
Defensin Interactions in Relation to Monoclonal and Disease-Related Proteinase 3 Antibodies Binding at the Catalytic Site
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. PR3 Purification
2.3. SDS-PAGE
2.4. Western Blotting
2.5. PR3 ELISA
2.6. PR3 Capture ELISA
2.7. PR3 Proteolytic Activity and Inhibition of Antibody-Antigen Binding
2.8. Peptide CovaLink-NH ELISA
2.9. Surface Plasmon Resonance (SPR) Assays
2.10. PR3 Ab Sequences
2.11. Amino Acid Analysis (AAA)
2.12. Mass Spectrometry (MS)
3. Results
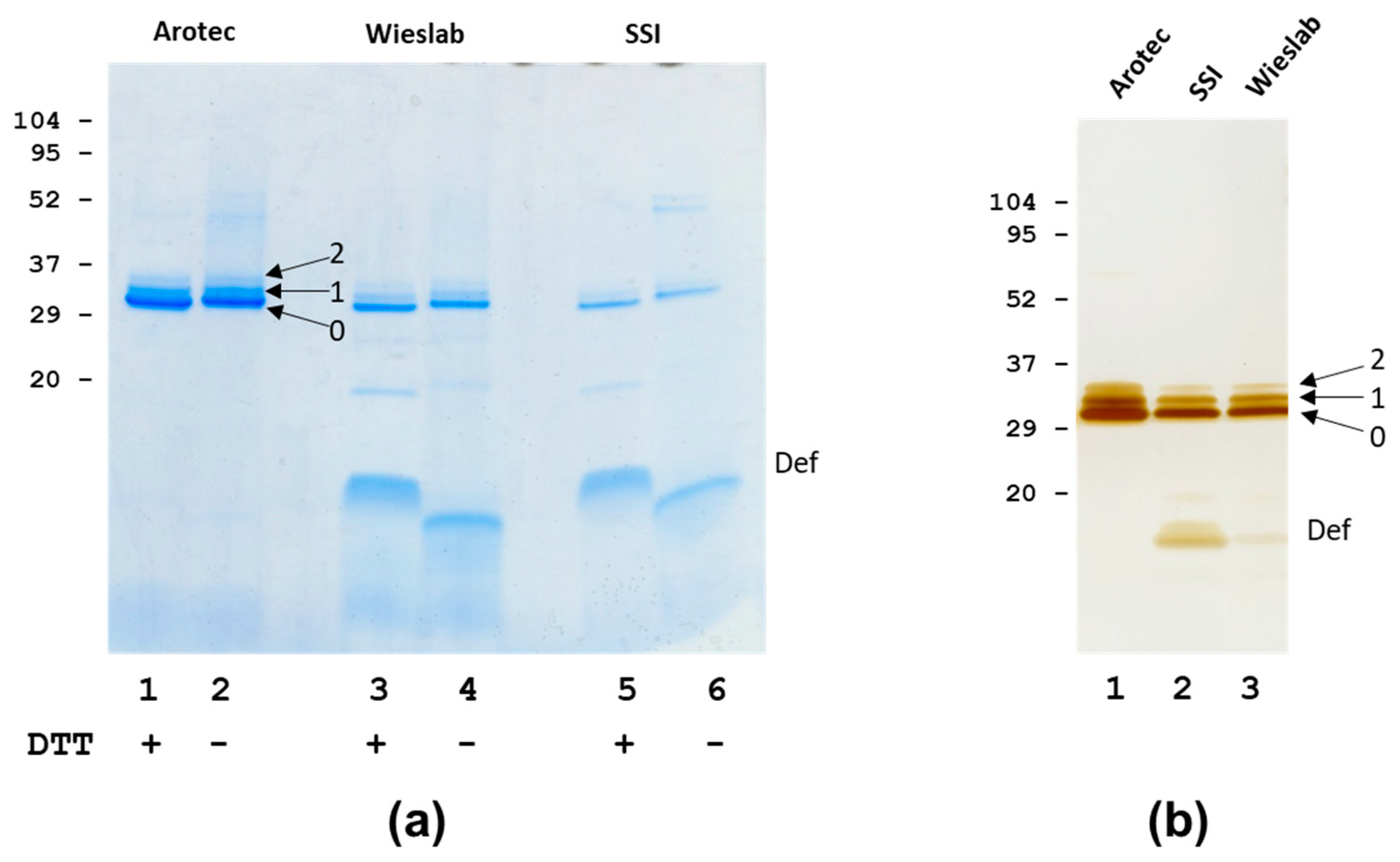
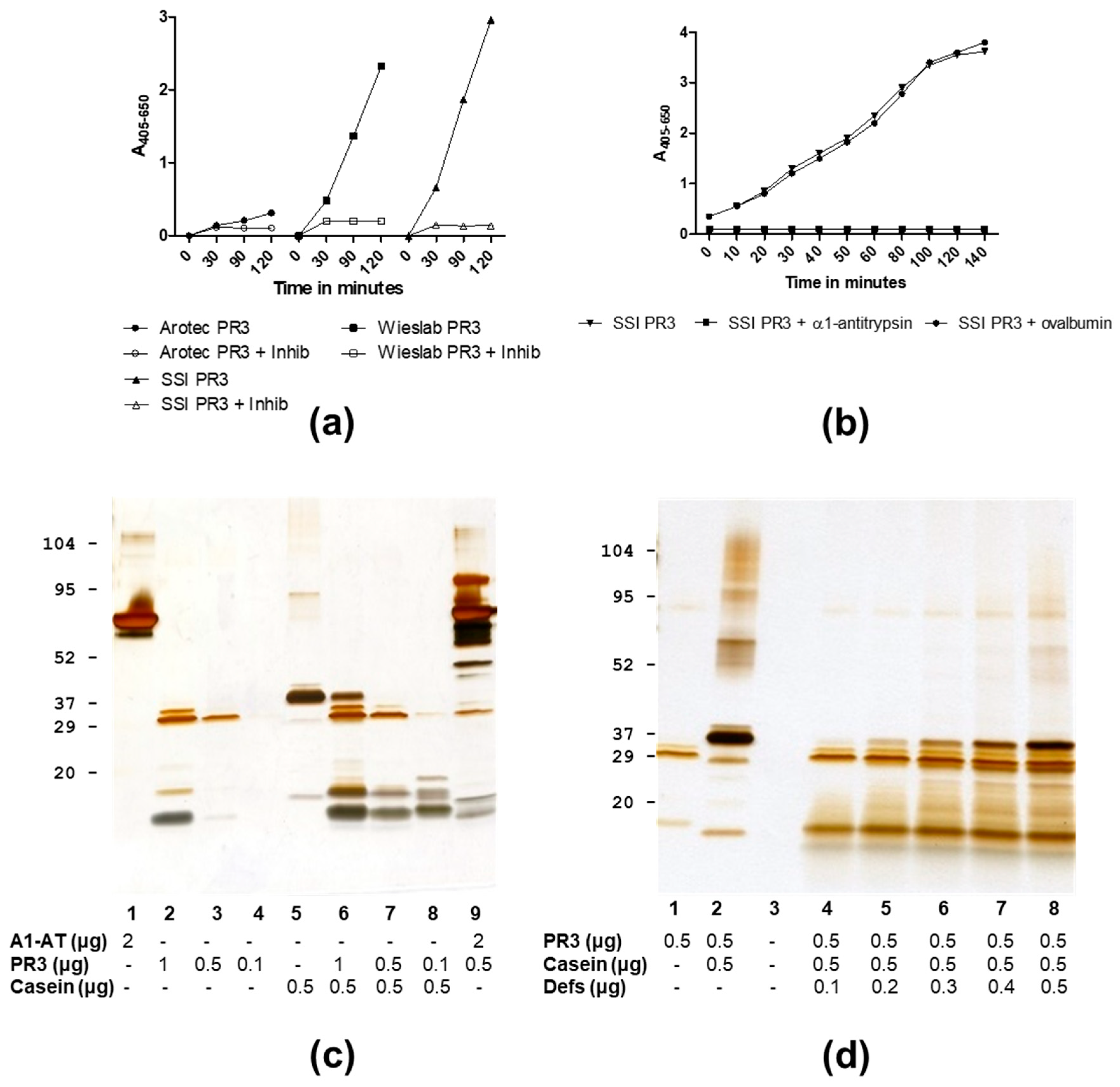
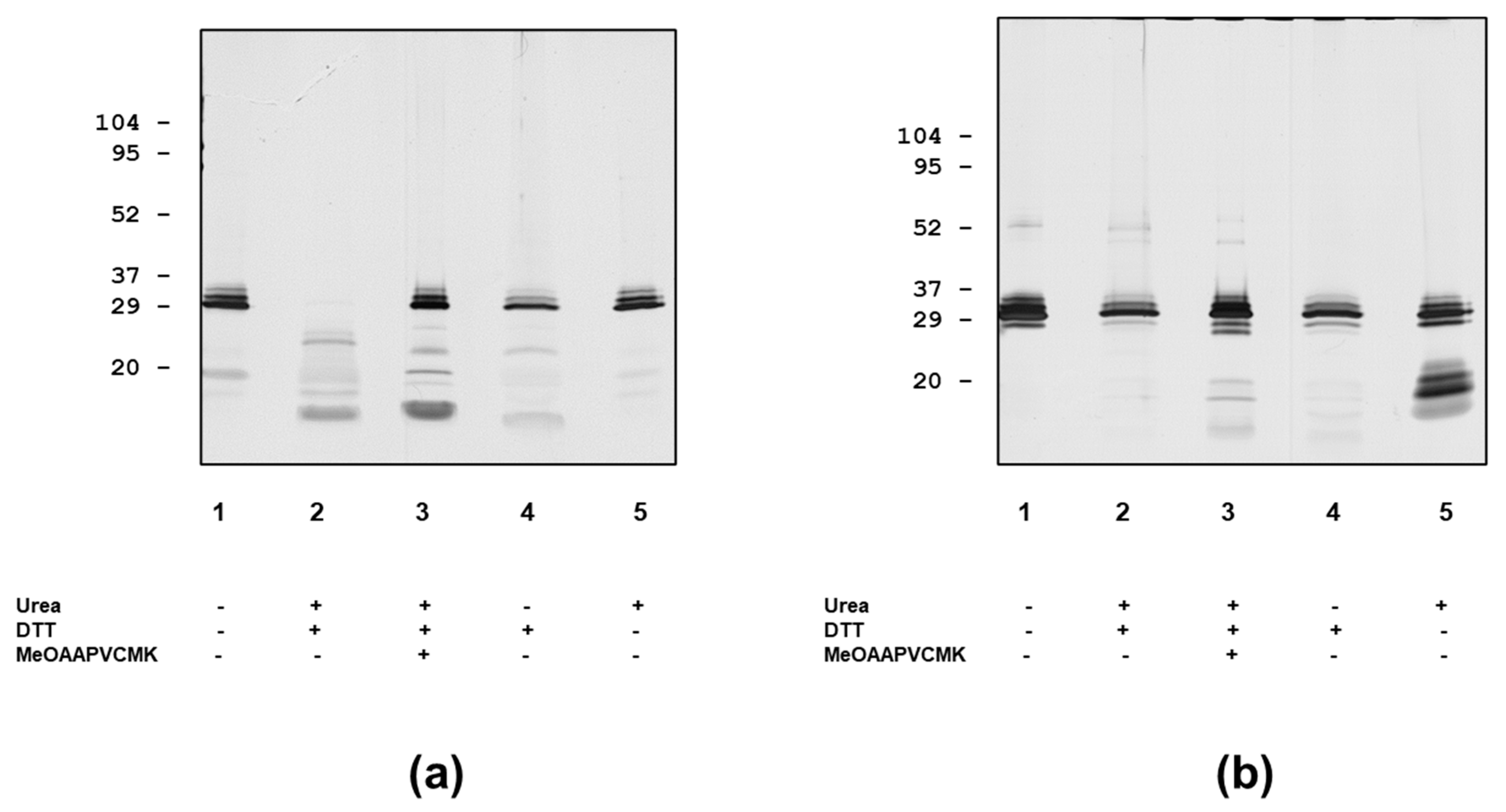
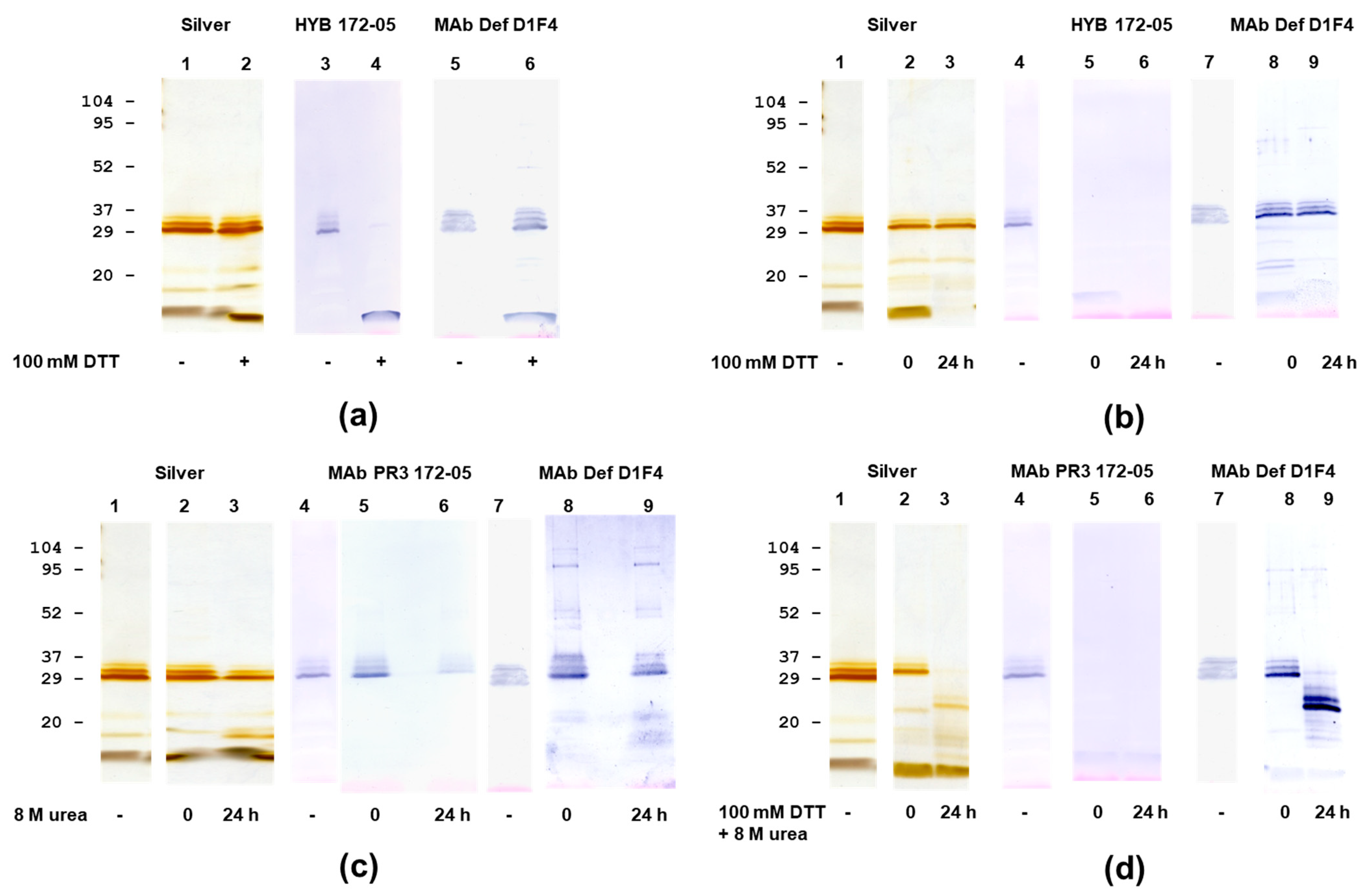
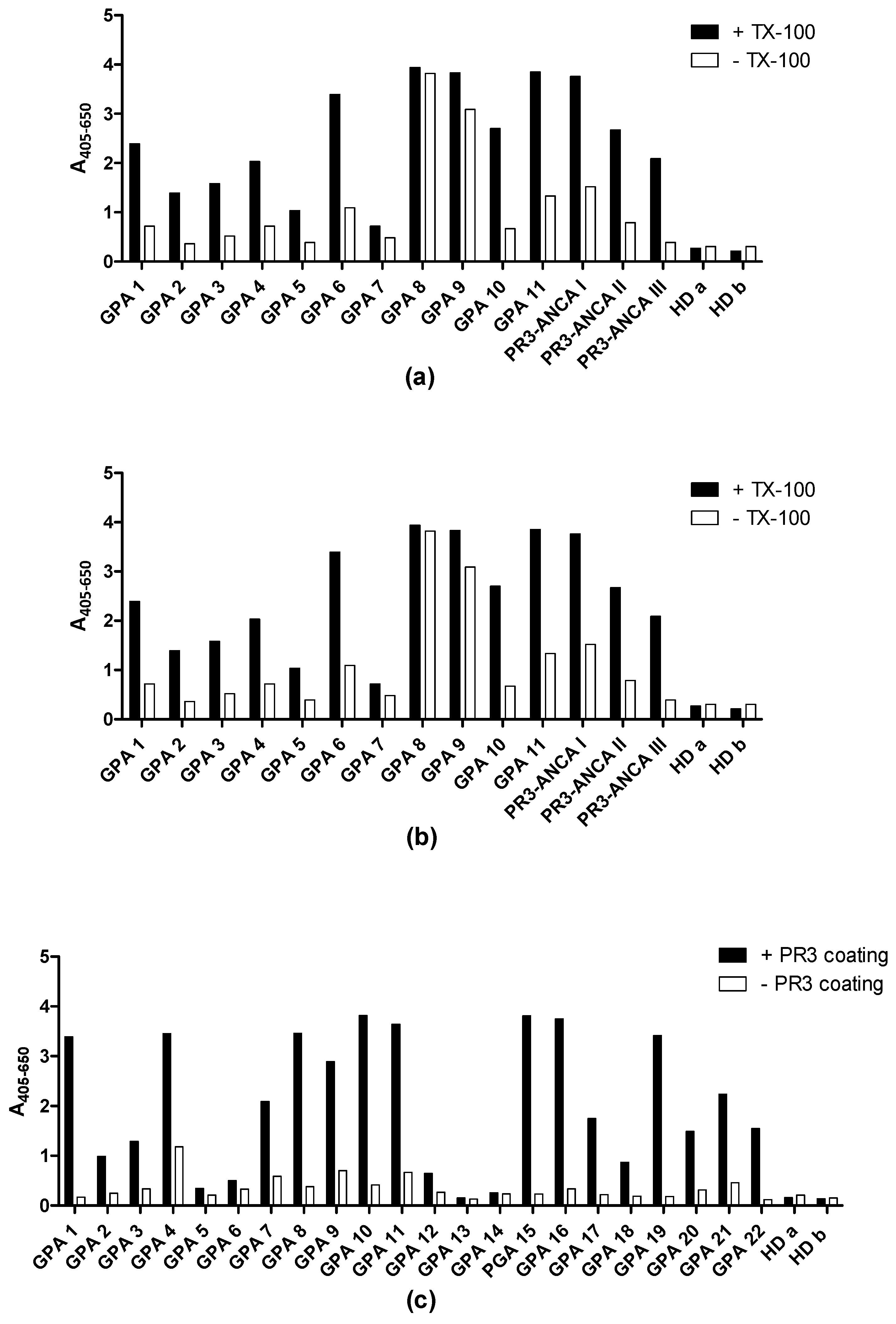
3.1. Analysis of PR3 Preparations
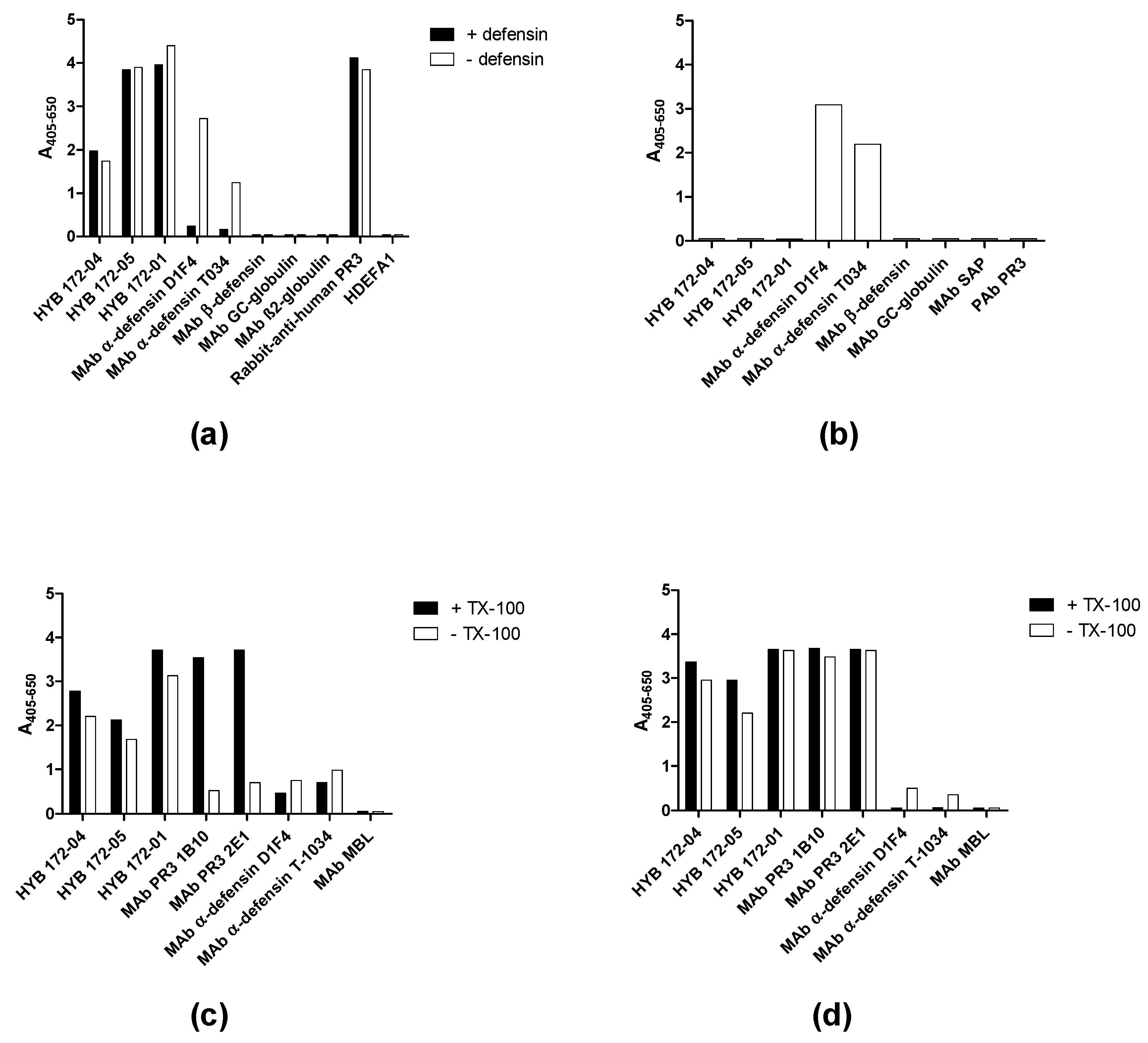
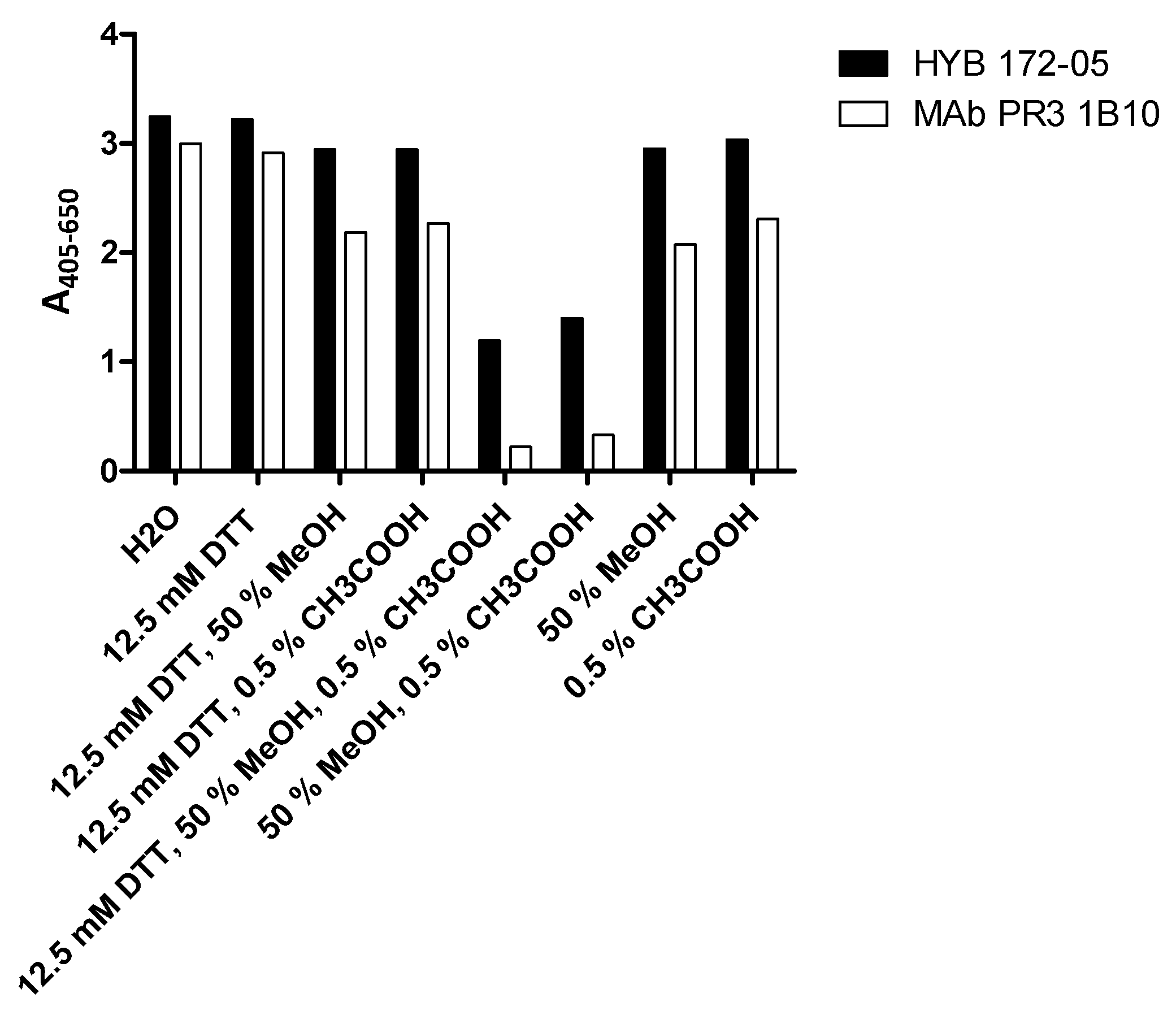
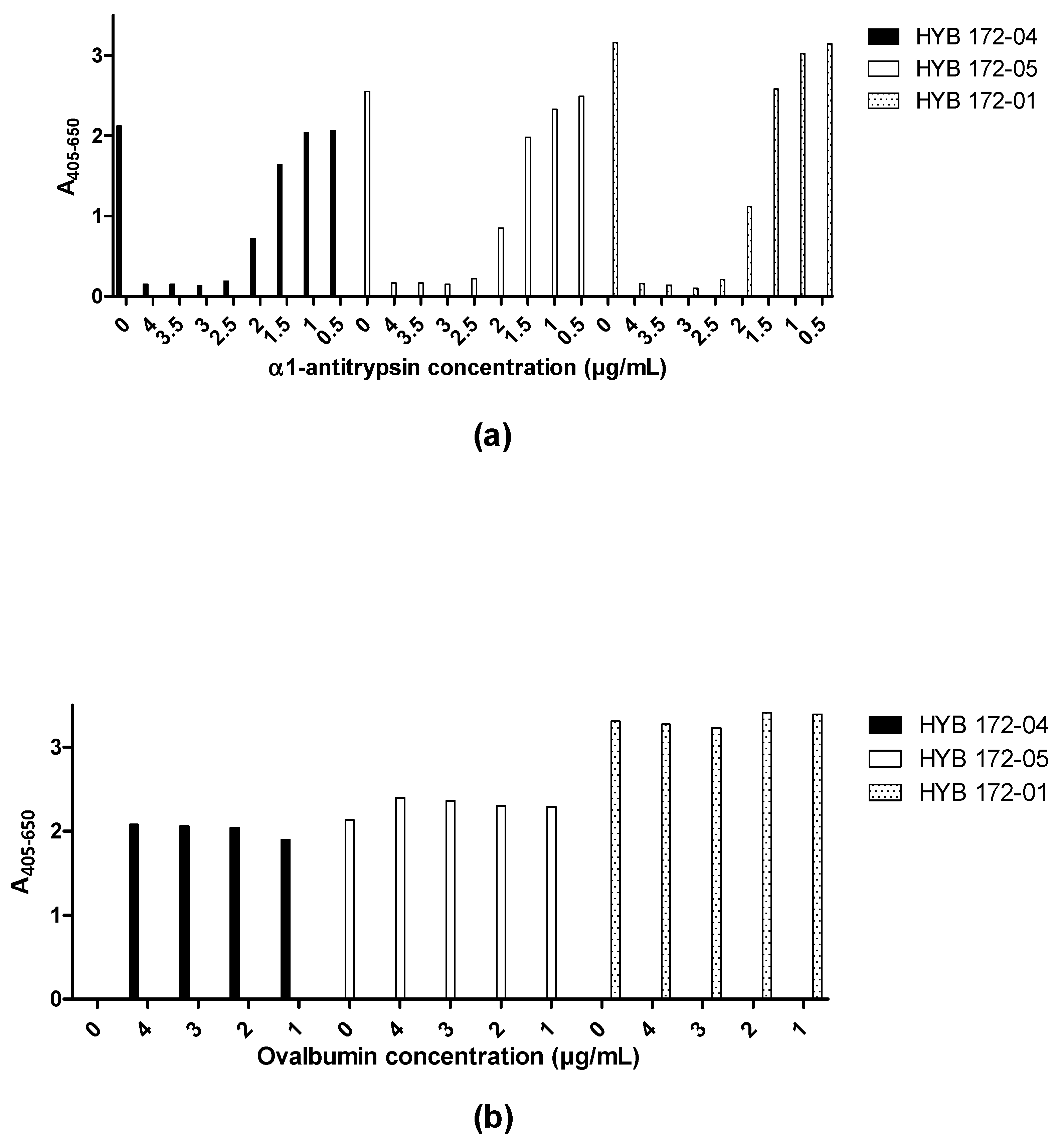
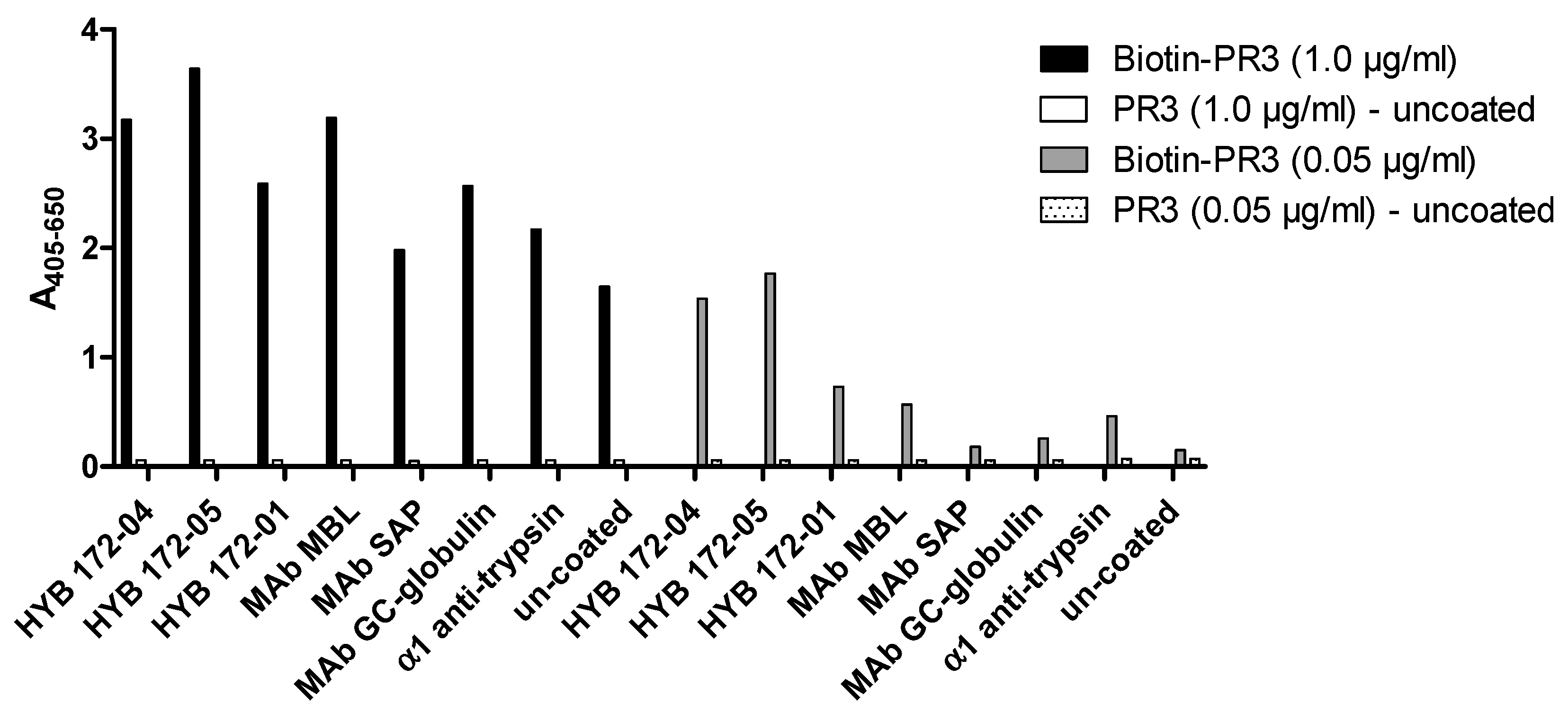
3.2. Characterisation of PR3 MAbs
3.3. PR3 Defensin Association
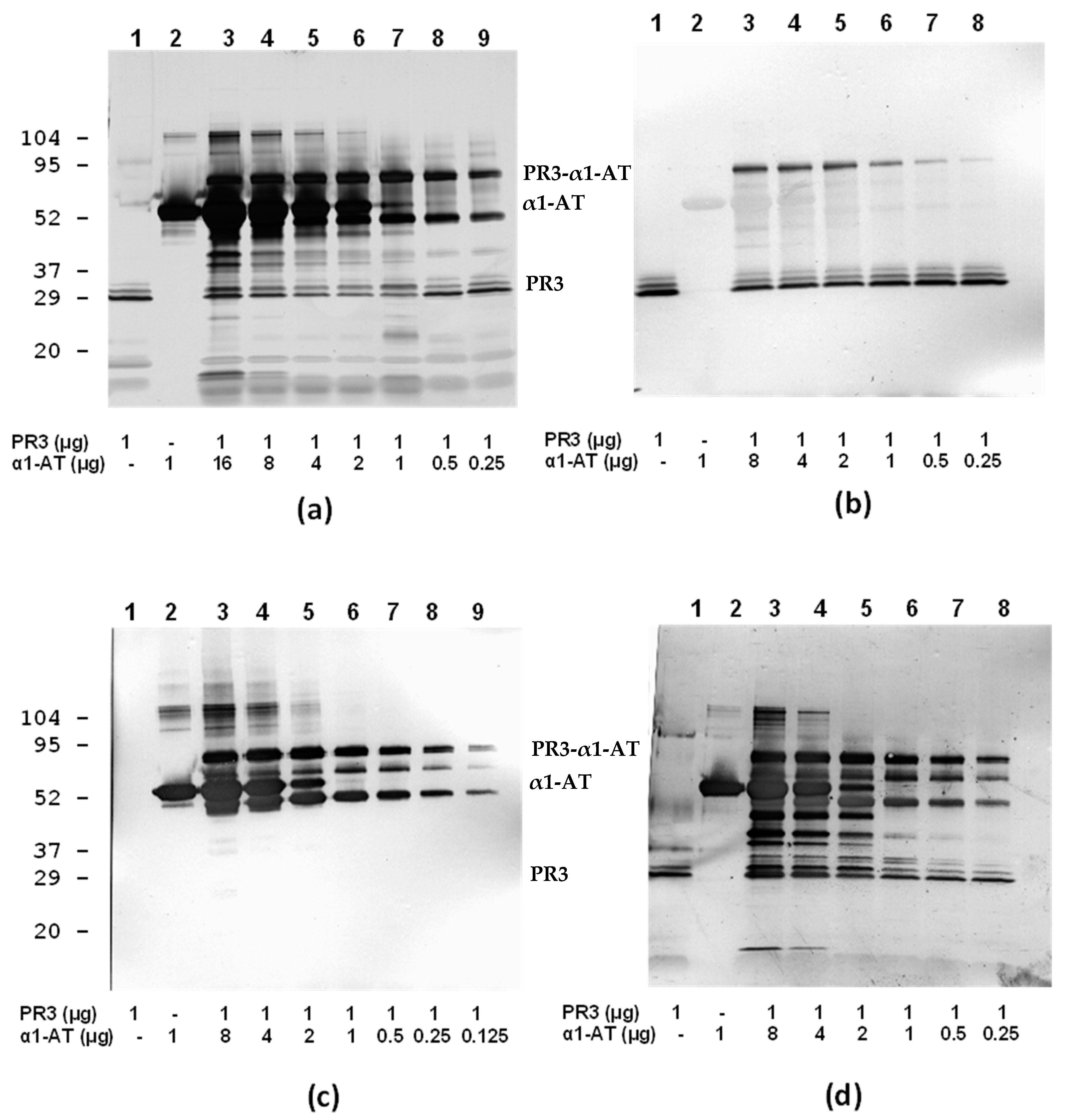
3.4. Inhibition of PR3 MAb Interactions
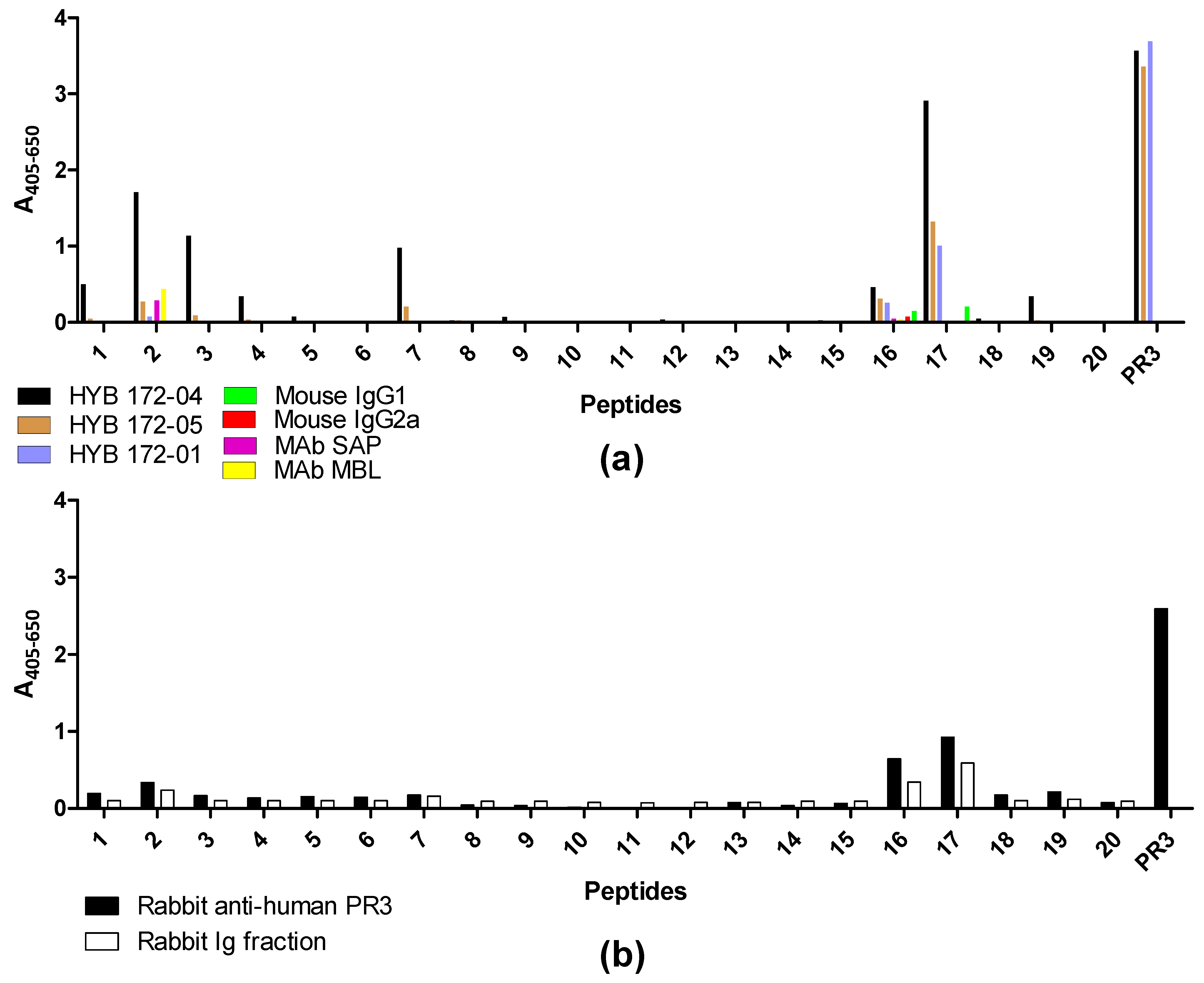
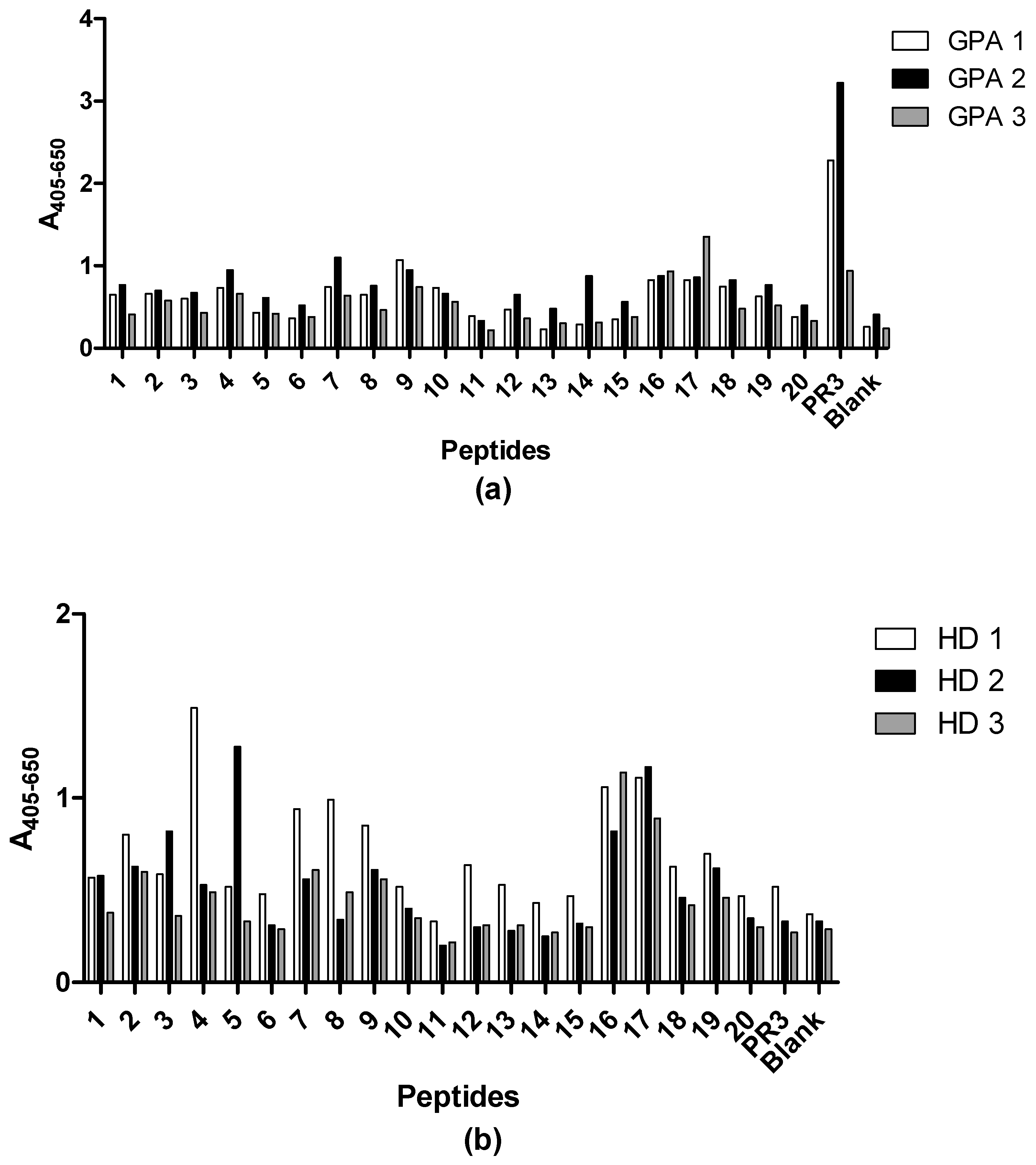
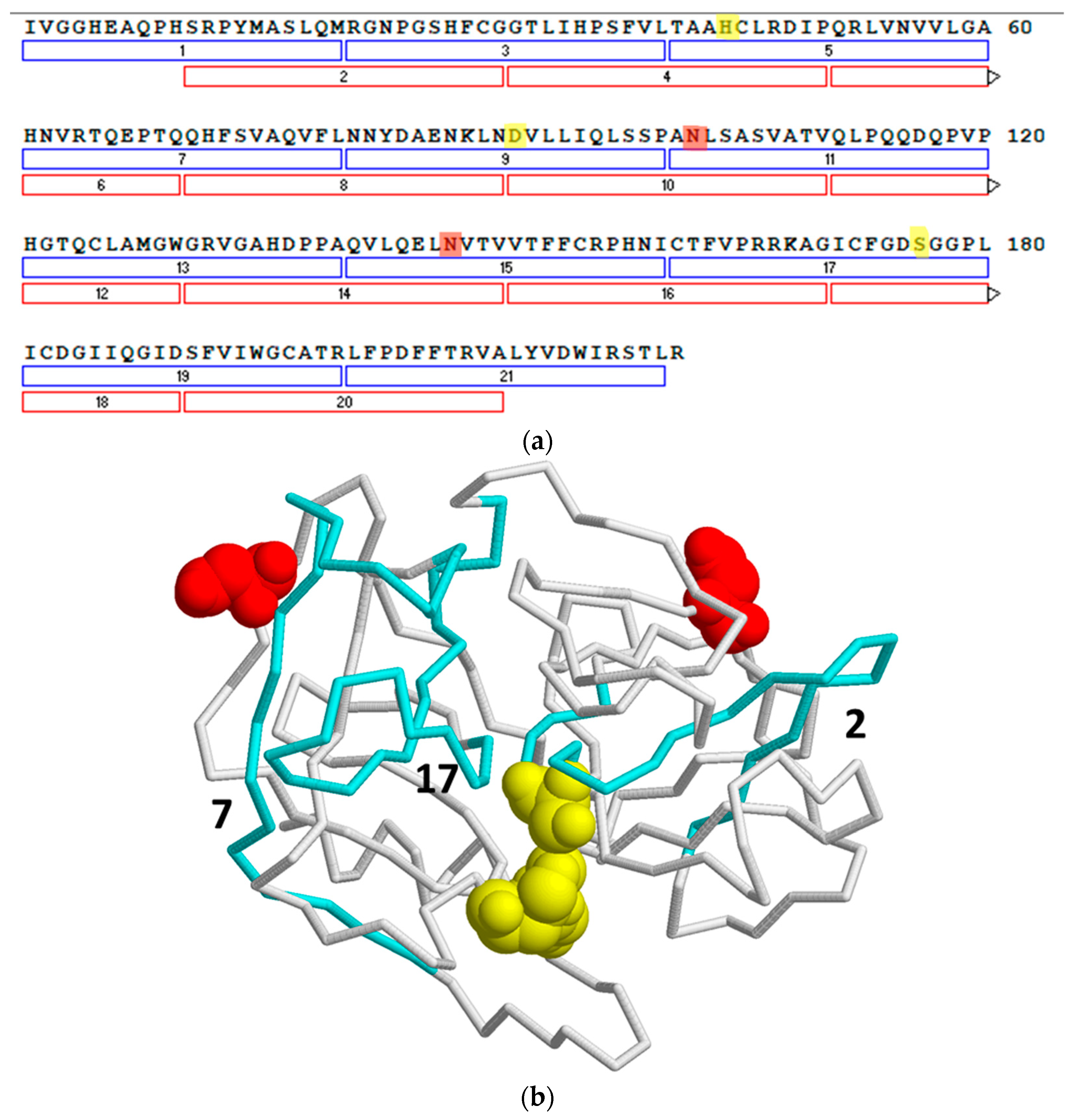
3.5. Epitope Mapping with Proteinase 3-Derived Peptides
3.6. Monoclonal Antibody Sequences/Modelling
3.7. Immuno Assays for PR3 Autoantibodies
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cowland, J.B.; Borregaard, N. Granulopoiesis and granules of human neutrophils. Immunol. Rev. 2016, 273, 11–28. [Google Scholar] [CrossRef] [PubMed]
- Korkmaz, B.; Moreau, T.; Gauthier, F. Neutrophil elastase, proteinase 3 and cathepsin G: Physicochemical properties, activity and physiopathological functions. Biochimie 2007, 90, 227–242. [Google Scholar] [CrossRef] [PubMed]
- Thieblemont, N.; Wright, H.L.; Edwards, S.W.; Witko-Sarsat, V. Human neutrophils in autoimmunity. Semin. Immunol. 2016, 28, 159–173. [Google Scholar] [CrossRef] [PubMed]
- Weiner, M.; Segelmark, M. The clinical presentation and therapy of diseases related to anti-neutrophil cytoplasmic antibodies (ANCA). Autoimmun. Rev. 2016, 15, 978–982. [Google Scholar] [CrossRef] [Green Version]
- Jennette, J.C. Nomenclature and classification of vasculitis: Lessons learned from granulomatosis with polyangiitis (Wegener’s granulomatosis). Clin. Exp. Immunol. 2011, 164 (Suppl. S1), 7–10. [Google Scholar] [CrossRef] [PubMed]
- Granel, J.; Korkmaz, B.; Nouar, D.; Weiss, S.A.I.; Jenne, D.E.; Lemoine, R.; Hoarau, C. Pathogenicity of proteinase 3-anti-neutrophil cytoplasmic antibody in granulomatosis with polyangiitis: Implications as biomarker and future therapies. Front. Immunol. 2021, 12, 571933. [Google Scholar] [CrossRef] [PubMed]
- van de Wiel, B.A.; Dolman, K.M.; van der Meer-Gerritsen, C.H.; Hack, C.E.; von dem Borne, A.E.; Goldschmeding, R. Interference of Wegener’s granulomatosis autoantibodies with neutrophil proteinase 3 activity. Clin. Exp. Immunol. 1992, 90, 409–414. [Google Scholar] [CrossRef]
- Dolman, K.M.; Stegeman, C.A.; van de Wiel, B.A.; Hack, C.E.; von dem Borne, A.E.; Kallenberg, C.G.; Goldschmeding, R. Relevance of classic anti-neutrophil cytoplasmic autoantibody (C-ANCA)-mediated inhibition of proteinase 3-alpha 1-antitrypsin complexation to disease activity in Wegener’s granulomatosis. Clin. Exp. Immunol. 1993, 93, 405–410. [Google Scholar] [CrossRef]
- Bini, P.; Gabay, J.E.; Teitel, A.; Melchior, M.; Zhou, J.L.; Elkon, K.B. Antineutrophil cytoplasmic autoantibodies in Wegener’s granulomatosis recognize conformational epitope(s) on proteinase 3. J. Immunol. 1992, 149, 1409–1415. [Google Scholar] [CrossRef]
- Hajjar, E.; Broemstrup, T.; Kantari, C.; Witko-Sarsat, V.; Reuter, N. Structures of human proteinase 3 and neutrophil elastase—So similar yet so different. FEBS J. 2010, 277, 2238–2254. [Google Scholar] [CrossRef]
- van der Geld, Y.M.; Limburg, P.C.; Kallenberg, C.G. Proteinase 3, Wegener’s autoantigen: From gene to antigen. J. Leukoc. Biol. 2001, 69, 177–190. [Google Scholar] [CrossRef] [PubMed]
- Campanelli, D.; Melchior, M.; Fu, Y.; Nakata, M.; Shuman, H.; Nathan, C.; Gabay, J.E. Cloning of cDNA for proteinase 3: A serine protease, antibiotic, and autoantigen from human neutrophils. J. Exp. Med. 1990, 172, 1709–1715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmer, M.; Medcalf, R.L.; Fink, T.M.; Mattmann, C.; Lichter, P.; Jenne, D.E. Three human elastase-like genes coordinately expressed in the myelomonocyte lineage are organized as a single genetic locus on 19pter. Proc. Natl. Acad. Sci. USA 1992, 89, 8215–8219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujinaga, M.; Chernaia, M.M.; Halenbeck, R.; Koths, K.; James, M.N. The crystal structure of PR3, a neutrophil serine proteinase antigen of Wegener’s granulomatosis antibodies. J. Mol. Biol. 1996, 261, 267–278. [Google Scholar] [CrossRef]
- Rao, N.V.; Wehner, N.G.; Marshall, B.C.; Gray, W.R.; Gray, B.H.; Hoidal, J.R. Characterization of proteinase-3 (PR-3), a neutrophil serine proteinase. Structural and functional properties. J. Biol. Chem. 1991, 266, 9540–9548. [Google Scholar] [CrossRef]
- Duranton, J.; Bieth, J.G. Inhibition of proteinase 3 by [alpha]1-antitrypsin in vitro predicts very fast inhibition in vivo. Am. J. Respir. Cell Mol. Biol. 2003, 29, 57–61. [Google Scholar] [CrossRef]
- Ballieux, B.E.; Hagen, E.C.; van der Keur, C.; Zegers, N.D.; van Es, L.A.; van der Woude, F.J.; Daha, M.R. Isolation of a protein complex from purulent sputum consisting of proteinase-3 and alpha 1-antitrypsin reactive with anti neutrophil cytoplasmic antibodies. J. Immunol. Methods 1993, 159, 63–70. [Google Scholar] [CrossRef]
- Chang, L.; Binos, S.; Savige, J. Epitope mapping of anti-proteinase 3 and anti-myeloperoxidase antibodies. Clin. Exp. Immunol. 1995, 102, 112–119. [Google Scholar] [CrossRef]
- Sun, J.; Fass, D.N.; Viss, M.A.; Hummel, A.M.; Tang, H.; Homburger, H.A.; Specks, U. A proportion of proteinase 3 (PR3)-specific anti-neutrophil cytoplasmic antibodies (ANCA) only react with PR3 after cleavage of its N-terminal activation dipeptide. Clin. Exp. Immunol. 1998, 114, 320–326. [Google Scholar] [CrossRef]
- Specks, U.; Fass, D.N.; Finkielman, J.D.; Hummel, A.M.; Viss, M.A.; Litwiller, R.D.; McDonald, C.J. Functional significance of Asn-linked glycosylation of proteinase 3 for enzymatic activity, processing, targeting, and recognition by anti-neutrophil cytoplasmic antibodies. J. Biochem. 2007, 141, 101–112. [Google Scholar] [CrossRef]
- Finkielman, J.D.; Merkel, P.A.; Schroeder, D.; Hoffman, G.S.; Spiera, R.; St Clair, E.W.; Davies, J.C., Jr.; McCune, W.J.; Lears, A.; Ytterberg, S.R.; et al. Glycosylation of proteinase 3 (PR3) is not required for its reactivity with antineutrophil cytoplasmic antibodies (ANCA) in Wegener’s granulomatosis. Clin. Exp. Rheumatol. 2009, 27, S45–S52. [Google Scholar]
- Witko-Sarsat, V.; Halbwachs-Mecarelli, L.; Almeida, R.P.; Nusbaum, P.; Melchior, M.; Jamaleddine, G.; Lesavre, P.; Descamps-Latscha, B.; Gabay, J.E. Characterization of a recombinant proteinase 3, the autoantigen in Wegener’s granulomatosis and its reactivity with anti-neutrophil cytoplasmic autoantibodies. FEBS Lett. 1996, 382, 130–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Geld, Y.M.; Oost-Kort, W.; Limburg, P.C.; Specks, U.; Kallenberg, C.G. Recombinant proteinase 3 produced in different expression systems: Recognition by anti-PR3 antibodies. J. Immunol. Methods 2000, 244, 117–131. [Google Scholar] [CrossRef] [PubMed]
- Harmsen, M.C.; Heeringa, P.; van der Geld, Y.M.; Huitema, M.G.; Klimp, A.; Tiran, A.; Kallenberg, C.G. Recombinant proteinase 3 (Wegener’s antigen) expressed in Pichia pastoris is functionally active and is recognized by patient sera. Clin. Exp. Immunol. 1997, 110, 257–264. [Google Scholar] [CrossRef]
- Szymkowiak, C.H.; Johnston, T.W.; Csernok, E.; Gross, W.L. Expression of the human autoantigen of Wegener’s granulomatosis (PR3) in a baculovirus expression system. Biochem. Biophys. Res. Commun. 1996, 219, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Specks, U.; Wiegert, E.M.; Homburger, H.A. Human mast cells expressing recombinant proteinase 3 (PR3) as substrate for clinical testing for anti-neutrophil cytoplasmic antibodies (ANCA). Clin. Exp. Immunol. 1997, 109, 286–295. [Google Scholar] [CrossRef] [PubMed]
- Silva, F.; Hummel, A.M.; Jenne, D.E.; Specks, U. Discrimination and variable impact of ANCA binding to different surface epitopes on proteinase 3, the Wegener’s autoantigen. J. Autoimmun. 2010, 35, 299–308. [Google Scholar] [CrossRef] [Green Version]
- Selga, D.; Segelmark, M.; Wieslander, J.; Gunnarsson, L.; Hellmark, T. Epitope mapping of anti-PR3 antibodies using chimeric human/mouse PR3 recombinant proteins. Clin. Exp. Immunol. 2004, 135, 164–172. [Google Scholar] [CrossRef] [Green Version]
- Kuhl, A.; Korkmaz, B.; Utecht, B.; Kneipert, A.; Schönermarck, U.; Specks, U.; Jenne, D.E. Mapping of conformational epitopes on human proteinase 3, the autoantigen of Wegener’s granulomatosis. J. Immunol. 2010, 185, 387–399. [Google Scholar] [CrossRef] [Green Version]
- Bruner, B.F.; Vista, E.S.; Wynn, D.M.; Harley, J.B.; James, J.A. Anti-neutrophil cytoplasmic antibodies target sequential functional proteinase 3 epitopes in the sera of patients with Wegener’s granulomatosis. Clin. Exp. Immunol. 2010, 162, 262–270. [Google Scholar] [CrossRef]
- Williams, R.C., Jr.; Staud, R.; Malone, C.C.; Payabyab, J.; Byres, L.; Underwood, D. Epitopes on proteinase-3 recognized by antibodies from patients with Wegener’s granulomatosis. J. Immunol. 1994, 152, 4722–4737. [Google Scholar] [CrossRef]
- Griffith, M.E.; Coulthart, A.; Pemberton, S.; George, A.J.; Pusey, C.D. Anti-neutrophil cytoplasmic antibodies (ANCA) from patients with systemic vasculitis recognize restricted epitopes of proteinase 3 involving the catalytic site. Clin. Exp. Immunol. 2001, 123, 170–177. [Google Scholar] [CrossRef]
- van der Geld, Y.M.; Simpelaar, A.; Van Der Zee, R.; Tervaert, J.W.; Stemgeman, C.A.; Limberg, P.C.; Kallenberg, C.G. Antineutrophil cytoplasmic antibodies to proteinase 3 in Wegener’s granulomatosis: Epitope analysis using synthetic peptides. Kidney Int. 2001, 59, 147–159. [Google Scholar] [CrossRef] [Green Version]
- Sommarin, Y.; Rasmussen, N.; Wieslander, J. Characterization of monoclonal antibodies to proteinase-3 and application in the study of epitopes for classical anti-neutrophil cytoplasm antibodies. Exp. Nephrol. 1995, 3, 249–256. [Google Scholar]
- van der Geld, Y.M.; Limburg, P.C.; Kallenberg, C.G. Characterization of monoclonal antibodies to proteinase 3 (PR3) as candidate tools for epitope mapping of human anti-PR3 autoantibodies. Clin. Exp. Immunol. 1999, 118, 487–496. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Fass, D.N.; Hudson, J.A.; Viss, M.A.; Wieslander, J.; Homburger, H.A.; Specks, U. Capture-ELISA based on recombinant PR3 is sensitive for PR3-ANCA testing and allows detection of PR3 and PR3-ANCA/PR3 immunecomplexes. J. Immunol. Methods 1998, 211, 111–123. [Google Scholar] [CrossRef] [PubMed]
- Hinkofer, L.C.; Seidel, S.A.; Korkmaz, B.; Silva, F.; Hummel, A.M.; Braun, D.; Jenne, D.E.; Specks, U. A monoclonal antibody (MCPR3-7) interfering with the activity of proteinase 3 by an allosteric mechanism. J. Biol. Chem. 2013, 288, 26635–26648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zoega, M.; Ravnsborg, T.; Højrup, P.; Houen, G.; Schou, C. Proteinase 3 carries small unusual carbohydrates and associates with α-defensins. J. Proteom. 2012, 75, 1472–1485. [Google Scholar] [CrossRef] [PubMed]
- Trier, N.H.; Mortensen, A.; Schiolborg, A.; Friis, T. Production and Screening of Monoclonal Peptide Antibodies. Methods Mol. Biol. 2015, 1348, 109–126. [Google Scholar] [CrossRef]
- Stummann, L.; Wiik, A. A simple high yield procedure for purification of human proteinase 3, the main molecular target of cANCA. J. Immunol. Methods 1997, 206, 35–42. [Google Scholar] [CrossRef]
- Heegaard, N.H.; Jakobsen, D.R.; Klattschou, D. Purification of Wegener’s granulomatosis autoantigen, proteinase 3, from neutrophils by Triton X-114 extraction of azurophilic granules. Anal. Biochem. 1997, 253, 259–262. [Google Scholar] [CrossRef] [PubMed]
- Shevchenko, A.; Wilm, M.; Vorm, O.; Mann, M. Mass spectrometric sequencing of proteins silver-stained polyacrylamide gels. Anal. Chem. 1996, 68, 850–858. [Google Scholar] [CrossRef] [PubMed]
- Højrup, P. Analysis of peptides and conjugates by amino acid analysis. Meth. Mol. Biol. 2015, 1348, 65–76. [Google Scholar]
- Ganz, T. Defensins: Antimicrobial peptides of innate immunity. Nat. Rev. Immunol. 2003, 3, 710–720. [Google Scholar] [CrossRef]
- Schneider, J.J.; Unholzer, A.; Schaller, M.; Schäfer-Korting, M.; Korting, H.C. Human defensins. J. Mol. Med. 2005, 83, 587–595. [Google Scholar] [CrossRef]
- Lehrer, R.I.; Lu, W. α-Defensins in human innate immunity. Immunol. Rev. 2012, 245, 84–112. [Google Scholar] [CrossRef]
- Ravnsborg, T.; Houen, G.; Højrup, P. The glycosylation of myeloperoxidase. Biochim. Et Biophys. Acta (BBA)—Proteins Proteom. 2010, 1804, 2046–2053. [Google Scholar] [CrossRef]
- Loke, I.; Østergaard, O.; Heegaard, N.H.; Packer, N.H.; Thaysen-Andersen, M. Paucimannose-Rich N-glycosylation of spatiotemporally regulated human neutrophil elastase modulates its immune functions. Mol. Cell. Proteomics 2017, 16, 1507–1527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiik, A.; Rasmussen, N.; Wieslander, J. Methods to detect autoantibodies to neutrophilic granulocytes. In Manual of Biological Markers of Disease; van Venrooij, W.J., Maini, R.N., Eds.; Springer: Dordrecht, The Netherlands, 1993. [Google Scholar]
- Radice, A.; Sinico, R.A. Antineutrophil cytoplasmic antibodies (ANCA). Autoimmunity 2005, 38, 93–103. [Google Scholar] [CrossRef]
- Rasmussen, N.; Wiik, A.; Jayne, D.R. A historical essay on detection of anti-neutrophil cytoplasmic antibodies. Nephrol. Dial. Transplant. 2015, 30 (Suppl. S1), i8–i13. [Google Scholar] [CrossRef] [Green Version]
- Lehrer, R.I.; Jung, G.; Ruchala, P.; Andre, S.; Gabius, H.J.; Lu, W. Multivalent binding of carbohydrates by the human alpha-defensin, HD5. J. Immunol. 2009, 183, 480–490. [Google Scholar] [CrossRef] [Green Version]
- Gooptu, B.; Lomas, D.A. Conformational pathology of the serpins: Themes, variations, and therapeutic strategies. Annu. Rev. Biochem. 2009, 78, 147–176. [Google Scholar] [CrossRef]
- Dobó, J.; Gettins, P.G. alpha1-Proteinase inhibitor forms initial non-covalent and final covalent complexes with elastase analogously to other serpin-proteinase pairs, suggesting a common mechanism of inhibition. J. Biol. Chem. 2004, 279, 9264–9269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dementiev, A.; Dobó, J.; Gettins, P.G. Active site distortion is sufficient for proteinase inhibition by serpins: Structure of the covalent complex of alpha1-proteinase inhibitor with porcine pancreatic elastase. J. Biol. Chem. 2006, 281, 3452–3457. [Google Scholar] [CrossRef] [Green Version]
- Stratikos, E.; Gettins, P.G. Formation of the covalent serpin-proteinase complex involves translocation of the proteinase by more than 70 A and full insertion of the reactive center loop into beta-sheet A. Proc. Natl. Acad. Sci. USA 1999, 96, 4808–4813. [Google Scholar] [CrossRef] [Green Version]
- Huntington, J.A.; Stein, P.E. Structure and properties of ovalbumin. J. Chromatogr. B Biomed. Sci. Appl. 2001, 756, 189–198. [Google Scholar] [CrossRef]
- Lee, K.N.; Im, H.; Kang, S.W.; Yu, M.H. Characterization of a human alpha1-antitrypsin variant that is as stable as ovalbumin. J. Biol. Chem. 1998, 273, 2509–2516. [Google Scholar] [CrossRef] [Green Version]
- van der Geld, Y.M.; Tool, A.T.; Videler, J.; de Haas, M.; Cohen Tervaert, J.W.; Stegeman, C.A.; Limburg, P.C.; Kallenberg, C.G.M.; Roos, D. Interference of PR3-ANCA with the enzymatic activity of PR3: Differences in patients during active disease or remission of Wegener’s granulomatosis. Clin. Exp. Immunol. 2002, 129, 562–570. [Google Scholar] [CrossRef]
- Dolman, K.M.; van de Wiel, B.A.; Kam, C.M.; Kerrigan, J.E.; Hack, C.E.; von dem Borne, A.E.; Powers, J.C.; Goldschmeding, R. Proteinase 3: Substrate specificity and possible pathogenetic effect of Wegener’s granulomatosis autoantibodies (c-ANCA) by dysregulation of the enzyme. Adv. Exp. Med. Biol. 1993, 336, 55–60. [Google Scholar] [PubMed]
- Daouk, G.H.; Palsson, R.; Arnaout, M.A. Inhibition of proteinase 3 by ANCA and its correlation with disease activity in Wegener’s granulomatosis. Kidney Int. 1995, 47, 1528–1536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arranz, O.; Ara, J.; Rodriguez, R.; Quintó, L.; Font, J.; Mirapeix, E.; Darnell, A. Comparison of anti-PR3 capture and anti-PR3 direct ELISA for detection of antineutrophil cytoplasmic antibodies (ANCA) in long-term clinical follow-up of PR3-ANCA-associated vasculitis patients. Clin. Nephrol. 2001, 56, 295–301. [Google Scholar]
- Csernok, E.; Holle, J.; Hellmich, B.; Willem, J.; Tervart, C.; Kallenberg, C.G.M.; Limburg, P.C.; Niles, J.; Pan, G.; Specks, U.; et al. Evaluation of capture ELISA for detection of antineutrophil cytoplasmic antibodies directed against proteinase 3 in Wegener’s granulomatosis: First results from a multicentre study. Rheumatol. 2004, 43, 174–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, A.S.; Finkielman, J.D.; Peikert, T.; Hummel, A.M.; Viss, M.A.; Specks, U. A novel capture-ELISA for detection of anti-neutrophil cytoplasmic antibodies (ANCA) based on c-myc peptide recognition in carboxy-terminally tagged recombinant neutrophil serine proteases. J. Immunol. Methods 2005, 307, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.; Liu, P.; Li, Z.; Sui, B. Clinical relevance of anti-PR3 capture ELISA in diagnosing Wegener’s granulomatosis. J. Clin. Lab. Anal. 2008, 22, 73–76. [Google Scholar] [CrossRef]
- Bossuyt, X.; Cohen Tervaert, J.W.; Arimura, Y.; Blockmans, D.; Flores-Suárez, L.F.; Guillevin, L.; Hellmich, B.; Jayne, D.; Jennette, J.C.; Kallenberg, C.G.M.; et al. Position paper: Revised 2017 international consensus on testing of ANCAs in granulomatosis with polyangiitis and microscopic polyangiitis. Nat. Rev. Rheumatol. 2017, 13, 683–692. [Google Scholar] [CrossRef] [PubMed]
- Damoiseaux, J.; Dähnrich, C.; Rosemann, A.; Probst, C.; Komorowski, L.; Stegeman, C.A.; Egerer, K.; Hiepe, F.; van Paassen, P.; Stöcker, W.; et al. A novel enzyme-linked immunosorbent assay using a mixture of human native and recombinant proteinase-3 significantly improves the diagnostic potential for antineutrophil cytoplasmic antibody-associated vasculitis. Ann. Rheum. Dis. 2009, 68, 228–233. [Google Scholar] [CrossRef]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
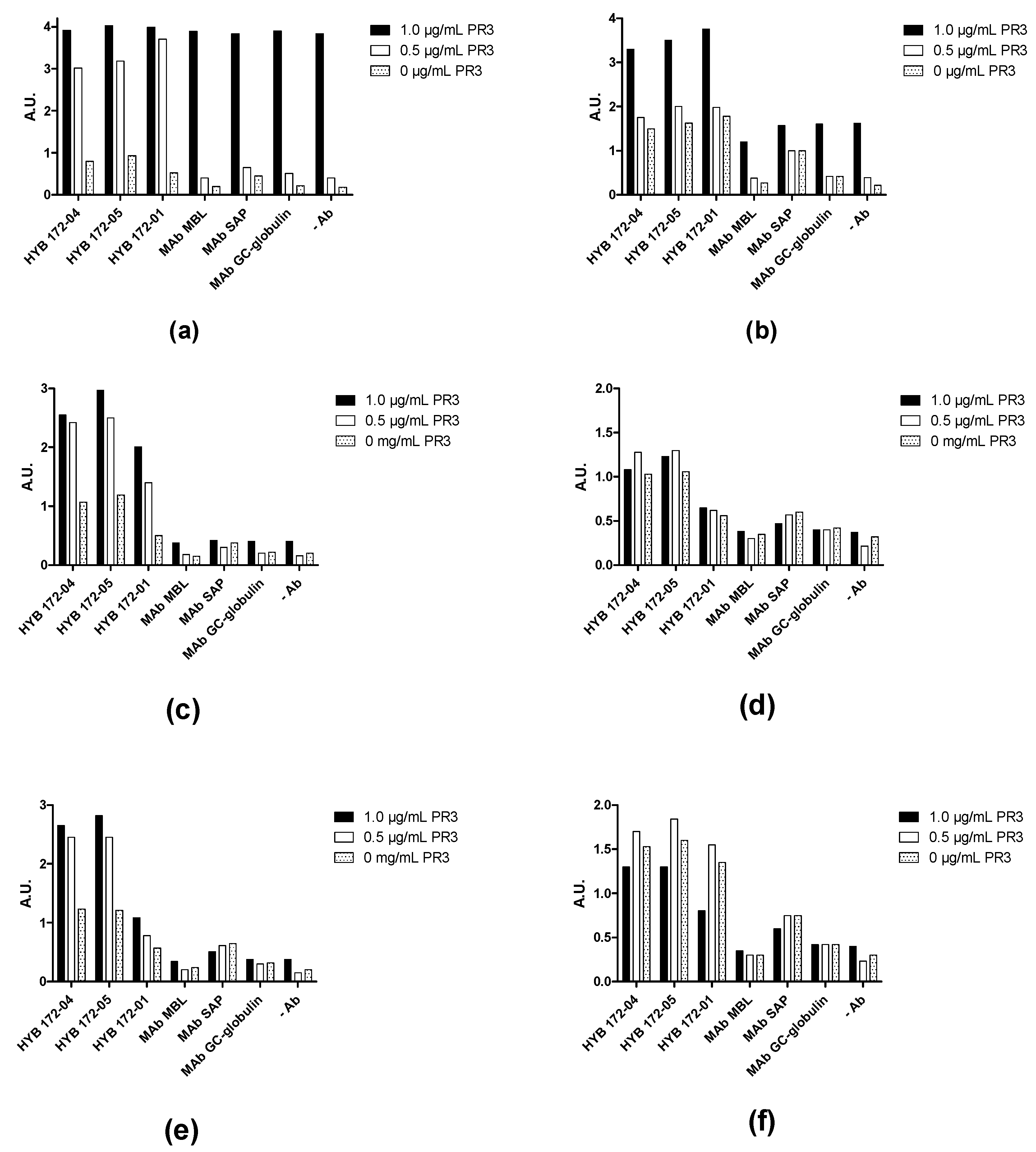{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Preparation | Mr/SDS-PAGE | Mw/MS | Defensins a | Activity b | Inhibition c | α1-AT Binding d |
|---|---|---|---|---|---|---|
| SSI | 26, 29, 32 | 25.5, 29.0, 32.3 | 5–6 | + | +/+/+ | + |
| Wieslab | 26, 29, 32 | 25.5, 28.9, 32.4 | 5–6 | + | +/+/+ | + |
| Arotec | 26, 29, 32 | 25.6, −, − | 1–2 | − | NA | + |
| (a) | ||||||
| PR3 MAb | Type | 50% Binding (µg/mL) | Elution pH (Low/High) | Inhibition of Binding * | α1-AT Inhibition | Immuno Blotting # |
| HYB 172-01 | IgG1,κ (6A6) | 0.6 | 4/12 | −/−/− | + | 26, 29 |
| HYB 172-04 | IgG2a,κ (4A3) | 0.6 | 2/12 | −/−/− | + | 26, 29, 32 |
| HYB 172-05 | IgG2a,κ (4A5) | 0.6 | 2/12 | −/−/− | + | 26, 29, 32 |
| (b) | ||||||
| PR3 Mab | ka (1/Ms) | kd (1/s) | Ka (1/M) | Kd (M) | Chi2 | |
| HYB 172-01 | 2.72 × 106 | 5.21 × 10−8 | 5.23 × 1013 | 1.91 × 10−14 | 4.03 | |
| HYB 172-04 | 1.03 × 107 | 5.71 × 10−4 | 1.8 × 1010 | 5.55 × 10−11 | 0.152 | |
| HYB 172-05 | 1.24 × 107 | 6.84 × 10−5 | 1.81 × 1011 | 1.81× 1011 | 4.8 | |
| (a) | |||||
| HYB | 172-05 | 206-1 | 172-03 | 172-01 | 172-04 |
| 172-05 | 100.00 | 92.66 | 63.55 | 60.38 | 60.38 |
| 206-1 | 92.66 | 100.00 | 65.42 | 61.32 | 61.32 |
| 172-03 | 63.55 | 65.42 | 100.00 | 55.77 | 55.77 |
| 172-01 | 60.38 | 61.32 | 55.77 | 100.00 | 100.00 |
| 172-04 | 60.38 | 61.32 | 55.77 | 100.00 | 100.00 |
| (b) | |||||
| HYB | 172-04 | 172-05 | 172-03 | 206-1 | 172-01 |
| 172-04 | 100.00 | 98.28 | 62.93 | 57.76 | 48.28 |
| 172-05 | 98.28 | 100.00 | 64.66 | 59.48 | 48.28 |
| 172-03 | 62.93 | 64.66 | 100.00 | 64.66 | 45.30 |
| 206-1 | 57.76 | 59.48 | 64.66 | 100.00 | 50.86 |
| 172-01 | 48.28 | 48.28 | 45.30 | 50.86 | 100.00 |
| Indirect Immunofluorescence Microscopy | Enzyme-Linked Immunosorbent Assay | |||||
|---|---|---|---|---|---|---|
| N = 2112 | N = 41 | N = 176 | ||||
| Antibody | C-ANCA Positive | P-ANCA Positive | PR3 Positive | PR3 Negative | MPO Positive | MPO Negative |
| Samples | 41 | 176 | 24 | 17 | 70 | 106 |
| % | 2 | 8 | 59 | 41 | 40 | 60 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zoega, M.; Trier, N.H.; Nejrup, R.G.; Chailyan, A.; Friis, T.; Højrup, P.; Houen, G. Defensin Interactions in Relation to Monoclonal and Disease-Related Proteinase 3 Antibodies Binding at the Catalytic Site. Antibodies 2023, 12, 23. https://doi.org/10.3390/antib12010023
Zoega M, Trier NH, Nejrup RG, Chailyan A, Friis T, Højrup P, Houen G. Defensin Interactions in Relation to Monoclonal and Disease-Related Proteinase 3 Antibodies Binding at the Catalytic Site. Antibodies. 2023; 12(1):23. https://doi.org/10.3390/antib12010023
Chicago/Turabian StyleZoega, Morten, Nicole Hartwig Trier, Rikke Guldhammer Nejrup, Anna Chailyan, Tina Friis, Peter Højrup, and Gunnar Houen. 2023. "Defensin Interactions in Relation to Monoclonal and Disease-Related Proteinase 3 Antibodies Binding at the Catalytic Site" Antibodies 12, no. 1: 23. https://doi.org/10.3390/antib12010023