
Weed Seed Banks in Intensive Farmland and the Influence of Tillage, Field Position, and Sown Flower Strips
Abstract
:1. Introduction
2. Materials and Methods
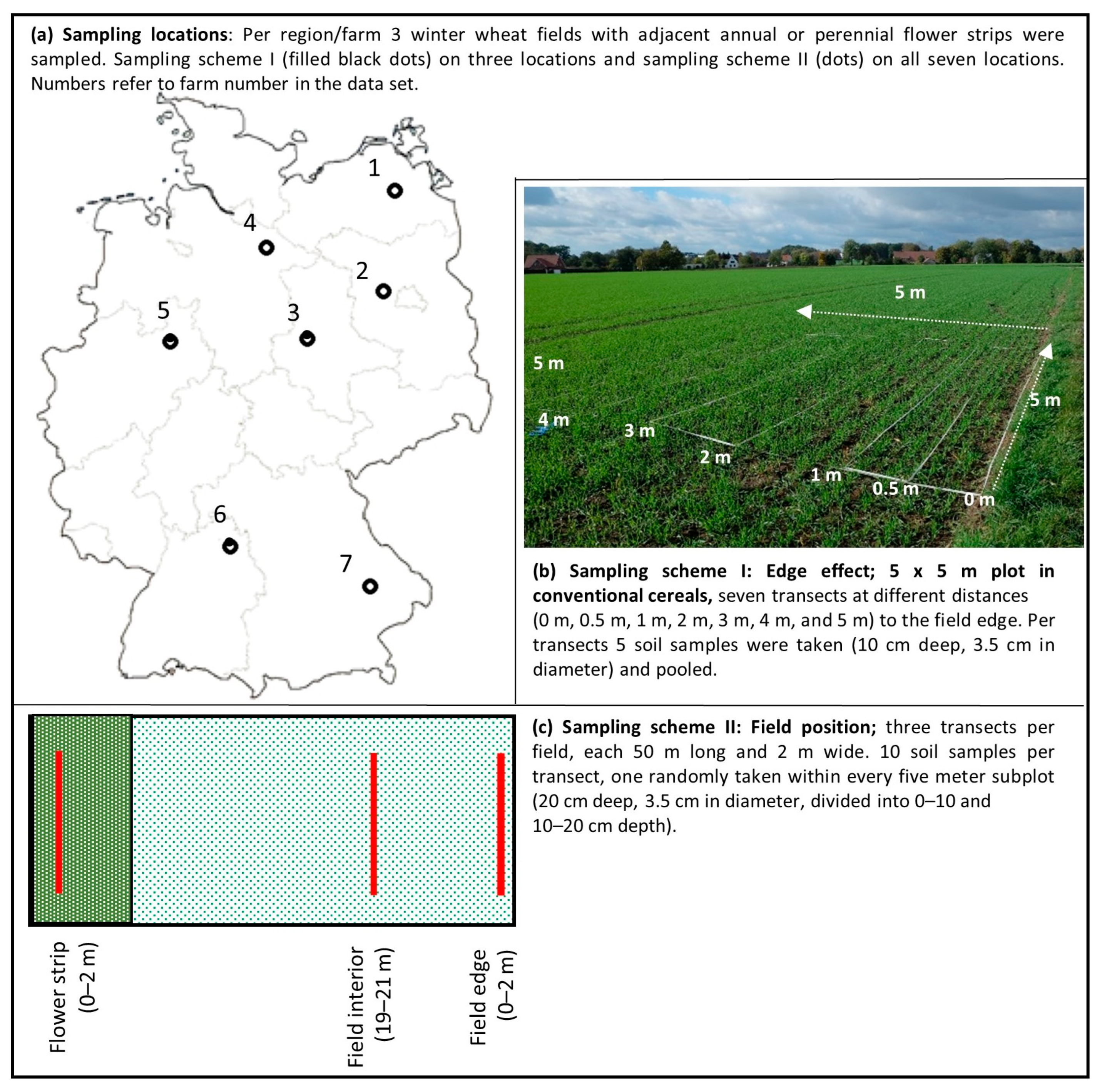
2.1. Locations and Sampling Design
2.2. Sampling Scheme I Edge Effect: Analysis of the Effect of Field Edge
2.3. Sampling Scheme II Field Position: Seed Bank Enrichment in Flower Strips
2.4. Seedling Emergence Method
3. Statistical Analysis
4. Results
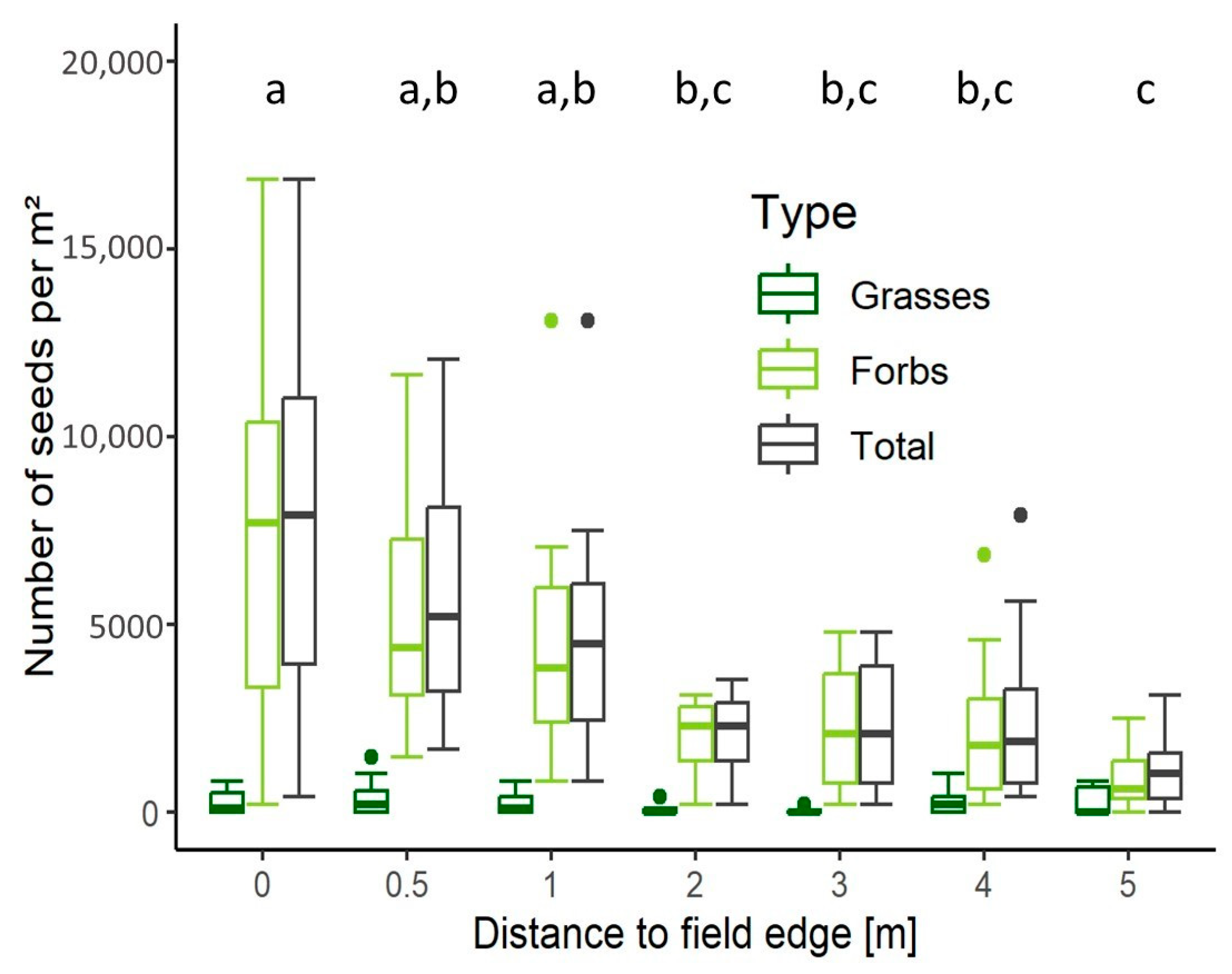
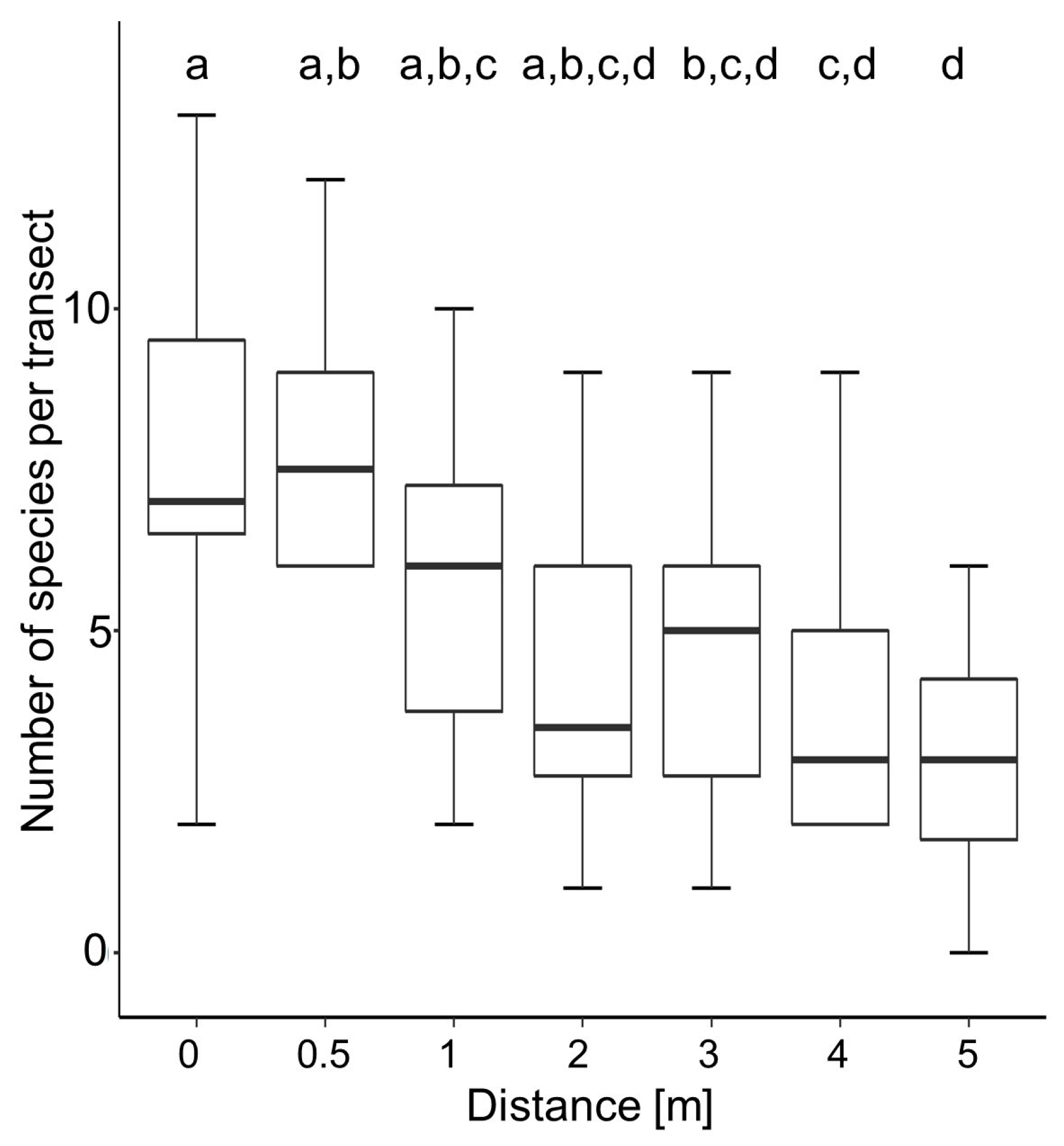
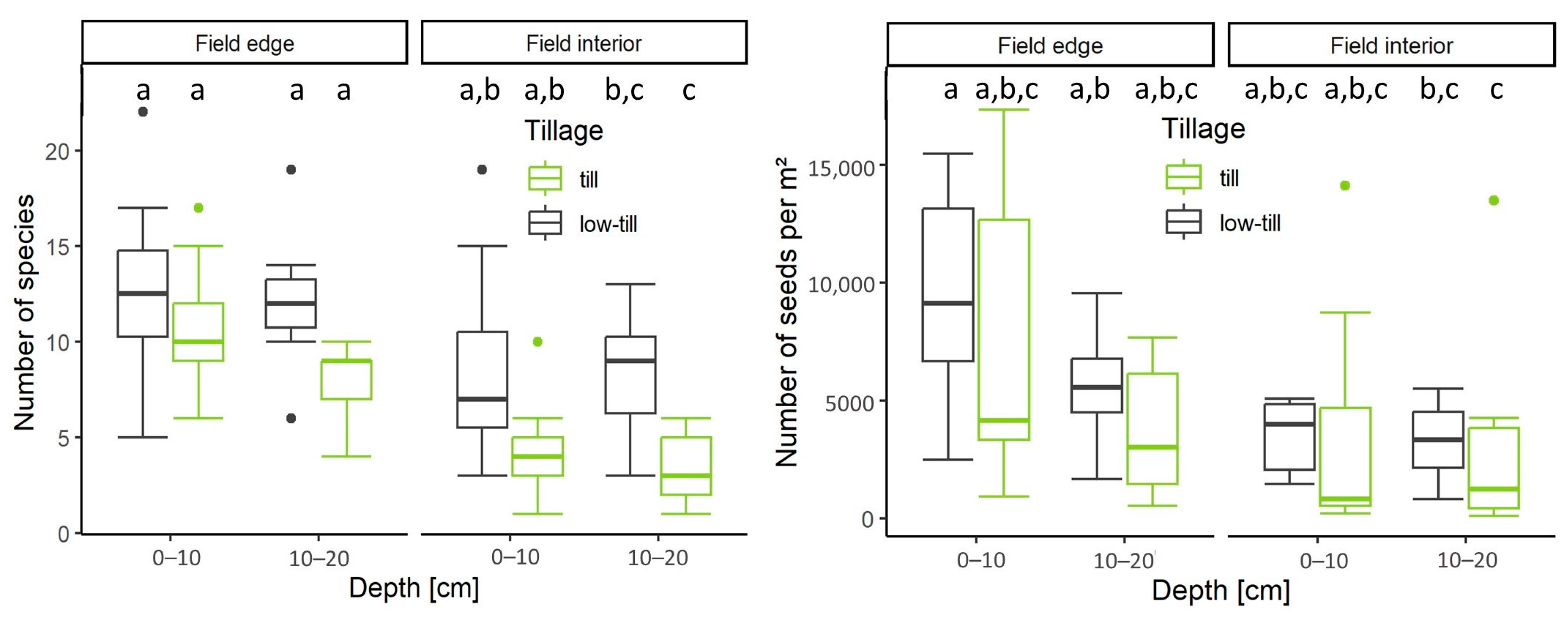
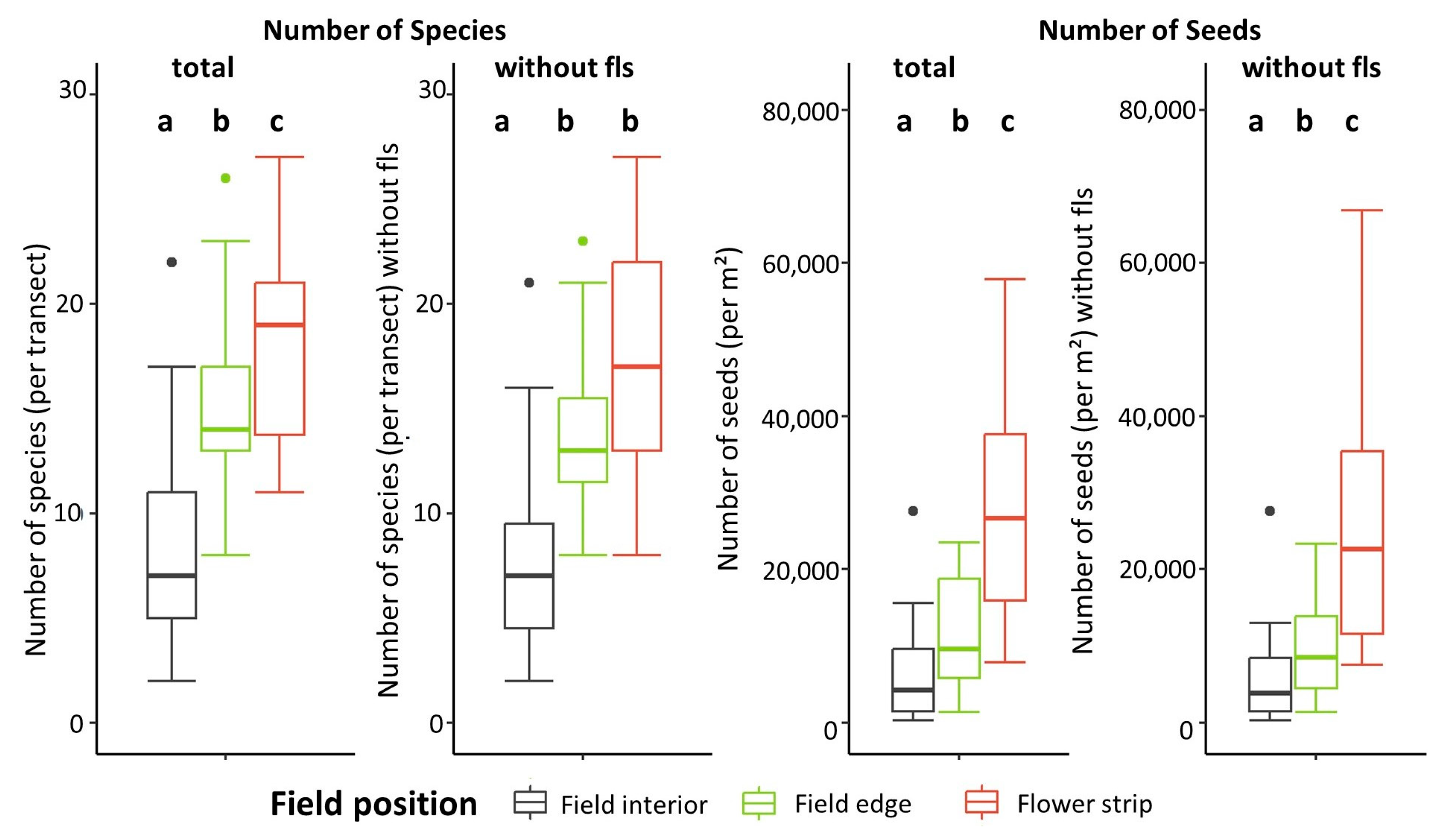
4.1. Number of Species and Seeds in Relation to Tillage Regime, Soil Depth, and Distance to Field Edge
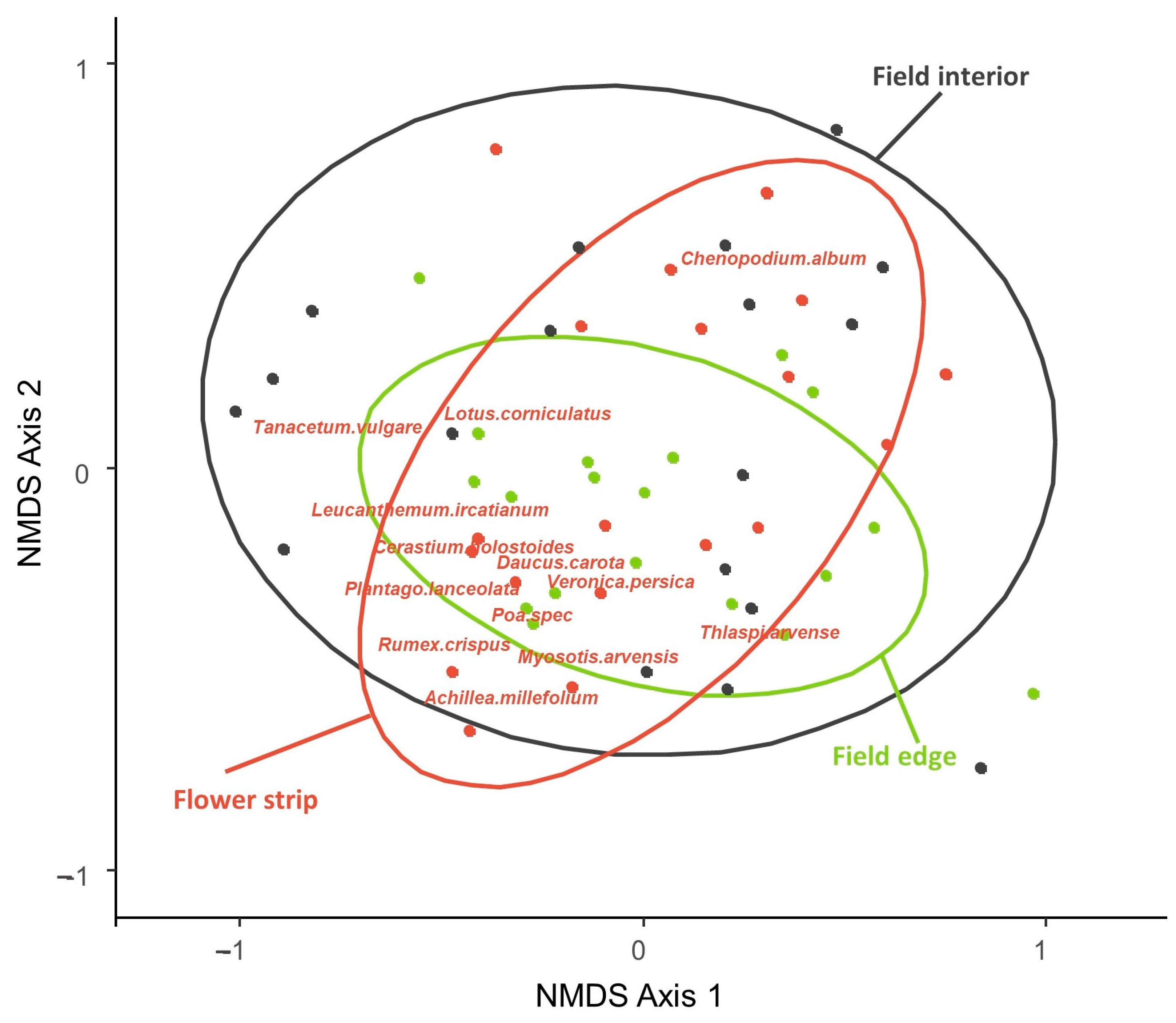
4.2. Differences in Abundance, Species Numbers, and Species Composition of the Seed Bank According to Field Location
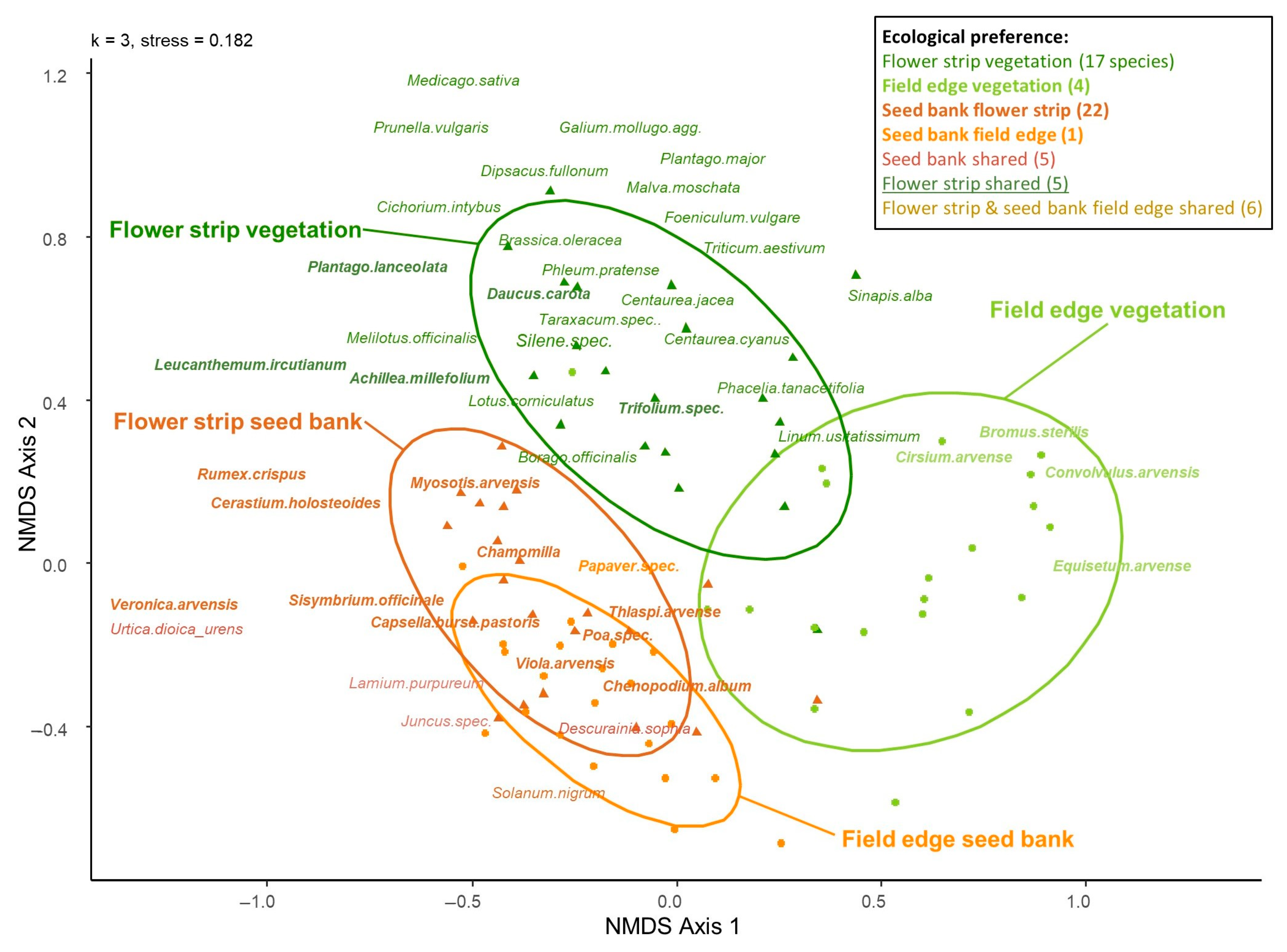
4.3. Differences in the Composition of Seed Banks and Associated Above-Ground Vegetation
5. Discussion
5.1. Edge Effect and Low-Tillage Increase Seed Bank Density in Arable Soils
5.2. Flower Strips Enrich the Quality and Quantity of Seed Banks
5.3. Larger Differences between Aboveground Vegetation and Seed Bank in the Field than in Flower Strips
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rollin, O.; Benelli, G.; Benvenuti, S.; Decourtye, A.; Wratten, S.D.; Canale, A.; Dexneus, N. Weed-insect pollinator networks as bio-indicators of ecological sustainability in agriculture. A review. Agron. Sustain. Dev. 2016, 36, 8. [Google Scholar] [CrossRef]
- Zerbe, S. Species-Rich Arable Land. In Restoration of Ecosystems–Bridging Nature and Humans: A Transdisciplinary Approach; Springer: Berlin/Heidelberg, Germany, 2023; pp. 393–407. [Google Scholar]
- Leuschner, C.; Ellenberg, H. Ecology of Central European Non-Forest Vegetation: Coastal to Alpine, Natural to Man-Made Habitats: Vegetation Ecology of Central Europe, Volume II.; Springer: Cham, Switzerland, 2017. [Google Scholar]
- Richner, N.; Holderegger, R.; Linder, H.P.; Walter, T. Reviewing change in the arable flora of Europe: A meta-analysis. Weed Res. 2015, 55, 1–13. [Google Scholar] [CrossRef]
- Meyer, S.; Wesche, K.; Krause, B.; Leuschner, C. Dramatic losses of specialist arable plants in Central Germany since the 1950s/60s—A cross-regional analysis. Divers. Distrib. 2013, 19, 1175–1187. [Google Scholar] [CrossRef]
- Fischer, A.; Milberg, P. Effects on the flora of extensified use of field margins. Swed. J. Agric. Res. 1997, 27, 105–112. [Google Scholar]
- Wietzke, A.; Albert, K.; Bergmeier, E.; Sutcliffe, L.M.E.; van Waveren, C.-S.; Leuschner, C. Flower strips, conservation field margins and fallows promote the arable flora in intensively farmed landscapes: Results of a 4-year study. Agric. Ecosyst. Environ. 2020, 304, 107142. [Google Scholar] [CrossRef]
- Hoffmann, H.; Peter, F.; Herrmann, J.D.; Donath, T.W.; Diekoetter, T. Benefits of wildflower areas as overwintering habitats for ground-dwelling arthropods depend on landscape structural complexity. Agric. Ecosyst. Environ. 2021, 314, 107421. [Google Scholar] [CrossRef]
- Barreiro-Hurlé, J.; Espinosa-Goded, M.; Dupraz, P. Does intensity of change matter? Factors affecting adoption of agri-environmental schemes in Spain. J. Environ. Plan. Manag. 2010, 53, 891–905. [Google Scholar] [CrossRef]
- Joormann, I.; Schmidt, T. F.R.A.N.Z.-Studie—Hinternisse und Perspektiven für mehr Biodiversität in der Agrarlandschaft: Thünen Working Paper 75; Thünen-Institut: Braunschweig, Germany, 2017. [Google Scholar]
- Espinosa-Goded, M.; Barreiro-Hurlé, J.; Ruto, E. What do farmers want from agri-environmental scheme design? A choice experiment approach. J. Agric. Econ. 2010, 61, 259–273. [Google Scholar] [CrossRef]
- Kleijn, D.; Baquero, R.A.; Clough, Y.; Díaz, M.; de Esteban, J.; Fernández, F.; Gabriel, D.; Herzog, F.; Holzschuh, A.; Jöhl, R.; et al. Mixed biodiversity benefits of agri-environment schemes in five European countries. Ecol. Lett. 2006, 9, 243–254. [Google Scholar] [CrossRef]
- Melander, B.; Munier-Jolain, N.; Charles, R.; Wirth, J.; Schwarz, J.; van der Weide, R.; Bonin, L.; Jensen, P.K.; Kudsk, P. European perspectives on the adoption of nonchemical weed management in reduced-tillage systems for arable crops. Weed Technol. 2013, 27, 231–240. [Google Scholar] [CrossRef]
- Albrecht, H.; Auerswald, K. Seed traits in arable weed seed banks and their relationship to land-use changes. Basic Appl. Ecol. 2009, 10, 516–524. [Google Scholar] [CrossRef]
- Thompson, K.; Bakker, J.P.; Bekker, R.M.; Hodgson, J.G. Ecological correlates of seed persistence in soil in the north-west European flora. J. Ecol. 1998, 86, 163–169. [Google Scholar] [CrossRef]
- Lang, M.; Prestele, J.; Wiesinger, K.; Kollmann, J.; Albrecht, H. Reintroduction of rare arable plants: Seed production, soil seed banks, and dispersal 3 years after sowing. Restor. Ecol. 2018, 26, S170–S178. [Google Scholar] [CrossRef]
- Görzen, E.; Diekötter, T.; Meyerink, M.; Kretzschmar, H.; Donath, T.W. The Potential to Save Agrestal Plant Species in an Intensively Managed Agricultural Landscape through Organic Farming—A Case Study from Northern Germany. Land 2021, 10, 219. [Google Scholar] [CrossRef]
- Armengot, L.; Blanco-Moreno, J.M.; Bàrberi, P.; Bocci, G.; Carlesi, S.; Aendekerk, R.; Berner, A.; Celette, F.; Grosse, M.; Huiting, H.; et al. Tillage as a driver of change in weed communities: A functional perspective. Agric. Ecosyst. Environ. 2016, 222, 276–285. [Google Scholar] [CrossRef]
- Derpsch, R. Conservation Tillage, No-Tillage and Related Technologies. In Conservation Agriculture; Springer: Berlin/Heidelberg, Germany, 2003; pp. 181–190. [Google Scholar]
- Ghersa, C.M.; Martınez-Ghersa, M.A. Ecological correlates of weed seed size and persistence in the soil under different tilling systems: Implications for weed management. Field Crops Res. 2000, 67, 141–148. [Google Scholar] [CrossRef]
- Burmeier, S.; Eckstein, R.L.; Donath, T.W.; Otte, A. Plant pattern development during early post-restoration succession in Grasslands—A case study of Arabis nemorensis. Restor. Ecol. 2011, 19, 648–659. [Google Scholar] [CrossRef]
- Wilson, P.J.; Aebischer, N.J. The distribution of dicotyledonous arable weeds in relation to distance from the field edge. J. Appl. Ecol. 1995, 32, 295–310. [Google Scholar] [CrossRef]
- Fried, G.; Petit, S.; Dessaint, F.; Reboud, X. Arable weed decline in Northern France: Crop edges as refugia for weed conservation? Biol. Conserv. 2009, 142, 238–243. [Google Scholar] [CrossRef]
- Yvoz, S.; Cordeau, S.; Zuccolo, C.; Petit, S. Crop type and within-field location as sources of intraspecific variations in the phenology and the production of floral and fruit resources by weeds. Agric. Ecosyst. Environ. 2020, 302, 107082. [Google Scholar] [CrossRef]
- Simmering, D.; Waldhardt, R.; Otte, A. Erfassung und Analyse der Pflanzenartenvielfalt in der “Normallandschaft”—Ein Beispiel aus Mittelhessen. Ber. Reinhard-Tüxen-Ges. 2013, 25, 73–94. [Google Scholar]
- Kleijn, D.; van der Voort, L.A.C. Conservation headlands for rare arable weeds: The effects of fertilizer application and light penetration on plant growth. Biol. Conserv. 1997, 81, 57–67. [Google Scholar] [CrossRef]
- Devlaeminck, R.; Bossuyt, B.; Hermy, M. Inflow of seeds through the forest edge: Evidence from seed bank and vegetation patterns. Plant Ecol. 2005, 176, 1–17. [Google Scholar] [CrossRef]
- Albrecht, H.; Pilgram, M. The weed seed bank of soils in a landscape segment in southern Bavaria–II. Relation to environmental variables and to the surface vegetation. Plant Ecol. 1997, 131, 31–43. [Google Scholar] [CrossRef]
- Bossuyt, B.; Honnay, O. Can the seed bank be used for ecological restoration? An overview of seed bank characteristics in European communities. J. Veg. Sci. 2008, 19, 875–884. [Google Scholar] [CrossRef]
- Hopfensperger, K.N. A review of similarity between seed bank and standing vegetation across ecosystems. Oikos 2007, 116, 1438–1448. [Google Scholar] [CrossRef]
- Lang, M.; Kollmann, J.; Prestele, J.; Wiesinger, K.; Albrecht, H. Reintroduction of rare arable plants in extensively managed fields: Effects of crop type, sowing density and soil tillage. Agric. Ecosyst. Environ. 2021, 306, 107187. [Google Scholar] [CrossRef]
- Ter Heerdt, G.N.; Verweij, G.L.; Bekker, R.M.; Bakker, J.P. An improved method for seed-bank analysis: Seedling emergence after removing the soil by sieving. Funct. Ecol. 1996, 10, 144–151. [Google Scholar] [CrossRef]
- Hanf, M. Ackerunkräuter Europas mit Ihren Keimlingen und Samen, 2nd ed.; BLV Verlagsgesellschaft: Frankfrut, Germany, 1984. [Google Scholar]
- Jäger, E.J.; Werner, K. Rothmaler-Exkursionsflora von Deutschland; Springer: Berlin/Heidelberg, Germany, 2011. [Google Scholar]
- Ellenberg, H. Zeigerwerte von Pflanzen in Mitteleuropa. Scr. Geobot. 1991, 18, 1–248. [Google Scholar]
- Sutcliffe, L.; Leuschner, C. Auswirkungen von Biodiversitätsmaßnahmen auf die Segetalflo-ra auf intensiv bewirtschafteten landwirtschaftlichen Flächen—Ergebnisse aus dem F.R.A.N.Z.-Projekt. Naturschutz Landsch. 2022, 54, 22–29. [Google Scholar]
- Bates, D.; Sarkar, D.; Bates, M.D.; Matrix, L. The lme4 package. R Package Version 1.1.31 2007, 2, 74. [Google Scholar]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. Package ‘lmertest’. R Package Version 3.1.3 2015, 2, 734. [Google Scholar]
- RStudio Team. RStudio: Integrated Development Environment for R. RStudio; P.B.C.: Boston, MA, USA, 2021. [Google Scholar]
- Lenth, R.V.; Buerkner, P.; Herve, M.; Love, J.; Riebl, H.; Singmann, H. Emmeans: Estimated Marginal Means, aka Least-Squares Means; 2021. Available online: https://cran.microsoft.com/snapshot/2018-01-13/web/packages/emmeans/emmeans.pdf (accessed on 1 July 2022).
- Hartig, F. DHARMa: Residual Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models. 2021. Available online: http://florianhartig.github.io/DHARMa/ (accessed on 1 July 2022).
- McCune, B.; Grace, J.B. Analysis of Ecological Communities; Wild Blueberry Media LLC: Corvallis, OR, USA, 2002. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Roeland, K.; Pierre, L.; Dan, M.; Minchin, P.R.; O’Hara, R.; Simpson, G.S.; Solymos, P.; et al. Package ‘vegan’; 2022. Available online: https://cran.ism.ac.jp/web/packages/vegan/vegan.pdf (accessed on 1 November 2022).
- Legendre, P.; Gallagher, E.D. Ecologically meaningful transformations for ordination of species data. Oecologia 2001, 129, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Zelený, D.; Schaffers, A.P. Too good to be true: Pitfalls of using mean Ellenberg indicator values in vegetation analyses. J. Veg. Sci. 2012, 23, 419–431. [Google Scholar] [CrossRef]
- Wei, T.; Simko, V.; Levy, M.; Xie, Y.; Jin, Y.; Zemla, J. Package ‘corrplot’. Statistician 2017, 56, e24. [Google Scholar]
- de Cáceres, M.; Legendre, P. Associations between species and groups of sites: Indices and statistical inference. Ecology 2009, 90, 3566–3574. [Google Scholar] [CrossRef] [PubMed]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Slowikowski, K.; Schep, A.; Hughes, S.; Dang, T.K.; Lukauskas, S.; Irisson, J.-O.; Kamvar, Z.N.; Ryan, T.; Christophe, D.; Hiroaki, Y.; et al. Package Ggrepel. Automatically Position Non-Overlapping Text Labels with ’ggplot2’. 2018. Available online: https://github.com/slowkow/ggrepel (accessed on 1 July 2022).
- Wesolowski, M. Type composition and number of weed seeds in the soils of South-East Poland. Ann. Univ. Mariae Curie Sect. E 1979, 34, 23–47. [Google Scholar]
- Andreasen, C.; Jensen, H.A.; Jensen, S.M. Decreasing diversity in the soil seed bank after 50 years in Danish arable fields. Agric. Ecosyst. Environ. 2018, 259, 61–71. [Google Scholar] [CrossRef]
- Kropàč, Z. Estimation of weed seeds in arable soil. Pedobiologia 1966, 6, 106–130. [Google Scholar] [CrossRef]
- Berbeć, A.K.; Feledyn-Szewczyk, B. Biodiversity of weeds and soil seed bank in organic and conventional farming systems. Res. Rural Dev. 2018, 2, 12–19. [Google Scholar]
- Albrecht, H. Untersuchungen zur Veränderung der Segetalflora an sieben bayerischen Ackerstandorten zwischen den Erhebungszeiträumen 1951/68 und 1986/88. In Mit 31 Tabellen: Dissert Bot 141, 1-201. Cramer in D. Borntraeger-Verl.-Buchh. Berlin; Schweizerbart Science Publishers: Stuttgart, Germany, 1989. [Google Scholar]
- Metcalfe, H.; Hassall, K.L.; Boinot, S.; Storkey, J. The contribution of spatial mass effects to plant diversity in arable fields. J. Appl. Ecol. 2019, 56, 1560–1574. [Google Scholar] [CrossRef] [PubMed]
- O’Donovan, J.T.; McAndrew, D.W. Effect of tillage on weed populations in continuous barley (Hordeum vulgare). Weed Technol. 2000, 14, 726–733. [Google Scholar] [CrossRef]
- Swanton, C.J.; Clements, D.R.; Derksen, D.A. Weed succession under conservation tillage: A hierarchical framework for research and management. Weed Technol. 1993, 7, 286–297. [Google Scholar] [CrossRef]
- Murphy, S.D.; Clements, D.R.; Belaoussoff, S.; Kevan, P.G.; Swanton, C.J. Promotion of weed species diversity and reduction of weed seedbanks with conservation tillage and crop rotation. Weed Sci. 2006, 54, 69–77. [Google Scholar] [CrossRef]
- Albrecht, H. Langfristige Veränderung des Bodensamenvorrates bei pflugloser Bodenbearbeitung. J. Plant Dis. Prot. 2004, 19, 97–104. [Google Scholar]
- Shmida, A.V.; Wilson, M.V. Biological determinants of species diversity. J. Biogeogr. 1985, 12, 1–20. [Google Scholar] [CrossRef]
- Armengot, L.; Sans, F.X.; Fischer, C.; Flohre, A.; José-María, L.; Tscharntke, T.; Thies, C. The β-diversity of arable weed communities on organic and conventional cereal farms in two contrasting regions. Appl. Veg. Sci. 2012, 15, 571–579. [Google Scholar] [CrossRef]
- Roschewitz, I.; Gabriel, D.; Tscharntke, T.; Thies, C. The effects of landscape complexity on arable weed species diversity in organic and conventional farming. J. Appl. Ecol. 2005, 42, 873–882. [Google Scholar] [CrossRef]
- Matthies, D. Plasticity of reproductive components at different stages of development in the annual plant Thlaspi arvense L. Oecologia 1990, 83, 105–116. [Google Scholar] [CrossRef]
- Leguizamón, E.S.; Roberts, H.A. Seed production by an arable weed community. Weed Res. 1982, 22, 35–39. [Google Scholar] [CrossRef]
- Cavers, P.B.; Harper, J.L. Rumex obtusifolius L. and R. crispus L. J. Ecol. 1964, 52, 737–766. [Google Scholar] [CrossRef]
- Wiles, L.; Brodahl, M. Exploratory data analysis to identify factors influencing spatial distributions of weed seed banks. Weed Sci. 2004, 52, 936–947. [Google Scholar] [CrossRef]
- Stroot, L.; Brinkert, A.; Hölzel, N.; Rüsing, A.; Bucharova, A. Establishment of wildflower strips in a wide range of environments: A lesson from a landscape-scale project. Restor. Ecol. 2022, 30, e13542. [Google Scholar] [CrossRef]
- Bakker, J.P.; Berendse, F. Constraints in the restoration of ecological diversity in grassland and heathland communities. Trends Ecol. Evol. 1999, 14, 63–68. [Google Scholar] [CrossRef]
- Korneck, D.; Schnittler, M.; Vollmer, I. Rote Liste der Farn- und Blütenpflanzen (Pteridophyta et Spermatophyta) Deutschlands; German Red Data Book of Plants; Bundesamt für Naturschutz: Bonn, Germany, 1998. [Google Scholar]
- Thompson, K.; Bekker, R.M.; Bakker, J.P. Weed seed banks; evidence from the north-west European seed bank database. Asp. Appl. Biol. 1998, 51, 105–112. [Google Scholar]
- Albrecht, H. Development of arable weed seedbanks during the 6 years after the change from conventional to organic farming. Weed Res. 2005, 45, 339–350. [Google Scholar] [CrossRef]
- Uyttenbroeck, R.; Hatt, S.; Piqueray, J.; Paul, A.; Bodson, B.; Francis, F.; Monty, A. Creating perennial flower strips: Think functional! Agric. Agric. Sci. Procedia 2015, 6, 95–101. [Google Scholar] [CrossRef]
- Shaukat, S.S.; Siddiqui, I.A. Spatial pattern analysis of seeds of an arable soil seed bank and its relationship with above-ground vegetation in an arid region. J. Arid Environ. 2004, 57, 311–327. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sum Sq | Num DF | Den DF | F | p | |
|---|---|---|---|---|---|
| Number of species | |||||
| Depth | 0.233 | 1 | 45 | 2.884 | 0.096 |
| Field position | 6.321 | 1 | 45 | 78.215 | <0.001 |
| Tillage | 0.621 | 1 | 15 | 7.682 | 0.014 |
| Depth × field position | 0.0146 | 1 | 45 | 0.203 | 0.654 |
| Depth × tillage | 0.215 | 1 | 45 | 2.661 | 0.100 |
| Field position × tillage | 0.768 | 1 | 45 | 0.91 | 0.004 |
| Depth × field position × tillage | 0.008 | 1 | 1 | 0.101 | 0.752 |
| Number of seeds per m2 | |||||
| Depth | 2.624 | 1 | 45 | 1.9 | 0.0711 |
| Field position | 13.236 | 1 | 45 | 10.2 | <0.001 |
| Tillage | 3.651 | 1 | 15 | 4.3 | 0.046 |
| Depth × field position | 0.4122 | 1 | 45 | 0.5367 | 0.468 |
| Depth × tillage | 0.0014 | 1 | 45 | 0.0018 | 0.967 |
| Field position × tillage | 0.3966 | 1 | 45 | 0.5164 | 0.476 |
| Depth × field position × tillage | 0.0856 | 1 | 1 | 0.1115 | 0.740 |
| Sum Sq | Num DF | Den DF | F-Value | p | |
|---|---|---|---|---|---|
| Number of species | |||||
| Field location (total) | 8.1 | 2 | 36 | 34.9 | <0.0001 |
| Field location (without fls) | 4.5 | 2 | 36 | 18.2 | <0.0001 |
| Number of seeds | |||||
| Field location (total) | 32.6 | 2 | 36 | 24.9 | <0.0001 |
| Field location (without fls) | 30.7 | 2 | 36 | 23.5 | <0.0001 |
| Sørensen Similarity Index | Both | Seed Bank Only | Vegetation Only | |
|---|---|---|---|---|
| Mean ± SE | Mean ± SE | Mean ± SE | Mean ± SE | |
| Field interior | 0.17 ± 0.04 | 1.05 ± 0.44 | 6.40 ± 0.90 | 2.25 ± 0.67 |
| Field edge | 0.20 ± 0.02 | 2.40 ± 0.34 | 11.50 ± 0.89 | 5.90 ± 0.84 |
| Flower strip | 0.33 ± 0.02 | 8.10 ± 0.84 | 13.55 ± 1.36 | 17.85 ± 1.58 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schnee, L.; Sutcliffe, L.M.E.; Leuschner, C.; Donath, T.W. Weed Seed Banks in Intensive Farmland and the Influence of Tillage, Field Position, and Sown Flower Strips. Land 2023, 12, 926. https://doi.org/10.3390/land12040926
Schnee L, Sutcliffe LME, Leuschner C, Donath TW. Weed Seed Banks in Intensive Farmland and the Influence of Tillage, Field Position, and Sown Flower Strips. Land. 2023; 12(4):926. https://doi.org/10.3390/land12040926
Chicago/Turabian StyleSchnee, Liesa, Laura M. E. Sutcliffe, Christoph Leuschner, and Tobias W. Donath. 2023. "Weed Seed Banks in Intensive Farmland and the Influence of Tillage, Field Position, and Sown Flower Strips" Land 12, no. 4: 926. https://doi.org/10.3390/land12040926