
Serological Cross-Reactivity in Zoonotic Flaviviral Infections of Medical Importance

 , ,
, ,
Abstract
:1. Introduction
2. Epidemiology and Geographic Distribution of Medically Important Flaviviruses
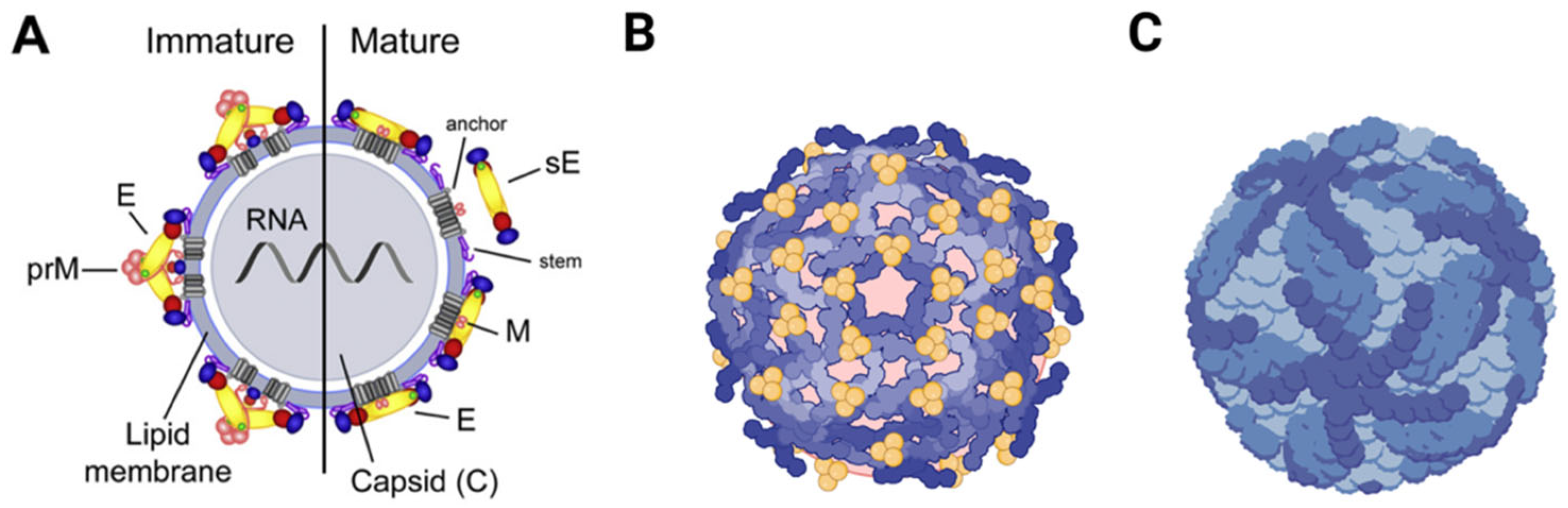
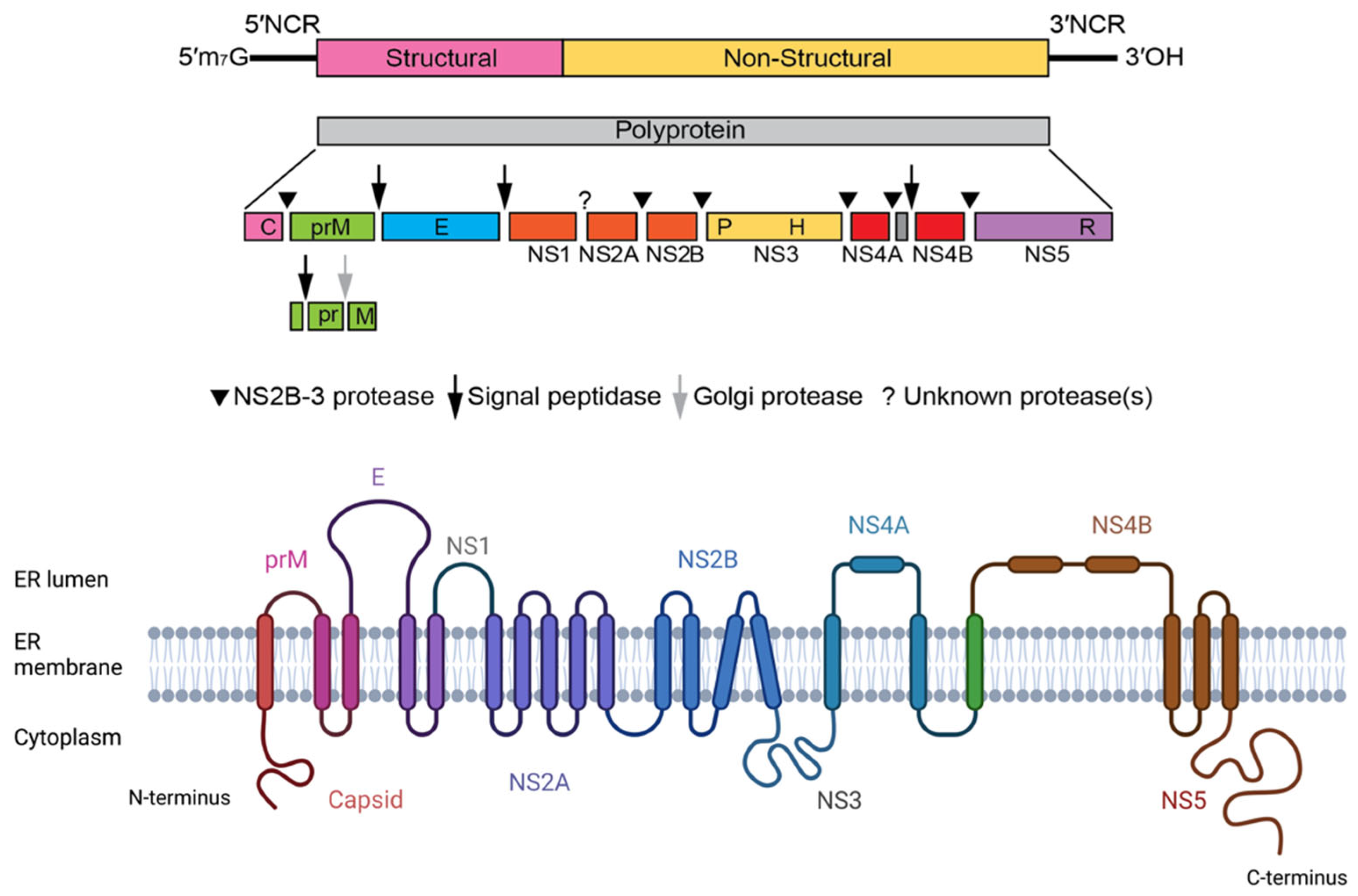
3. Structure of Flaviviruses
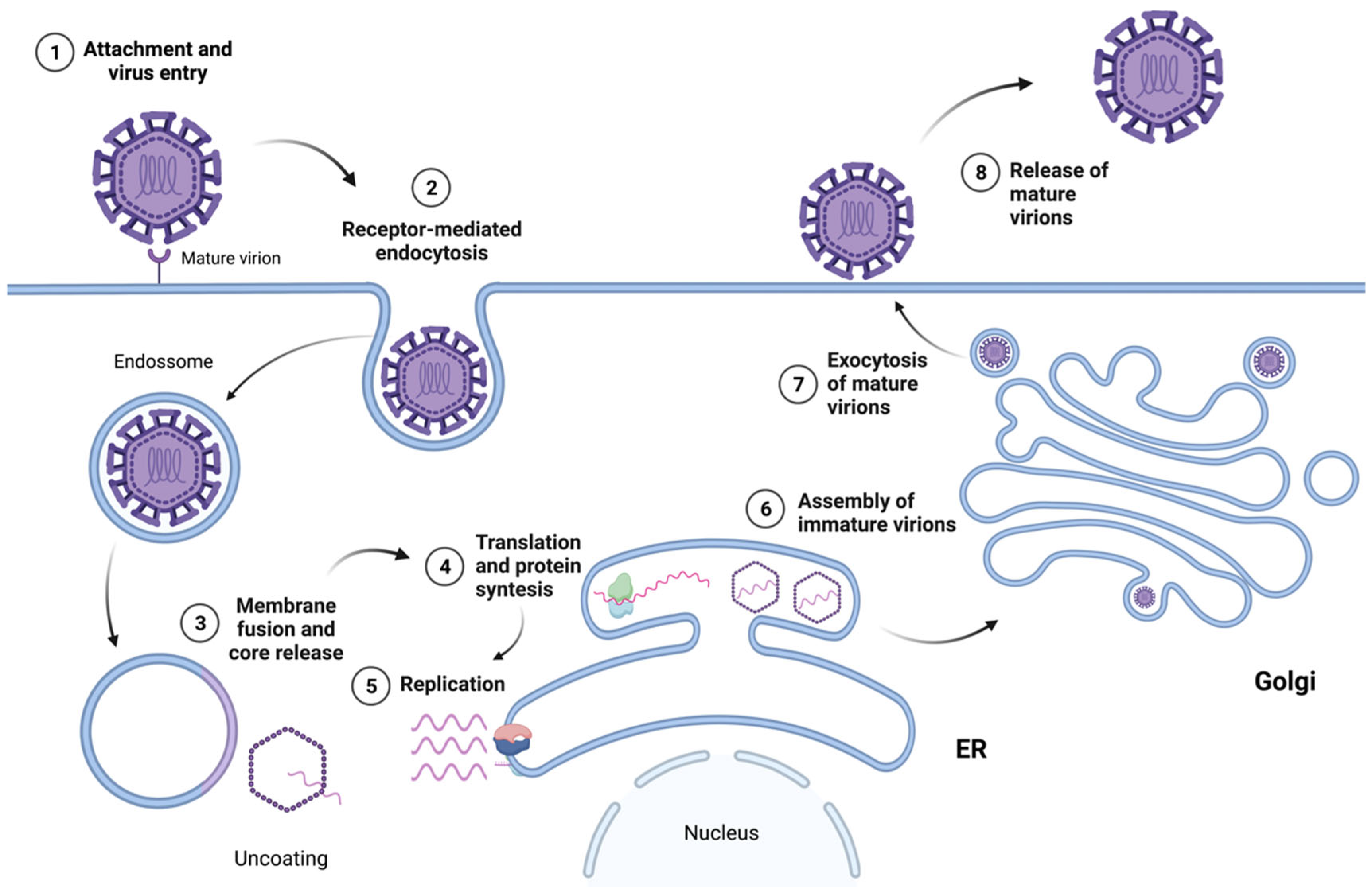
4. Flavivirus Replication Cycle
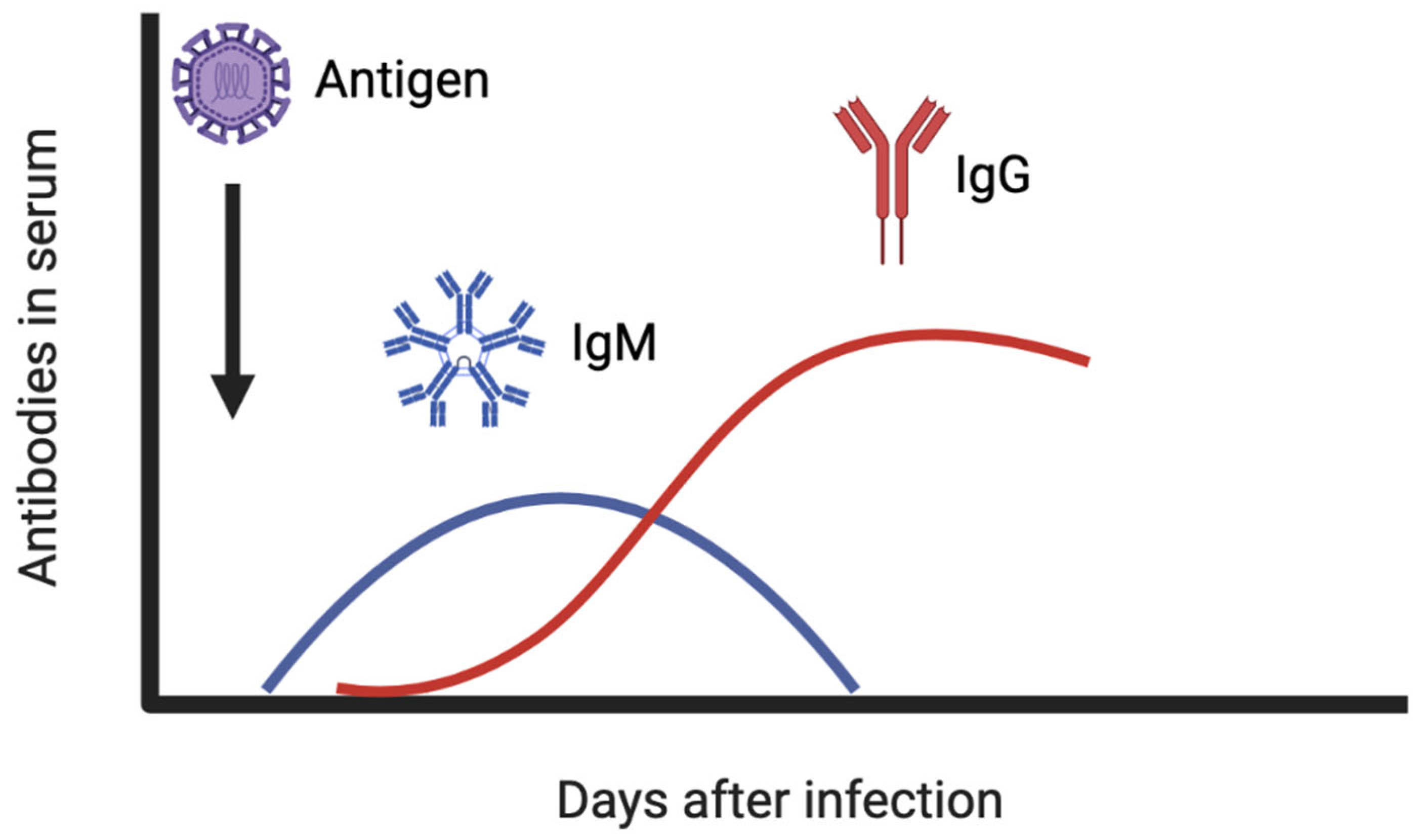
5. Immune Response against Viruses and the Importance of Neutralizing Antibodies
- Steric interference with virus–receptor binding;
- Blocking of endocytosis;
- Blocking of the uncoating process;
- Blocking of the uncoating process inside a cell after replication has started;
- Aggregation.
6. The “Original Antigenic Sin” Cross-Reactive Neutralizing Antibodies
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Payne, S. Viruses: From Understanding to Investigation; Elsevier Inc.: Amsterdam, The Netherlands, 2018; ISBN 978-0-12-803109-4. [Google Scholar]
- Saron, W.A.A.; Rathore, A.P.S.; Ting, L.; Ooi, E.E.; Low, J.; Abraham, S.N.; St. John, A.L. Flavivirus Serocomplex Cross-Reactive Immunity Is Protective by Activating Heterologous Memory CD4 T Cells. Sci. Adv. 2018, 4, eaar4297. [Google Scholar] [CrossRef] [Green Version]
- Rey, F.A.; Stiasny, K.; Vaney, M.; Dellarole, M.; Heinz, F.X. The Bright and the Dark Side of Human Antibody Responses to Flaviviruses: Lessons for Vaccine Design. EMBO Rep. 2017, 19, 206–224. [Google Scholar] [CrossRef]
- Solomon, T.; Mallewa, M. Dengue and Other Emerging Flaviviruses. J. Infect. 2001, 42, 104–115. [Google Scholar] [CrossRef] [PubMed]
- Cleton, N.B.; Godeke, G.J.; Reimerink, J.; Beersma, M.F.; van Doorn, H.R.; Franco, L.; Goeijenbier, M.; Jimenez-Clavero, M.A.; Johnson, B.W.; Niedrig, M.; et al. Spot the Difference—Development of a Syndrome Based Protein Microarray for Specific Serological Detection of Multiple Flavivirus Infections in Travelers. PLoS Negl. Trop. Dis. 2015, 9, e0003580. [Google Scholar] [CrossRef] [PubMed]
- Rathore, A.P.S.; St. John, A.L. Cross-Reactive Immunity among Flaviviruses. Front. Immunol. 2020, 11, 334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porterfield, J.S.; Brandt, W.E.; Shope, R.E.; Westaway, E.G.; Karabatsos, N.; Calisher, C.H.; Dalrymple, J.M. Antigenic Relationships between Flaviviruses as Determined by Cross-Neutralization Tests with Polyclonal Antisera. J. Gen. Virol. 1989, 70, 37–43. [Google Scholar] [CrossRef]
- Priyamvada, L.; Quicke, K.M.; Hudson, W.H.; Onlamoon, N.; Sewatanon, J.; Edupuganti, S.; Pattanapanyasat, K.; Chokephaibulkit, K.; Mulligan, M.J.; Wilson, P.C.; et al. Human Antibody Responses after Dengue Virus Infection Are Highly Cross-Reactive to Zika Virus. Proc. Natl. Acad. Sci. USA 2016, 113, 7852–7857. [Google Scholar] [CrossRef] [Green Version]
- Dejnirattisa, W.; Jumnainsong, A.; Onsirisakul, N.; Fitton, P.; Vasanawathana, S.; Limpitikul, W.; Puttikhunt, C.; Edwards, C.; Duangchinda, T.; Supasa, S.; et al. Cross-Reacting Antibodies Enhance Dengue Virus Infection in Humans. Science 2010, 328, 745–749. [Google Scholar] [CrossRef] [Green Version]
- Dejnirattisai, W.; Supasa, P.; Wongwiwat, W.; Rouvinski, A.; Barba-Spaeth, G.; Duangchinda, T.; Sakuntabhai, A.; Cao-Lormeau, V.M.; Malasit, P.; Rey, F.A.; et al. Dengue Virus Sero-Cross-Reactivity Drives Antibody-Dependent Enhancement of Infection with Zika Virus. Nat. Immunol. 2016, 17, 1102–1108. [Google Scholar] [CrossRef]
- Barba-Spaeth, G.; Dejnirattisai, W.; Rouvinski, A.; Vaney, M.C.; Medits, I.; Sharma, A.; Simon-Lorière, E.; Sakuntabhai, A.; Cao-Lormeau, V.M.; Haouz, A.; et al. Structural Basis of Potent Zika-Dengue Virus Antibody Cross-Neutralization. Nature 2016, 536, 48–53. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.-S.; Ng, L.-C.; Thein, T.-L.; Leo, Y.-S.; Yung, C.-F.; Wong, J.G.X.; Lye, D.C.; Tan, L.-K.; Gan, V.C. Dengue Serotype-Specific Differences in Clinical Manifestation, Laboratory Parameters and Risk of Severe Disease in Adults, Singapore. Am. J. Trop. Med. Hyg. 2015, 92, 999–1005. [Google Scholar] [CrossRef]
- Chan, K.W.K.; Tan, N.W.W.; Vasudevan, S.G.; Watanabe, S. Dengue Virus and Zika Virus Serological Cross-Reactivity and Their Impact on Pathogenesis in Mice. J. Infect. Dis. 2018, 219, 223–233. [Google Scholar] [CrossRef]
- Guzman, M.G.; Vazquez, S. The Complexity of Antibody-Dependent Enhancement of Dengue Virus Infection. Viruses 2010, 2, 2649–2662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martín-Acebes, M.A.; Saiz, J.C.; de Oya, N.J. Antibody-Dependent Enhancement and Zika: Real Threat or Phantom Menace? Front. Cell. Infect. Microbiol. 2018, 8, 2014–2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halstead, S.B.; Cohen, S.N. Dengue Hemorrhagic Fever at 60 Years: Early Evolution of Concepts of Causation and Treatment. Microbiol. Mol. Biol. Rev. 2015, 79, 281–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katzelnick, L.C.; Bos, S.; Harris, E. Protective and Enhancing Interactions among Dengue Viruses 1-4 and Zika Virus. Curr. Opin. Virol. 2020, 43, 59–70. [Google Scholar] [CrossRef]
- Khandia, R.; Munjal, A.; Dhama, K.; Karthik, K.; Tiwari, R.; Malik, Y.S.; Singh, R.K.; Chaicumpa, W. Modulation of Dengue/Zika Virus Pathogenicity by Antibody-Dependent Enhancement and Strategies to Protect Against Enhancement in Zika Virus Infection. Front. Immunol. 2018, 9, 597. [Google Scholar] [CrossRef]
- Makino, Y.; Tadano, M.; Saito, M.; Fukunaga, T.; Maneekarn, N.; Sittisombut, N.; Sirisanthana, V.; Ponepraser, B. Studies on Serological Cross-Reaction in Sequential Flavivirus Infections. Microbiol. Immunol. 1994, 38, 951–955. [Google Scholar] [CrossRef] [Green Version]
- Heinz, F.X.; Stiasny, K. Flaviviruses and Their Antigenic Structure. J. Clin. Virol. 2012, 55, 289–295. [Google Scholar] [CrossRef]
- DeSA, U. World Population Prospects: The 2012 Revision; United Nations Department of Economic and Social Affairs: New York, NY, USA, 2013. [Google Scholar]
- Braack, L.; Gouveia De Almeida, A.P.; Cornel, A.J.; Swanepoel, R.; De Jager, C. Mosquito-Borne Arboviruses of African Origin: Review of Key Viruses and Vectors. Parasites Vectors 2018, 11, 29. [Google Scholar] [CrossRef]
- Chong, H.Y.; Leow, C.Y.; Abdul Majeed, A.B.; Leow, C.H. Flavivirus Infection—A Review of Immunopathogenesis, Immunological Response, and Immunodiagnosis. Virus Res. 2019, 274, 197770. [Google Scholar] [CrossRef] [PubMed]
- Daep, C.A.; Muñoz-Jordán, J.L.; Eugenin, E.A. Flaviviruses, an Expanding Threat in Public Health: Focus on Dengue, West Nile, and Japanese Encephalitis Virus. J. Neurovirol. 2014, 20, 539–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cleton, N.; Koopmans, M.; Reimerink, J.; Godeke, G.J.; Reusken, C. Come Fly with Me: Review of Clinically Important Arboviruses for Global Travelers. J. Clin. Virol. 2012, 55, 191–203. [Google Scholar] [CrossRef] [PubMed]
- Barrett, A.D.T.; Higgs, S. Yellow Fever: A Disease That Has Yet To Be Conquered. Annu. Rev. Entomol. 2007, 52, 209–229. [Google Scholar] [CrossRef] [Green Version]
- Mirzaian, E.; Durham, M.J.; Hess, K.; Goad, J.A. Mosquito-Borne Illnesses in Travelers: A Review of Risk and Prevention. Pharmacotherapy 2010, 30, 1031–1043. [Google Scholar] [CrossRef]
- Romero, J.R.; Simonsen, K.A. Powassan Encephalitis and Colorado Tick Fever. Infect. Dis. Clin. N. Am. 2008, 22, 545–559. [Google Scholar] [CrossRef]
- Günther, G.; Haglund, M. Tick-Borne Encephalopathies: Epidemiology, Diagnosis, Treatment and Prevention. CNS Drugs 2005, 19, 1009–1032. [Google Scholar] [CrossRef]
- Maltezou, H.C.; Andonova, L.; Andraghetti, R.; Bouloy, M.; Ergonul, O.; Jongejan, F.; Kalvatchev, N.; Nichol, S.; Niedrig, M.; Platonov, A.; et al. Crimean-Congo Hemorrhagic Fever in Europe: Current Situation Calls for Preparedness. Euro Surveill. Bull. Eur. sur les Mal. Transm. Eur. Commun. Dis. Bull. 2010, 15, 19504. [Google Scholar] [CrossRef]
- MacKenzie, J.S.; Williams, D.T. The Zoonotic Flaviviruses of Southern, South-Eastern and Aastern Asia, and Australasia: The Potential for Emergent Viruses. Zoonoses Public Health 2009, 56, 338–356. [Google Scholar] [CrossRef]
- Pugliese, A.; Beltramo, T.; Torre, D. Emerging and Re-Emerging Viral Infections in Europe. Cell Biochem. Funct. 2007, 25, 1–13. [Google Scholar] [CrossRef]
- Blitvich, B.J.; Firth, A.E. Insect-specific flaviviruses: A systematic review of their discovery, host range, mode of transmission, superinfection exclusion potential and genomic organization. Viruses 2015, 7, 1927–1959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endale, A.; Medhin, G.; Darfiro, K.; Kebede, N.; Legesse, M. Magnitude of Antibody Cross-Reactivity in Medically Important Mosquito-Borne Flaviviruses: A Systematic Review. Infect. Drug Resist. 2021, 14, 4291–4299. [Google Scholar] [CrossRef] [PubMed]
- Gould, E.A.; Solomon, T. Pathogenic Flaviviruses. Lancet 2008, 371, 500–509. [Google Scholar] [CrossRef] [PubMed]
- Pierson, T.C.; Diamond, M.S. Degrees of Maturity: The Complex Structure and Biology of Flaviviruses. Curr. Opin. Virol. 2012, 2, 168–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Corver, J.; Chipman, P.R.; Zhang, W.; Pletnev, S.V.; Sedlak, D.; Baker, T.S.; Strauss, J.H.; Kuhn, R.J.; Rossmann, M.G. Structures of Immature Flavivirus Particles. EMBO J. 2003, 22, 2604–2613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Jia, R.; Shen, H.; Wang, M.; Yin, Z.; Cheng, A. Structures and Functions of the Envelope Glycoprotein in Flavivirus Infections. Viruses 2017, 9, 338. [Google Scholar] [CrossRef] [Green Version]
- Chávez, J.H.; Silva, J.R.; Amarilla, A.A.; Moraes Figueiredo, L.T. Domain III Peptides from Flavivirus Envelope Protein Are Useful Antigens for Serologic Diagnosis and Targets for Immunization. Biologicals 2010, 38, 613–618. [Google Scholar] [CrossRef]
- Ng, W.C.; Soto-Acosta, R.; Bradrick, S.S.; Garcia-Blanco, M.A.; Ooi, E.E. The 5’ and 3’ Untranslated Regions of the Flaviviral Genome. Viruses 2017, 9, 137. [Google Scholar] [CrossRef] [Green Version]
- Lindenbach, B.D.; Rice, C.M. The Viruses and Their Replication. In Fields Virology, 5th ed.; Lippincot William & Wilkins: Philadelphia, PA, USA, 2007; pp. 1101–1113. [Google Scholar]
- Mukhopadhyay, S.; Kuhn, R.J.; Rossmann, M.G. A Structural Perspective of the Flavivirus Life Cycle. Nat. Rev. Microbiol. 2005, 3, 13–22. [Google Scholar] [CrossRef]
- Morita, E.; Suzuki, Y. Membrane-Associated Flavivirus Replication Complex—Its Organization and Regulation. Viruses 2021, 13, 1060. [Google Scholar] [CrossRef]
- Pierson, T.C.; Diamond, M.S. The Continued Threat of Emerging Flaviviruses. Nat. Microbiol. 2020, 5, 796–812. [Google Scholar] [CrossRef] [PubMed]
- Van Den Elsen, K.; Quek, J.P.; Luo, D. Molecular Insights into the Flavivirus Replication Complex. Viruses 2021, 13, 956. [Google Scholar] [CrossRef] [PubMed]
- Bollati, M.; Alvarez, K.; Assenberg, R.; Baronti, C.; Canard, B.; Cook, S.; Coutard, B.; Decroly, E.; de Lamballerie, X.; Gould, E.A.; et al. Structure and Functionality in Flavivirus NS-Proteins: Perspectives for Drug Design. Antiviral Res. 2010, 87, 125–148. [Google Scholar] [CrossRef] [PubMed]
- Murray, C.L.; Jones, C.T.; Rice, C.M. Architects of Assembly: Roles of Flaviviridae Non-Structural Proteins in Virion Morphogenesis. Nat. Rev. Microbiol. 2008, 6, 699–708. [Google Scholar] [CrossRef] [Green Version]
- Egloff, M.-P.; Decroly, E.; Malet, H.; Selisko, B.; Benarroch, D.; Ferron, F.; Canard, B. Structural and Functional Analysis of Methylation and 5′-RNA Sequence Requirements of Short Capped RNAs by the Methyltransferase Domain of Dengue Virus NS5. J. Mol. Biol. 2007, 372, 723–736. [Google Scholar] [CrossRef]
- Heinz, F.X.; Stiasny, K. Flaviviruses and Flavivirus Vaccines. Vaccine 2012, 30, 4301–4306. [Google Scholar] [CrossRef]
- Cann, A.J. Principles of Molecular Virology, 5th ed.; Elsevier: Amsterdam, The Netherlands, 2012; ISBN 9780123849397. [Google Scholar]
- Jensen, S.; Thomsen, A.R. Sensing of RNA Viruses: A Review of Innate Immune Receptors Involved in Recognizing RNA Virus Invasion. J. Virol. 2012, 86, 2900–2910. [Google Scholar] [CrossRef] [Green Version]
- Braciale, T.J.; Hahn, Y.S. Immunity to Viruses. Immunol. Rev. 2013, 255, 5–12. [Google Scholar] [CrossRef] [Green Version]
- Swain, S.S.; Kai McKinstry, K.; Strutt, T.M. Expanding Roles for CD4+ T Cells in Immunity to Viruses. Nat. Immunol. 2011, 12, 136–148. [Google Scholar] [CrossRef]
- Vatti, A.; Monsalve, D.M.; Pacheco, Y.; Chang, C.; Anaya, J.M.; Gershwin, M.E. Original Antigenic Sin: A Comprehensive Review. J. Autoimmun. 2017, 83, 12–21. [Google Scholar] [CrossRef]
- Huber, S.R.; van Beek, J.; de Jonge, J.; Luytjes, W.; van Baarle, D. T Cell Responses to Viral Infections—Opportunities for Peptide Vaccination. Front. Immunol. 2014, 5, 171. [Google Scholar] [CrossRef]
- Flint, J.; Racaniello, V.R.; Rall, G.F.; Marie Skalka, A.; Enquist, L.W. Principles of Virology, 4th ed.; American Society for Microbiology: Washington, DC, USA, 2015; ISBN 9781555819330. [Google Scholar]
- Calvert, A.E.; Horiuchi, K.; Boroughs, K.L.; Ong, Y.T.; Anderson, K.M.; Biggerstaff, B.J.; Stone, M.; Simmons, G.; Busch, M.P.; Huang, C.Y.-H. The Specificity of the Persistent IgM Neutralizing Antibody Response in Zika Virus Infections among Individuals with Prior Dengue Virus Exposure. J. Clin. Microbiol. 2021, 59, e0040021. [Google Scholar] [CrossRef] [PubMed]
- Edara, V.-V.; Pinsky, B.A.; Suthar, M.S.; Lai, L.; Davis-Gardner, M.E.; Floyd, K.; Flowers, M.W.; Wrammert, J.; Hussaini, L.; Ciric, C.R.; et al. Infection and Vaccine-Induced Neutralizing-Antibody Responses to the SARS-CoV-2 B.1.617 Variants. N. Engl. J. Med. 2021, 385, 664–666. [Google Scholar] [CrossRef] [PubMed]
- Morales-Núñez, J.J.; Muñoz-Valle, J.F.; Torres-Hernández, P.C.; Hernández-Bello, J. Overview of Neutralizing Antibodies and Their Potential in COVID-19. Vaccines 2021, 9, 1376. [Google Scholar] [CrossRef] [PubMed]
- Heinz, F.X.; Stiasny, K. The Antigenic Structure of Zika Virus and Its Relation to Other Flaviviruses: Implications for Infection and Immunoprophylaxis. Microbiol. Mol. Biol. Rev. 2017, 81, e00055-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henry, C.; Palm, A.K.E.; Krammer, F.; Wilson, P.C. From Original Antigenic Sin to the Universal Influenza Virus Vaccine. Trends Immunol. 2018, 39, 70–79. [Google Scholar] [CrossRef]
- Francis, T. On the Doctrine of Original Antigenic Sin. Proc. Am. Philos. Soc. 1960, 104, 572–578. [Google Scholar]
- Howard, F.H.N.; Kwan, A.; Winder, N.; Mughal, A.; Collado-Rojas, C.; Muthana, M. Understanding Immune Responses to Viruses—Do Underlying Th1/Th2 Cell Biases Predict Outcome? Viruses 2022, 14, 1493. [Google Scholar] [CrossRef]
- MacLachlan, N.J.; Dubovi, E.J.B.T.-F.V.V. Chapter 29—Flaviviridae, 5th ed.; Academic Press: Boston, MA, USA, 2017; pp. 525–545. ISBN 978-0-12-800946-8. [Google Scholar]
- Chan, K.R.; Ismail, A.A.; Thergarajan, G.; Raju, C.S.; Yam, H.C.; Rishya, M.; Sekaran, S.D. Serological Cross-Reactivity among Common Flaviviruses. Front. Cell. Infect. Microbiol. 2022, 12, 1–14. [Google Scholar] [CrossRef]
- Houghton-Triviño, N.; Montaña, D.; Castellanos, J. Dengue-Yellow Fever Sera Cross-Reactivity; Challenges for Diagnosis. Rev. Salud Pública 2009, 10, 299–307. [Google Scholar] [CrossRef] [Green Version]
- Mansfield, K.L.; Horton, D.L.; Johnson, N.; Li, L.; Barrett, A.D.T.; Smith, D.J.; Galbraith, S.E.; Solomon, T.; Fooks, A.R. Flavivirus-Induced Antibody Cross-Reactivity. J. Gen. Virol. 2011, 92, 2821–2829. [Google Scholar] [CrossRef] [PubMed]
- Maeki, T.; Tajima, S.; Ikeda, M.; Kato, F.; Taniguchi, S.; Nakayama, E.; Takasaki, T.; Lim, C.K.; Saijo, M. Analysis of Cross-Reactivity between Flaviviruses with Sera of Patients with Japanese Encephalitis Showed the Importance of Neutralization Tests for the Diagnosis of Japanese Encephalitis. J. Infect. Chemother. 2019, 25, 786–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, B.; Chen, H.; Gao, N.; An, J. Cross-Reactive Immunity among Five Medically Important Mosquito-Borne Flaviviruses Related to Human Diseases. Viruses 2022, 14, 1213. [Google Scholar] [CrossRef] [PubMed]
- Vojtek, I.; Buchy, P.; Doherty, T.M.; Hoet, B. Would Immunization Be the Same without Cross-Reactivity? Vaccine 2019, 37, 539–549. [Google Scholar] [CrossRef]
- Sirohi, D.; Chen, Z.; Sun, L.; Klose, T.; Pierson, T.C.; Rossmann, M.G.; Kuhn, R.J. The 3.8Å Resolution Cryo-EM Structure of Zika Virus. Science 2016, 352, 467–470. [Google Scholar] [CrossRef] [Green Version]
- Gunther, V.J.; Putnak, R.; Eckels, K.H.; Mammen, M.P.; Scherer, J.M.; Lyons, A.; Sztein, M.B.; Sun, W. A Human Challenge Model for Dengue Infection Reveals a Possible Protective Role for Sustained Interferon Gamma Levels during the Acute Phase of Illness. Vaccine 2011, 29, 3895–3904. [Google Scholar] [CrossRef]
- Izurieta, R.; Macaluso, M.; Watts, D.; Tesh, R.; Guerra, B.; Cruz, L.; Galwankar, S.; Vermund, S. Anamnestic Immune Response to Dengue and Decreased Severity of Yellow Fever. J. Glob. Infect. Dis. 2009, 1, 111. [Google Scholar] [CrossRef]
- Zhu, H.-B.; Tesh, R.B.; Xiao, S.-Y.; Guzman, H.; da Rosa, A.P.A.T. Alteration of Clinical Outcome and Histopathology of Yellow Fever Virus Infection in a Hamster Model By Previous Infection with Heterologous Flaviviruses. Am. J. Trop. Med. Hyg. 2018, 68, 695–703. [Google Scholar] [CrossRef]
- Thomas, S.J.; Yoon, I.K. A Review of Dengvaxia®: Development to Deployment. Hum. Vaccines Immunother. 2019, 15, 2295–2314. [Google Scholar] [CrossRef] [Green Version]
- McArthur, M.A.; Sztein, M.B.; Edelman, R. Dengue Vaccines: Recent Developments, Ongoing Challenges and Current Candidates. Expert Rev. Vaccines 2013, 12, 933–953. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
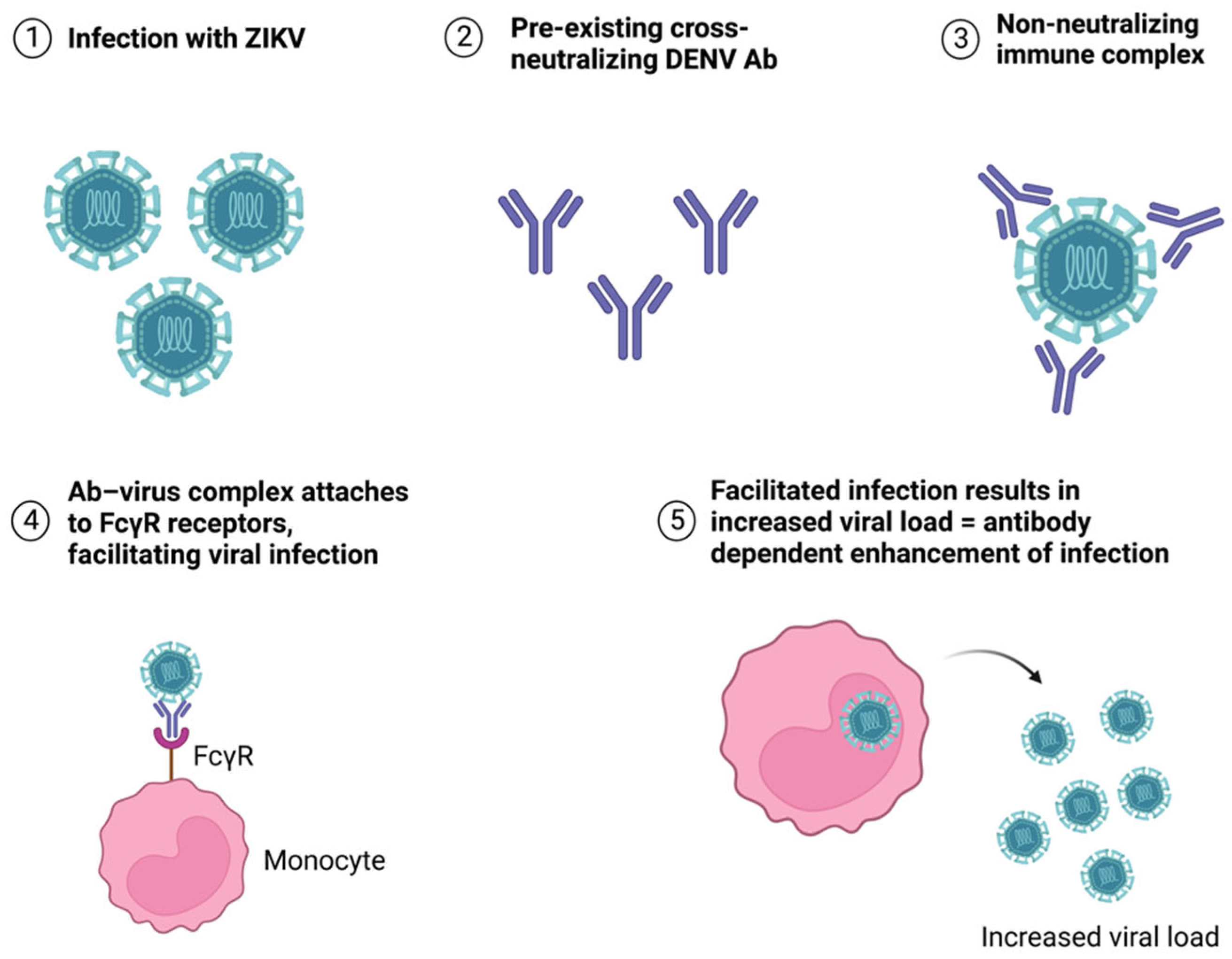{kind=link}
| Antigenic Complex (Serocomplex) | Viruses |
|---|---|
| Tick-borne encephalitis | Russian spring-summer encephalitis |
| Central European encephalitis | |
| Omsk hemorrhagic fever | |
| Louping Ill | |
| Kyasanur Forest disease | |
| Langat | |
| Phnom Penh bat | |
| Carey Island | |
| Negishi | |
| Powassan | |
| Karshi | |
| Royal Farm | |
| Rio Bravo | Rio Bravo |
| Entebbe bat | |
| Dakar bat | |
| Bukalasa bat | |
| Saboya | |
| Apoi | |
| Japanese encephalitis | Japanese encephalitis |
| Murray Valley encephalitis | |
| Kokobera | |
| Alfuy | |
| Stratford | |
| St. Louis encephalitis | |
| Usutu | |
| West Nile | |
| Kunjin | |
| Koutango | |
| Tyuleniy | Tyuleniy, Saumarez |
| Reef | |
| Meaban | |
| Ntaya | Ntaya |
| Temusu | |
| Yokose | |
| Israel turkey meningoencephalitis | |
| Bagaza | |
| Uganda S | Uganda S |
| Banzi | |
| Bouboui | |
| Edge Hill | |
| Dengue | Dengue 1 |
| Dengue 2 | |
| Dengue 3 | |
| Dengue 4 | |
| Modoc virus complex | Modoc |
| Cowbone Ridge | |
| Jutiapa Sal | |
| Vieja | |
| San Perlita |
| Virus | Country Where It Was Discovered | Geographic Distribution | Main Vector spp. | Main Host spp. | Human Disease |
|---|---|---|---|---|---|
| Alkhurma | Saudi Arabia | Arabian Peninsula a | Ornithodorus savignyia | Humans, sheep, camels | Haemorrhagic fever |
| Apoi | Japan | Japan | Unknown | Rodents a | Encephalitis |
| Bagaza | Central African Republic | Africa | Culex spp. | Unknown | Fever |
| Banzi | South Africa | Africa | Culex spp. | Unknown | Fever |
| Bussuquara | Brazil | Brazil | Culex spp. | Unknown | Fever |
| Dakar bat | Senegal | Africa | Unknown | Bats a | Fever |
| Dengue 1 | Hawaii | Tropics, subtropics | Aedes aegypti | Humans | Fever, rash, vasculopathy |
| Dengue 2 | New Guinea | Tropics, subtropics | Aedes aegypti | Humans | Fever, rash, vasculopathy |
| Dengue 3 | Philippines | Tropics, subtropics | Aedes aegypti | Humans | Fever, rash, vasculopathy |
| Dengue 4 | Philippines | Tropics, subtropics | Aedes aegypti | Humans | Fever, rash, vasculopathy |
| Ilheus | Brazil | South and Central America | Culex spp. a | Birds | Fever |
| Japanese encephalitis | Japan | Asia and Oceania | Culex trita-eniorhynchus | Birds | Encephalitis |
| Koutango | Senegal | Senegal | Unknown | Rodents a | Fever, rash |
| Kyasanur Forest disease | India | India | Haemaphysalis spinigera | Monkeys | Haemorrhagic fever |
| Langat | Malaysia | Malaysia, Thailand, Siberia | Ixodes granulatus | Unknown | Encephalitis |
| Louping ill | Scotland | UK, Ireland; it has also been reported in Norway and one region in far eastern Russia, and on the island of Bornholm in Denmark | Ixodes spp. | Sheep, grouse, hares | Encephalitis |
| Modoc | USA | USA | Unknown | Peromyscus maniculatus | Encephalitis |
| Murrat Valley encephalitis | Australia | Australia, New Guinea | Culex annulirostris | Birds | Encephalitis |
| Ntaya | Uganda | Africa | Mosquitos | Unknown | Fever |
| Omsk haemorrhagic fever | Russia | Western Siberia | Dermacentor pictus | Muskrats, rodents a | Haemorrhagic fever |
| Powassan | Russia, USA, Canada | Russia, USA, Canada | Ixodes spp. | Small mammals | Encephalitis |
| Rio Bravo | USA | USA, Mexico | Unknown | Tadanida braziliensis mexicana | Fever |
| Rocio | Brazil | Brazil | Culex spp. a | Birds | Encephalitis |
| St Louis encephalitis | USA | North, Central, and South America | Culex spp. | Birds | Encephalitis |
| Sepik | New Guinea | New Guinea | Mosquitos | Unknown | Fever |
| Spondweni | South Africa | Africa | Aedes circumluteolus | Unknown | Fever |
| Tick-borne encephalitis | Russia | Many parts of Europe and Asia | Ixodes spp. | Rodents a | Encephalitis |
| Usutu | South Africa | Africa | Mosquitoes | Birds | Fever, rash |
| Wesselsbron | South Africa | Parts of Africa, Madagascar, and Thailand | Aedes spp. | Unknown | Unknown |
| West Nile | Uganda | Worldwide | Mosquitos, ticks | Birds | Encephalitis |
| Yellow Fever | Ghana | Tropical and subtropical areas of Africa and South America | Aedes spp/Haemagogus spp. | Monkeys | Pantropic |
| Zika | Uganda | Africa, the Americas, Southern Asia and Western Pacific | Aedes spp. | Monkeys a | Fever, rash |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gomes da Silva, P.; Seixas dos Reis, J.A.; Nogueira Rodrigues, M.; da Silva Ardaya, Q.; Mesquita, J.R. Serological Cross-Reactivity in Zoonotic Flaviviral Infections of Medical Importance. Antibodies 2023, 12, 18. https://doi.org/10.3390/antib12010018
Gomes da Silva P, Seixas dos Reis JA, Nogueira Rodrigues M, da Silva Ardaya Q, Mesquita JR. Serological Cross-Reactivity in Zoonotic Flaviviral Infections of Medical Importance. Antibodies. 2023; 12(1):18. https://doi.org/10.3390/antib12010018
Chicago/Turabian StyleGomes da Silva, Priscilla, José Augusto Seixas dos Reis, Marcio Nogueira Rodrigues, Quézia da Silva Ardaya, and João Rodrigo Mesquita. 2023. "Serological Cross-Reactivity in Zoonotic Flaviviral Infections of Medical Importance" Antibodies 12, no. 1: 18. https://doi.org/10.3390/antib12010018