
Chemical Models for Understanding the Emergence of Homo-Chirality of Phospholipids for Origin of Life Studies


Institut de Chimie et de Biochimie Moléculaires et Supramoléculaires (UMR 5246), Université Claude Bernard Lyon 1, Université de Lyon, Bât. Edgar Lederer, 1 rue Victor Grignard, CEDEX, F-69622 Villeurbanne, France
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Symmetry 2022, 14(10), 2109; https://doi.org/10.3390/sym14102109
Submission received: 13 September 2022
/
Revised: 3 October 2022
/
Accepted: 8 October 2022
/
Published: 11 October 2022
(This article belongs to the Collection Feature Papers in Chemistry)
Abstract
:In the prebiotic world, the chemical assembly of biotic building blocks led to racemic mixtures; however, homo-chirality emerged in the racemic prebiotic soup. Polymers and other molecules assembled from mixtures of enantiomers rather than racemic ones. Understanding how symmetry breaking happens is one of the most challenging fields of research in origin of life studies. With this article, we aim to shed light on one of the problems: in the absence of physical examples for use in a laboratory scale, what are the best models to use to simulate the conditions and lead to homo-chiral symmetry breaking? In this perspective, we suggest looking to chemical models that can represent a poorly studied class of prebiotic compounds in the context of symmetry breaking: the phospholipids.
1. Introduction
The two forms of a molecule with a chiral center are called enantiomers. Chirality can be defined, for example, in cases such as the mirror image reflection of sp3-hybridized carbon, because this image cannot be superimposed on the original one, and the mirror images are then two different molecules [1]. Chiral molecules in living organisms exist almost exclusively, as single enantiomers, a property that is critical for molecular recognition and replication processes, and would thus seem to be a prerequisite for the threshold of life. Genetic polymers (DNA/RNA) are composed of the same chirality (handedness) and are able to act as templates in the replication of functional polymers (proteins/RNA) and to fold into appropriate structures. Enantiomers can have identical physical and chemical properties, but can variously interact with other chiral molecules [1,2].
1.1. Geochemical Scenarios
The geology and the chemistry of Earth before the advent of life was completely different from what we know today. At that time, sunlight, volcanic heat, and hydrothermal sites were the main energy sources that were able to drive the synthesis of many molecules, including nucleosides, peptides, sugars and amphiphilic compounds. The atmosphere was mostly nitrogen (N2), as today, with a substantial amount of carbon dioxide (CO2) and much smaller amounts of carbon monoxide, ammonia, and methane (CO, NH3, CH4). It is also likely that water, present in locally limited amounts, contained hydrogen cyanide (HCN), formaldehyde (HCHO) and formamide (HCONH2) [3]. Intriguingly, those molecules are found in interstellar space, together with many other that can be considered as building blocks for the assembling of biomolecules such as water (H2O), formic acid (HCOOH), methanol (CH3OH) cyanamide (NH2CN), acetic acid (CH3COOH), acetamide (CH3CONH2), ethylene glycol (HOCH2CH2OH) and glycine [4]. We can imagine that at the beginning of its existence, our planet—and, for instance, this is true for any planet with a similarity with planet Earth and having liquid water-based Life—was an ensemble of active volcanic land with some salty oceans. Fresh water was also present because the evaporation of the water generated rain that, after falling on landscapes, accumulated in depressions. The origin of liquid water on our planet is controversial on several levels. The Deuterium/Hydrogen ratio of the water present in today’s oceans, compared with that present on comets, excludes the possibility that the total amount of this water came from the impact of several comets on the early planet Earth [5]. Water was produced by internal chemical processes in the crust. Evaporation followed by recondensation (rain) delivered to the hot volcanic land masses (cooling them down), and starting to accumulate in lava depressions, forming primordial oceans [6]. Two different and plausible geochemical scenarios have been depicted in the last sixty years. The plausible scenarios have a common leitmotiv: the presence of liquid water, a source of energy (geothermal but also sun-light/UV radiation), and the presence of minerals and organic carbon. These two scenarios are extremely different, and scientist called them hydrothermal vents and hydrothermal fields [7,8]. Hydrothermal vents (HV), also present today at the bottom of the oceans), are systems whose heat source is the underlying magma or hot water generated by convection currents, due to high thermal gradients. HV, also called hydrothermal black smokers or submarine hot springs [9], are alkaline, far-from-equilibrium environments, and until their discoveries they were proposed as sites at which chemical reactions could initiate primitive metabolism involving the reduction of CO2 by dissolved H2. The alternatives to HV are hydrothermal fields (HF), known as hydrothermal pools [7,10,11]. Recently, Damer and Deamer pointed out that fluctuating hydrothermal pools (FHPs) could be considered as plausibly prebiotic reactors for the synthesis of several key molecules for the development of life, including lipids, nucleic acids and peptides. Concerning the origin of life studies domain, more than one model, reproduced in a laboratory scale and where possible in the field or messy environments (see a recent article by David Deamer [12]) have been speculated upon. In addition, it is important not to forget that, as chemical substrates, clays, pyrite and sulfur containing minerals represent the most plausible 2D models for simulating prebiotic reactions, and that eventually substrates can be used for prebiotic symmetry breaking experiments [13,14].
1.2. Homochirality Emergence and Origin of Life Studies
In origin of life studies (OOL), homochirality in biotic molecules is reputed to be one of the signatures of life, and the question raised is how this property emerged. The simulation of prebiotic conditions for the preparation of biotic building blocks shows the formation of molecules in racemic mixtures. Moreover, in a prebiotic chemical world, the small molecules, from which these macromolecules could be synthesized, were plausibly formed as racemic mixtures. Models have been suggested to incorporate chirogenesis into chemical evolution, structured by prebiotic building blocks [15,16]. In 1848, Pasteur discovered the crystallization of sodium ammonium tartrate C4H8NNaO6, and two types of large crystals, visible under microscopic light, were separated manually. Once separated, they showed two different optical activities, rotating the plane of polarized light in two opposite directions. Pasteur correctly interpreted the crystal structures as representing an asymmetry in both molecules, which could exist in left-handed or right-handed forms. Later, Pasteur speculated that understanding the origin of the asymmetry might provide a key to the nature of the emergence of life, and several subsequent discoveries confirmed his speculations [17]. Surprisingly, experimental validation and proposed models for the origin of homochirality in living organisms were not explored for more than half a century. The question that needs to be answered has two parts. The first regards “symmetry breaking”: what could have distorted the production towards one enantiomer? The second concerns “amplification”: given this imbalance of enantiomers, how is it possible to be sustained and generated to establish homochirality in life [18]? One abnormality on natural symmetry breaking that led to an excess of l- over d-amino acids, has been discovered on carbonaceous meteorites [19,20,21,22], with the relative preference of one enantiomer, depending on meteorite origin, whereas glycine has been found in very small amounts in interstellar dust [23]. In addition, several scientists have speculated on how the different abundancies of l- over d-amino acids in interstellar space bodies and congeners have been generated. In a recent review, Burton and Berger summarized the most relevant experiments carried out using five carbon amino acids [24]. In all cases, and to the best of our knowledge, deracemization happens in different conditions: the exposition of samples to circular polarized light (chiral light, ~0.5%), electrical discharges and particle irradiation, whereas a difference in femto- to picomole excess was measured whereas exposed to parity violation energies. However, for a laboratory scale experiment, those experiments are poorly applicable. Despite this downside, we have reported in Table 1 a comparison between data from the first Murchison meteorite analysis and those obtained by a re-analysis of one of Miller’s experiments [25], confirming that prebiotic simulated condition reactions can lead only to racemic mixtures, and concluding that one or a combination of physical events led to the formation of scalemic mixtures of amino acids in space bodies A complete speculation on the presence of scalemic mixtures of amino acids has been recently summarized elsewhere, in Chieffo et al., 2022 (submitted to Life).
However, these data only partly explain the protein’s l-chirality, assuming only that proteins could be made of short polypeptides [31] and that the concentration of l- over d-AA has been enriched only by feedstock from interstellar media. However, the presence of d-amino acids in prebiotic proteins cannot be excluded. The analysis of the venom of some marine organisms such as Conidaae carnivorous, that live close to sub-marine chimneys, showed the presence of d-amino acids in toxic proteins. As reported by Solokov and co-workers [32], this fact cannot exclude the fact that in the early stage of life, mixtures of both type of proteins or a scalemic mixture of l- and d-amino acids, were present in the first forms of protocells [33].
Concerning nucleotides, homochirality is due to the presence of ribose and 2-deoxyribose in RNA and DNA, respectively, both solely in a d-configuration. The phenomenon of symmetry breaking in chiral molecules has lately been pointing towards phospholipids, whereas chirality regards carbon C-2 of the glycerol backbone [34].
2. Autocatalysis and Symmetry Breaking
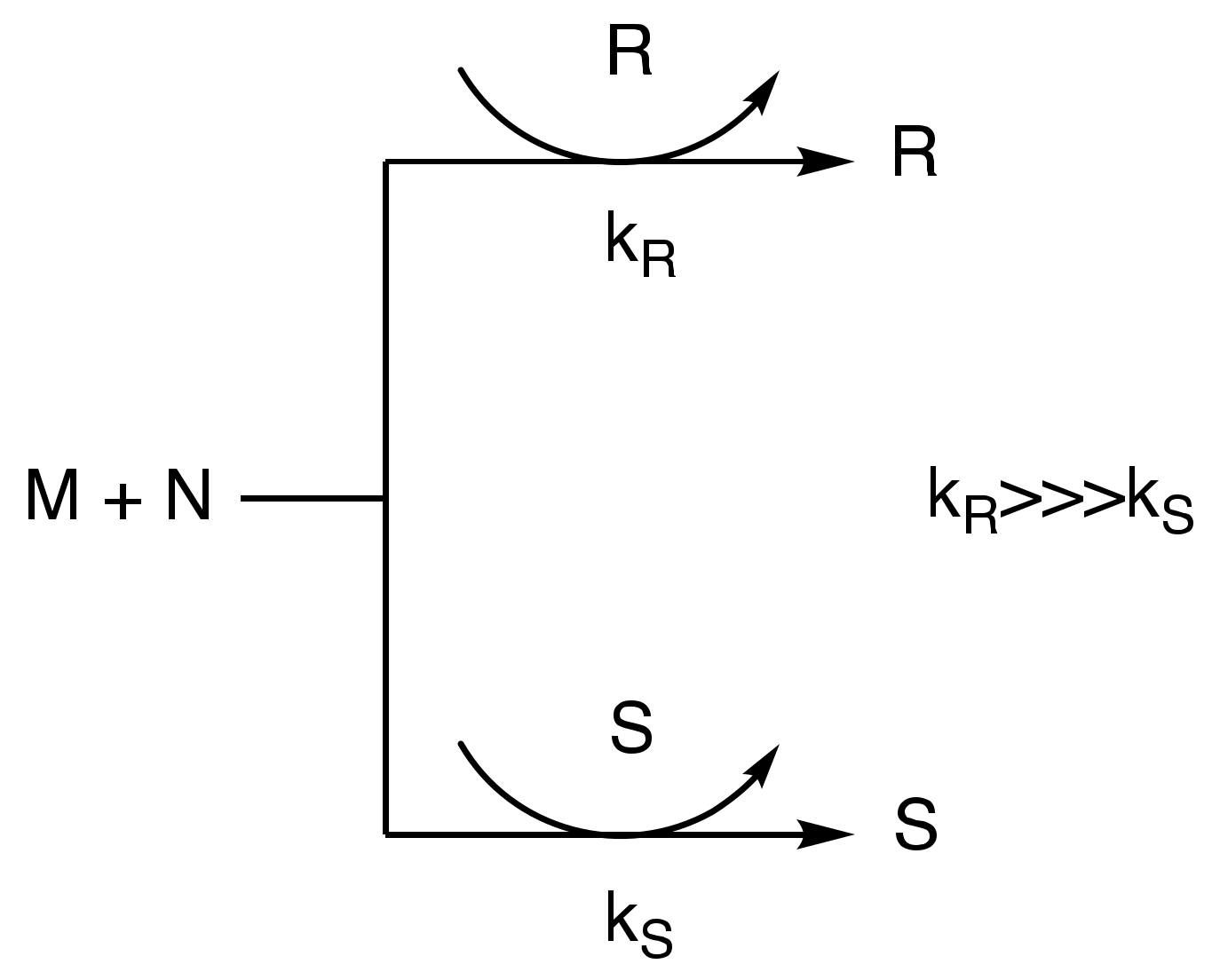
In the early fifties of the last century, Frank proposed a mathematical model for the predominance of homochirality during chemical synthesis. The model is simple: a chiral molecule catalyzes its own formation, suppressing the formation of the opposite enantiomer with, for example, the exclusive formation of an R or S enantiomer for a non-chiral starting material (Figure 1). The model, which is widely accepted today, proposes the amplification of a small initial asymmetry in the chiral reactions, and concludes that it is the key element for this simple scheme of reactions [35].
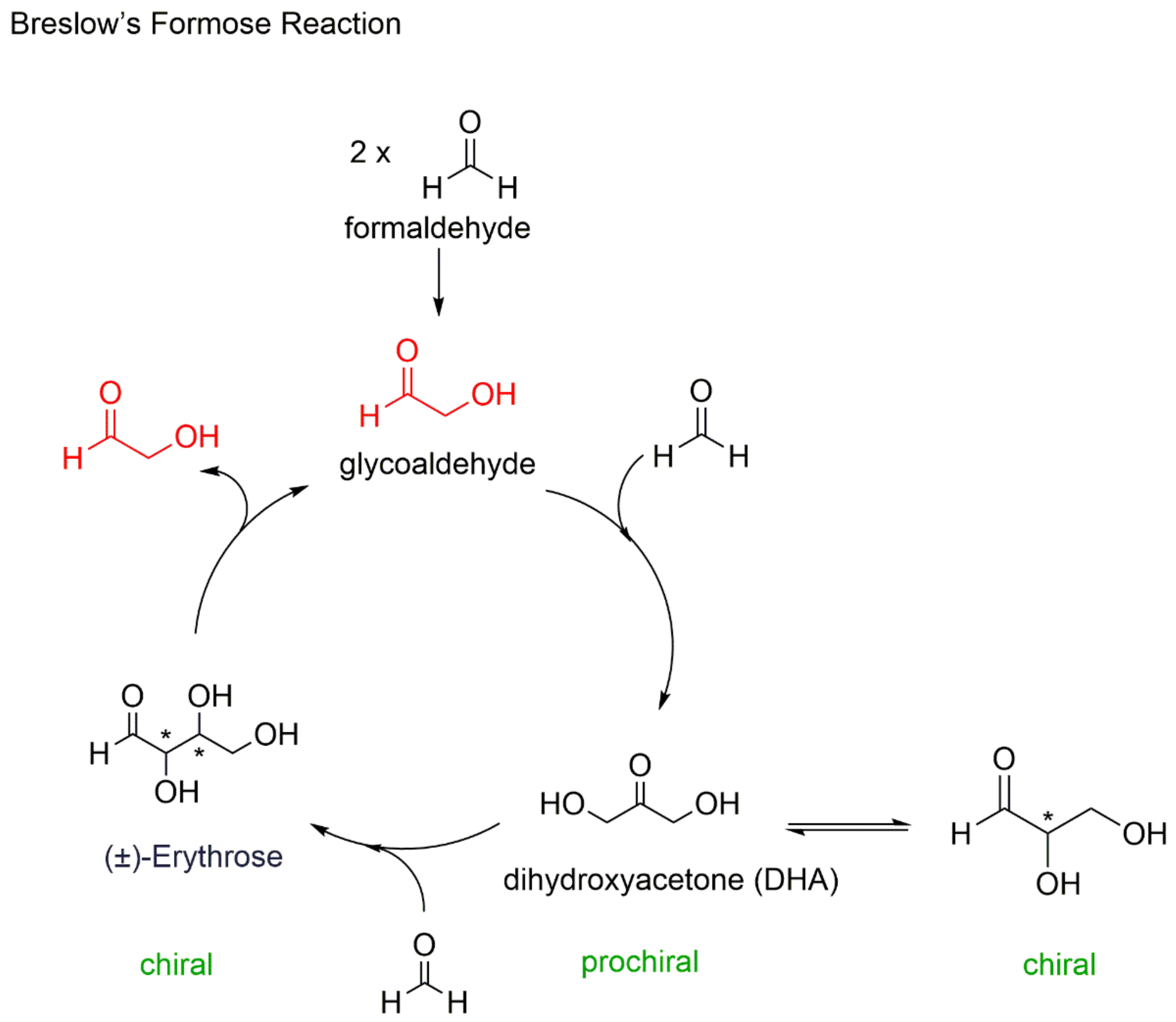
In an autocatalytic system, the catalyst, which is the same species as the reaction product, must be capable of self-replication, but this activity must also be accompanied by suppression of the activity of its enantiomer, in what Frank referred to, in the proposed model, as mutual antagonism. The fundamental ways in which asymmetric autocatalysis exceeds the non-autocatalytic asymmetric catalysis can be mainly described in the three following sentences: (i) There is high efficiency due to the self-replication process. (ii) There is no decrease in the amount and the activity of the catalyst as a result of the analogous increase of the catalytic amount and the product of the reaction. Finally, (iii) separation of the product from the catalyst is not necessary, because of the conformity of the catalyst and product structures [36]. Following the publication of Frank’s mathematical model, organic chemists attempted to associate an autocatalytic reaction system with prebiotic chemistry. However, this model is not applicable to all self-catalytic processes. For example, in Breslow’s formose reaction [37], even though achiral glycoaldehyde catalyzes its own formation, this system does not conform to the suggested autocatalytic model, and therefore will not be associated with the emergence of homochirality (Figure 2), with one exception: the introduction of a catalytic (<1%) amount of l-proline for the formation of d-sugars [38]. Therefore, we are back to square one. Where did the homochiral proline come from in a prebiotic land where chemical synthesis led only to racemic mixtures?
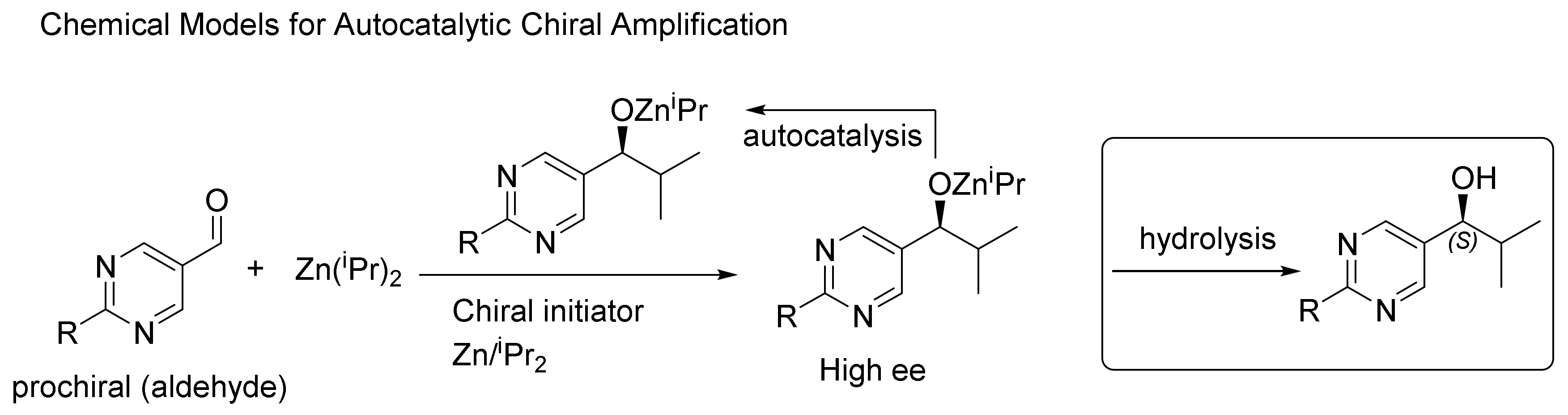
Several groups undertook the description of reactions involving alkyl metal reagents, but the challenge was not met for the next 40 years, until Kenso Soai’s first work was published in 1995. The first experimental proof became known when Soai and colleagues reported the autocatalytic alkylation of pyrimidyl aldehydes with (pyrimidyl) dialkylzincs (cf. Figure 3). The Soai reaction is, up until today, the only fitting example for chiral autocatalysis that follows Frank’s model (Figure 1). The rate of the reaction is not only accelerated by the autocatalytic product which, starting from scalemic mixtures (with a non-equal ratio between two enantiomers), but may ultimately appear in very “high” enantiomeric excess, starting from a very “low” enantiomeric excess of the original autocatalyst (Figure 3). Finally, Soai and colleagues also demonstrated a variety of initiators and directing agents for the product enantiomeric excess in this reaction, including exposure to circularly polarized light [39,40,41]. The Soai alkylation of pyrimidyl aldehydes, which is of interest due to its unique effect of chiral amplification via autocatalysis, requires conditions far from those on primitive Earth, in a prebiotic world. However, studying this reaction serves as a model for understanding how homochirality might have emerged and may help us search for prebiotically plausible reactions with similar mechanisms. In 2019, in a more recent paper, Soai, Kawasaki and Matsumoto employed an asymmetric autocatalysis of amplification of ee% to examine the several proposed mechanisms of the origin of chirality [39]. Subsequently, Denmark and colleagues have added a new model aldehyde (Figure 3, last row, right-hand side) that serves to give more support for the comprehension of the mechanism of the Soai reaction, introducing a “quaternary” complex rather than the “ternary” ones supposed before [42]. This model reaction has been particularly instructive in helping to understand how autocatalysis could lead to a homochiral state. Nonetheless, the discovery of a prebiotically plausible reaction, or a systems reaction from a bottom-up approach that obeys the Frank model of autocatalysis, is yet to be brought to light [7,18,19].
3. Crystallization Led to Homochiral Symmetry Breaking
In 1990, Kondepudi et al. reported an experimental example of such spontaneous chiral symmetry breaking for the stirred crystallization of a homogeneous solution of achiral NaClO3. Symmetry breaking in this case appears in a simple way: if NaClO3 is crystallized from an unstirred solution, statistically equal numbers of l and d crystals are obtained. If the crystallization is performed accompanied with rapid stirring, more than 99% are obtained with the same chirality. Viedma et al., in 2003, described in their report an experiment to explore the phenomenon. They reported that in primary nucleation, crystals occur in the solution itself. On the other hand, secondary nucleation is the process by which a “mother crystal” generates secondary crystals at a fast rate if the solution is stirred; in the case of NaClO3, a first l or d “mother crystal” forms new crystals with similar chirality in a few minutes. It is quite evident that the model of an initial single chiral crystal cannot explain the complete chiral purity in Viedma’s experiments. Considering the role of grinding glass balls in the symmetry breaking phenomenon, Viedma suggests that under these conditions a continuous dissolution–crystallization phenomenon is highly enhanced by the grinding process, making the system very similar to the homogeneous autocatalytic reaction. These results can provide a more prebiotically plausible extension of this process, indicating essentially chiral molecules that form conglomerate solids [18,43,44]. The formation of separate l and d crystals, as in those Pasteur separated with his tweezers, are known as “conglomerates”. The “preferential crystallization” method is well known as a way to “untangle” a racemic conglomerate of homochiral R and S crystals, by repeated crystallization from the supersaturated solution. In 1998, Tamura et al. proposed a phenomenon referred to as preferential enrichment, which is completely opposed to preferential crystallization: in preferential enrichment, it is in the mother liquor that substantial enantiomeric enrichment occurs by recrystallization, and, at the same time, a slight enrichment of the opposite enantiomer always occurs [45]. This new phenomenon has two fundamental characteristics: (1) recurring recrystallization of the racemate eventuates in the alternating enrichment of the two enantiomeric crystal forms up to 100% ee, at each crop of the deposited crystals; (2) in the event of recrystallization of nonracemic crystals, the sequential crystals consistently have the opposite chirality. Tamura et al., in one of their publications, reported an experimental case of preferential enrichment, in which the stable crystalline form of the racemate is the mixed crystal, which is composed of a weakly ordered arrangement of the two enantiomers and contains the centrosymmetric heterochiral cyclic dimer (R, S) as the major component in the crystal lattice (Figure 4) [46].
4. A Chemical Model for the Study of Symmetry Breaking of Phospholipids
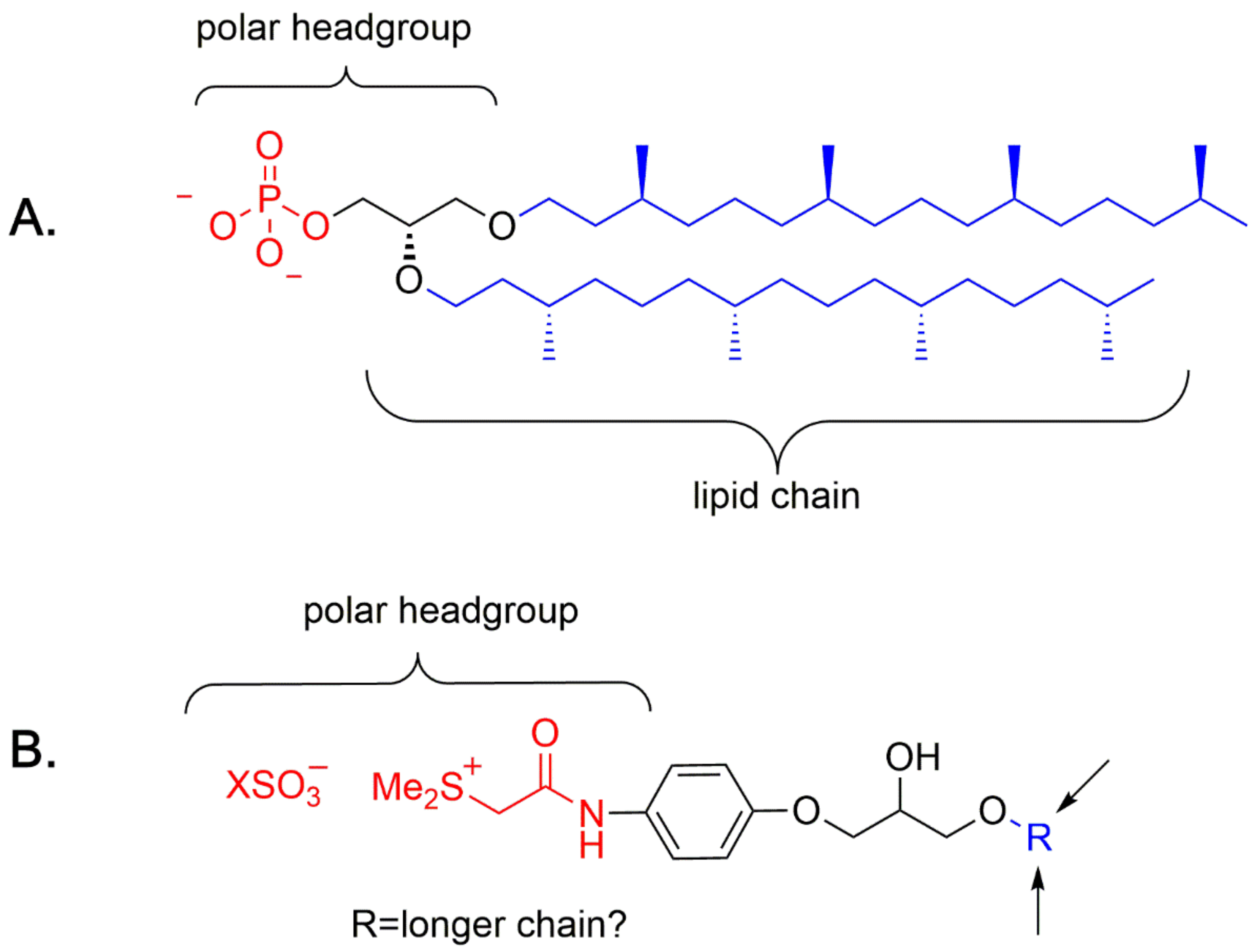
Nucleosides and, in particular, the prebiotic formation of ribo-nucleosides and the prebiotic synthesis of amino acids, have a central role in OOL studies [47]. Phospholipids and their precursors have not been the objects of research, with few exceptions [48]. Although a major step was taken by Sutherland and colleagues [49], the question of how symmetry breaking of phospholipids has occurred still remains an unsolved problem [34]. John Sutherland and colleagues have shown that several important classes of biomolecules, including phospholipids precursors, can be prepared under the same prebiotic conditions (see Figure A1, adapted from [49]) [49,50,51,52]. In a previous publication in this journal [34], one of the present authors pointed out that the lack of homochirality in phospholipid prebiotic synthesis leaves a gap in the comprehension of the role of encapsulation of biotic molecules, for example [53,54]. It is our opinion, however, that this problem can be solved using a model as a starting point and considering that phospholipids have been subjected to symmetry breaking in the solid state, as crystals. Looking at Tamura and colleagues’ structures, we have found an interesting analogy with phospholipid ethers, an analogy that can be extended to lyso-phospholipid esters, covering the spectra of phospholipids from Archaea to Eukarya and Bacteria. The possible model compounds should differentiate on the length of the hydrophobic alkyl chain (Figure 5, in blue) where a longer chain replaces the ethyl chain, whereas the polar head bears a sulfate salt (Figure 5, in red). On phospholipid esters only, the unique chiral center is located at the C-2 (sn-2) of the glycerol backbone (Figure 5, in black), whereas archaea phospholipids can possess chiral lipid chains, too, as shown in Figure 5 [26]. In addition, as previously reported, prebiotic synthesis of phospholipid ethers is practically absent in the list of experiments performed for the preparation of phospholipids, giving a crucial contribution to the research carried out previously [48]. For this reason, in Appendix A there is reported a plausible retrosynthesis of a “Tamura-like Archaea phospholipid-model”, starting from racemic alcohol mixtures.
Untangle a Racemic Conglomerate of Phospholipids
In previous research, we prepared a racemic dioleoyl phosphatidic acid (rac-DOPA) and dioleoyl phosphoethanolamines (rac-DOPE) from racemic dioleoyl glycerol (rac-DOG) [55]. These compounds served to prepare plausible prebiotic liposomes in the form of giant vesicles (GVs) [56], simulating the prebiotic membrane of protocells, see ref. [33] for a comprehensive summary. The main conclusion was that no difference between a mixture of pure DOPA plus a plausible prebiotic mixture of co-surfactants or rac-DOPA in prebiotic mixtures could be observed. The most relevant difference for the formation of GVs was the nature of the polar head and the concentration of the “vesiculation helpers”. A few years later we were able to publish the follow-up to this research, showing that pure POPC (1-palmitoyl-sn-2-oleoyl-glycero-3-phosphocholine) had a 10% difference in the retention of a polar hydrophilic dye with respect to pure rac-POPC or scalemic (2R:1S) scal-POPC vesicles. To the best of our knowledge, other experiments have not been carried out with biotic polymers such as proteins or RNA (or DNA). In the light of these results, we think that symmetry breaking of phospholipids occurred before the formation of the first protocells capable of retaining prebiotic enantiopure polymers. This fact does not exclude the mixed nature of the first protocell membrane, as speculated by Jordan and colleagues [57].
5. Conclusions
This short article summarizes the most relevant chemical models used to shed light on the plausible (prebiotic) symmetry breaking of biotic molecules. For the moment, the valuable Frank model cannot be applied to known prebiotic pathways such as the formose reaction that is relevant for the synthesis of sugars, including riboses, due to the lack of a chiral intermediate that leads to the formation of one enantiomer (erythrose, for example), yielding only racemic mixtures. Prebiotic simulated soups instead lead only to racemic mixtures of amino acids, sugars and phospholipids precursors. Instead, crystallization seems to be a solution to untangle a racemic conglomerate, using a phospholipid model. This essay is intended to be a precursor for the publication of some, in our opinion, breakthrough results obtained just before the submission of the first version of the manuscript. Too embryonal to be the object of publication, we can affirm that using models for understanding the origin of life, modeling the prebiotic environment, using bottom-up experiments in systems chemistry philosophy is the best perspective for understanding the emergence of homo-chirality.
Author Contributions
M.F. had the main idea and wrote the manuscript; F.S. and C.C. wrote the manuscript together with M.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
M.F. dedicates this work to his daughter Océane. We wish also to thank referee n3 for the following words, “Manuscript of Fiore and co-workers summarize the chemical models to justify the born of chirality in the Earth, taking as example phospholipids. The topic of this perspective is probably one of the most interesting that I have read”, his thought is more encouraging than any prize my students and I can receive. All the authors have approved the manuscript and consented to the acknowledgements.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A
Figure A1.
Possible retrosynthesis of a Tamura-like Archaea phospholipid-model [46]; retrosynthetic conditions: (a) p-nitrophenol (in red), toluene, PhCH2NMe3Cl, reflux, o.n.; (b) NH2NH2·H2O, FeCl3·6H2O, 90 °C, MeOH, o.n; or Pd(OH)2/C/H2, MeOH, r.t., o.n. (c) MeSCH2CH2COCl, NaOH, CH2Cl2, r.t., o.n.; (d) R2SO3Me, acetone, 48h, reflux); dashed lines indicate a disconnection between the possible bonds formed during the reactions.
Figure A1.
Possible retrosynthesis of a Tamura-like Archaea phospholipid-model [46]; retrosynthetic conditions: (a) p-nitrophenol (in red), toluene, PhCH2NMe3Cl, reflux, o.n.; (b) NH2NH2·H2O, FeCl3·6H2O, 90 °C, MeOH, o.n; or Pd(OH)2/C/H2, MeOH, r.t., o.n. (c) MeSCH2CH2COCl, NaOH, CH2Cl2, r.t., o.n.; (d) R2SO3Me, acetone, 48h, reflux); dashed lines indicate a disconnection between the possible bonds formed during the reactions.

References
- Suh, I.-H.; Park, K.H.; Jensen, W.P.; Lewis, D.E. Molecules, Crystals, and Chirality. J. Chem. Educ. 1997, 74, 800. [Google Scholar] [CrossRef]
- Müller, F.; Escobar, L.; Xu, F.; Węgrzyn, E.; Nainytė, M.; Amatov, T.; Chan, C.; Pichler, A.; Carell, T. A prebiotically plausible scenario of an RNA–peptide world. Nature 2022, 605, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Airapetian, V.S.; Glocer, A.; Gronoff, G.; Hébrard, E.; Danchi, W. Prebiotic chemistry and atmospheric warming of early Earth by an active young Sun. Nat. Geosci. 2016, 9, 452–455. [Google Scholar] [CrossRef]
- Schmitt-Kopplin, P.; Gabelica, Z.; Gougeon, R.D.; Fekete, A.; Kanawati, B.; Harir, M.; Gebefuegi, I.; Eckel, G.; Hertkorn, N. High molecular diversity of extraterrestrial organic matter in Murchison meteorite revealed 40 years after its fall. Proc. Natl. Acad. Sci. USA 2010, 107, 2763–2768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitadai, N.; Maruyama, S. Origins of building blocks of life: A review. Geosci. Front. 2018, 9, 1117–1153. [Google Scholar] [CrossRef]
- Ross, D.S.; Deamer, D. Dry/wet cycling and the thermodynamics and kinetics of prebiotic polymer synthesis. Life 2016, 6, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damer, B.; Deamer, D. The hot spring hypothesis for an origin of life. Astrobiology 2020, 20, 429–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reeves, E.P.; Seewald, J.S.; Saccocia, P.; Bach, W.; Craddock, P.R.; Shanks, W.C.; Sylva, S.P.; Walsh, E.; Pichler, T.; Rosner, M. Geochemistry of hydrothermal fluids from the PACMANUS, Northeast Pual and Vienna Woods hydrothermal fields, Manus Basin, Papua New Guinea. Geochim. Cosmochim. Acta 2011, 75, 1088–1123. [Google Scholar] [CrossRef] [Green Version]
- Miller, S.L.; Bada, J.L. Submarine hot springs and the origin of life. Nature 1988, 334, 609–611. [Google Scholar] [CrossRef] [PubMed]
- Damer, B.; Deamer, D. Coupled Phases and Combinatorial Selection in Fluctuating Hydrothermal Pools: A Scenario to Guide Experimental Approaches to the Origin of Cellular Life. Life 2015, 5, 872–887. [Google Scholar] [CrossRef]
- Deamer, D. The Role of Lipid Membranes in Life’s Origin. Life 2017, 7, 5. [Google Scholar] [CrossRef] [PubMed]
- Deamer, D. Origins of Life Research: The Conundrum between Laboratory and Field Simulations of Messy Environments. Life 2022, 12, 1429. [Google Scholar] [CrossRef] [PubMed]
- Schwarzenbach, E.M.; Früh-Green, G.L.; Bernasconi, S.M.; Alt, J.C.; Shanks, W.C.; Gaggero, L.; Crispini, L. Sulfur geochemistry of peridotite-hosted hydrothermal systems: Comparing the Ligurian ophiolites with oceanic serpentinites. Geochim. Cosmochim. Acta 2012, 91, 283–305. [Google Scholar] [CrossRef]
- Rickard, D. Pyrite and the Origins of Life. In Pyrite; Oxford University Press: Oxford, UK, 2015. [Google Scholar]
- Avalos, M.; Babiano, R.; Cintas, P.; Jimenez, J.L.; Palacios, J.C. Symmetry Breaking by Spontaneous Crystallization—Is it the Most Plausible Source of Terrestrial Handedness we have Long Been Looking for?—A Reappraisal. Orig. Life Evol. Biosph. 2004, 34, 391–405. [Google Scholar] [CrossRef]
- Chen, Y.; Ma, W. The origin of biological homochirality along with the origin of life. PLoS Comput. Biol. 2020, 16, e1007592. [Google Scholar] [CrossRef] [PubMed]
- Gal, J. When did Louis Pasteur present his memoir on the discovery of molecular chirality to the Académie des sciences? Analysis of a discrepancy. Chirality 2008, 20, 1072–1084. [Google Scholar] [CrossRef]
- Blackmond, D.G. The origin of biological homochirality. Cold Spring Harb. Perspect. Biol. 2019, 2, a002147. [Google Scholar] [CrossRef] [Green Version]
- Cronin, J.R.; Gandy, W.E.; Pizzarello, S. Amino acids of the murchison meteorite: I. Six Carbon Acyclic Primary α-Amino Alkanoic Acids. J. Mol. Evol. 1981, 17, 265–272. [Google Scholar] [CrossRef]
- Engel, M.H.; Nagy, B.; Bada, J.L.; Cronin, J.R.; Ho, M.-S.S.; Kvenvolden, K.A.; Lawless, J.G.; Miller, S.L.; Oro, J.; Steinberg, S. On the reported optical activity of amino acids in the Murchison meteorite (reply). Nature 1983, 301, 496–497. [Google Scholar] [CrossRef]
- Cronin, J.R.; Pizzarello, S. Enantiomeric excesses in meteoritic amino acids. Science 1997, 275, 951–955. [Google Scholar] [CrossRef]
- Cronin, J.R.; Pizzarello, S. Amino acids of the Murchison meteorite. III. Seven carbon acyclic primary α-amino alkanoic acids1. Geochim. Cosmochim. Acta 1986, 50, 2419–2427. [Google Scholar] [CrossRef]
- Garcia, A.; Meinert, C.; Sugahara, H.; Jones, N.; Hoffmann, S.; Meierhenrich, U. The Astrophysical Formation of Asymmetric Molecules and the Emergence of a Chiral Bias. Life 2019, 9, 29. [Google Scholar] [CrossRef] [Green Version]
- Burton, A.; Berger, E. Insights into Abiotically-Generated Amino Acid Enantiomeric Excesses Found in Meteorites. Life 2018, 8, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker, E.T.; Cleaves, H.J.; Dworkin, J.P.; Glavin, D.P.; Callahan, M.; Aubrey, A.; Lazcano, A.; Bada, J.L. Primordial synthesis of amines and amino acids in a 1958 Miller H 2S-rich spark discharge experiment. Proc. Natl. Acad. Sci. USA 2011, 108, 5526–5531. [Google Scholar] [CrossRef] [Green Version]
- Engel, M.H.; Macko, S.A.; Silfer, J.A. Carbon isotope composition of individual amino acids in the Murchison meteorite. Nature 1990, 348, 47–49. [Google Scholar] [CrossRef] [PubMed]
- Engel, M.H.; Nagy, B. Distribution and enantiomeric composition of amino acids in the Murchison meteorite. Nature 1982, 296, 837–840. [Google Scholar] [CrossRef]
- Cronin, J.R.; Pizzarello, S.; Yuen, G.U. Amino acids of the Murchison meteorite: II. Five carbon acyclic primary β-, γ-, and δ-amino alkanoic acids. Geochim. Cosmochim. Acta 1985, 49, 2259–2265. [Google Scholar] [CrossRef]
- Kvenvolden, K.A.; Lawless, J.G.; Ponnamperuma, C. Nonprotein Amino Acids in the Murchison Meteorite. Proc. Natl. Acad. Sci. USA 1971, 68, 486–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, S.L. A Production of Amino Acids Under Possible Primitive Earth Conditions. Science 1953, 117, 528–529. [Google Scholar] [CrossRef] [Green Version]
- Skolnick, J.; Zhou, H.; Gao, M. On the possible origin of protein homochirality, structure, and biochemical function. Proc. Natl. Acad. Sci. USA 2019, 116, 26571–26579. [Google Scholar] [CrossRef] [PubMed]
- Grishin, D.V.; Zhdanov, D.D.; Pokrovskaya, M.V.; Sokolov, N.N. D-amino acids in nature, agriculture and biomedicine. All Life 2020, 13, 11–22. [Google Scholar] [CrossRef] [Green Version]
- Lopez, A.; Fiore, M. Investigating Prebiotic Protocells for a Comprehensive Understanding of the Origins of Life: A Prebiotic Systems Chemistry Perspective. Life 2019, 9, 49. [Google Scholar] [CrossRef] [Green Version]
- Fiore, M.; Buchet, R. Symmetry breaking of phospholipids. Symmetry 2020, 12, 1488. [Google Scholar] [CrossRef]
- Frank, F.C. On spontaneous asymmetric synthesis. Biochim. Biophys. Acta 1953, 11, 459–463. [Google Scholar] [CrossRef]
- Micheau, J.-C.; Coudret, C.; Cruz, J.-M.; Buhse, T. Amplification of enantiomeric excess, mirror-image symmetry breaking and kinetic proofreading in Soai reaction models with different oligomeric orders. Phys. Chem. Chem. Phys. 2012, 14, 13239. [Google Scholar] [CrossRef]
- Breslow, R. On the mechanisms of formose reaction. Tetrahedron Lett. 1959, 1, 22–26. [Google Scholar] [CrossRef]
- Breslow, R.; Cheng, Z.L. L-amino acids catalyze the formation of an excess of D-glyceraldehyde, and thus of other D sugars, under credible prebiotic conditions. Proc. Natl. Acad. Sci. USA 2010, 107, 5723–5725. [Google Scholar] [CrossRef] [Green Version]
- Soai, K.; Kawasaki, T.; Matsumoto, A. Role of Asymmetric Autocatalysis in the Elucidation of Origins of Homochirality of Organic Compounds. Symmetry 2019, 11, 694. [Google Scholar] [CrossRef] [Green Version]
- Hawbaker, N.A.; Blackmond, D.G. Energy threshold for chiral symmetry breaking in molecular self-replication. Nat. Chem. 2019, 11, 957–962. [Google Scholar] [CrossRef]
- Geiger, Y. One Soai reaction, two mechanisms? Chem. Soc. Rev. 2022, 51, 1206–1211. [Google Scholar] [CrossRef]
- Athavale, S.V.; Simon, A.; Houk, K.N.; Denmark, S.E. Demystifying the asymmetry-amplifying, autocatalytic behaviour of the Soai reaction through structural, mechanistic and computational studies. Nat. Chem. 2020, 12, 412–423. [Google Scholar] [CrossRef]
- Viedma, C. Experimental evidence of chiral symmetry breaking in crystallization from primary nucleation. J. Cryst. Growth 2004, 261, 118–121. [Google Scholar] [CrossRef] [Green Version]
- Kondepudi, D.K.; Kaufman, R.J.; Singh, N. Chiral Symmetry Breaking in Sodium Chlorate Crystallization. Science 1990, 250, 975–976. [Google Scholar] [CrossRef]
- Tamura, R.; Iwama, S.; Takahashi, H. Chiral Symmetry Breaking Phenomenon Caused by a Phase Transition. Symmetry 2010, 2, 112–135. [Google Scholar] [CrossRef]
- Tamura, R.; Takahashi, H.; Hirotsu, K.; Nakajima, Y.; Ushio, T.; Toda, F. Unusual disordered crystal structure of a racemate exhibiting a novel enantiomeric resolution: Preferential enrichment. Angew. Chemie-Int. Ed. 1998, 37, 2876–2878. [Google Scholar] [CrossRef]
- Ruiz-Mirazo, K.; Briones, C.; De La Escosura, A. Prebiotic systems chemistry: New perspectives for the origins of life. Chem. Rev. 2014, 114, 285–366. [Google Scholar] [CrossRef]
- Fiore, M.; Chieffo, C.; Lopez, A.; Fayolle, D.; Ruiz, J.; Soulère, L.; Oger, P.; Altamura, E.; Popowycz, F.; Buchet, R. Synthesis of Phospholipids Under Plausible Prebiotic Conditions and Analogies with Phospholipid Biochemistry for Origin of Life Studies. Astrobiology 2022, 22, 598–627. [Google Scholar] [CrossRef]
- Patel, B.H.; Percivalle, C.; Ritson, D.J.; Duffy, C.D.; Sutherland, J.D. Common origins of RNA, protein and lipid precursors in a cyanosulfidic protometabolism. Nat. Chem. 2015, 7, 301–307. [Google Scholar] [CrossRef] [Green Version]
- Bonfio, C.; Valer, L.; Scintilla, S.; Shah, S.; Evans, D.J.; Jin, L.; Szostak, J.W.; Sasselov, D.D.; Sutherland, J.D.; Mansy, S.S. UV-light-driven prebiotic synthesis of iron-sulfur clusters. Nat. Chem. 2017, 9, 1229–1234. [Google Scholar] [CrossRef]
- Bonfio, C.; Caumes, C.; Duffy, C.D.; Patel, B.H.; Percivalle, C.; Tsanakopoulou, M.; Sutherland, J.D. Length-Selective Synthesis of Acylglycerol-Phosphates through Energy-Dissipative Cycling. J. Am. Chem. Soc. 2019, 141, 3934–3939. [Google Scholar] [CrossRef]
- Liu, Z.; Wu, L.; Xu, J.; Bonfio, C.; Russell, D.A.; Pascal, R.; Sutherland, J.D. Harnessing Chemical Energy for the Activation and Joining of Prebiotic Building Blocks. Nat. Chem. 2019, 12, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Lopez, A.; Chieffo, C.; Fiore, M. Abiotic Synthesis and Role of Amphiphiles in the Encapsulation Process in Life’s Origin. In Prebiotic Chemistry and The Origin of Life; Neubeck Anna, M.S., Ed.; Springer: Berlin/Heidelberg, Germany, 2021; pp. 139–164. ISBN 978-3-030-81038-2. [Google Scholar]
- Altamura, E.; Comte, A.; D’Onofrio, A.; Roussillon, C.; Fayolle, D.; Buchet, R.; Mavelli, F.; Stano, P.; Fiore, M.; Strazewski, P. Racemic Phospholipids for Origin of Life Studies. Symmetry 2020, 12, 1108. [Google Scholar] [CrossRef]
- Fayolle, D.; Altamura, E.; D’Onofrio, A.; Madanamothoo, W.; Fenet, B.; Mavelli, F.; Buchet, R.; Stano, P.; Fiore, M.; Strazewski, P. Crude phosphorylation mixtures containing racemic lipid amphiphiles self-assemble to give stable primitive compartments. Sci. Rep. 2017, 7, 18106. [Google Scholar] [CrossRef] [Green Version]
- Wick, R.; Walde, P.; Luisi, P.L. Giant Vesicles. In Self-Production of Supramolecular Structures; Springer: Dordrecht, The Netherlands, 1994; pp. 295–299. [Google Scholar]
- Jordan, S.F.; Rammu, H.; Zheludev, I.N.; Hartley, A.M.; Maréchal, A.; Lane, N. Promotion of protocell self-assembly from mixed amphiphiles at the origin of life. Nat. Ecol. Evol. 2019, 3, 1705–1714. [Google Scholar] [CrossRef]
Figure 1.
Frank’s model for autocatalysis.

Figure 2.
Breslow’s formose reaction mechanism, where glycoaldehyde (in red) catalyses its own formation.
Figure 2.
Breslow’s formose reaction mechanism, where glycoaldehyde (in red) catalyses its own formation.

Figure 3.
An example of asymmetry-amplifying, autocatalytic alkylation of pyrimidine-5-carbaldehydes with diisopropylzinc, named the ‘Soai reaction’. Elucidation of its mechanism and the role of each chiral initiator used is recently summarized in ref [41].
Figure 3.
An example of asymmetry-amplifying, autocatalytic alkylation of pyrimidine-5-carbaldehydes with diisopropylzinc, named the ‘Soai reaction’. Elucidation of its mechanism and the role of each chiral initiator used is recently summarized in ref [41].

Figure 4.
Enantiomeric resolution by successive recrystallization of (±)-SN (in black) in R or S enantiomers by preferential enrichment. Crystallization conditions are reported from ethanol at r.t. in 24 h (a–d) or from evaporation of ethanol (e).
Figure 4.
Enantiomeric resolution by successive recrystallization of (±)-SN (in black) in R or S enantiomers by preferential enrichment. Crystallization conditions are reported from ethanol at r.t. in 24 h (a–d) or from evaporation of ethanol (e).

Figure 5.
Structural analogies between a phospholipid ether ((A), from archaea) and a general structure from research carried out by Tamura and co-workers and (B) possible modification.
Figure 5.
Structural analogies between a phospholipid ether ((A), from archaea) and a general structure from research carried out by Tamura and co-workers and (B) possible modification.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Differences between the analysis of prebiotic prepared mixtures of amino acids and the content of Murchison meteorite samples. Only the latter contains an excess of l-amino acids, whereas the former contain racemic mixtures.
Table 1.
Differences between the analysis of prebiotic prepared mixtures of amino acids and the content of Murchison meteorite samples. Only the latter contains an excess of l-amino acids, whereas the former contain racemic mixtures.
| Year | Amino Acids | Comments/References |
|---|---|---|
| 2011 a | Gly, Ala, Ser, Thr, Asp, Glu, Met, Val, Leu, Phe, β-Ala, Iso-serine, Homocysteic acid, α-ABA β-ABA, α-AIB, β-AIB, γ-ABI, S-Methyl Cysteine, Iso-valine, Methionine sulfoxide, Methionine sulfone, Iso-leucine, Ethionine, MA, EA, Cysteamine, Ethanolamine | Analysis of the original samples from Miller—Bada experiments. Reactions were carried out in the presence of H2S in addition to CH4, NH3 and H2O [25] |
| From 1969 | Glu (0.322); Asp (0.202); Pro (0.342); Leu (0.166); Ala (0.682) | Murchinson meteorite extracts [26,27]: predominancy of l-configuration; more than 20 amino acids were found in the Murchinson meteorite [19,20,22,23,28,29] |
a the reported results are the most representative re-analysis of one of Miller’s original samples [30].
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Skorda, F.; Chieffo, C.; Fiore, M. Chemical Models for Understanding the Emergence of Homo-Chirality of Phospholipids for Origin of Life Studies. Symmetry 2022, 14, 2109. https://doi.org/10.3390/sym14102109
AMA Style
Skorda F, Chieffo C, Fiore M. Chemical Models for Understanding the Emergence of Homo-Chirality of Phospholipids for Origin of Life Studies. Symmetry. 2022; 14(10):2109. https://doi.org/10.3390/sym14102109
Chicago/Turabian StyleSkorda, Fryni, Carolina Chieffo, and Michele Fiore. 2022. "Chemical Models for Understanding the Emergence of Homo-Chirality of Phospholipids for Origin of Life Studies" Symmetry 14, no. 10: 2109. https://doi.org/10.3390/sym14102109
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.