
In Vitro and In Vivo Characterization of a Broadly Neutralizing Anti-SARS-CoV-2 Antibody Isolated from a Semi-Immune Phage Display Library
 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Monoclonal Antibody Production and Purification
2.2. Developability Assessment
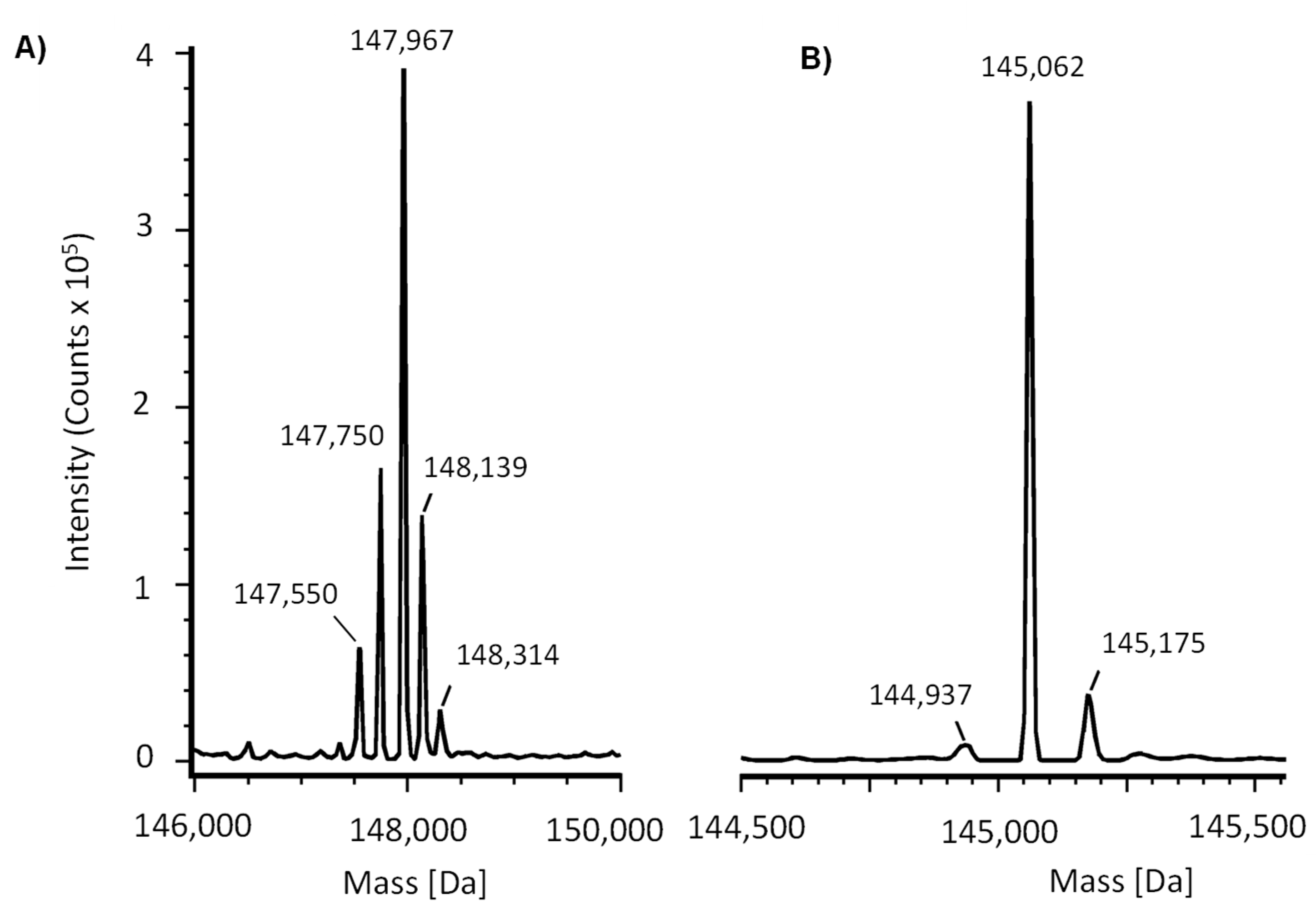
2.3. Mass Spectrometry Characterization
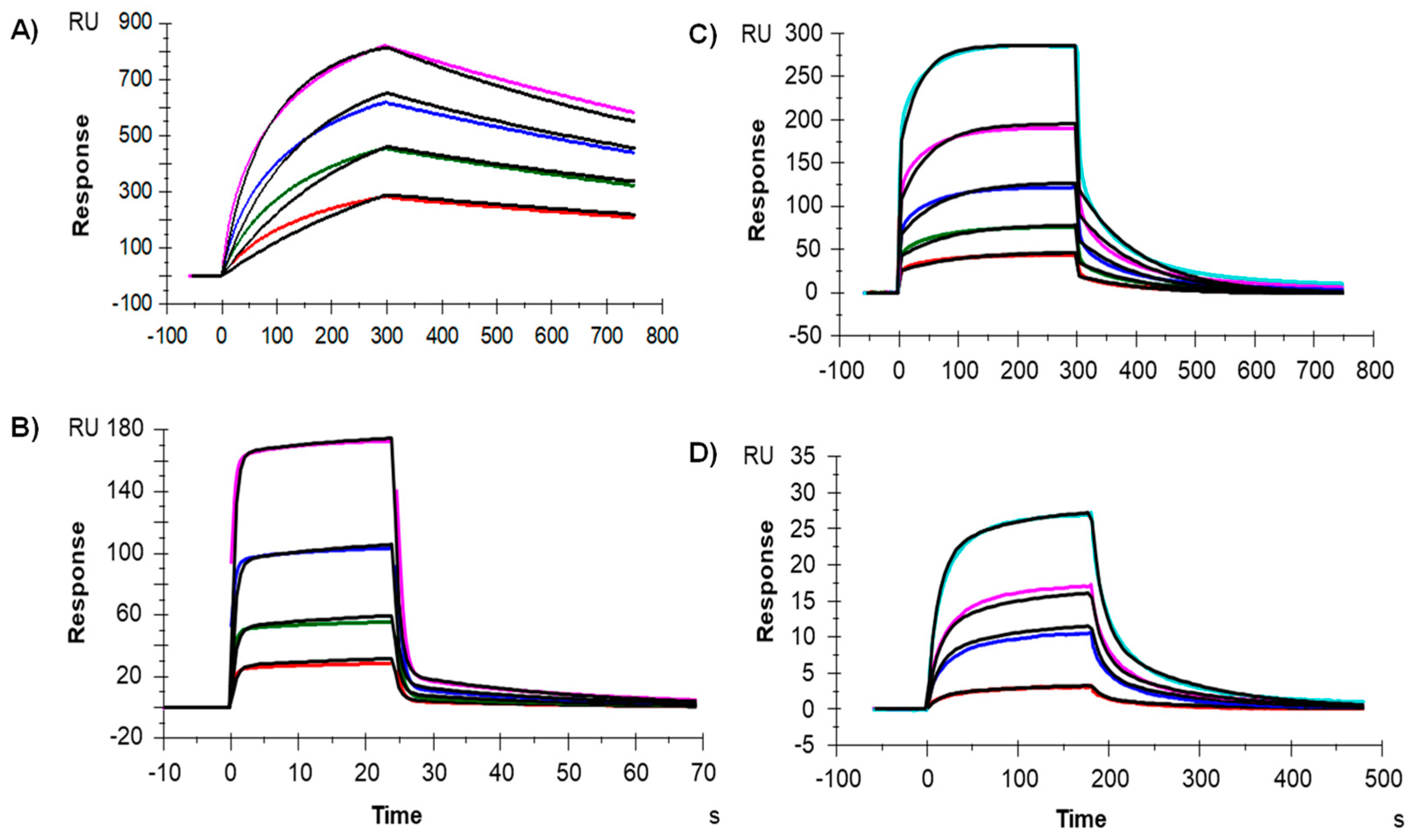
2.4. Binding to the Fcγ Receptors and FcRn by Surface Plasmon Resosnance (SPR)
2.5. SARS-CoV-2 Isolates
2.6. Plaque Reduction Neutralization Test (PRNT)
2.7. Efficacy Assay
3. Results
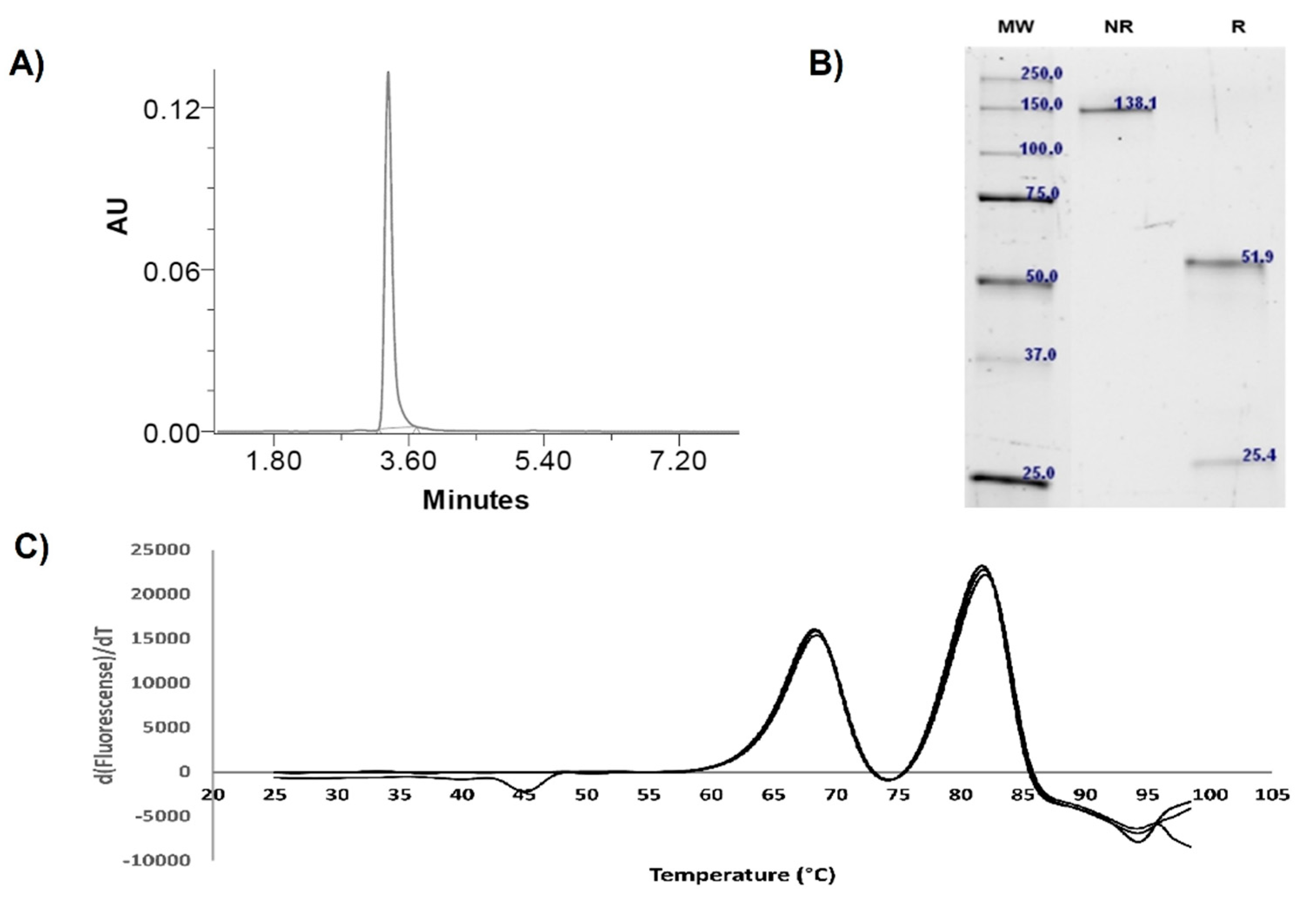
3.1. Expression and Physicochemical Characterization of IgG-A7
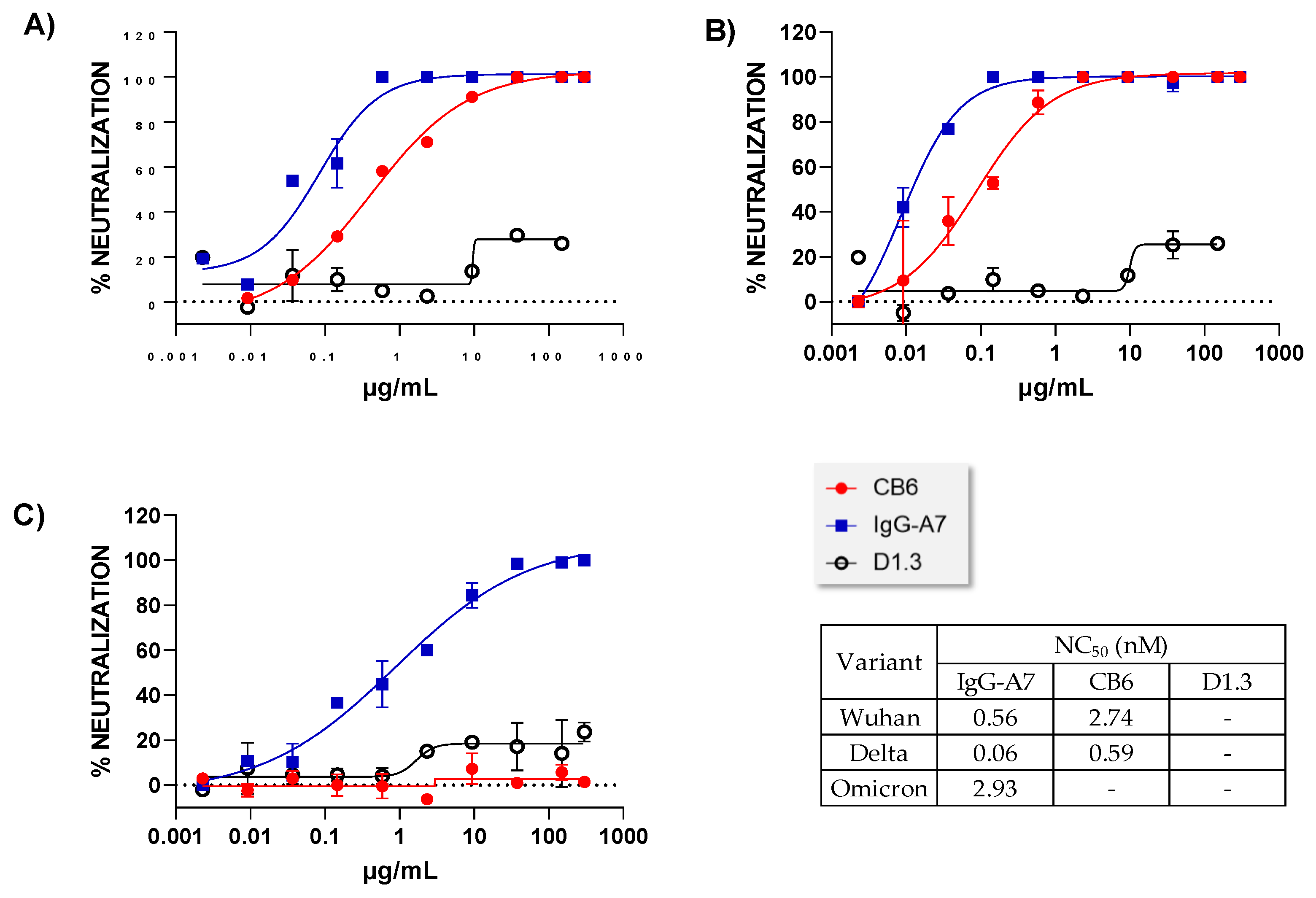
3.2. Neutralization Assay In Vitro
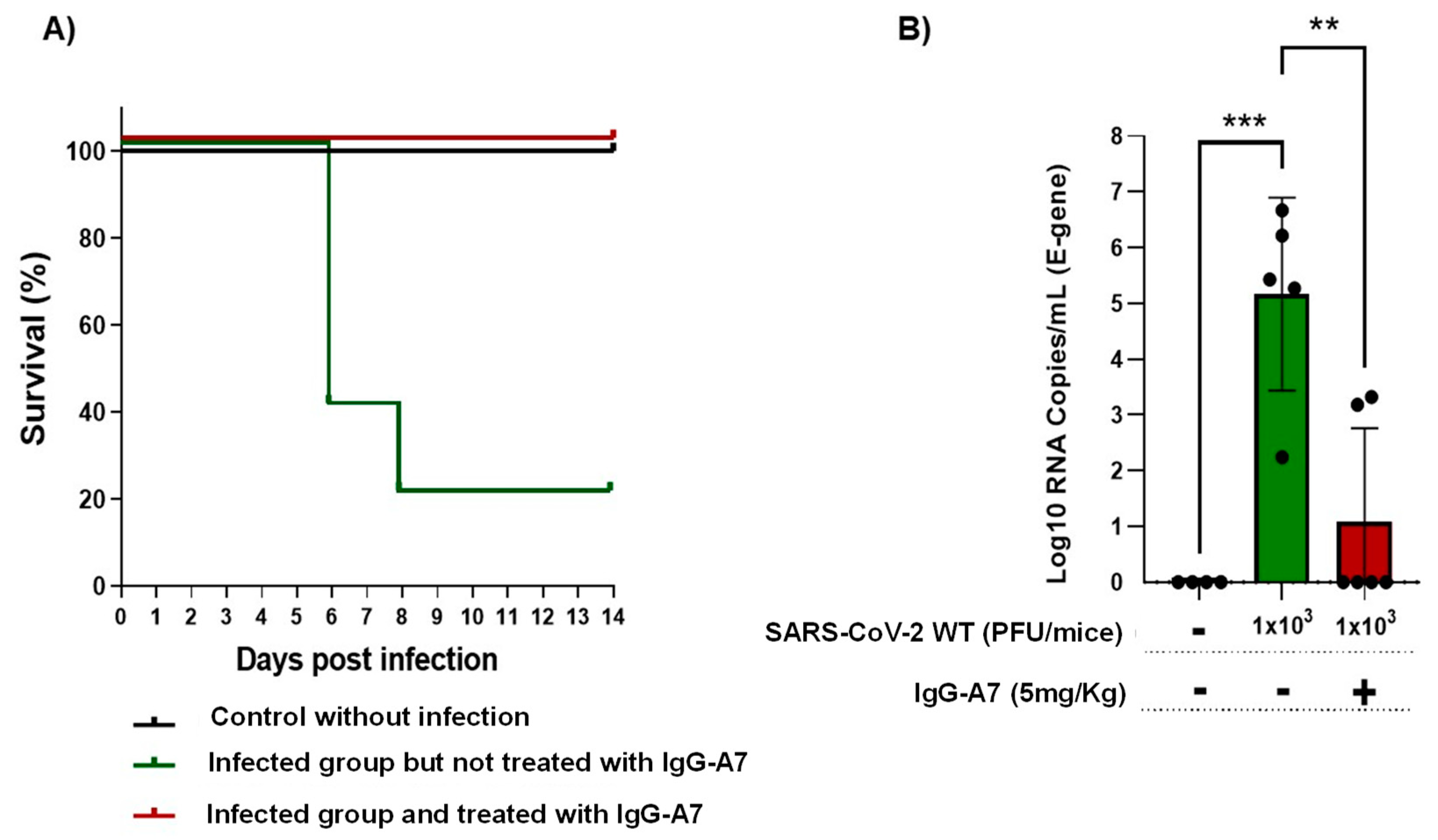
3.3. Efficacy in Transgenic Mice
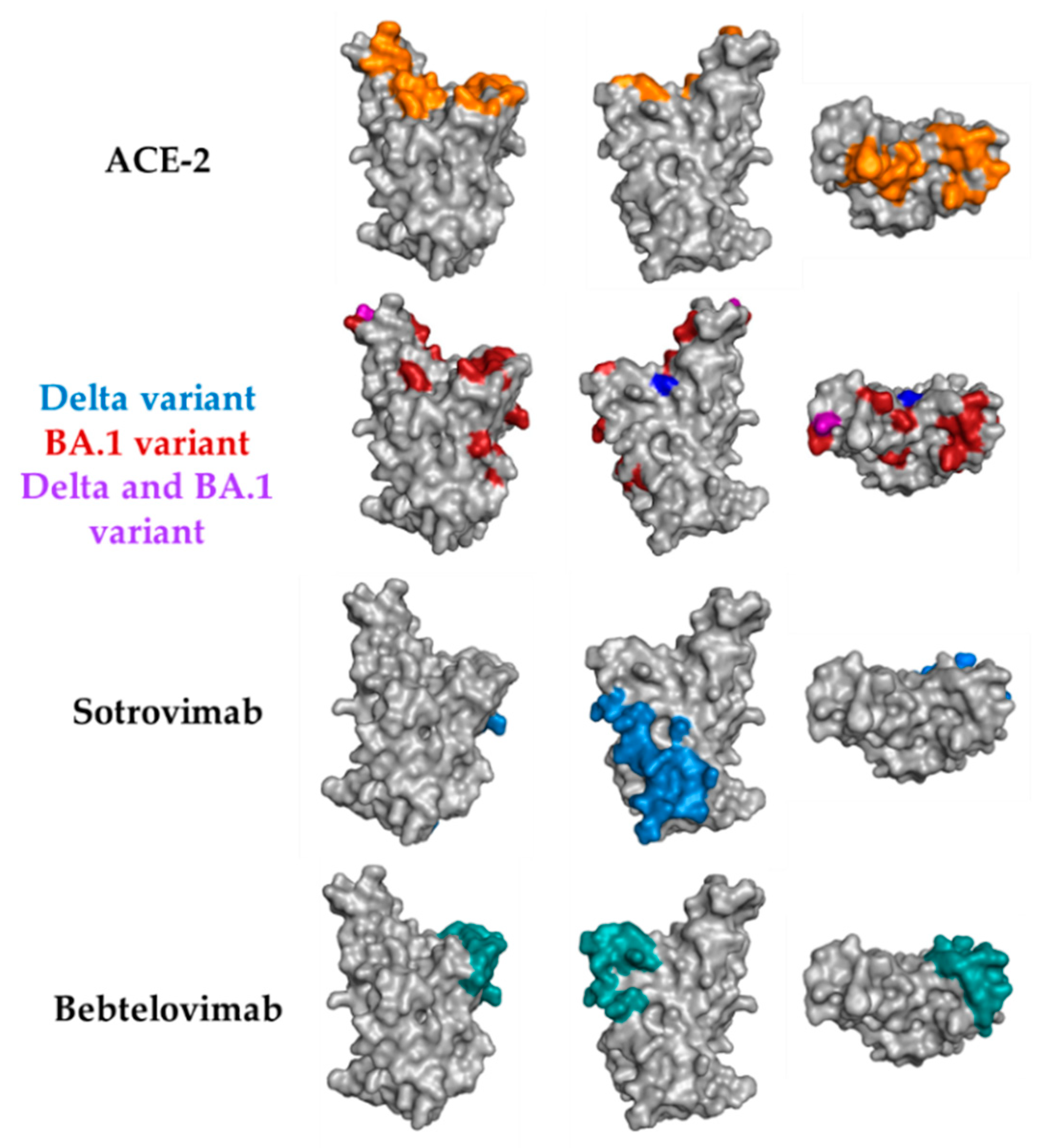
4. Discussion

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Attribute | Units | IgG-A7 | Success Criteria |
|---|---|---|---|
| Expression yield | mg/L | 20 | 12–16 |
| Endotoxin | UE/mL | <0.125 | <15 |
| Purity (SEC) | % | 100 | >95 |
| kDa | 205 | ~150 | |
| Integrity (SDS-PAGE) | Heavy chain (kDa) | 51.9 | ~50 |
| Light chain (kDa) | 25.4 | ~25 | |
| Whole molecule (kDa) | 138.1 | ~150 | |
| Thermal stability | Tm1 (°C) | 68.5 | 68 |
| Tm2 (°C) | 82.1 | 68–83 | |
| Intact Mass (Main peak) | Da [ppm] | 147,967 | Should correspond to the calculated mass based on the amino acid sequence plus glycans |
| Deglycosylated (Main peak) | Da [ppm] | 145,062 | Should correspond to the calculated mass based on the amino acid sequence |
| Fcγ receptors (reference values are those measured with a similar capture reagent, i.e., anti-HIS antibody) | FcγRI (µM) | 0.02 | 0.0009–0.052 [31] |
| FcγRIIA (µM) | 0.42 | 4.20–6.00 [31] | |
| FcγRIIIA (µM) | 0.38 | 0.089–2.166 [31] | |
| FcRn | µM | 2.74 | 0.9–4.3 [32] |
5. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cucinotta, D.; Vanelli, M. WHO Declares COVID-19 a Pandemic. Acta Biomed. 2020, 91, 157–160. [Google Scholar] [CrossRef] [PubMed]
- COVID Live—Coronavirus Statistics—Worldometer. Available online: https://www.worldometers.info/coronavirus/ (accessed on 11 June 2022).
- Zimmer, C.J.; Wee, S.; Kristoffersen, M. Coronavirus Vaccine Tracker, The New York Times. Available online: https://www.nytimes.com/interactive/2020/science/coronavirus-vaccine-tracker.html (accessed on 2 June 2022).
- Lazarevic, I.; Pravica, V.; Miljanovic, D.; Cupic, M. Immune Evasion of SARS-CoV-2 Emerging Variants: What Have We Learnt So Far? Viruses 2021, 13, 1192. [Google Scholar] [CrossRef] [PubMed]
- Almagro, J.C.; Mellado-Sánchez, G.; Pedraza-Escalona, M.; Pérez-Tapia, S.M. Evolution of approved anti-SARS-COV-2 therapeutic antibodies. Int. J. Mol. Sci. 2022, 23, 9763. [Google Scholar] [CrossRef]
- Deb, P.; Molla, M.M.A.; Saif-Ur-Rahman, K.M. An update to monoclonal antibody as therapeutic option against COVID-19. Biosaf. Health 2021, 3, 87–91. [Google Scholar] [CrossRef]
- Research C for DE and FDA Authorizes REGEN-COV Monoclonal Antibody Therapy for Post-Exposure Prophylaxis (Prevention) for COVID-19. FDA. Available online: https://www.fda.gov/drugs/drug-safety-and-availability/fda-authorizes-regen-cov-monoclonal-antibody-therapy-post-exposure-prophylaxis-prevention-COVID-19 (accessed on 16 June 2022).
- Touret, F.; Baronti, C.; Bouzidi, H.S.; de Lamballerie, X. In vitro evaluation of therapeutic antibodies against a SARS-CoV-2 Omicron B.1.1.529 isolate. Sci. Rep. 2022, 12, 4683. [Google Scholar] [CrossRef]
- Lusvarghi, S.; Pollett, S.D.; Neerukonda, S.N.; Wang, W.; Wang, R.; Vassell, R.; Epsi, N.J.; Fries, A.C.; Agan, B.K.; Lindholm, D.A.; et al. SARS-CoV-2 Omicron neutralization by therapeutic antibodies, convalescent sera, and post-mRNA vaccine booster. bioRxiv 2021. [Google Scholar] [CrossRef]
- Table: Characteristics of Anti-SARS-CoV-2 Antibody Products. COVID-19 Treatment Guidelines. Available online: https://www.covid19treatmentguidelines.nih.gov/tables/mab-characteristics/ (accessed on 2 June 2022).
- Tracking SARS-CoV-2 Variants. Available online: https://www.who.int/activities/tracking-SARS-CoV-2-variants (accessed on 17 June 2022).
- Mendoza-Salazar, I.; Gomez-Castellano, K.M.; Gonzalez-Gonzalez, E.; Gamboa-Suasnavart, R.; Rodriguez-Luna, S.D.; Santiago-Casas, G.; Cortes-Paniagua, M.I.; Perez-Tapia, S.M.; Almagro, J.C. Anti-SARS-CoV-2 Omicron Antibodies Isolated from a SARS-CoV-2 Delta Semi-Immune Phage Display Library. Antibodies 2022, 11, 13. [Google Scholar] [CrossRef]
- Research C for DE and FDA Authorizes Bamlanivimab and Etesevimab Monoclonal Antibody Therapy for Post-Exposure Prophylaxis (Prevention) for COVID-19. FDA. Available online: https://www.fda.gov/drugs/drug-safety-and-availability/fda-authorizes-bamlanivimab-and-etesevimab-monoclonal-antibody-therapy-post-exposure-prophylaxis (accessed on 15 June 2022).
- Shi, R.; Shan, C.; Duan, X.; Chen, Z.; Liu, P.; Song, J.; Song, T.; Bi, X.; Han, C.; Wu, L.; et al. A human neutralizing antibody targets the receptor-binding site of SARS-CoV-2. Nature 2020, 584, 120–124. [Google Scholar] [CrossRef]
- Wu, X.; Li, N.; Wang, G.; Liu, W.; Yu, J.; Cao, G.; Wang, J.; Chen, Y.; Ma, J.; Wu, J.; et al. Tolerability, Safety, Pharmacokinetics, and Immunogenicity of a Novel SARS-CoV-2 Neutralizing Antibody, Etesevimab, in Chinese Healthy Adults: A Randomized, Double-Blind, Placebo-Controlled, First-in-Human Phase 1 Study. Antimicrob. Agents Chemother. 2021, 65, e0035021. [Google Scholar] [CrossRef]
- Harper, M.L.F.; Boulot, G.; Poljak, R.J. Antigen specificity and cross-reactivity of monoclonal anti-lysozyme antibodies. Mol. Immunol. 1987, 24, 97–108. [Google Scholar] [CrossRef]
- Perdomo-Abundez, F.C.; Vallejo-Castillo, L.; Vazquez-Leyva, S.; Lopez-Morales, C.A.; Velasco-Velazquez, M.; Pavon, L.; Perez-Tapia, S.M.; Medina-Rivero, E. Development and validation of a mass spectrometric method to determine the identity of rituximab based on its microheterogeneity profile. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2020, 1139, 121885. [Google Scholar] [CrossRef] [PubMed]
- Menzen, T.; Friess, W. High-throughput melting-temperature analysis of a monoclonal antibody by differential scanning fluorimetry in the presence of surfactants. J. Pharm. Sci. 2013, 102, 415–428. [Google Scholar] [CrossRef] [PubMed]
- Camacho-Sandoval, R.; Nieto-Patlan, A.; Carballo-Uicab, G.; Montes-Luna, A.; Jimenez-Martinez, M.C.; Vallejo-Castillo, L.; Gonzalez-Gonzalez, E.; Arrieta-Oliva, H.I.; Gomez-Castellano, K.; Guzman-Bringas, O.U.; et al. Development and Evaluation of a Set of Spike and Receptor Binding Domain-Based Enzyme-Linked Immunosorbent Assays for SARS-CoV-2 Serological Testing. Diagnostics 2021, 11, 1506. [Google Scholar] [CrossRef] [PubMed]
- Bewley, K.R.; Coombes, N.S.; Gagnon, L.; McInroy, L.; Baker, N.; Shaik, I.; St-Jean, J.R.; St-Amant, N.; Buttigieg, K.R.; Humphries, H.E.; et al. Quantification of SARS-CoV-2 neutralizing antibody by wild-type plaque reduction neutralization, microneutralization and pseudotyped virus neutralization assays. Nat. Protoc. 2021, 16, 3114–3140. [Google Scholar] [CrossRef]
- WMA—The World Medical Association-Declaración de Helsinki de la AMM—Principios Éticos Para las Investigaciones Médicas en Seres Humanos. Available online: https://www.wma.net/es/policies-post/declaracion-de-helsinki-de-la-amm-principios-eticos-para-las-investigaciones-medicas-en-seres-humanos/ (accessed on 11 June 2022).
- Guidance-for-BSL2+ COVID-Laboratories-Anschutz.pdf. Available online: https://research.cuanschutz.edu/docs/librariesprovider148/ehs_documents/default-library/guidance-for-bsl2-covid-laboratories-anschutz.pdf?sfvrsn=79c7e4b9_0 (accessed on 21 June 2022).
- Biosafety in Microbiological and Biomedical Laboratories—6th Edition. Available online: https://www.cdc.gov/labs/pdf/SF__19_308133-A_BMBL6_00-BOOK-WEB-final-3.pdf (accessed on 11 June 2022).
- Laboratory Biosafety Manual, 4th Edition. Available online: https://www.who.int/publications-detail-redirect/9789240011311 (accessed on 11 June 2022).
- CDC. Labs. Centers for Disease Control and Prevention. Published 11 February 2020. Available online: https://www.cdc.gov/coronavirus/2019-ncov/lab/lab-safety-practices.html (accessed on 21 June 2022).
- Norma Oficial Mexicana NOM-062-ZOO-1999; Especificaciones técnicas para la producción, cuidado y uso de los animales de laboratorio. Diario Oficial de la Federación: Ciudad de México, Mexico, 2001.
- Gilliland, G.L.; Luo, J.; Vafa, O.; Almagro, J.C. Leveraging SBDD in protein therapeutic development: Antibody engineering. Methods Mol. Biol. 2012, 841, 321–349. [Google Scholar] [CrossRef]
- Junker, F.; Gordon, J.; Qureshi, O. Fc Gamma Receptors and Their Role in Antigen Uptake, Presentation, and T Cell Activation. Front. Immunol. 2020, 11, 1393. [Google Scholar] [CrossRef]
- Iketani, S.; Liu, L.; Guo, Y.; Liu, L.; Chan, J.F.; Huang, Y.; Wang, M.; Luo, Y.; Yu, J.; Chu, H.; et al. Antibody evasion properties of SARS-CoV-2 Omicron sublineages. Nature 2022, 604, 553–556. [Google Scholar] [CrossRef]
- Fact Sheet for Health Care Providers Emergency Use Authorization (Eua) of Bamlanivimab and Etesevimab. Available online: https://www.fda.gov/media/145802/download (accessed on 11 June 2022).
- Forest-Nault, C.; Gaudreault, J.; Henry, O.; Durocher, Y.; De Crescenzo, G. On the Use of Surface Plasmon Resonance Biosensing to Understand IgG-FcγR Interactions. Int. J. Mol. Sci. 2021, 22, 6616. [Google Scholar] [CrossRef]
- Wang, X.; McKay, P.; Yee, L.T.; Dutina, G.; Hass, P.E.; Nijem, I.; Allison, D.; Cowan, K.J.; Lin, K.; Quarmby, V.; et al. Impact of SPR biosensor assay configuration on antibody: Neonatal Fc receptor binding data. MAbs 2017, 9, 319–332. [Google Scholar] [CrossRef]
- VanBlargan, L.A.; Errico, J.M.; Halfmann, P.J.; Zost, S.J.; Crowe, J.E., Jr.; Purcell, L.A.; Kawaoka, Y.; Corti, D.; Fremont, D.H.; Diamond, M.S. An infectious SARS-CoV-2 B.1.1.529 Omicron virus escapes neutralization by therapeutic monoclonal antibodies. Nat. Med. 2022, 28, 490–495. [Google Scholar] [CrossRef]
- Westendorf, K.; Zentelis, S.; Wang, L.; Foster, D.; Vaillancourt, P.; Wiggin, M.; Lovett, E.; van der Lee, R.; Hendle, J.; Pustilnik, A.; et al. LY-CoV1404 (bebtelovimab) potently neutralizes SARS-CoV-2 variants. Cell Rep. 2022, 39, 110812. [Google Scholar] [CrossRef] [PubMed]






| (INN) | D614G | (Omicron) |
|---|---|---|
| Casirivimab plus Imdevimab | ++++ | − |
| Bamlanivimab plus Etesivimab | ++++ | − |
| Cilgavimab plus Tixagevimab | ++++ | Cilgavimab (++) |
| Tixagevimab (+) | ||
| Regdanvimab | ++++ | − |
| Sotrovimab | ++ | ++ |
| Bebtelovimab | ++++ | ++++ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-González, E.; Carballo-Uicab, G.; Salinas-Trujano, J.; Cortés-Paniagua, M.I.; Vázquez-Leyva, S.; Vallejo-Castillo, L.; Mendoza-Salazar, I.; Gómez-Castellano, K.; Pérez-Tapia, S.M.; Almagro, J.C. In Vitro and In Vivo Characterization of a Broadly Neutralizing Anti-SARS-CoV-2 Antibody Isolated from a Semi-Immune Phage Display Library. Antibodies 2022, 11, 57. https://doi.org/10.3390/antib11030057
González-González E, Carballo-Uicab G, Salinas-Trujano J, Cortés-Paniagua MI, Vázquez-Leyva S, Vallejo-Castillo L, Mendoza-Salazar I, Gómez-Castellano K, Pérez-Tapia SM, Almagro JC. In Vitro and In Vivo Characterization of a Broadly Neutralizing Anti-SARS-CoV-2 Antibody Isolated from a Semi-Immune Phage Display Library. Antibodies. 2022; 11(3):57. https://doi.org/10.3390/antib11030057
Chicago/Turabian StyleGonzález-González, Edith, Gregorio Carballo-Uicab, Juana Salinas-Trujano, María I. Cortés-Paniagua, Said Vázquez-Leyva, Luis Vallejo-Castillo, Ivette Mendoza-Salazar, Keyla Gómez-Castellano, Sonia M. Pérez-Tapia, and Juan C. Almagro. 2022. "In Vitro and In Vivo Characterization of a Broadly Neutralizing Anti-SARS-CoV-2 Antibody Isolated from a Semi-Immune Phage Display Library" Antibodies 11, no. 3: 57. https://doi.org/10.3390/antib11030057