
Identification of the Binding Epitope of an Anti-Mouse CCR6 Monoclonal Antibody (C6Mab-13) Using 1× Alanine Scanning
 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Antibodies
2.2. Peptides
2.3. ELISA
2.4. Measurement of Dissociation Constants Using Surface Plasmon Resonance (SPR)
3. Results
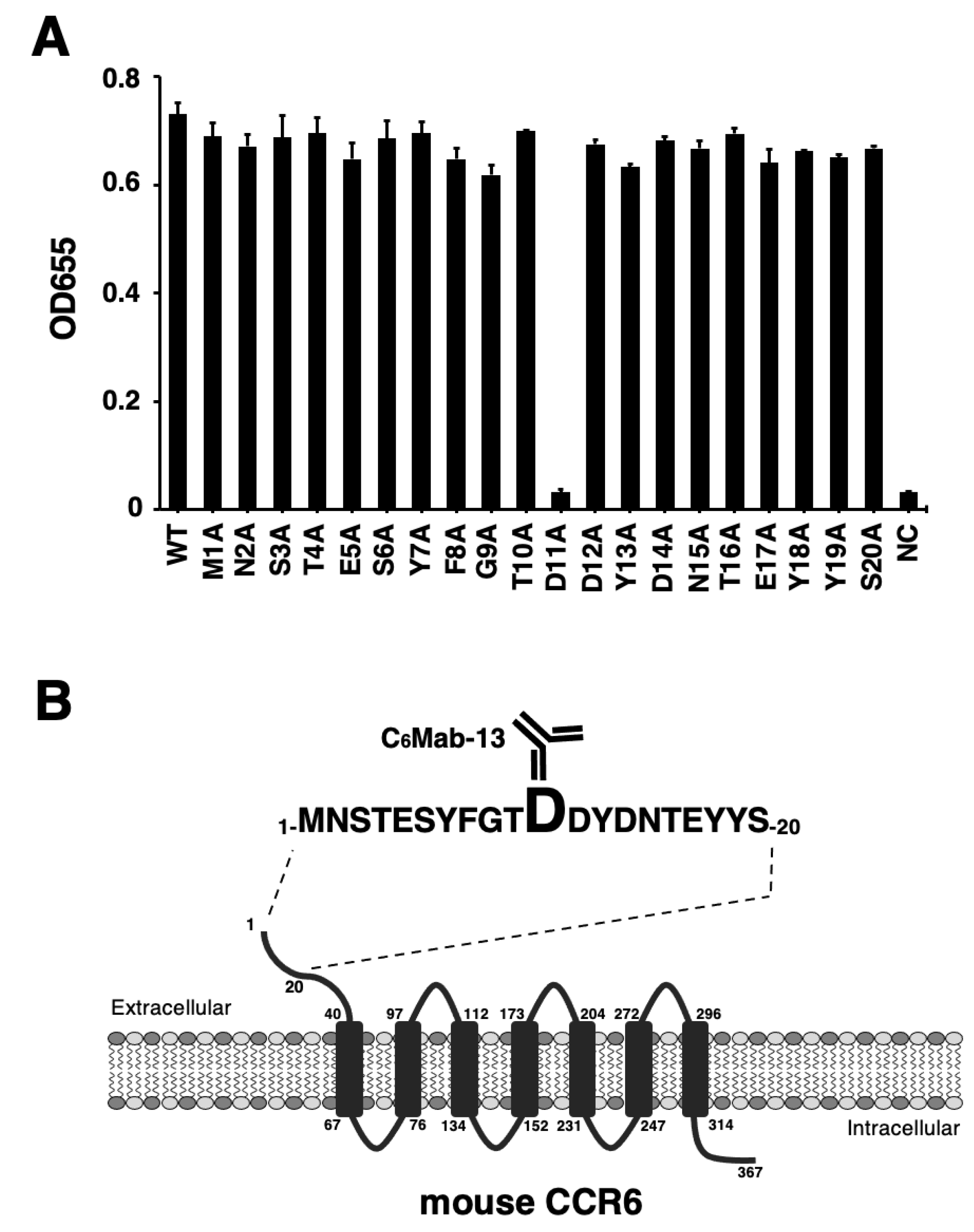
3.1. Epitope Identification of C6Mab-13 by ELISA Using 1× Alanine-Substituted mCCR6 Peptides
3.2. Epitope Identification of C6Mab-13 by SPR Using 1× Alanine-Substituted mCCR6 Peptides
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Morein, D.; Erlichman, N.; Ben-Baruch, A. Beyond Cell Motility: The Expanding Roles of Chemokines and Their Receptors in Malignancy. Front. Immunol. 2020, 11, 952. [Google Scholar] [CrossRef]
- Mollica Poeta, V.; Massara, M.; Capucetti, A.; Bonecchi, R. Chemokines and Chemokine Receptors: New Targets for Cancer Immunotherapy. Front. Immunol. 2019, 10, 379. [Google Scholar] [CrossRef]
- Hughes, C.E.; Nibbs, R.J.B. A guide to chemokines and their receptors. Febs. J. 2018, 285, 2944–2971. [Google Scholar] [CrossRef]
- Zlotnik, A.; Yoshie, O. The chemokine superfamily revisited. Immunity 2012, 36, 705–716. [Google Scholar] [CrossRef] [PubMed]
- Schutyser, E.; Struyf, S.; Van Damme, J. The CC chemokine CCL20 and its receptor CCR6. Cytokine Growth Factor. Rev. 2003, 14, 409–426. [Google Scholar] [CrossRef] [PubMed]
- Baba, M.; Imai, T.; Nishimura, M.; Kakizaki, M.; Takagi, S.; Hieshima, K.; Nomiyama, H.; Yoshie, O. Identification of CCR6, the specific receptor for a novel lymphocyte-directed CC chemokine LARC. J. Biol. Chem. 1997, 272, 14893–14898. [Google Scholar] [CrossRef] [PubMed]
- Korbecki, J.; Grochans, S.; Gutowska, I.; Barczak, K.; Baranowska-Bosiacka, I. CC Chemokines in a Tumor: A Review of Pro-Cancer and Anti-Cancer Properties of Receptors CCR5, CCR6, CCR7, CCR8, CCR9, and CCR10 Ligands. Int. J. Mol. Sci. 2020, 21, 7619. [Google Scholar] [CrossRef]
- Kadomoto, S.; Izumi, K.; Mizokami, A. The CCL20-CCR6 Axis in Cancer Progression. Int. J. Mol. Sci. 2020, 21, 5186. [Google Scholar] [CrossRef]
- Chen, W.; Qin, Y.; Liu, S. CCL20 Signaling in the Tumor Microenvironment. Adv. Exp. Med. Biol. 2020, 1231, 53–65. [Google Scholar] [CrossRef]
- Gómez-Melero, S.; Caballero-Villarraso, J. CCR6 as a Potential Target for Therapeutic Antibodies for the Treatment of Inflammatory Diseases. Antibodies 2023, 12, 30. [Google Scholar] [CrossRef]
- Misra, D.P.; Agarwal, V. Th17.1 lymphocytes: Emerging players in the orchestra of immune-mediated inflammatory diseases. Clin. Rheumatol. 2022, 41, 2297–2308. [Google Scholar] [CrossRef] [PubMed]
- Meitei, H.T.; Jadhav, N.; Lal, G. CCR6-CCL20 axis as a therapeutic target for autoimmune diseases. Autoimmun. Rev. 2021, 20, 102846. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Yang, G.; Xiao, F.; Xie, J.; Wang, S.; Lu, L.; Cui, D. Role of Th22 Cells in the Pathogenesis of Autoimmune Diseases. Front. Immunol. 2021, 12, 688066. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, S.; Xing, M.; Hong, S.; Liu, L.; Ding, X.J.; Sun, X.Y.; Luo, Y.; Wang, C.X.; Zhang, M.; et al. Current evidence on the role of lipid lowering drugs in the treatment of psoriasis. Front. Med. 2022, 9, 900916. [Google Scholar] [CrossRef]
- Furue, M.; Furue, K.; Tsuji, G.; Nakahara, T. Interleukin-17A and Keratinocytes in Psoriasis. Int. J. Mol. Sci. 2020, 21, 1275. [Google Scholar] [CrossRef] [PubMed]
- Furue, K.; Ito, T.; Tsuji, G.; Nakahara, T.; Furue, M. The CCL20 and CCR6 axis in psoriasis. Scand. J. Immunol. 2020, 91, e12846. [Google Scholar] [CrossRef] [PubMed]
- Hedrick, M.N.; Lonsdorf, A.S.; Hwang, S.T.; Farber, J.M. CCR6 as a possible therapeutic target in psoriasis. Expert. Opin. Ther. Targets 2010, 14, 911–922. [Google Scholar] [CrossRef] [PubMed]
- Martina, M.G.; Giorgio, C.; Allodi, M.; Palese, S.; Barocelli, E.; Ballabeni, V.; Szpakowska, M.; Chevigné, A.; Piet van Hamburg, J.; Davelaar, N.; et al. Discovery of small-molecules targeting the CCL20/CCR6 axis as first-in-class inhibitors for inflammatory bowel diseases. Eur. J. Med. Chem. 2022, 243, 114703. [Google Scholar] [CrossRef]
- Trivedi, P.J.; Adams, D.H. Chemokines and Chemokine Receptors as Therapeutic Targets in Inflammatory Bowel Disease; Pitfalls and Promise. J. Crohns. Colitis. 2018, 12, S641–S652. [Google Scholar] [CrossRef]
- Skovdahl, H.K.; Granlund, A.; Østvik, A.E.; Bruland, T.; Bakke, I.; Torp, S.H.; Damås, J.K.; Sandvik, A.K. Expression of CCL20 and Its Corresponding Receptor CCR6 Is Enhanced in Active Inflammatory Bowel Disease, and TLR3 Mediates CCL20 Expression in Colonic Epithelial Cells. PLoS ONE 2015, 10, e0141710. [Google Scholar] [CrossRef] [PubMed]
- Cheluvappa, R. Experimental appendicitis and appendectomy modulate the CCL20-CCR6 axis to limit inflammatory colitis pathology. Int. J. Colorectal. Dis. 2014, 29, 1181–1188. [Google Scholar] [CrossRef] [PubMed]
- Kaser, A.; Ludwiczek, O.; Holzmann, S.; Moschen, A.R.; Weiss, G.; Enrich, B.; Graziadei, I.; Dunzendorfer, S.; Wiedermann, C.J.; Mürzl, E.; et al. Increased expression of CCL20 in human inflammatory bowel disease. J. Clin. Immunol. 2004, 24, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.Y.S.; Körner, H. The CCR6-CCL20 axis in humoral immunity and T-B cell immunobiology. Immunobiology 2019, 224, 449–454. [Google Scholar] [CrossRef] [PubMed]
- Kondo, T.; Takata, H.; Takiguchi, M. Functional expression of chemokine receptor CCR6 on human effector memory CD8+ T cells. Eur. J. Immunol. 2007, 37, 54–65. [Google Scholar] [CrossRef]
- Rescigno, M. CCR6(+) dendritic cells: The gut tactical-response unit. Immunity 2006, 24, 508–510. [Google Scholar] [CrossRef]
- Annunziato, F.; Cosmi, L.; Santarlasci, V.; Maggi, L.; Liotta, F.; Mazzinghi, B.; Parente, E.; Filì, L.; Ferri, S.; Frosali, F.; et al. Phenotypic and functional features of human Th17 cells. J. Exp. Med. 2007, 204, 1849–1861. [Google Scholar] [CrossRef]
- Essien, K.I.; Katz, E.L.; Strassner, J.P.; Harris, J.E. Regulatory T Cells Require CCR6 for Skin Migration and Local Suppression of Vitiligo. J. Invest. Dermatol. 2022, 142, 3158–3166.e3157. [Google Scholar] [CrossRef]
- Varona, R.; Villares, R.; Carramolino, L.; Goya, I.; Zaballos, A.; Gutiérrez, J.; Torres, M.; Martínez, A.C.; Márquez, G. CCR6-deficient mice have impaired leukocyte homeostasis and altered contact hypersensitivity and delayed-type hypersensitivity responses. J. Clin. Invest. 2001, 107, R37–R45. [Google Scholar] [CrossRef]
- Power, C.A.; Church, D.J.; Meyer, A.; Alouani, S.; Proudfoot, A.E.; Clark-Lewis, I.; Sozzani, S.; Mantovani, A.; Wells, T.N. Cloning and characterization of a specific receptor for the novel CC chemokine MIP-3alpha from lung dendritic cells. J. Exp. Med. 1997, 186, 825–835. [Google Scholar] [CrossRef]
- Hieshima, K.; Imai, T.; Opdenakker, G.; Van Damme, J.; Kusuda, J.; Tei, H.; Sakaki, Y.; Takatsuki, K.; Miura, R.; Yoshie, O.; et al. Molecular cloning of a novel human CC chemokine liver and activation-regulated chemokine (LARC) expressed in liver. Chemotactic activity for lymphocytes and gene localization on chromosome 2. J. Biol. Chem. 1997, 272, 5846–5853. [Google Scholar] [CrossRef]
- Hromas, R.; Gray, P.W.; Chantry, D.; Godiska, R.; Krathwohl, M.; Fife, K.; Bell, G.I.; Takeda, J.; Aronica, S.; Gordon, M.; et al. Cloning and characterization of exodus, a novel beta-chemokine. Blood 1997, 89, 3315–3322. [Google Scholar] [PubMed]
- Kulkarni, N.; Meitei, H.T.; Sonar, S.A.; Sharma, P.K.; Mujeeb, V.R.; Srivastava, S.; Boppana, R.; Lal, G. CCR6 signaling inhibits suppressor function of induced-Treg during gut inflammation. J. Autoimmun. 2018, 88, 121–130. [Google Scholar] [CrossRef]
- Oh, H.; Ghosh, S. NF-κB: Roles and regulation in different CD4(+) T-cell subsets. Immunol. Rev. 2013, 252, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Noubade, R.; Krementsov, D.N.; Del Rio, R.; Thornton, T.; Nagaleekar, V.; Saligrama, N.; Spitzack, A.; Spach, K.; Sabio, G.; Davis, R.J.; et al. Activation of p38 MAPK in CD4 T cells controls IL-17 production and autoimmune encephalomyelitis. Blood 2011, 118, 3290–3300. [Google Scholar] [CrossRef] [PubMed]
- Haxhinasto, S.; Mathis, D.; Benoist, C. The AKT-mTOR axis regulates de novo differentiation of CD4+Foxp3+ cells. J. Exp. Med. 2008, 205, 565–574. [Google Scholar] [CrossRef]
- Yang, X.O.; Panopoulos, A.D.; Nurieva, R.; Chang, S.H.; Wang, D.; Watowich, S.S.; Dong, C. STAT3 regulates cytokine-mediated generation of inflammatory helper T cells. J. Biol. Chem. 2007, 282, 9358–9363. [Google Scholar] [CrossRef]
- Harper, E.G.; Guo, C.; Rizzo, H.; Lillis, J.V.; Kurtz, S.E.; Skorcheva, I.; Purdy, D.; Fitch, E.; Iordanov, M.; Blauvelt, A. Th17 cytokines stimulate CCL20 expression in keratinocytes in vitro and in vivo: Implications for psoriasis pathogenesis. J. Invest. Dermatol. 2009, 129, 2175–2183. [Google Scholar] [CrossRef]
- Ito, T.; Carson, W.F.T.; Cavassani, K.A.; Connett, J.M.; Kunkel, S.L. CCR6 as a mediator of immunity in the lung and gut. Exp. Cell. Res. 2011, 317, 613–619. [Google Scholar] [CrossRef]
- Ranasinghe, R.; Eri, R. Modulation of the CCR6-CCL20 Axis: A Potential Therapeutic Target in Inflammation and Cancer. Medicina (Kaunas) 2018, 54, 88. [Google Scholar] [CrossRef]
- Lee, A.Y.S.; Körner, H. CCR6/CCL20 chemokine axis in human immunodeficiency virus immunity and pathogenesis. J. Gen. Virol. 2017, 98, 338–344. [Google Scholar] [CrossRef]
- Lee, A.Y.; Körner, H. CCR6 and CCL20: Emerging players in the pathogenesis of rheumatoid arthritis. Immunol. Cell. Biol. 2014, 92, 354–358. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, R.; Ichimura, Y.; Kubota, N.; Konishi, R.; Nakamura, Y.; Mizuno, S.; Takahashi, S.; Fujimoto, M.; Nomura, T.; Okiyama, N. The Role of PD-L1 on Langerhans Cells in the Regulation of Psoriasis. J. Invest. Dermatol. 2022, 142, 3167–3174.e3169. [Google Scholar] [CrossRef]
- Kadomoto, S.; Izumi, K.; Hiratsuka, K.; Nakano, T.; Naito, R.; Makino, T.; Iwamoto, H.; Yaegashi, H.; Shigehara, K.; Kadono, Y.; et al. Tumor-Associated Macrophages Induce Migration of Renal Cell Carcinoma Cells via Activation of the CCL20-CCR6 Axis. Cancers 2019, 12, 89. [Google Scholar] [CrossRef] [PubMed]
- Han, G.; Wu, D.; Yang, Y.; Li, Z.; Zhang, J.; Li, C. CrkL meditates CCL20/CCR6-induced EMT in gastric cancer. Cytokine 2015, 76, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Lou, X.M.; He, Y. Preferential recruitment of Th17 cells to cervical cancer via CCR6-CCL20 pathway. PLoS ONE 2015, 10, e0120855. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Zhao, X.; Zhu, J.; Zhang, L.; Chen, Y.; Zhang, B.; Li, Y.; Wang, M.; Zhang, Z.; Wang, C. lncRNA-u50535 promotes the progression of lung cancer by activating CCL20/ERK signaling. Oncol. Rep. 2019, 42, 1946–1956. [Google Scholar] [CrossRef]
- Zhang, X.P.; Hu, Z.J.; Meng, A.H.; Duan, G.C.; Zhao, Q.T.; Yang, J. Role of CCL20/CCR6 and the ERK signaling pathway in lung adenocarcinoma. Oncol. Lett. 2017, 14, 8183–8189. [Google Scholar] [CrossRef]
- Lee, J.J.; Kao, K.C.; Chiu, Y.L.; Jung, C.J.; Liu, C.J.; Cheng, S.J.; Chang, Y.L.; Ko, J.Y.; Chia, J.S. Enrichment of Human CCR6(+) Regulatory T Cells with Superior Suppressive Activity in Oral Cancer. J. Immunol. 2017, 199, 467–476. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Li, G.; Asano, T.; Saito, M.; Kaneko, M.K.; Suzuki, H.; Kato, Y. Development of a Novel Anti-Mouse CCR2 Monoclonal Antibody (C(2)Mab-6) by N-Terminal Peptide Immunization. Monoclon. Antib. Immunodiagn. Immunother. 2022, 41, 80–86. [Google Scholar] [CrossRef]
- Asano, T.; Suzuki, H.; Tanaka, T.; Saito, M.; Li, G.; Goto, N.; Nanamiya, R.; Kaneko, M.K.; Kato, Y. C(3)Mab-3: A Monoclonal Antibody for Mouse CC Chemokine Receptor 3 for Flow Cytometry. Monoclon. Antib. Immunodiagn. Immunother. 2022, 41, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Takei, J.; Suzuki, H.; Asano, T.; Tanaka, T.; Kaneko, M.K.; Kato, Y. Development of a Novel Anti-Mouse CCR4 Monoclonal Antibody (C(4)Mab-1) by N-Terminal Peptide Immunization. Monoclon. Antib. Immunodiagn. Immunother. 2022, 41, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Asano, T.; Tanaka, T.; Suzuki, H.; Li, G.; Nanamiya, R.; Tateyama, N.; Isoda, Y.; Okada, Y.; Kobayashi, H.; Yoshikawa, T.; et al. Development of a Novel Anti-Mouse CCR6 Monoclonal Antibody (C(6)Mab-13) by N-Terminal Peptide Immunization. Monoclon. Antib. Immunodiagn. Immunother. 2022, 41, 343–349. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, H.; Asano, T.; Suzuki, H.; Tanaka, T.; Yoshikawa, T.; Kaneko, M.K.; Kato, Y. Establishment of a Sensitive Monoclonal Antibody Against Mouse CCR9 (C(9)Mab-24) for Flow Cytometry. Monoclon. Antib. Immunodiagn. Immunother. 2023, 42, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, K.; Suzuki, H.; Kaneko, M.K.; Kato, Y. Cx(6)Mab-1: A Novel Anti-Mouse CXCR6 Monoclonal Antibody Established by N-Terminal Peptide Immunization. Monoclon. Antib. Immunodiagn. Immunother. 2022, 41, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Chamorro, S.; Vela, M.; Franco-Villanueva, A.; Carramolino, L.; Gutiérrez, J.; Gómez, L.; Lozano, M.; Salvador, B.; García-Gallo, M.; Martínez, A.C.; et al. Antitumor effects of a monoclonal antibody to human CCR9 in leukemia cell xenografts. MAbs 2014, 6, 1000–1012. [Google Scholar] [CrossRef] [PubMed]
- Chain, B.; Arnold, J.; Akthar, S.; Brandt, M.; Davis, D.; Noursadeghi, M.; Lapp, T.; Ji, C.; Sankuratri, S.; Zhang, Y.; et al. A Linear Epitope in the N-Terminal Domain of CCR5 and Its Interaction with Antibody. PLoS ONE 2015, 10, e0128381. [Google Scholar] [CrossRef]
- Gómez-Melero, S.; García-Maceira, F.I.; García-Maceira, T.; Luna-Guerrero, V.; Montero-Peñalvo, G.; Túnez-Fiñana, I.; Paz-Rojas, E. Amino terminal recognition by a CCR6 chemokine receptor antibody blocks CCL20 signaling and IL-17 expression via β-arrestin. BMC Biotechnol. 2021, 21, 41. [Google Scholar] [CrossRef] [PubMed]
- Aguilera-Durán, G.; Romo-Mancillas, A. Behavior of Chemokine Receptor 6 (CXCR6) in Complex with CXCL16 Soluble form Chemokine by Molecular Dynamic Simulations: General Protein—Ligand Interaction Model and 3D-QSAR Studies of Synthetic Antagonists. Life 2021, 11, 346. [Google Scholar] [CrossRef]
- Wasilko, D.J.; Johnson, Z.L.; Ammirati, M.; Che, Y.; Griffor, M.C.; Han, S.; Wu, H. Structural basis for chemokine receptor CCR6 activation by the endogenous protein ligand CCL20. Nat. Commun. 2020, 11, 3031. [Google Scholar] [CrossRef]
- Dragan, P.; Merski, M.; Wiśniewski, S.; Sanmukh, S.G.; Latek, D. Chemokine Receptors-Structure-Based Virtual Screening Assisted by Machine Learning. Pharmaceutics 2023, 15, 516. [Google Scholar] [CrossRef]
- Yu, X.; Orr, C.M.; Chan, H.T.C.; James, S.; Penfold, C.A.; Kim, J.; Inzhelevskaya, T.; Mockridge, C.I.; Cox, K.L.; Essex, J.W.; et al. Reducing affinity as a strategy to boost immunomodulatory antibody agonism. Nature 2023, 614, 539–547. [Google Scholar] [CrossRef]
- Frick, V.O.; Rubie, C.; Keilholz, U.; Ghadjar, P. Chemokine/chemokine receptor pair CCL20/CCR6 in human colorectal malignancy: An overview. World J. Gastroenterol. 2016, 22, 833–841. [Google Scholar] [CrossRef] [PubMed]
- Isoda, Y.; Tanaka, T.; Suzuki, H.; Asano, T.; Yoshikawa, T.; Kitamura, K.; Kudo, Y.; Ejima, R.; Ozawa, K.; Kaneko, M.K.; et al. Epitope Mapping Using the Cell-Based 2 × Alanine Substitution Method About the Anti-mouse CXCR6 Monoclonal Antibody, Cx(6)Mab-1. Monoclon. Antib. Immunodiagn. Immunother. 2023, 42, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Korbecki, J.; Kojder, K.; Simińska, D.; Bohatyrewicz, R.; Gutowska, I.; Chlubek, D.; Baranowska-Bosiacka, I. CC Chemokines in a Tumor: A Review of Pro-Cancer and Anti-Cancer Properties of the Ligands of Receptors CCR1, CCR2, CCR3, and CCR4. Int. J. Mol. Sci. 2020, 21, 8412. [Google Scholar] [CrossRef] [PubMed]
- Rutihinda, C.; Haroun, R.; Saidi, N.E.; Ordoñez, J.P.; Naasri, S.; Lévesque, D.; Boisvert, F.M.; Fortier, P.H.; Belzile, M.; Fradet, L.; et al. Inhibition of the CCR6-CCL20 axis prevents regulatory T cell recruitment and sensitizes head and neck squamous cell carcinoma to radiation therapy. Cancer Immunol. Immunother. 2023, 72, 1089–1102. [Google Scholar] [CrossRef]
- Jia, S.N.; Han, Y.B.; Yang, R.; Yang, Z.C. Chemokines in colon cancer progression. Semin. Cancer Biol. 2022, 86, 400–407. [Google Scholar] [CrossRef]
- Chen, K.J.; Lin, S.Z.; Zhou, L.; Xie, H.Y.; Zhou, W.H.; Taki-Eldin, A.; Zheng, S.S. Selective recruitment of regulatory T cell through CCR6-CCL20 in hepatocellular carcinoma fosters tumor progression and predicts poor prognosis. PLoS ONE 2011, 6, e24671. [Google Scholar] [CrossRef]
- Jin, L.; Cao, L.; Zhu, Y.; Cao, J.; Li, X.; Zhou, J.; Liu, B.; Zhao, T. Enhance anti-lung tumor efficacy of chimeric antigen receptor-T cells by ectopic expression of C-C motif chemokine receptor 6. Sci. Bull. 2021, 66, 803–812. [Google Scholar] [CrossRef]
- Zhong, C.; Chen, J. CAR-T cell engineering with CCR6 exhibits superior anti-solid tumor efficacy. Sci. Bull. 2021, 66, 755–756. [Google Scholar] [CrossRef]
- Jeon, S.H.; Kang, M.; Jeon, M.; Chung, Y.; Kim, A.R.; Lee, Y.J.; Kim, E.S.; Nam, H.; Park, J.; Lee, J.Y.; et al. CEACAM1 marks highly suppressive intratumoral regulatory T cells for targeted depletion therapy. Clin. Cancer Res. 2023, in press. [CrossRef]


{kind=link}
| Peptides | Sequences | C6Mab-13 Reactivity |
|---|---|---|
| p1–20 (WT) | MNSTESYFGTDDYDNTEYYS | +++ |
| M1A | ANSTESYFGTDDYDNTEYYS | +++ |
| N2A | MASTESYFGTDDYDNTEYYS | +++ |
| S3A | MNATESYFGTDDYDNTEYYS | +++ |
| T4A | MNSAESYFGTDDYDNTEYYS | +++ |
| E5A | MNSTASYFGTDDYDNTEYYS | +++ |
| S6A | MNSTEAYFGTDDYDNTEYYS | +++ |
| Y7A | MNSTESAFGTDDYDNTEYYS | +++ |
| F8A | MNSTESYAGTDDYDNTEYYS | +++ |
| G9A | MNSTESYFATDDYDNTEYYS | +++ |
| T10A | MNSTESYFGADDYDNTEYYS | +++ |
| D11A | MNSTESYFGTADYDNTEYYS | - |
| D12A | MNSTESYFGTDAYDNTEYYS | +++ |
| Y13A | MNSTESYFGTDDADNTEYYS | +++ |
| D14A | MNSTESYFGTDDYANTEYYS | +++ |
| N15A | MNSTESYFGTDDYDATEYYS | +++ |
| T16A | MNSTESYFGTDDYDNAEYYS | +++ |
| E17A | MNSTESYFGTDDYDNTAYYS | +++ |
| Y18A | MNSTESYFGTDDYDNTEAYS | +++ |
| Y19A | MNSTESYFGTDDYDNTEYAS | +++ |
| S20A | MNSTESYFGTDDYDNTEYYA | +++ |
| Peptides | ka (/ms) | kd (/s) | KD (M) |
|---|---|---|---|
| p1_20 (WT) | 6.84 × 103 | 3.77 × 10−3 | 5.52 × 10−7 |
| M1A | 6.94 × 103 | 4.15 × 10−3 | 5.99 × 10−7 |
| N2A | 7.86 × 103 | 4.23 × 10−3 | 5.38 × 10−7 |
| S3A | 7.62 × 103 | 4.53 × 10−3 | 5.94 × 10−7 |
| T4A | 7.92 × 103 | 4.55 × 10−3 | 5.75 × 10−7 |
| E5A | 8.20 × 103 | 4.64 × 10−3 | 5.65 × 10−7 |
| S6A | 9.05 × 103 | 5.25 × 10−3 | 5.81 × 10−7 |
| Y7A | 8.16 × 103 | 3.45 × 10−3 | 4.23 × 10−7 |
| F8A | 1.43 × 103 | 1.23 × 10−2 | 8.55 × 10−6 |
| G9A | ND | ND | ND |
| T10A | 1.31 × 104 | 3.15 × 10−2 | 2.40 × 10−6 |
| D11A | ND | ND | ND |
| D12A | 7.43 × 103 | 7.09 × 10−3 | 9.55 × 10−7 |
| Y13A | 1.43 × 103 | 1.30 × 10−2 | 9.12 × 10−6 |
| D14A | 6.87 × 103 | 1.05 × 10−2 | 1.53 × 10−6 |
| N15A | 6.19 × 103 | 5.61 × 10−3 | 9.06 × 10−7 |
| T16A | 6.23 × 103 | 5.17 × 10−3 | 8.30 × 10−7 |
| E17A | 6.38 × 103 | 6.67 × 10−3 | 1.05 × 10−6 |
| Y18A | 5.23 × 103 | 5.56 × 10−3 | 1.06 × 10−6 |
| Y19A | 5.75 × 103 | 6.02 × 10−3 | 1.05 × 10−6 |
| S20A | 4.68 × 103 | 5.96 × 10−3 | 1.27 × 10−6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tanaka, T.; Tawara, M.; Suzuki, H.; Kaneko, M.K.; Kato, Y. Identification of the Binding Epitope of an Anti-Mouse CCR6 Monoclonal Antibody (C6Mab-13) Using 1× Alanine Scanning. Antibodies 2023, 12, 32. https://doi.org/10.3390/antib12020032
Tanaka T, Tawara M, Suzuki H, Kaneko MK, Kato Y. Identification of the Binding Epitope of an Anti-Mouse CCR6 Monoclonal Antibody (C6Mab-13) Using 1× Alanine Scanning. Antibodies. 2023; 12(2):32. https://doi.org/10.3390/antib12020032
Chicago/Turabian StyleTanaka, Tomohiro, Mayuki Tawara, Hiroyuki Suzuki, Mika K. Kaneko, and Yukinari Kato. 2023. "Identification of the Binding Epitope of an Anti-Mouse CCR6 Monoclonal Antibody (C6Mab-13) Using 1× Alanine Scanning" Antibodies 12, no. 2: 32. https://doi.org/10.3390/antib12020032