
Adaptability of Bony Armor Elements of the Threespine Stickleback Gasterosteus aculeatus (Teleostei: Gasterosteidae): Ecological and Evolutionary Insights from Symmetry Analyses

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling Sites
2.2. Predation Pressure
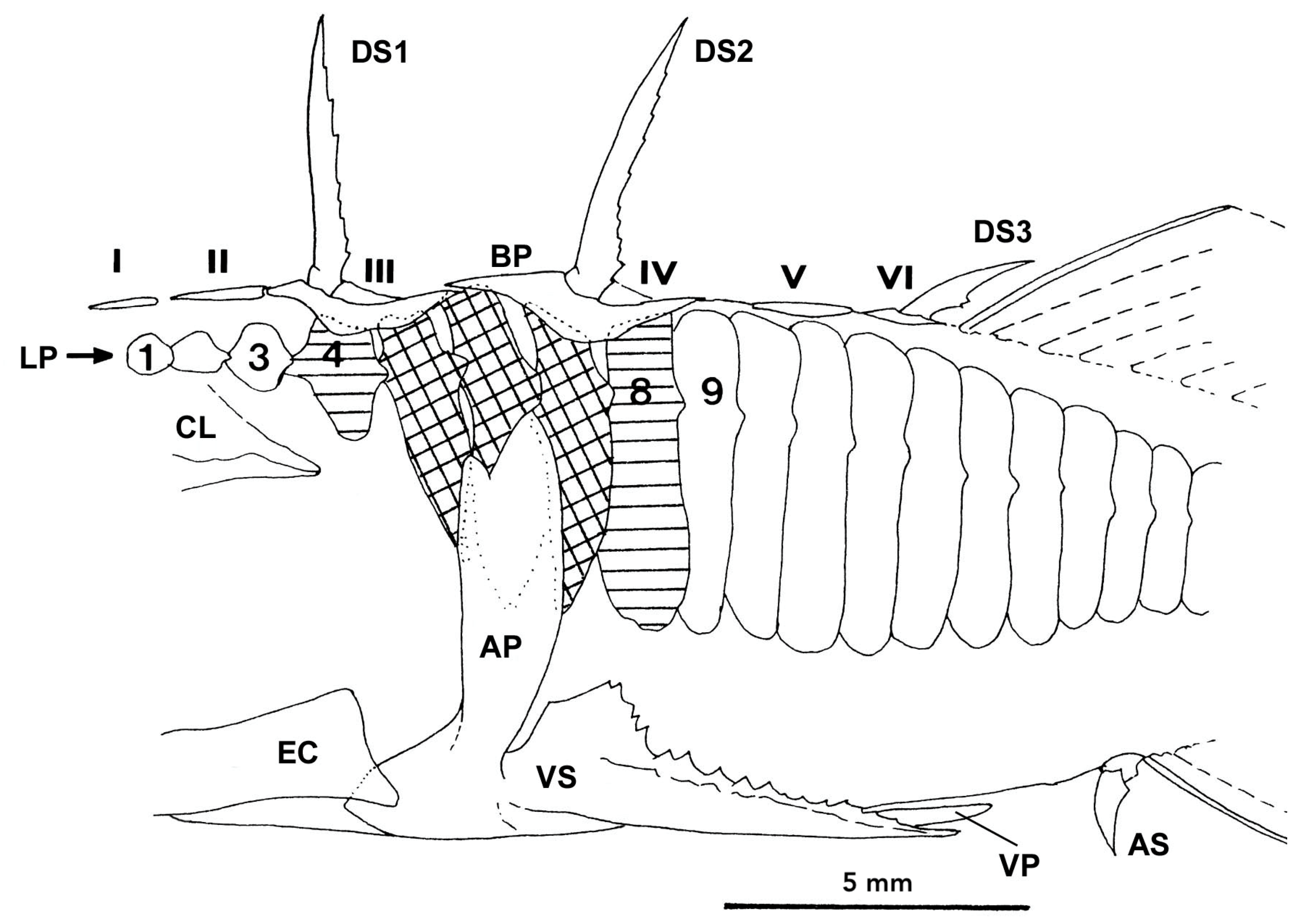
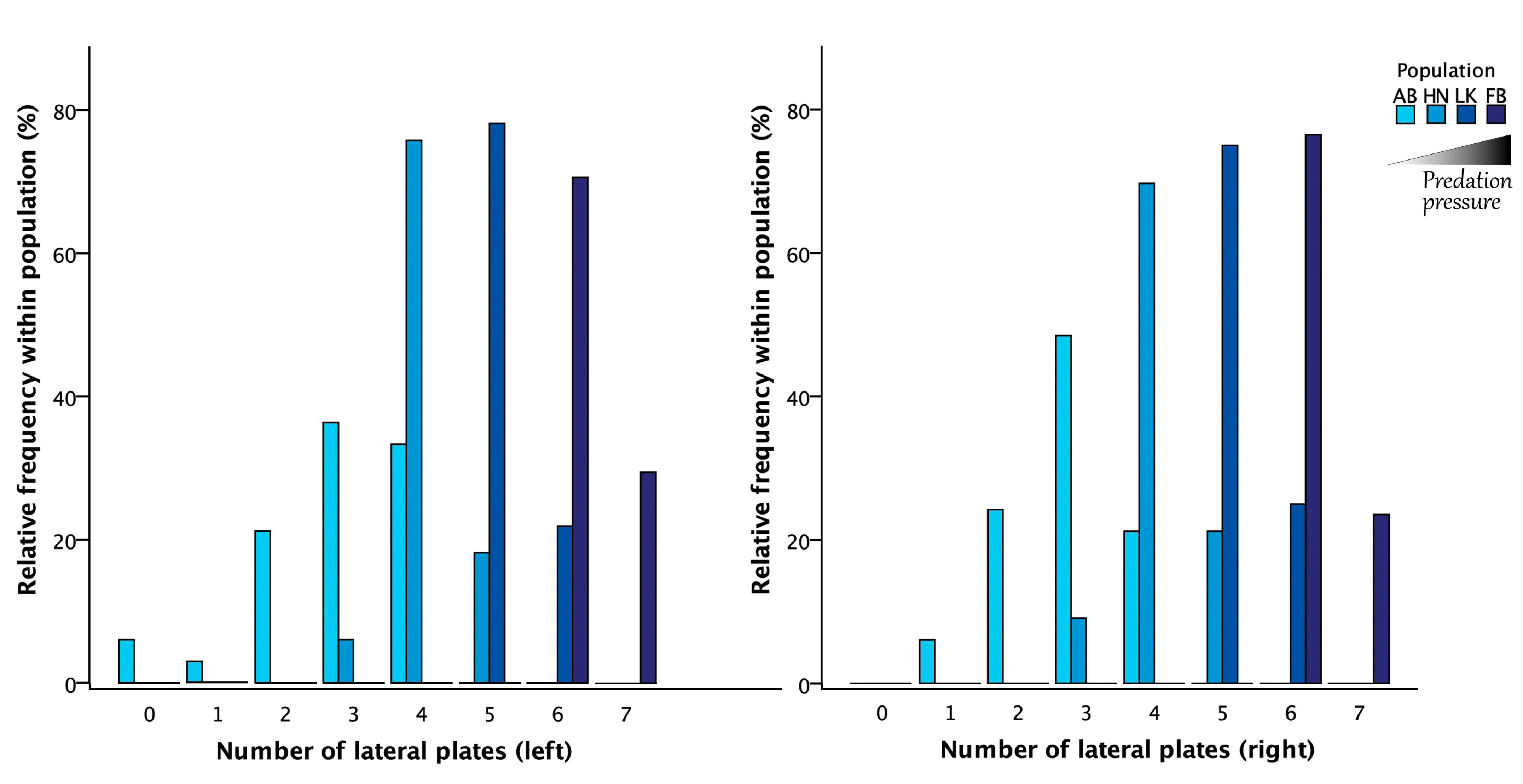
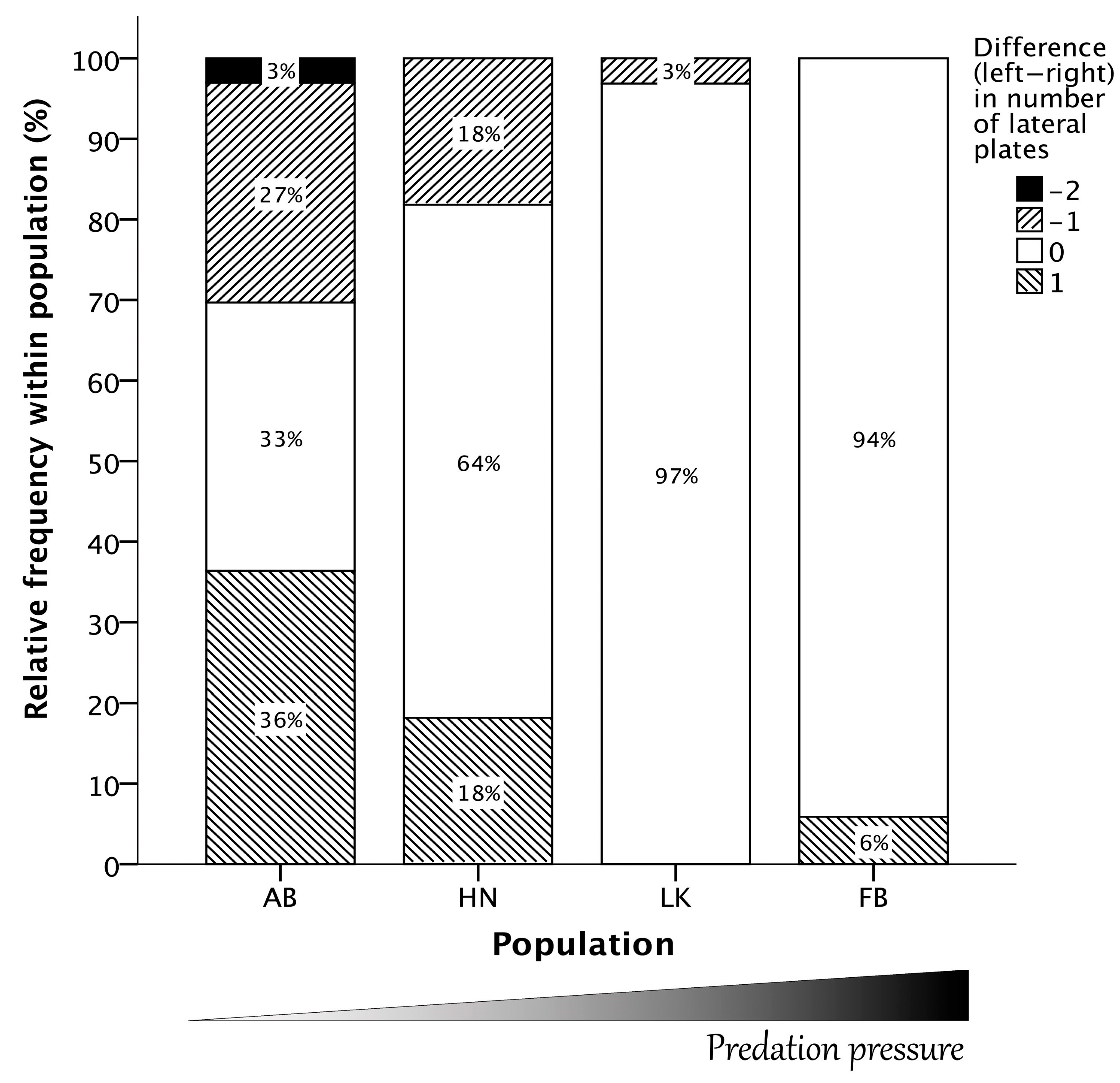
2.3. The Lateral Plates
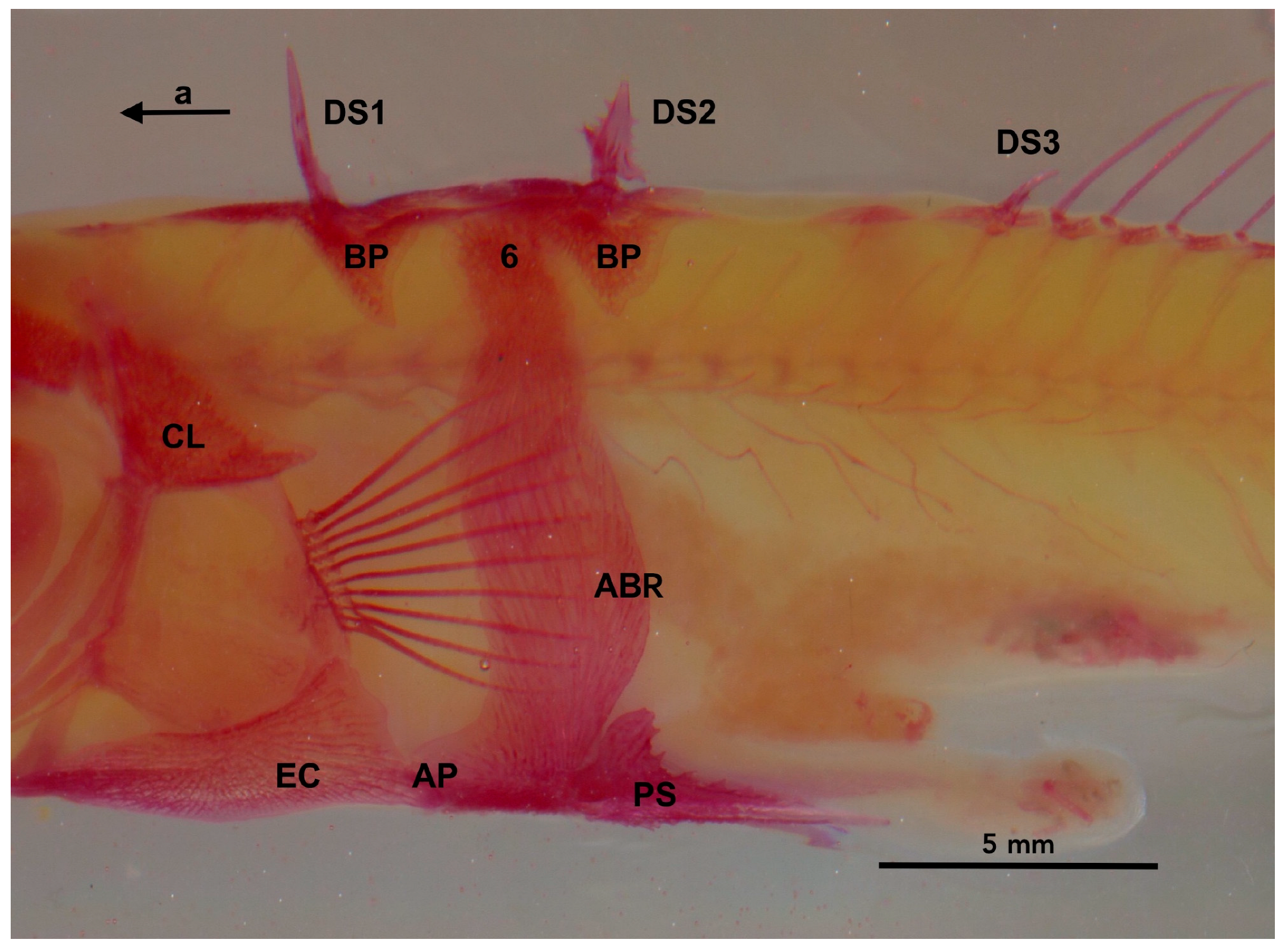
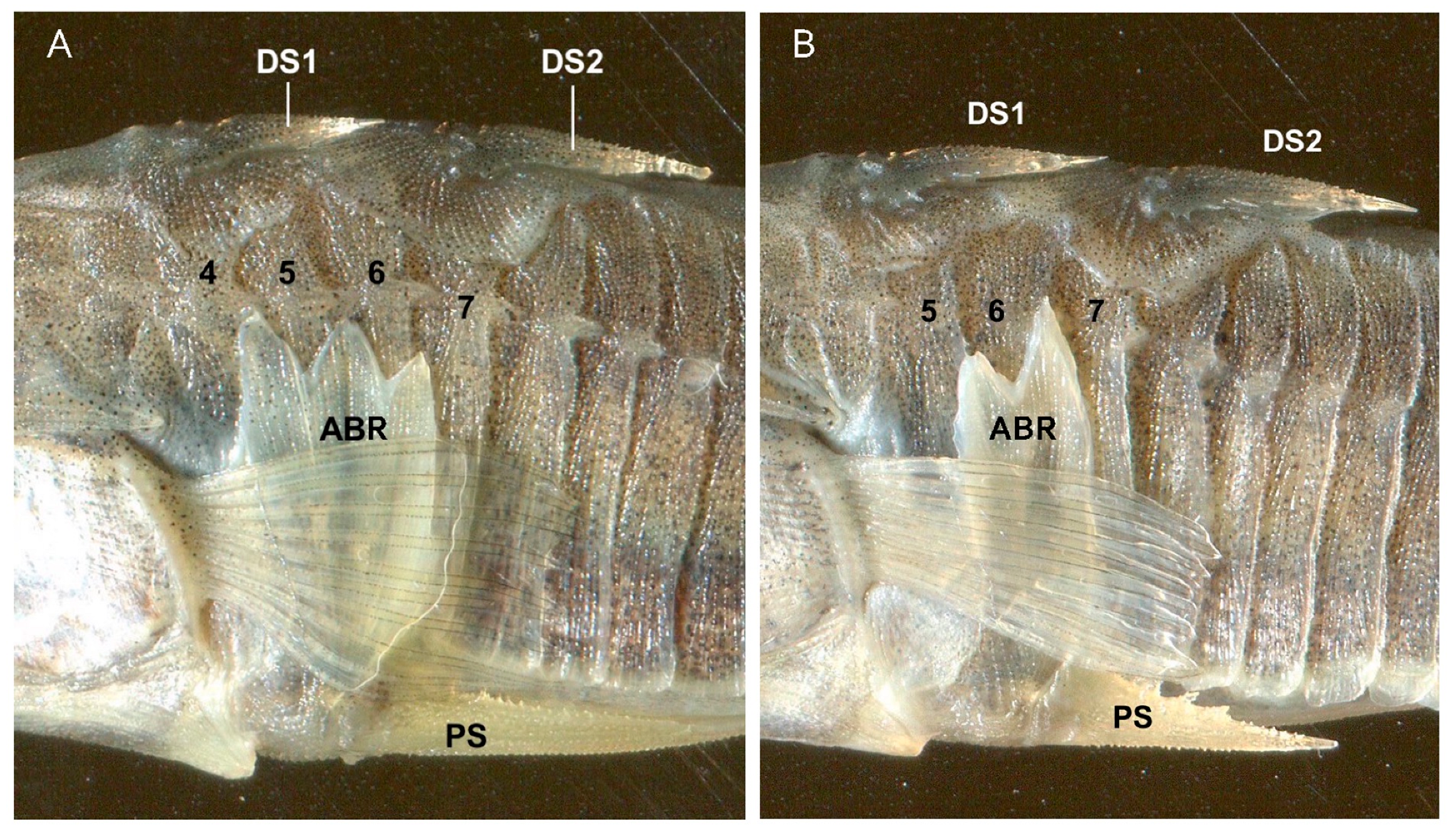
2.4. The Defensive Complex (DC)
2.4.1. Structural Lateral Plates
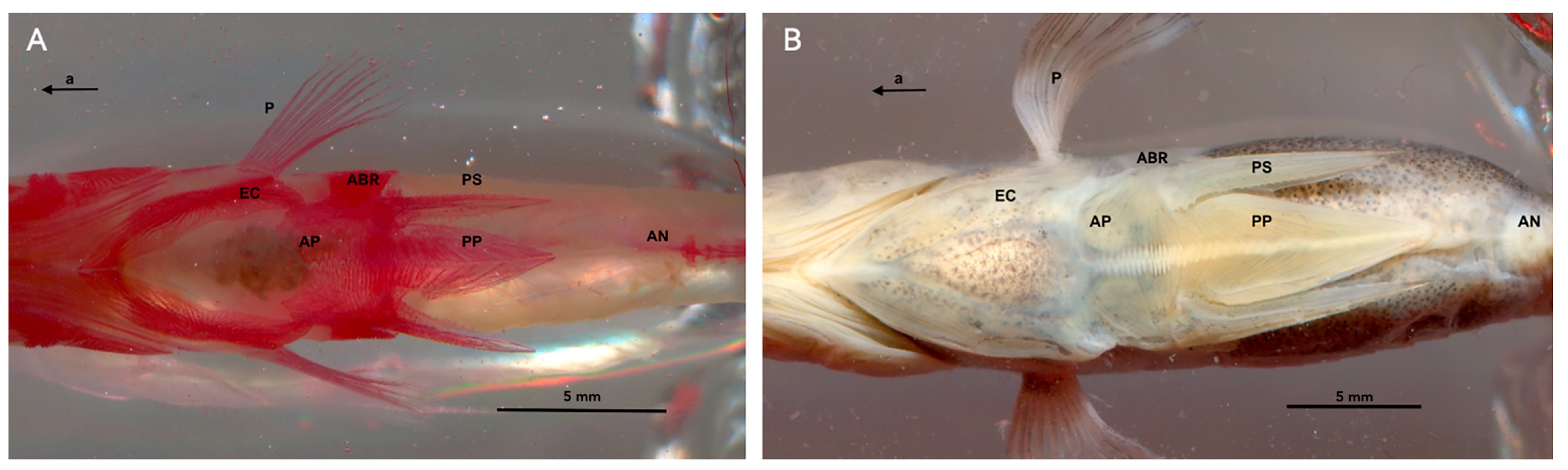
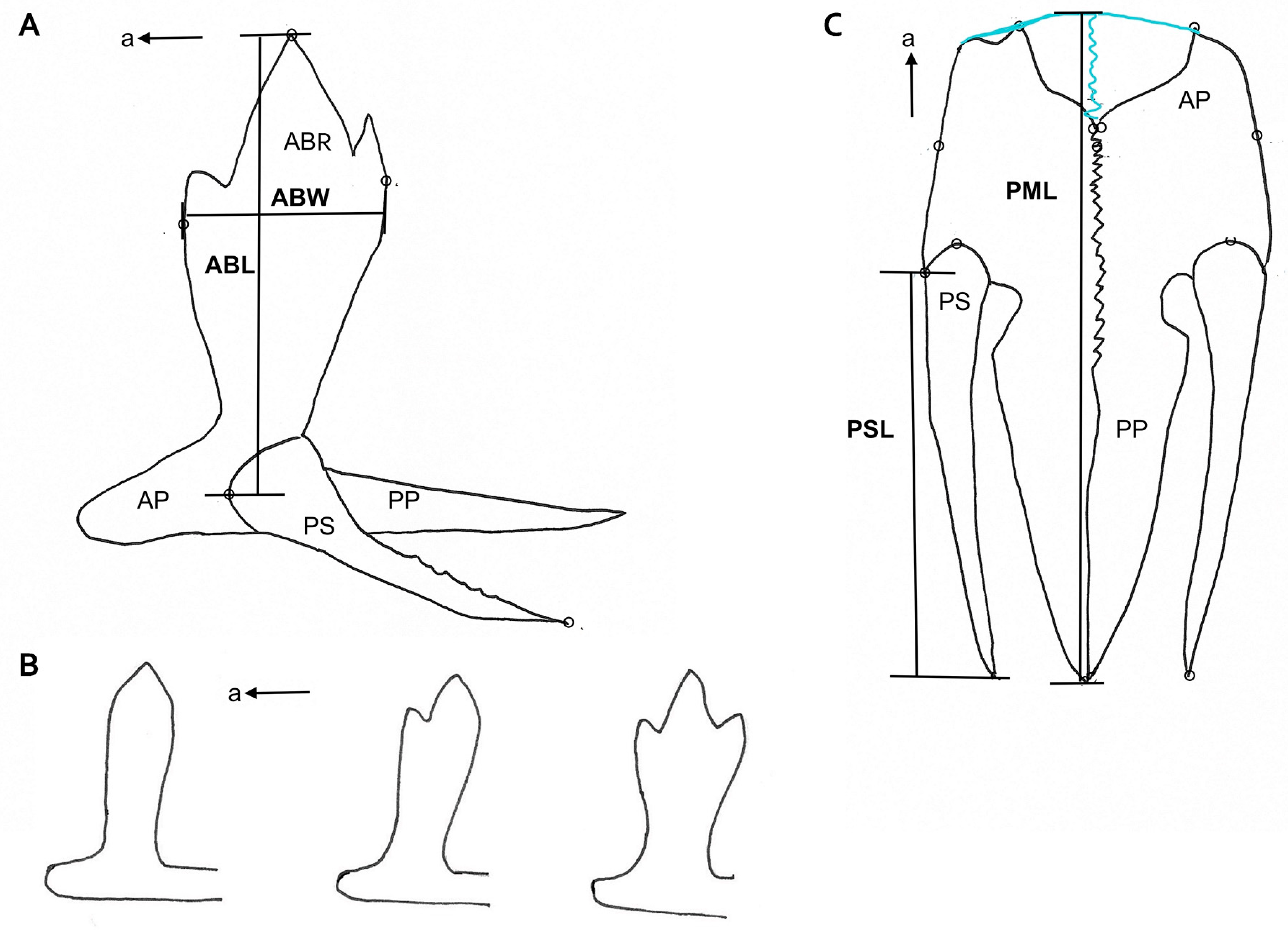
2.4.2. The Pelvic Complex
2.5. Data Acquisition
2.6. Statistical Analyses
3. Results
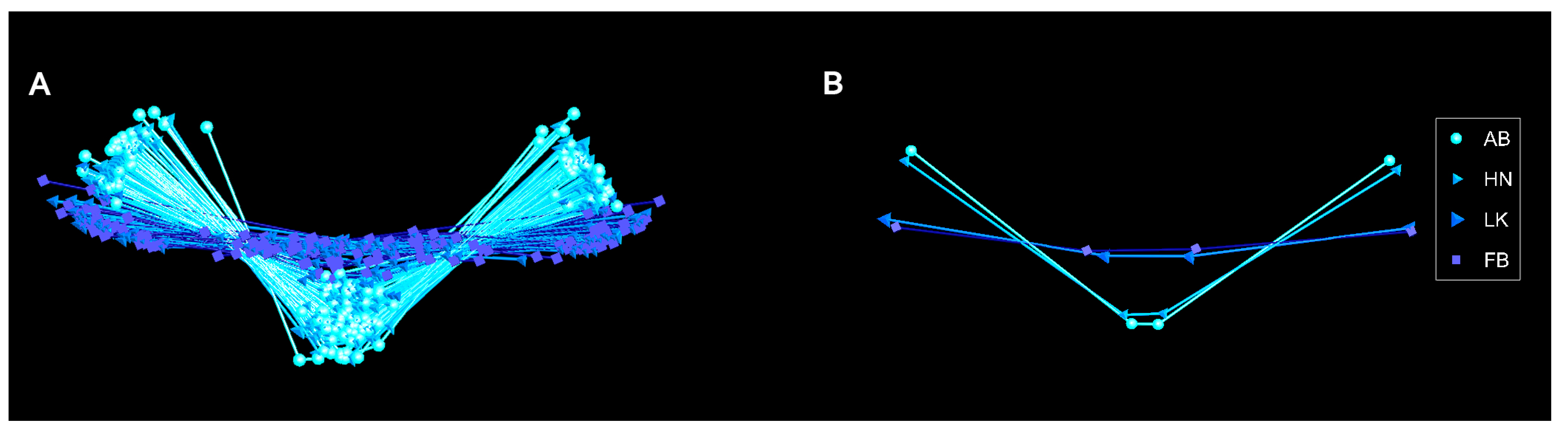
3.1. Symmetry Analyses of the Structural Lateral Plate Numbers
3.2. Symmetry Analyses of the Pelvic Complex
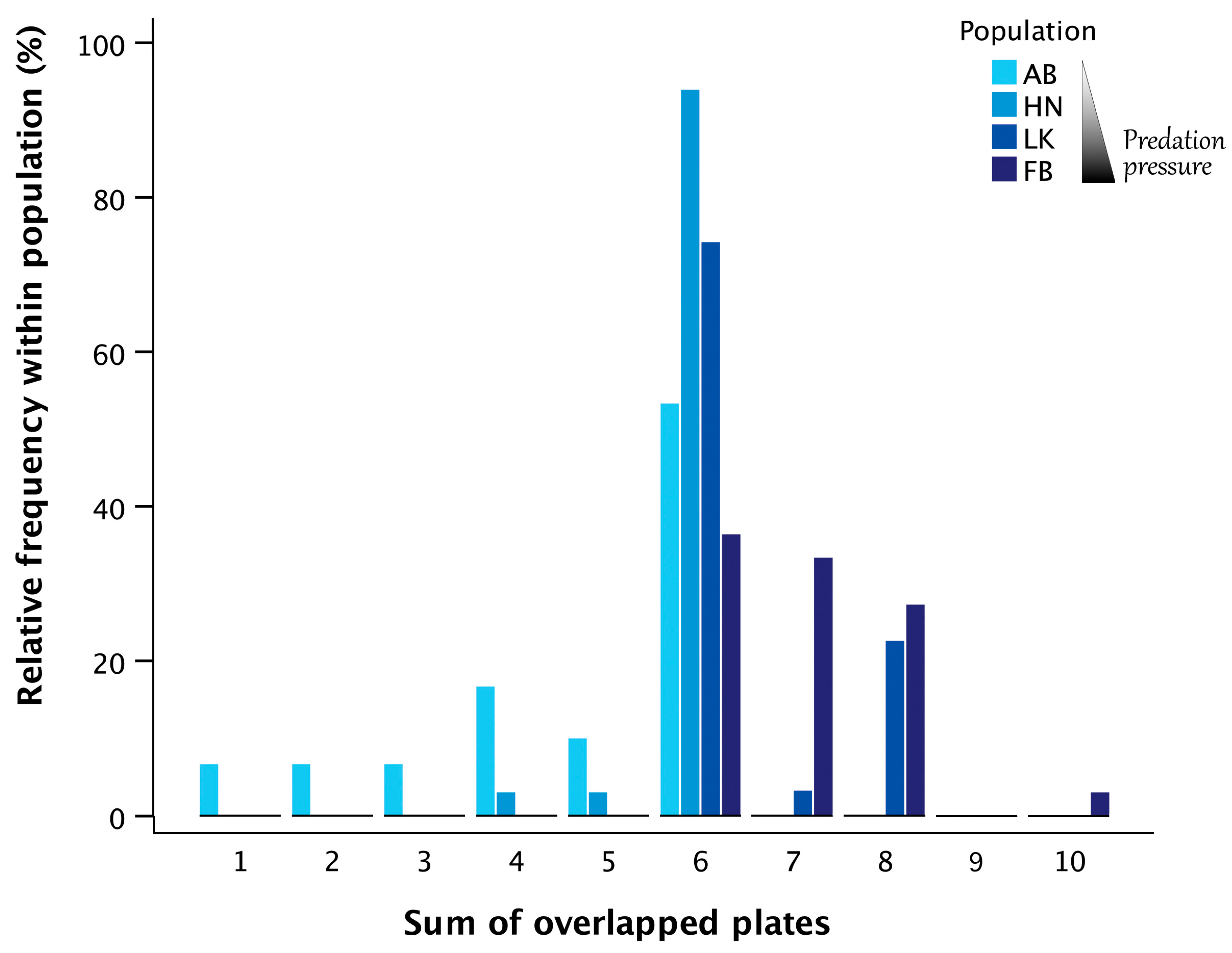
3.3. Symmetry Analyses of the Structural Plates of the Central Defensive Complex (CDC)
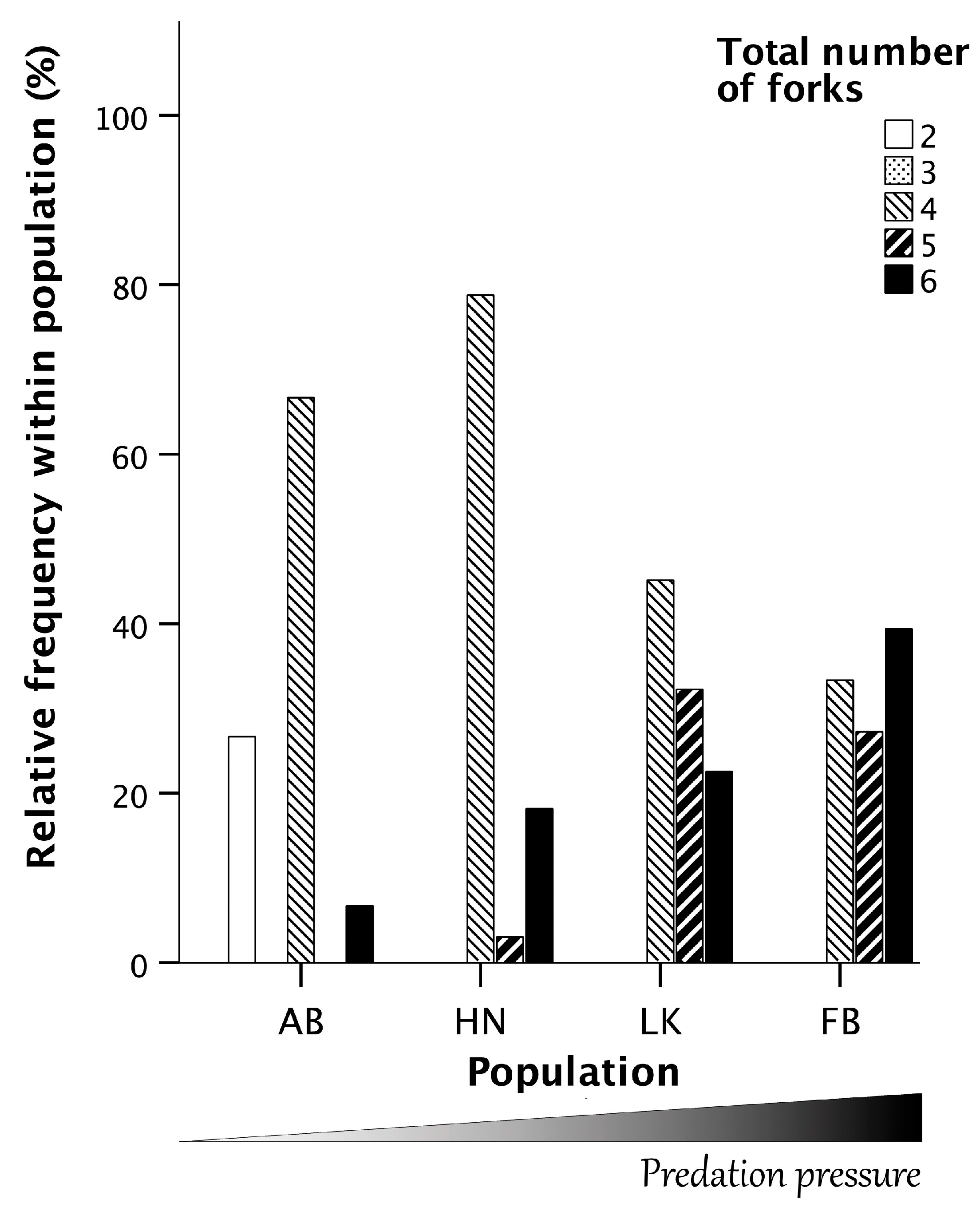
3.4. Symmetry Analyses of the Forks of the Ascending Branch of the Pelvis
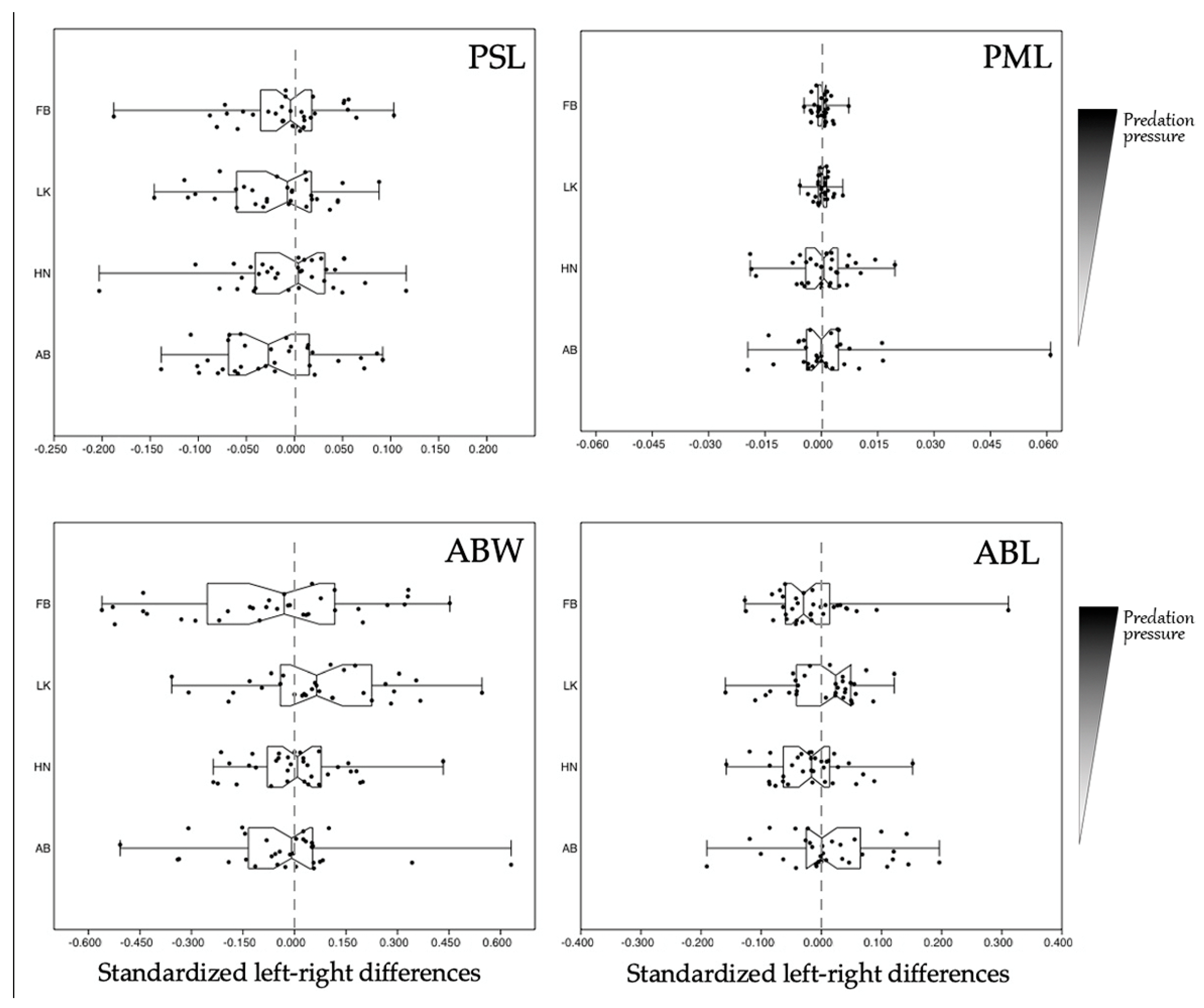
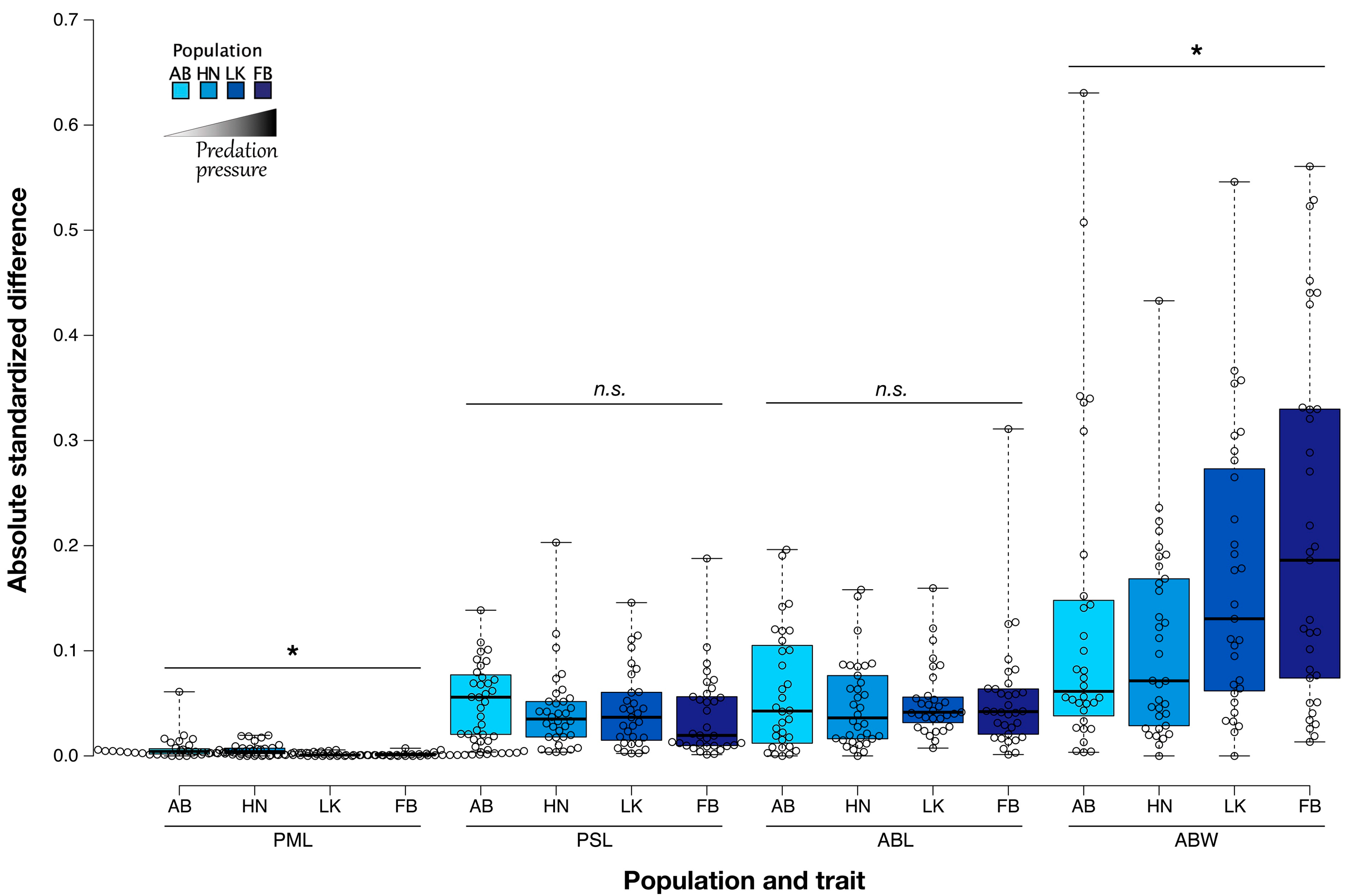
3.5. Pelvis Length
3.6. Pelvic Spine Length
3.7. Fish Size
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (I) Population | (J) Pop. | Mean Difference (I–J) | Std. Error | Sig. | 95% Confidence Interval | |
|---|---|---|---|---|---|---|
| Lower Bound | Upper Bound | |||||
| FB | LK | 1.1 | 0.11 | <0.001 | 0.78 | 1.37 |
| HN | 2.2 | 0.12 | <0.001 | 1.86 | 2.49 | |
| AB | 3.4 | 0.21 | <0.001 | 2.84 | 3.99 | |
| LK | FB | −1.1 | 0.11 | <0.001 | −1.37 | −0.78 |
| HN | 1.1 | 0.11 | <0.001 | 0.79 | 1.40 | |
| AB | 2.3 | 0.21 | <0.001 | 1.77 | 2.91 | |
| HN | FB | −2.2 | 0.12 | <0.001 | −2.49 | −1.86 |
| LK | −1.1 | 0.11 | <0.001 | −1.40 | −0.79 | |
| AB | 1.2 | 0.21 | <0.001 | 0.66 | 1.82 | |
| AB | FB | −3.4 | 0.21 | <0.001 | −3.99 | −2.84 |
| LK | −2.3 | 0.21 | <0.001 | −2.91 | −1.77 | |
| HN | −1.2 | 0.21 | <0.001 | −1.82 | −0.66 | |
| (I) Population | (J) Pop. | Mean Difference (I–J) | Std. Error | Sig. | 95% Confidence Interval | |
| Lower Bound | Upper Bound | |||||
| FB | LK | 1.0 | 0.11 | <0.001 | 0.69 | 1.28 |
| HN | 2.1 | 0.12 | <0.001 | 1.79 | 2.44 | |
| AB | 3.4 | 0.16 | <0.001 | 2.94 | 3.83 | |
| LK | FB | −1.0 | 0.11 | <0.001 | −1.28 | −0.69 |
| HN | 1.1 | 0.12 | <0.001 | 0.80 | 1.46 | |
| AB | 2.4 | 0.17 | <0.001 | 1.95 | 2.85 | |
| HN | FB | −2.1 | 0.12 | <0.001 | −2.44 | −1.79 |
| LK | −1.1 | 0.12 | <0.001 | −1.46 | −0.80 | |
| AB | 1. 3 | 0.17 | <0.001 | 0.80 | 1.75 | |
| AB | FB | −3.4 | 0.16 | <0.001 | −3.83 | −2.94 |
| LK | −2.4 | 0.17 | <0.001 | −2.85 | −1.95 | |
| HN | −1. 3 | 0.17 | <0.001 | −1.75 | −0.80 | |
Appendix B
| Population | Left–Right Difference | |||
| −1 | 0 | 1 | ||
| AB | Count | 5 | 23 | 2 |
| % within population | 16.7% | 76.7% | 6.7% | |
| HN | Count | 0 | 32 | 1 |
| % within population | 0.0% | 97.0% | 3.0% | |
| LK | Count | 0 | 30 | 1 |
| % within population | 0.0% | 96.8% | 3.2% | |
| FB | Count | 7 | 22 | 4 |
| % within population | 21.2% | 66.7% | 12.1% | |
Appendix C
| Left–Right Difference: Forks | Total | |||||
| −1 | 0 | 1 | ||||
| Population | AB | Count | 0 | 30 | 0 | 30 |
| % within population | 0.0 | 100.0 | 0.0 | |||
| HN | Count | 0 | 32 | 1 | 33 | |
| % within population | 0.0 | 97.0 | 3.0 | |||
| LK | Count | 3 | 21 | 7 | 31 | |
| % within population | 9.7 | 67.7 | 22.6 | |||
| FB | Count | 3 | 24 | 6 | 33 | |
| % within population | 9.1 | 72.7 | 18.2 | |||
| Total | Count | 6 | 107 | 14 | 127 | |
| % | 4.7 | 84.3 | 11.0 | 100.0 | ||
References
- Palmer, A.R.; Strobeck, C. Fluctuating asymmetry as a measure of developmental stability: Implications of non-normal distributions and power of statistical tests. Acta Zool. Fennica 1992, 191, 57–72. [Google Scholar]
- Van Dongen, S. Fluctuating asymmetry and developmental instability in evolutionary biology: Past, present and future. European Soc. Evol. Biol. 2006, 19, 1727–1743. [Google Scholar] [CrossRef] [PubMed]
- Kenney, L.A.; von Hippel, F.A. Morphological asymmetry of insular freshwater populations of threespine stickleback. Environ. Biol. Fish. 2014, 97, 225–232. [Google Scholar] [CrossRef]
- Van Valen, L. A study of fluctuating asymmetry. Evolution 1962, 16, 125–142. [Google Scholar] [CrossRef]
- Palmer, A.R. Symmetry breaking and the evolution of development. Science 2004, 306, 828–833. [Google Scholar] [CrossRef] [Green Version]
- Martin, J.; López, P. Hindlimb asymmetry reduces escape performance in the lizard Psammodromus algirus. Physiol. Biochem. Zool. 2001, 74, 619–624. [Google Scholar] [CrossRef]
- Bergstrom, C.A.; Reimchen, T.E. Asymmetry in structural defenses: Insights into selective predation in the wild. Evolution 2003, 57, 2128–2138. [Google Scholar]
- Galeotti, P.; Sacchi, R.; Vicario, V. Fluctuating asymmetry in body traits increases predation risks: Tawny owl selection against asymmetric woodmice. Evol. Ecol. 2005, 19, 405–418. [Google Scholar] [CrossRef]
- Møller, A.P. Developmental stability and fitness: A review. Am. Nat. 1997, 149, 916–932. [Google Scholar] [CrossRef]
- Møller, A.P. Asymmetry as a predictor of growth, fecundity and survival. Ecol. Lett. 1999, 2, 149–156. [Google Scholar] [CrossRef]
- Lens, L.; van Dongen, S.; Kark, S.; Matthysen, E. Fluctuating asymmetry as an indicator of fitness: Can we bridge the gap between studies? Biol. Rev. 2002, 77, 27–38. [Google Scholar] [CrossRef] [Green Version]
- Trokovic, N.; Herczeg, G.; Ab Ghani, N.I.; Shikano, T.; Merilä, J. High levels of fluctuating asymmetry in isolated stickleback populations. BMC Evol. Biol. 2012, 12, 115. [Google Scholar] [CrossRef] [Green Version]
- Gummer, D.L.; Brigham, R.M. Does fluctuating asymmetry reflect the importance of traits in little brown bats (Myotis lucifugus)? Can. Zool. 1995, 73, 990–992. [Google Scholar] [CrossRef]
- Karvonen, E.; Merilä, J.; Rintamäki, P.T.; van Dongen, S. Geography of fluctuating asymmetry in the greenfinch, Carduelis chloris. Oikos 2003, 100, 507–516. [Google Scholar] [CrossRef]
- Reimchen, T.E.; Bergstrom, C.A. The ecology of asymmetry in stickleback defense structures. Evolution 2009, 63, 115–126. [Google Scholar] [CrossRef]
- Wootton, R.J. The Darwinian stickleback Gasterosteus aculeatus: A history of evolutionary studies. J. Fish. Biol. 2009, 75, 1919–1942. [Google Scholar] [CrossRef]
- Campbell, R.N. Morphological variation in the three-spined stickleback (Gasterosteus aculeatus) in Scotland. Behaviour 1985, 93, 161–168. [Google Scholar] [CrossRef] [Green Version]
- Bell, M.A.; Foster, S.A. The Evolutionary Biology of the Threespine Stickleback; Oxford University Press: Oxford, UK, 1994. [Google Scholar]
- Kingsley, D.M.; Zhu, B.; Osoegawa, K.; De Jong, P.J.; Schein, J.; Marra, M.; Peichel, C.; Amemiya, C.; Schluter, D.; Balhabhadra, S.; et al. New genomic tools for molecular studies of evolutionary change in threespine sticklebacks. Behaviour 2004, 141, 1331–1344. [Google Scholar]
- Raeymaekers, J.A.M.; Maes, G.E.; Audenaert, E.; Volckaert, F.A.M. Detecting Holocene divergence in the anadromous-freshwater three-spined stickleback (Gasterosteus aculeatus) system. Mol. Ecol. 2005, 14, 1001–1014. [Google Scholar] [CrossRef]
- Song, J.; Reichert, S.; Kalli, I.; Gazit, D.; Wund, M.; Boyce, M.C.; Ortiz, C. Quantitative microstructural studies of armor of the marine threespine stickleback (Gasterosteus aculeatus). J. Struct. Biol. 2010, 171, 318–331. [Google Scholar] [CrossRef]
- Defaveri, J.; Merilä, J. Evidence for adaptive phenotypic differentiation in Baltic Sea sticklebacks. J. Evol. Biol. 2013, 26, 1700–1715. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.; Zieba, G.; Spence, R.; Klepaker, T.; Przybylski, M. Three-spined stickleback armour predicted by body size, minimum winter temperature and pH. J. Zool. 2020, 311, 13–22. [Google Scholar] [CrossRef]
- Marchinko, K.B.; Schluter, D. Parallel evolution by correlated response: Lateral plate reduction in threespine stickleback. Evolution 2007, 61, 1084–1090. [Google Scholar] [CrossRef]
- Cresko, W.C. Armor development and fitness. Science 2008, 322, 204–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, E.B.; McPhail, J.D. Prolonged and burst swimming in anadromous and freshwater threespine stickleback, Gasterosteus aculeatus. Can. J. Zool. 1986, 64, 416–420. [Google Scholar] [CrossRef]
- Walker, J.A. Ecological morphology of lacustrine threespine stickleback Gasterosteus aculeatus L. (Gasterosteidae) body shape. Biol. J. Linn. Soc. 1997, 61, 3–50. [Google Scholar] [CrossRef] [Green Version]
- Bergstrom, C.A. Fast-start swimming performance and reduction in lateral plate number in threespine stickleback. Can. J. Zool. 2002, 80, 207–213. [Google Scholar] [CrossRef]
- Moodie, G.E.E.; Reimchen, T.E. Phenetic variation and habitat differences in Gasterosteus populations of the Queen Charlotte islands. Syst. Zool. 1976, 25, 49–61. [Google Scholar] [CrossRef]
- Gross, H.P. Natural selection by predators on the defensive apparatus of the three-spined stickleback, Gasterosteus aculeatus L. Can. J. Zool. 1978, 56, 398–413. [Google Scholar] [CrossRef]
- Reimchen, T.E. Structural relationships between spines and lateral plates in threespine stickleback (Gasterosteus aculeatus). Evolution 1983, 37, 931–946. [Google Scholar]
- Reimchen, T.E. Predators and morphological evolution in threespine stickleback. In The Evolutionary Biology of the Threespine Stickleback; Bell, M.A., Foster, S.A., Eds.; Oxford University Press: Oxford, UK, 1994; pp. 240–276. [Google Scholar]
- Ahnelt, H.; Pohl, H.; Hilgers, H.; Splechtna, H. The threespine stickleback in Austria (Gasterosteus aculeatus L., Pisces: Gasterosteidae) morphological variations. Ann. Naturhist. Mus. Wien 1998, 100B, 395–404. [Google Scholar]
- Bell, M.A.; Aguirre, W.E.; Buck, N.J. Twelve years of contemporary armor evolution in a threespine stickleback population. Evolution 2004, 58, 814–824. [Google Scholar]
- Francis, R.C.; Havens, A.C.; Bell, M.A. Unusual lateral plate variation of threespine sticklebacks (Gasterosteus aculeatus) from Knik Lake, Alaska. Copeia 1985, 1985, 619–624. [Google Scholar] [CrossRef]
- Reimchen, T.E.; Nosil, P. Lateral plate asymmetry, diet and parasitism in threespine stickleback. J. Evol. Biol. 2001, 14, 632–645. [Google Scholar] [CrossRef]
- Loehr, J.; Leinonen, T.; Herczeg, G.; O’Hara, R.B.; Merilä, J. Heritability of asymmetry and lateral plate number in the threespine stickleback. PLoS ONE 2012, 7, e39843. [Google Scholar] [CrossRef]
- Bergstrom, C.A.; Reimchen, T.E. Geographical variation in asymmetry in Gasterosteus aculeatus. Biol. J. Linn. Soc. 2002, 77, 9–22. [Google Scholar] [CrossRef] [Green Version]
- Hobson, E.S. Interactions between piscivorous fishes and their prey. In Predator-Prey Systems in Fisheries Management; Clepper, H., Ed.; Sport Fishing Institute: Washington, DC, USA, 1979; pp. 231–242. [Google Scholar]
- Sturat-Smith, R.D.; Richardson, A.M.M.; White, R.W.G. Increasing turbidity significantly alters the diet of brown trout: A multi-year longitudinal study. J. Fish Biol. 2004, 65, 376–388. [Google Scholar] [CrossRef]
- Turesson, H.; Brönmark, C. Predator-prey encounter rates in freshwater piscivores: Effects of prey density and water transparency. Oecologia 2007, 153, 281–290. [Google Scholar] [CrossRef]
- Kingsolver, J.G.; Hoekstra, H.E.; Hoekstra, J.M.; Berrigan, D.; Vignieri, S.N.; Hill, C.E.; Hoang, A.; Gilbert, P.; Berli, P. The strength of phenotypic selection in natural populations. Am. Nat. 2001, 157, 246–261. [Google Scholar] [CrossRef]
- Nosil, P.; Crespi, B.J. Experimental evidence that predation promotes divergence in adaptive radiation. Proc. Natl. Acad. Sci. USA 2006, 103, 9090–9095. [Google Scholar] [CrossRef] [Green Version]
- Ishikawa, M.; Kase, T.; Tsutsui, H. Deciphering deterministic factors of predation pressures in deep time. Sci. Rep. 2018, 8, 17532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bronmark, C.; Miner, J.G. Predator-induced phenotypical change in body morphology in Crucian carp. Science 1992, 258, 1348–1350. [Google Scholar] [CrossRef] [PubMed]
- Januszkiewicz, A.J.; Robinson, B.W. Divergent walleye (Sander vitreus)-mediated inducible defenses in the centrarchid sunfish (Lepomis gibbosus). Biol. J. Linn. Soc. 2007, 90, 25–36. [Google Scholar] [CrossRef]
- Persson, L.; Andersson, J.; Wahlstrom, E.; Eklov, P. Size-specific interactions in lake systems: Predator gape limitation and prey growth rate and mortality. Ecology 1996, 77, 900–911. [Google Scholar] [CrossRef]
- Baumgartner, J.V. Spatial variation of morphology in a freshwater population of thethreespine stickleback, Gasterosteus aculeatus. Can. J. Zool. 1992, 70, 1140–1148. [Google Scholar] [CrossRef]
- Ahnelt, H.; Pohl, H.; Milković, N.; Hilgers, H. Phenotypic diversity in the threespine stickleback Gasterosteus aculeatus Linnaeus 1758 (Teleostei: Gasterosteidae) in western Austria the four-spined form. Ann. Des Nat. Mus. Wien 2006, 107B, 25–38. [Google Scholar]
- Hoogland, R.; Morris, D.; Tinbergen, N. The spines of sticklebacks (Gasterosteus and Pygosteus) as means of defense against predators (Perca and Esox). Behaviour 1956, 10, 205–236. [Google Scholar]
- Bell, M.A. Directional asymmetry of pelvic vestiges in threespine sticklebacks. J. Exper. Zool. (Mol. Dev. Evol.) 2007, 308B, 189–199. [Google Scholar] [CrossRef]
- Vamosi, S.M. Predation sharpens the adaptive peaks: Survival trade-offs in sympatric sticklebacks. Ann. Zool. Fenn. 2002, 39, 237–248. [Google Scholar]
- Marchinko, K.B. Predation’s role in repeated phenotypic and genetic divergence of armor in threespine stickleback. Evolution 2009, 63, 127–138. [Google Scholar] [CrossRef]
- Miller, S.E.; Metcalf, D.; Schluter, D. Intraguild predation leads to genetically based character shifts in the threespine stickleback. Evolution 2015, 69, 3194–3203. [Google Scholar] [CrossRef]
- Bell, M.A. Bridging the gap between population biology and palaeobiology in stickleback fishes. Trends. Ecol. Evol. 1988, 3, 320–325. [Google Scholar] [CrossRef]
- Bowne, P.S. Systematics and morphology of the Gasterosteiformes. In The Evolutionary Biology of the Threespine Stickleback; Bell, M.A., Foster, S.A., Eds.; Oxford University Press: Oxford, UK, 1994. [Google Scholar]
- Vamosi, S.M.; Schluter, D. Character shifts in the defensive armor of sympatric sticklebacks. Evolution 2004, 58, 376–385. [Google Scholar]
- Bell, M.A. Reduction and loss of the pelvic girdle in Gasterosteus (Pisces). A case ofparallel evolution. Nat. Hist. Mus. Los Angeles Co. Contrib. Sci. 1974, 257, 1–36. [Google Scholar]
- Bergstrom, C.A.; Reimchen, T.E. Functional implications of fluctuating asymmetry among endemic populations of Gasterosteus aculeatus. Behaviour 2000, 137, 1097–1112. [Google Scholar]
- Reimchen, T.E. Predator handling failures of lateral plate morphs in Gasterosteus aculeatus: Functional implications for the ancestral plate condition. Behaviour 2000, 137, 1081–1096. [Google Scholar] [CrossRef] [Green Version]
- Klepaker, T.; Østbye, K.; Bell, M.A. Regressive evolution of the pelvic complex in stickleback fishes: A study of convergent evolution. Evol. Ecol. Res. 2013, 15, 413–435. [Google Scholar]
- Cresko, W.C.; McGuigan, K.L.; Phillips, P.C.; Postlethwait, J.H. Studies of threespine stickleback developmental evolution: Progress and promise. Genetica 2007, 129, 105–126. [Google Scholar] [CrossRef]
- Xie, K.T.; Wang, G.; Thompson, A.C.; Wucherpfennig, J.L.; Reimchen, T.E.; MacColl, A.D.C.; Schluter, D.; Bell, M.A.; Vasquez, K.M.; Kingsley, D.M. DNA fragility in the parallel evolution of pelvic reduction in stickleback fish. Science 2019, 362, 81–84. [Google Scholar] [CrossRef] [Green Version]
- Giles, N. The possible role of environmental calcium levels during the evolution of phenotypic diversity in Outer Hebridean populations of the Three-spined stickleback, Gasterosteus aculeatus. J. Zool. 1983, 199, 535–544. [Google Scholar] [CrossRef]
- Barrett, R.D.H. Adaptive evolution of lateral plates in three-spined stickleback Gasterosteus aculeatus: A case study in functional analysis of natural variation. J. Fish Biol. 2010, 77, 311–328. [Google Scholar] [CrossRef] [PubMed]
- Spence, R.; Wootton, R.J.; Barber, I.; Przybylski, M.; Smith, C. Ecological causes of morphological evolution in the three-spined stickleback. Ecol. Evol. 2013, 3, 1717–1726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wassermann, B.A.; Paccard, A.; Apgar, T.M.; Des Roches, S.; Barrett, R.D.H.; Hendry, A.P.; Palkovacs, E.P. Ecosystem size shapes antipredator trait evolution in estuarine threespine stickleback. Oikos 2020, 129, 1795–1806. [Google Scholar] [CrossRef]
- Vamosi, S.M. The presence of other fish species affects speciation in threespine sticklebacks. Evol. Ecol. Res. 2003, 5, 717–730. [Google Scholar]
- Le Rouzic, A.; Østbye, K.; Klepaker, T.O.; Hansen, T.F.; Bernatchez, L.; Schluter, D.; Vøllestad, A. Strong and consistent natural selection associated with armour reduction in sticklebacks. Mol. Ecol. 2011, 20, 2483–2493. [Google Scholar] [CrossRef]
- Ahnelt, H. Zum Vorkommen des Dreistachligen Stichlings (Gasterosteus aculeatus, Pisces: Gastrosteidae) im österreichischen Donauraum. Ann. Naturhist. Mus. Wien 1986, 88/89B, 309–314. [Google Scholar]
- Ahnelt, H.; Ramler, D.; Madsen, M.Ø.; Jensen, L.F.; Windhager, S. Diversity and sexual dimorphism in the head lateral line system in North Sea populations of threespine sticklebacks, Gasterosteus aculeatus (Teleostei: Gasterosteidae). Zoomorphology 2021, 140, 103–117. [Google Scholar] [CrossRef]
- Zhang, J.-D.; Sung, H.J.; Huang, W.-X. Specialization of tuna: A numerical study on the function of caudal keels. Phys. Fluids 2020, 32, 111902. [Google Scholar] [CrossRef]
- Walker, J.A. Dynamics of pectoral fin rowing in a fish with an extreme rowing stroke: The threespine stickleback (Gasterosteus aculeatus). J. Experiment. Biol. 2004, 207, 1925–1939. [Google Scholar] [CrossRef] [Green Version]
- Hagen, D.W.; Moodie, G.E.E. Polymorphism for plate morphs in Gasterosteus aculeatus on the east coast of Canada and a hypothesis for their global distribution. Can. J. Zool. 1982, 60, 1032–1042. [Google Scholar] [CrossRef]
- Bell, M.A. Lateral plate evolution in threespine stickleback: Getting nowhere fast. Genetica 2001, 112–113, 445–461. [Google Scholar] [CrossRef]
- Herler, J.; Lipej, L.; Makovec, T. A simple technique for digital imaging of live and preserved small fish specimens. Cybium 2007, 31, 39–44. [Google Scholar]
- Rohlf, F.J. The tps series of software. Hystrix 2015, 26, 9–12. [Google Scholar] [CrossRef]
- Field, A. Discovering Statistics Using IBM SPSS Statistics, 4th ed.; Sage: Los Angeles, CA, USA; London, UK; New Dehli, India, 2013. [Google Scholar]
- Feltz, C.J.; Miller, G.E. An asymptotic test for the equality of coefficients of variation from k populations. Stat. Med. 1996, 15, 646–658. [Google Scholar] [CrossRef]
- Marwick, B.; Krishnamoorthy, K. cvequality: Tests for the Equality of Coefficients of Variation from Multiple Groups. R Software Package Version 0.1. 7 January 2019. Available online: https://github.com/benmarwick/cvequality (accessed on 8 August 2019).
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis version. Palaeontol. Electron. 2001, 4, 1–9. Available online: http://palaeo-electronica.org/2001_1/past/issue1_01.htm (accessed on 14 March 2023).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016; Available online: https://www.R-project.org/ (accessed on 14 March 2023).
- Garren, S. Permutation tests for nonparametric statistics using R. Asian Res. J. Math. 2017, 5, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Bookstein, F. Morphometric Tools for Landmark Data: Geometry and Biology; Cambridge University Press: Cambridge, UK, 1991. [Google Scholar]
- Slice, D.E. Morpheus et al. Java Edition. [Computer software] Department of Scientific Computing, The Florida State University, Tallahassee, Florida, USA. Available online: http:/morphlab.sc.fsu.edu/ (accessed on 14 March 2023).
- Reimchen, T.E.; Ingram, T.; Hansen, S.C. Assessing niche differences of sex, armour and asymmetry phenotypes using stable isotope analyses in Haida Gwaii sticklebacks. Behaviour 2008, 145, 561–577. [Google Scholar] [CrossRef] [Green Version]
- Bell, M.A. Developmental osteology of the pelvic complex of Gasterosteus aculeatus. Copeia 1985, 1985, 789–792. [Google Scholar] [CrossRef]
- Reimchen, T.E. Spine deficiency and polymorphism in a population of Gasterosteus aculeatus: An adaptation to predators? Can. J. Zool. 1980, 58, 1232–1244. [Google Scholar] [CrossRef]
- Miller, S.E.; Barrueto, M.; Schluter, D. A comparative analysis of experimental selection on the stickleback pelvis. J. Evol. Biol. 2017, 30, 1165–1176. [Google Scholar] [CrossRef] [Green Version]
- Kitano, J.; Bolnick, D.I.; Beauchamp, D.A.; Mazur, M.M.; Mori, S.; Nakano, T.; Peichel, C.L. Reverse evolution of armor plates in the threespine stickleback. Current Biol. 2008, 18, 769–774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banbura, J.; Przybylski, M.; Frankiewicz, P. Selective predation of the pike Esox lucius: Comparison of lateral plates and some metric features of the three-spined stickleback Gasterosteus aculeatus. Zool. Scr. 1989, 18, 303–309. [Google Scholar] [CrossRef]
- Reimchen, T.E. Parasitism of asymmetrical pelvic phenotypes in stickleback. Can. J. Zool. 1997, 75, 2084–2094. [Google Scholar] [CrossRef]
- Mazzi, D.; Künzler, R.; Bakker, T.C.M. Female preference for symmetry in computer-animated three-spined stickleback, Gasterosteus aculeatus. Behav. Ecol. Sociobiol. 2003, 54, 156–161. [Google Scholar] [CrossRef]
- Symons, P.E.K. Analysis of spine-raising in the male three-spined stickleback. Behaviour 1966, 26, 1–75. [Google Scholar] [CrossRef]
- Huntingford, F.A. The relationship between anti-predator behavior and aggression among conspecifics in the three-spined stickleback, Gasterosteus aculeatus. Anim. Behav. 1976, 24, 245–260. [Google Scholar] [CrossRef]
- Reimchen, T.E.; Steeves, D.; Bergstrom, C.A. Sex matters for defense and trophic traits of threespine stickleback. Evol. Ecol. Res. 2016, 17, 459–485. [Google Scholar]
- Klepaker, T.; Østbye, K. Pelvic anti-predator armour reduction in Norwegian populations of the threespine stickleback: A rare phenomenon with adaptive implications? J. Zool. 2008, 276, 81–88. [Google Scholar] [CrossRef]
- Nelson, J.S. Absence of the pelvic complex in ninespine sticklebacks, Pungitius pungitius, collected in Ireland and Wood Buffalo National Park Region, Canada, with notes on meristic variation. Copeia 1971, 1971, 707–717. [Google Scholar] [CrossRef]
- Moodie, G.E.E.; Moodie, P.F. Do asymmetric sticklebacks make better fathers? Proc. R. Soc. Lond. B 1996, 263, 535–539. [Google Scholar]
- MacColl, A.D.C.; El Nagar, A.; de Roil, J. The evolutionary ecology of dwarfism in three-spined sticklebacks. J. Anim. Ecol. 2013, 82, 642–652. [Google Scholar] [CrossRef]
- Ramler, D.; Mitteroecker, P.; Shama, L.N.S.; Wegner, K.M.; Ahnelt, H. Non-linear effects of temperature on body form and on developmental canalization in the threespine stickleback. J. Evol. Biol. 2014, 27, 497–507. [Google Scholar] [CrossRef] [Green Version]
- Swain, M.W. A problem with the use of meristic characters to estimate developmental stability. Am. Nat. 1987, 129, 761–768. [Google Scholar] [CrossRef]
- Young, J.R. Removing bias for fluctuating asymmetry in meristic characters. J. Agric. Biol. Environ. St. 2007, 12, 485–497. [Google Scholar] [CrossRef]
- Burr, P.C.; Samiappan, S.; Hathcock, L.A.; Moorhead, R.J.; Dorr, B.S. Estimating waterbird abundance on catfish aquaculture ponds using an unmanned aerial system. Hum.-Wildl. Interact. 2019, 13, 317–330. [Google Scholar]
- Bakó, G.; Tolnai, M.; Takács, Á. Introduction and testing of a monitoring and colony-mapping method for waterbird populations that uses high-speed and ultra-detailed aerial remote sensing. Sensors 2014, 14, 12828–12846. [Google Scholar] [CrossRef]
- Able, K.W.; Grothues, T.M.; Rackovan, J.L.; Buderman, F.E. Application of mobile dual-frequency identification sonar (DIDSON) on fish in estuarine habitats. Northeast. Nat. 2014, 21, 192–209. [Google Scholar] [CrossRef]
- Jones, R.E.; Griffin, R.A.; Unsworth, R.K.F. Adaptive resolution imaging sonar (ARIS) as a tool for marine fish identification. Fish. Res. 2021, 243, 106092. [Google Scholar] [CrossRef]













| Number of Lateral Plates | Pairwise-Tests | |||||
|---|---|---|---|---|---|---|
| Population | Sample Size | Mean | Standard Deviation | Coefficient of Variation (%) | D’AD | p-Value (Uncorrected) |
| Left side of the fish | ||||||
| AB | 33 | 2.89 | 1.11 | 38.6 | AB–HN: 32.27 | 1.34 × 108 |
| HN | 33 | 4.12 | 0.48 | 11.8 | HN–LK: 4.31 | 0.038 |
| LK | 32 | 5.22 | 0.42 | 8.0 | LK–FB: 0.07 | 0.012 |
| FB | 34 | 6.29 | 0.46 | 7.3 | ||
| Right side of the fish | ||||||
| AB | 33 | 2.85 | 0.83 | 29.3 | AB–HN: 16.72 | 4.34 × 105 |
| HN | 33 | 4.12 | 0.55 | 13.2 | HN–LK: 6.16 | 0.013 |
| LK | 32 | 5.25 | 0.44 | 8.4 | LK–FB: 1.18 | 0.277 |
| FB | 34 | 6.24 | 0.43 | 6.9 | ||
| Population | Trait | t | df | p |
|---|---|---|---|---|
| AB (n = 32) | Pelvic spine length (PSL) | −2.4 | 31 | 0.021 |
| Pelvic length (PML) | 0.8 | 31 | 0.459 | |
| Ascending branch width (ABW) | −0.9 | 31 | 0.390 | |
| Ascending branch length (ABL) | 0.9 | 31 | 0.350 | |
| HN (n = 34) | Pelvic spine length (PSL) | −0.8 | 33 | 0.435 |
| Pelvic length (PML) | 0.1 | 33 | 0.889 | |
| Ascending branch width (ABW) | 0.3 | 33 | 0.778 | |
| Ascending branch length (ABL) | −1.8 | 33 | 0.088 | |
| LK (n = 31) | Pelvic spine length (PSL) | −2.2 | 30 | 0.038 |
| Pelvic length (PML) | 0.8 | 30 | 0.428 | |
| Ascending branch width (ABW) | 2.1 | 30 | 0.040 | |
| Ascending branch length (ABL) | 0.3 | 30 | 0.794 | |
| FB (n = 33) | Pelvic spine length (PSL) | −0.9 | 32 | 0.359 |
| Pelvic length (PML) | 0.6 | 32 | 0.576 | |
| Ascending branch width (ABW) | −1.3 | 32 | 0.205 | |
| Ascending branch length (ABL) | −1.2 | 32 | 0.255 |
| PML | ABW | |||||||
|---|---|---|---|---|---|---|---|---|
| Population | AB | HN | LK | FB | AB | HN | LK | FB |
| AB | ----- | 0.855 | 1 × 10−5 a | 2 × 10−5 b | ----- | 0.955 | 0.133 | 0.030 |
| HN | 0.321 | ----- | 4 × 10−5 c | 3 × 10−5 d | 0.690 | ----- | 0.176 | 0.010 |
| LK | 0.695 | 0.912 | ----- | 0.964 | 0.729 | 0.139 | ----- | 0.270 |
| FB | 0.130 | 0.488 | 0.469 | ----- | 0.639 | 0.724 | 0.248 | ----- |
| PSL | ABL | |||||||
| Population Difference | Test Statistic | Std. Error | Std. Test Statistic | Sig. | Adj. Sig. a |
|---|---|---|---|---|---|
| AB–HN | −20.342 | 7.921 | −2.568 | 0.010 | 0.061 |
| AB–LK | −38.390 | 8.042 | −4.774 | <0.001 | <0.001 |
| AB–FB | −57.509 | 7.921 | −7.260 | <0.001 | <0.001 |
| HN–LK | −18.048 | 7.854 | −2.298 | 0.022 | 0.129 |
| HN–FB | −37.167 | 7.730 | −4.808 | <0.001 | <0.001 |
| LK–FB | −19.119 | 7.854 | −2.434 | 0.015 | 0.090 |
| Total Number of Forks | Total | |||||
|---|---|---|---|---|---|---|
| 4 | 5 | 6 | ||||
| Population | AB | Count | 20 | 0 | 2 | 22 |
| Expected count | 13.1 | 3.7 | 5.2 | |||
| % within population | 90.9 | 0.0 | 9.1 | |||
| Std. residual | 1.9 | −1.9 | −1.4 | |||
| HN | Count | 26 | 1 | 6 | 33 | |
| Expected count | 19.7 | 5.5 | 7.8 | |||
| % within population | 78.8 | 3.0 | 18.2 | |||
| Std. residual | 1.4 | −1.9 | −0.6 | |||
| LK | Count | 14 | 10 | 7 | 31 | |
| Expected count | 18.5 | 5.2 | 7.3 | |||
| % within population | 45.2 | 32.3 | 22.6 | |||
| Std. residual | −1.0 | 2.1 | −0.1 | |||
| FB | Count | 11 | 9 | 13 | 33 | |
| Expected count | 19.7 | 5.5 | 7.8 | |||
| % within population | 33.3 | 27.3 | 39.4 | |||
| Std. residual | −2.0 | 1.5 | 1.9 | |||
| Total | Count | 71 | 20 | 28 | 119 | |
| Descriptive Statistics (%) | Pairwise-Tests | |||||
|---|---|---|---|---|---|---|
| Population | Sample Size | Mean | Standard Deviation | Coefficient of Variation | D’AD | p-Value (Uncorrected) |
| Pelvic length/standard length | ||||||
| AB | 32 | 18.22 | 1.99 | 10.9 | AB–HN: 4.05 | 0.044 |
| HN | 34 | 20.42 | 1.56 | 11.8 | HN–LK: 4.58 | 0.032 |
| LK | 31 | 24.18 | 1.25 | 7.6 | LK–FB: 4.30 | 0.038 |
| FB | 33 | 24.48 | 1.84 | 7.5 | ||
| Pelvic spine length/standard length | ||||||
| AB | 32 | 11.63 | 1.37 | 11.8 | Overall test n.s. | |
| HN | 34 | 13.80 | 1.40 | 10.2 | ||
| LK | 31 | 15.42 | 1.29 | 8.4 | ||
| FB | 33 | 14.85 | 1.57 | 10.6 | ||
| Population | Percentiles SL (in mm) | n | Predation | ||
|---|---|---|---|---|---|
| 25 | 50 | 75 | Pressure | ||
| AB | 43.2 | 45.1 | 48.4 | 32 | low |
| HN | 36.7 | 37.4 | 38.3 | 34 | moderate |
| LK | 47.9 | 49.8 | 51.8 | 31 | high |
| FB | 51.4 | 57.2 | 60.8 | 33 | very high |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schröder, M.; Windhager, S.; Schaefer, K.; Ahnelt, H. Adaptability of Bony Armor Elements of the Threespine Stickleback Gasterosteus aculeatus (Teleostei: Gasterosteidae): Ecological and Evolutionary Insights from Symmetry Analyses. Symmetry 2023, 15, 811. https://doi.org/10.3390/sym15040811
Schröder M, Windhager S, Schaefer K, Ahnelt H. Adaptability of Bony Armor Elements of the Threespine Stickleback Gasterosteus aculeatus (Teleostei: Gasterosteidae): Ecological and Evolutionary Insights from Symmetry Analyses. Symmetry. 2023; 15(4):811. https://doi.org/10.3390/sym15040811
Chicago/Turabian StyleSchröder, Margarethe, Sonja Windhager, Katrin Schaefer, and Harald Ahnelt. 2023. "Adaptability of Bony Armor Elements of the Threespine Stickleback Gasterosteus aculeatus (Teleostei: Gasterosteidae): Ecological and Evolutionary Insights from Symmetry Analyses" Symmetry 15, no. 4: 811. https://doi.org/10.3390/sym15040811