluded from the figure (mean = −1.38, s.d. = 1.17, 95% CI = −4.12, 0.60).
Figure 2.
The mean distribution and 95% credible intervals (CI) showing the association between antler damage and two measures of antler symmetry, number of fights and age (model R2 = 0.69). The black circle represents the mean value of the posterior distribution for the parameters. The intercept is excluded from the figure (mean = −1.38, s.d. = 1.17, 95% CI = −4.12, 0.60).


{kind=link}
{kind=link}
Table 1.
Posterior statistics for the model assessing the association between antler FA and contest tactics showing the mean effect sizes with their standard deviations, lower and upper 95% credible intervals (model R2 = 0.22), and the percentage of the posterior chain that crossed zero (Px).
Table 1.
Posterior statistics for the model assessing the association between antler FA and contest tactics showing the mean effect sizes with their standard deviations, lower and upper 95% credible intervals (model R2 = 0.22), and the percentage of the posterior chain that crossed zero (Px).
| Parameter | Mean | s.d. | 95% CI | %Px |
|---|---|---|---|---|
| Intercept | 20.76 | 2.11 | 16.55, 24.95 | |
| Duration | −1.13 | 2.18 | −5.39, 3.18 | 70.19 |
| Backward pushes | −0.15 | 2.87 | −5.41, 5.78 | 47.90 |
| Jump clashes | −0.66 | 2.81 | −6.21, 4.83 | 59.39 |
| Retreats | −1.99 | 2.20 | −6.23, 2.39 | 81.66 |
Table 2.
Posterior statistics for the model assessing the association between antler DA and contest tactics showing the mean effect sizes with their standard deviations, lower and upper 95% credible intervals (model R2 = 0.31), and the percentage of the posterior chain that crossed zero (Px).
Table 2.
Posterior statistics for the model assessing the association between antler DA and contest tactics showing the mean effect sizes with their standard deviations, lower and upper 95% credible intervals (model R2 = 0.31), and the percentage of the posterior chain that crossed zero (Px).
| Parameter | Mean | s.d. | 95% CI | %Px |
|---|---|---|---|---|
| Intercept | 1.76 | 0.84 | 0.11, 3.43 | |
| Duration | −0.85 | 0.92 | −2.68, 0.94 | 81.93 |
| Backward pushes | −0.76 | 1.23 | −3.14, 1.67 | 74.20 |
| Jump clashes | 0.76 | 1.19 | −3.15, 1.60 | 75.15 |
| Retreats | 0.98 | 0.89 | −0.76, 2.74 | 13.46 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Jennings, D.J.; Gammell, M.P. Fluctuating Asymmetry of Fallow Deer Antlers Is Associated with Weapon Damage but Not Tactical Behaviour during Fights. Symmetry 2023, 15, 829. https://doi.org/10.3390/sym15040829
AMA Style
Jennings DJ, Gammell MP. Fluctuating Asymmetry of Fallow Deer Antlers Is Associated with Weapon Damage but Not Tactical Behaviour during Fights. Symmetry. 2023; 15(4):829. https://doi.org/10.3390/sym15040829
Chicago/Turabian StyleJennings, Dómhnall J., and Martin P. Gammell. 2023. "Fluctuating Asymmetry of Fallow Deer Antlers Is Associated with Weapon Damage but Not Tactical Behaviour during Fights" Symmetry 15, no. 4: 829. https://doi.org/10.3390/sym15040829
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.