
Bioprocess Development and Characterization of a 13C-Labeled Hybrid Bispecific Antibody Produced in Escherichia coli
 ,
,
Abstract
:1. Introduction
2. Methods
2.1. Bacterial Strain and Plasmids
2.2. Fermentations
2.3. Analytical Measurements for Monitoring Fermentation
2.4. Downstream Processing
2.5. Mass Spectrometry Analysis and Determination of % 13C-Incorporation
3. Results
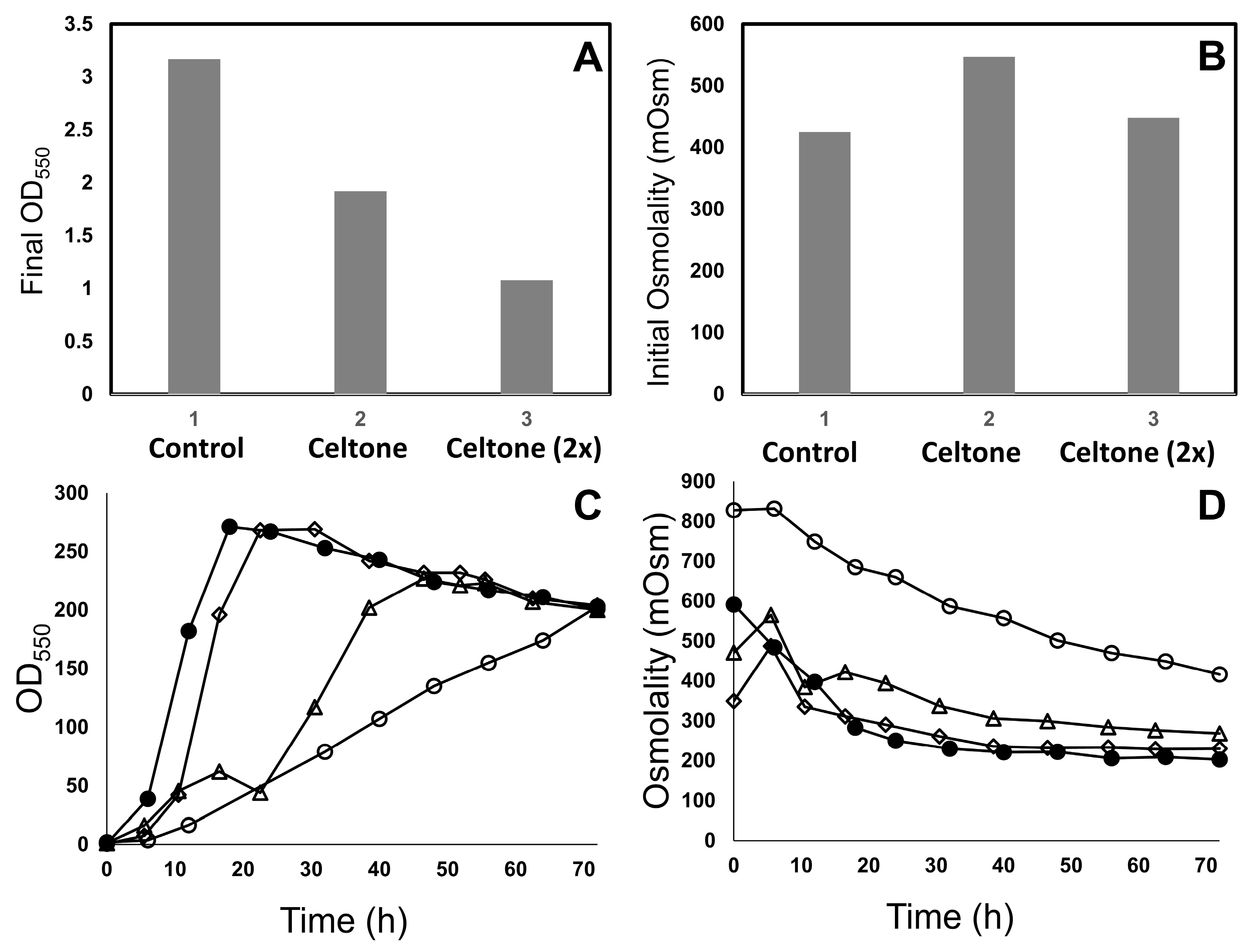
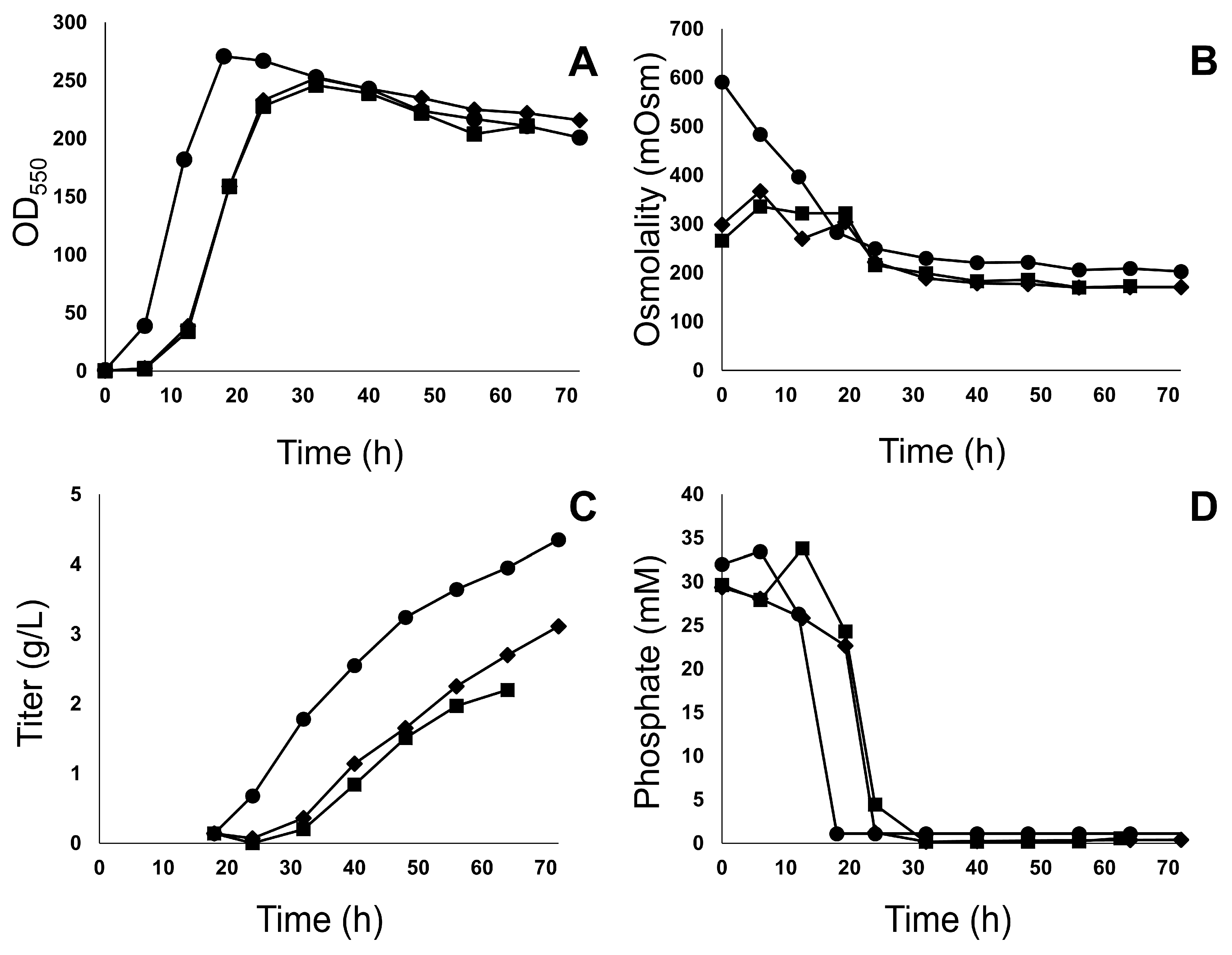
3.1. Fermentation
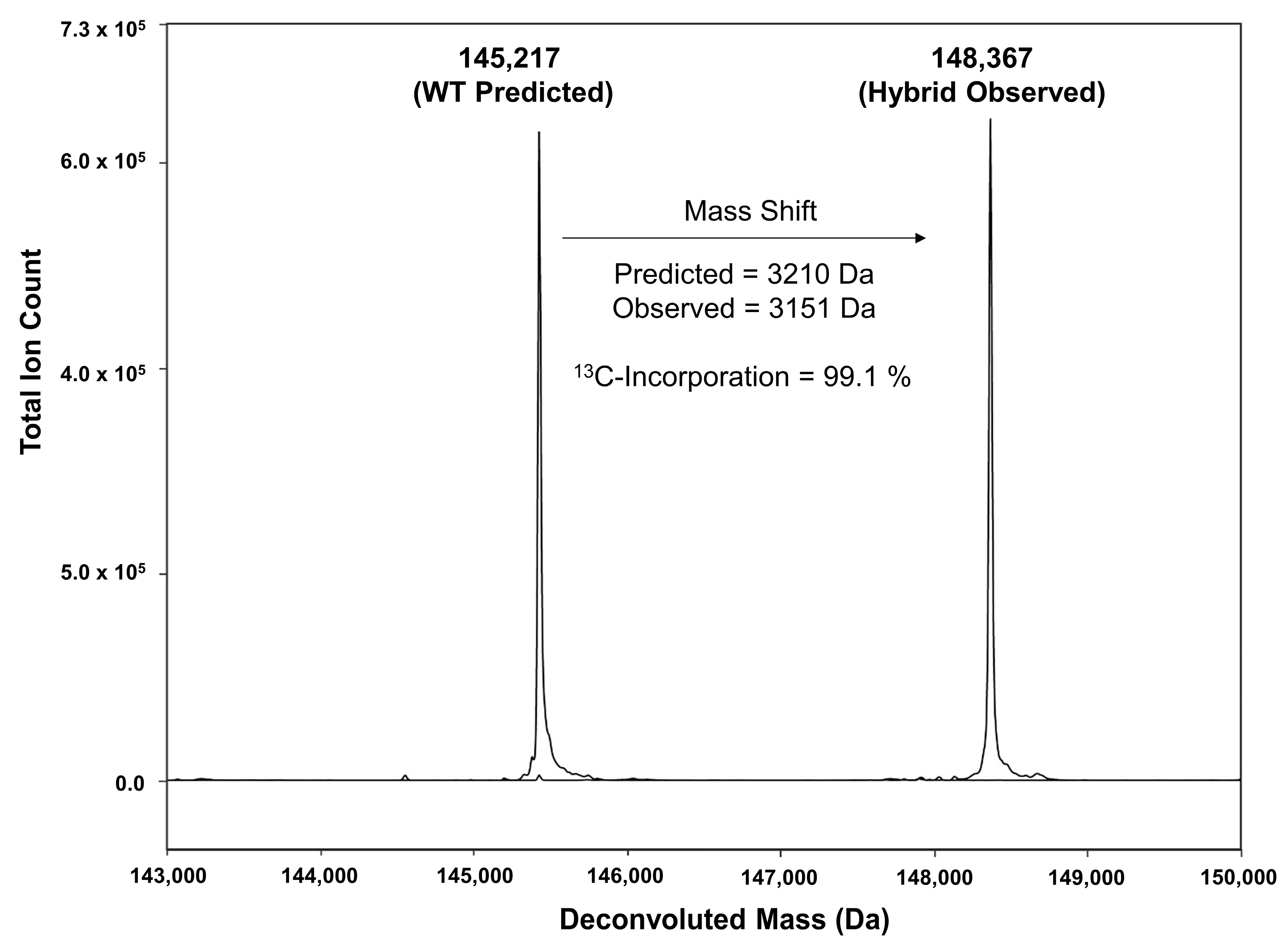
3.2. Mass Spectrometry Analysis and % 13C-Incorporation
4. Discussion and Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Lu, R.M.; Hwang, Y.C.; Liu, I.J.; Lee, C.C.; Tsai, H.Z.; Li, H.J.; Wu, H.C. Development of therapeutic antibodies for the treatment of diseases. J. Biomed. Sci. 2020, 27, 1. [Google Scholar] [CrossRef]
- Kaplon, H.; Chenoweth, A.; Crescioli, S.; Reichert, J.M. Antibodies to watch in 2022. MAbs 2022, 14, 2014296. [Google Scholar] [CrossRef]
- Carter, P.J.; Rajpal, A. Designing antibodies as therapeutics. Cell 2022, 185, 2789–2805. [Google Scholar] [CrossRef]
- Chiu, M.L.; Goulet, D.R.; Teplyakov, A.; Gilliland, G.L. Antibody Structure and Function: The Basis for Engineering Therapeutics. Antibodies 2019, 8, 55. [Google Scholar] [CrossRef]
- Sevy, A.M.; Meiler, J. Antibodies: Computer-Aided Prediction of Structure and Design of Function. Microbiol. Spectr. 2014, 2, 173–190. [Google Scholar] [CrossRef]
- Garidel, P.; Hegyi, M.; Bassarab, S.; Weichel, M. A rapid, sensitive and economical assessment of monoclonal antibody conformational stability by intrinsic tryptophan fluorescence spectroscopy. Biotechnol. J. 2008, 3, 1201–1211. [Google Scholar] [CrossRef]
- Thiagarajan, G.; Semple, A.; James, J.K.; Cheung, J.K.; Shameem, M. A comparison of biophysical characterization techniques in predicting monoclonal antibody stability. MAbs 2016, 8, 1088–1097. [Google Scholar] [CrossRef]
- Arbogast, L.W.; Brinson, R.G.; Marino, J.P. Mapping monoclonal antibody structure by 2D 13C NMR at natural abundance. Anal. Chem. 2015, 87, 3556–3561. [Google Scholar] [CrossRef]
- Harris, L.J.; Skaletsky, E.; McPherson, A. Crystallographic structure of an intact IgG1 monoclonal antibody. J. Mol. Biol. 1998, 275, 861–872. [Google Scholar] [CrossRef]
- Sears, A.E.; Albiez, S.; Gulati, S.; Wang, B.; Kiser, P.; Kovacik, L.; Engel, A.; Stahlberg, H.; Palczewski, K. Single particle cryo-EM of the complex between interphotoreceptor retinoid-binding protein and a monoclonal antibody. FASEB J. 2020, 34, 13918–13934. [Google Scholar] [CrossRef]
- Robotham, A.C.; Kelly, J.F. Chapter 1—LC-MS characterization of antibody-based therapeutics: Recent highlights and future prospects. In Approaches to the Purification, Analysis and Characterization of Antibody-Based Therapeutics; Matte, A., Ed.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 1–33. [Google Scholar]
- Kim, I.Y.; Suh, S.H.; Lee, I.K.; Wolfe, R.R. Applications of stable, nonradioactive isotope tracers in in vivo human metabolic research. Exp. Mol. Med. 2016, 48, e203. [Google Scholar] [CrossRef]
- Barbet, J.; Bardies, M.; Bourgeois, M.; Chatal, J.F.; Cherel, M.; Davodeau, F.; Faivre-Chauvet, A.; Gestin, J.F.; Kraeber-Bodere, F. Radiolabeled antibodies for cancer imaging and therapy. Methods Mol. Biol. 2012, 907, 681–697. [Google Scholar] [PubMed]
- Hiroaki, H. Recent applications of isotopic labeling for protein NMR in drug discovery. Expert Opin. Drug Discov. 2013, 8, 523–536. [Google Scholar] [CrossRef]
- Pan, S.; Aebersold, R.; Chen, R.; Rush, J.; Goodlett, D.R.; McIntosh, M.W.; Zhang, J.; Brentnall, T.A. Mass spectrometry based targeted protein quantification: Methods and applications. J. Proteome Res. 2009, 8, 787–797. [Google Scholar] [CrossRef] [PubMed]
- Reddy, P.T.; Brinson, R.G.; Hoopes, J.T.; McClung, C.; Ke, N.; Kashi, L.; Berkmen, M.; Kelman, Z. Platform development for expression and purification of stable isotope labeled monoclonal antibodies in Escherichia coli. MAbs 2018, 10, 992–1002. [Google Scholar] [PubMed]
- Martinovic, S.; Veenstra, T.D.; Anderson, G.A.; Pasa-Tolic, L.; Smith, R.D. Selective incorporation of isotopically labeled amino acids for identification of intact proteins on a proteome-wide level. J. Mass. Spectrom. 2002, 37, 99–107. [Google Scholar] [CrossRef]
- Ridgway, J.B.; Presta, L.G.; Carter, P. ‘Knobs-into-holes’ engineering of antibody CH3 domains for heavy chain heterodimerization. Protein Eng. 1996, 9, 617–621. [Google Scholar] [CrossRef]
- Bachmann, B.J. Pedigrees of some mutant strains of Escherichia coli K-12. Bacteriol. Rev. 1972, 36, 525–557. [Google Scholar] [CrossRef]
- Sutcliffe, J.G. Complete nucleotide sequence of the Escherichia coli plasmid pBR322. Cold Spring Harb. Symp. Quant. Biol. 1979, 43 Pt 1, 77–90. [Google Scholar] [CrossRef]
- Simmons, L.C.; Reilly, D.; Klimowski, L.; Raju, T.S.; Meng, G.; Sims, P.; Hong, K.; Shields, R.L.; Damico, L.A.; Rancatore, P.; et al. Expression of full-length immunoglobulins in Escherichia coli: Rapid and efficient production of aglycosylated antibodies. J. Immunol. Methods 2002, 263, 133–147. [Google Scholar] [CrossRef]
- Veeravalli, K.; Schindler, T.; Dong, E.; Yamada, M.; Hamilton, R.; Laird, M.W. Strain engineering to reduce acetate accumulation during microaerobic growth conditions in Escherichia coli. Biotechnol. Prog. 2018, 34, 303–314. [Google Scholar] [CrossRef] [PubMed]
- Taussky, H.H.; Shorr, E. A microcolorimetric method for the determination of inorganic phosphorus. J. Biol. Chem. 1953, 202, 675–685. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.J.; Giese, G.; Persson, J. Improved assembly of bispecific antibodies from knob and hole half-antibodies. Biotechnol. Prog. 2015, 31, 1315–1322. [Google Scholar] [CrossRef]
- Macchi, F.D.; Yang, F.; Li, C.; Wang, C.; Dang, A.N.; Marhoul, J.C.; Zhang, H.M.; Tully, T.; Liu, H.; Yu, X.C.; et al. Absolute Quantitation of Intact Recombinant Antibody Product Variants Using Mass Spectrometry. Anal. Chem. 2015, 87, 10475–10482. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Izadi, S.; Callahan, M.; Deperalta, G.; Wecksler, A.T. Antibody-receptor interactions mediate antibody-dependent cellular cytotoxicity. J. Biol. Chem. 2021, 297, 100826. [Google Scholar] [CrossRef]
- Klein, C.; Schaefer, W.; Regula, J.T. The use of CrossMAb technology for the generation of bi- and multispecific antibodies. MAbs 2016, 8, 1010–1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| WT Mass Predicted (Da) | Number of Carbons | Expected Mass Shift (Da) | 13C-labeling Mass Predicted (Da) | Mass Observed (Da) | 13C-Incorporation Calculated (%) | |
|---|---|---|---|---|---|---|
| hBsAb | 145,217 | 3210 | 3178 | 148,395 | 148,367 | 99.1 |
| Knob-hAb | 72,711 | --- | --- | --- | 72,712 | --- |
| Hole-hAb (Labeled) | 72,506 | 3210 | 3178 | 75,684 | 75,665 | 99.4 |
| Knob-HC | 48,912 | --- | --- | 48,912 | --- | |
| Hole-HC (labeled) | 48,707 | 2169 | 2147 | 50,854 | 50,835 | 99.1 |
| Knob-LC | 23,815 | --- | --- | --- | 23,815 | --- |
| Hole-LC (labeled) | 23,815 | 1041 | 1031 | 24,846 | 24,837 | 99.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wecksler, A.T.; Lundin, V.; Williams, A.J.; Veeravalli, K.; Reilly, D.E.; Grieco, S.-H. Bioprocess Development and Characterization of a 13C-Labeled Hybrid Bispecific Antibody Produced in Escherichia coli. Antibodies 2023, 12, 16. https://doi.org/10.3390/antib12010016
Wecksler AT, Lundin V, Williams AJ, Veeravalli K, Reilly DE, Grieco S-H. Bioprocess Development and Characterization of a 13C-Labeled Hybrid Bispecific Antibody Produced in Escherichia coli. Antibodies. 2023; 12(1):16. https://doi.org/10.3390/antib12010016
Chicago/Turabian StyleWecksler, Aaron T., Victor Lundin, Ambrose J. Williams, Karthik Veeravalli, Dorothea E. Reilly, and Sung-Hye Grieco. 2023. "Bioprocess Development and Characterization of a 13C-Labeled Hybrid Bispecific Antibody Produced in Escherichia coli" Antibodies 12, no. 1: 16. https://doi.org/10.3390/antib12010016