
The Spreading of Antibiotic-Resistant Bacteria in Terrestrial Ecosystems and the Formation of Soil Resistome

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil Sample Collection
2.2. Description of Localities
2.3. Microbial Assays
2.4. HPLC Analysis of Enrofloxacin
2.5. Statistics
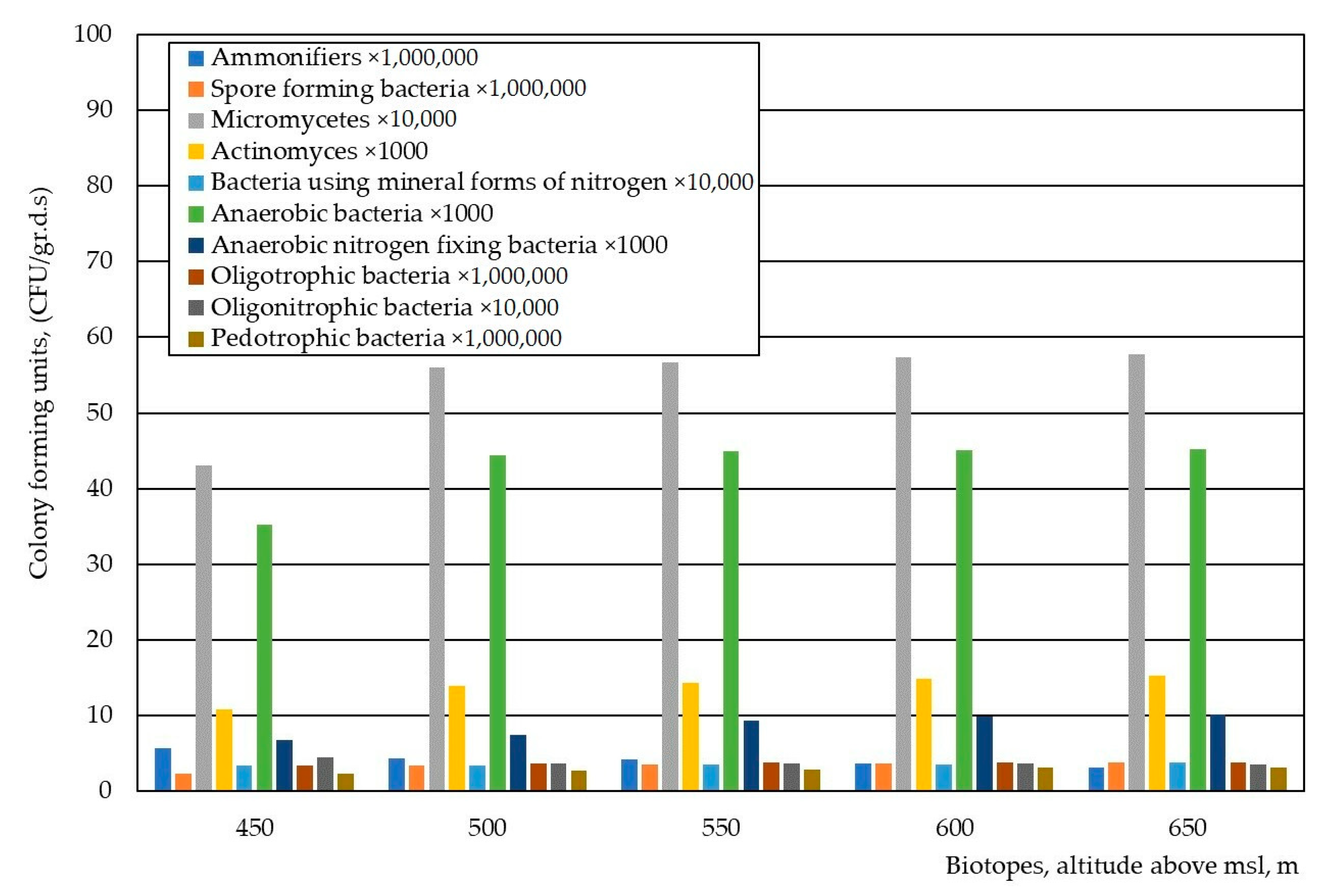
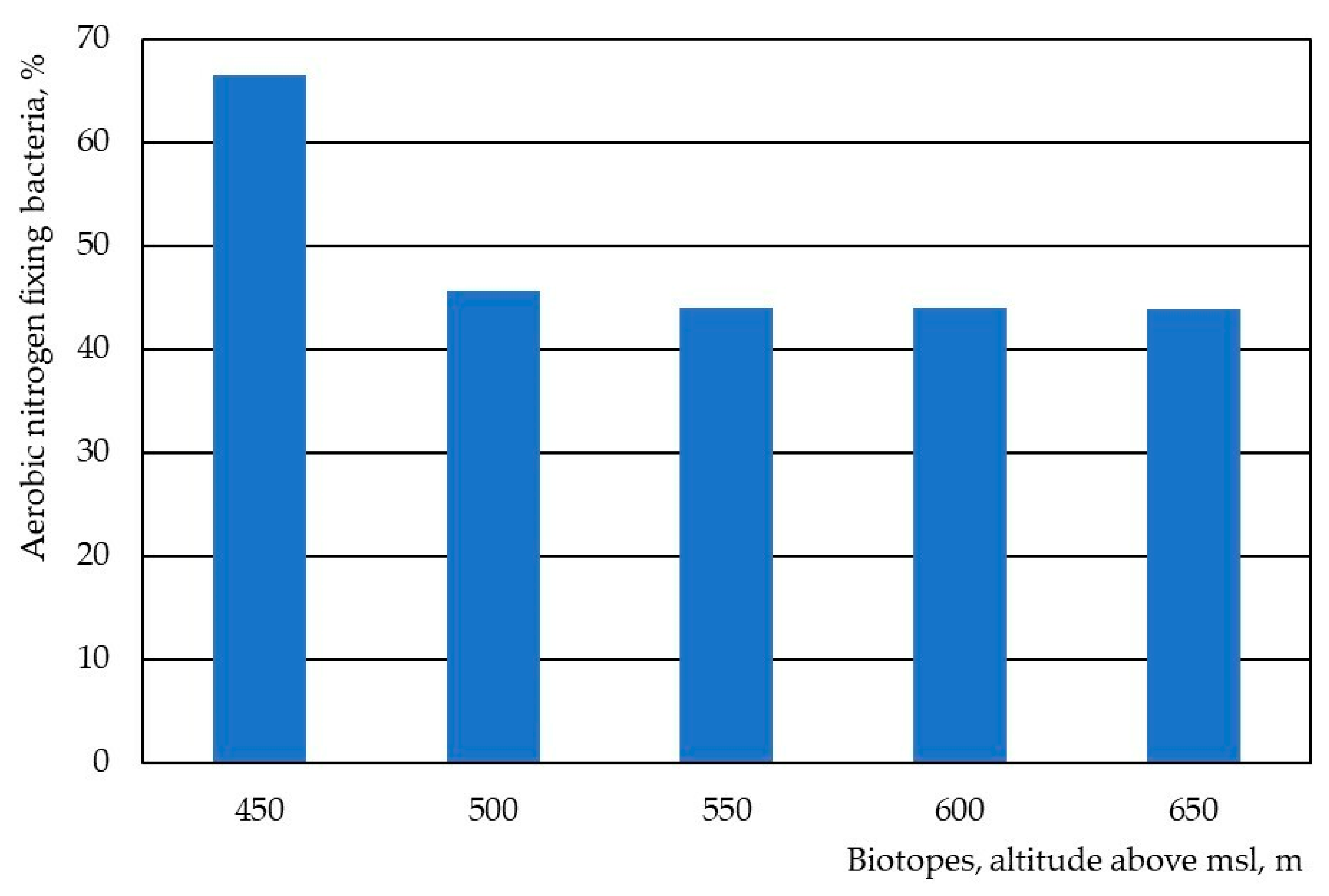
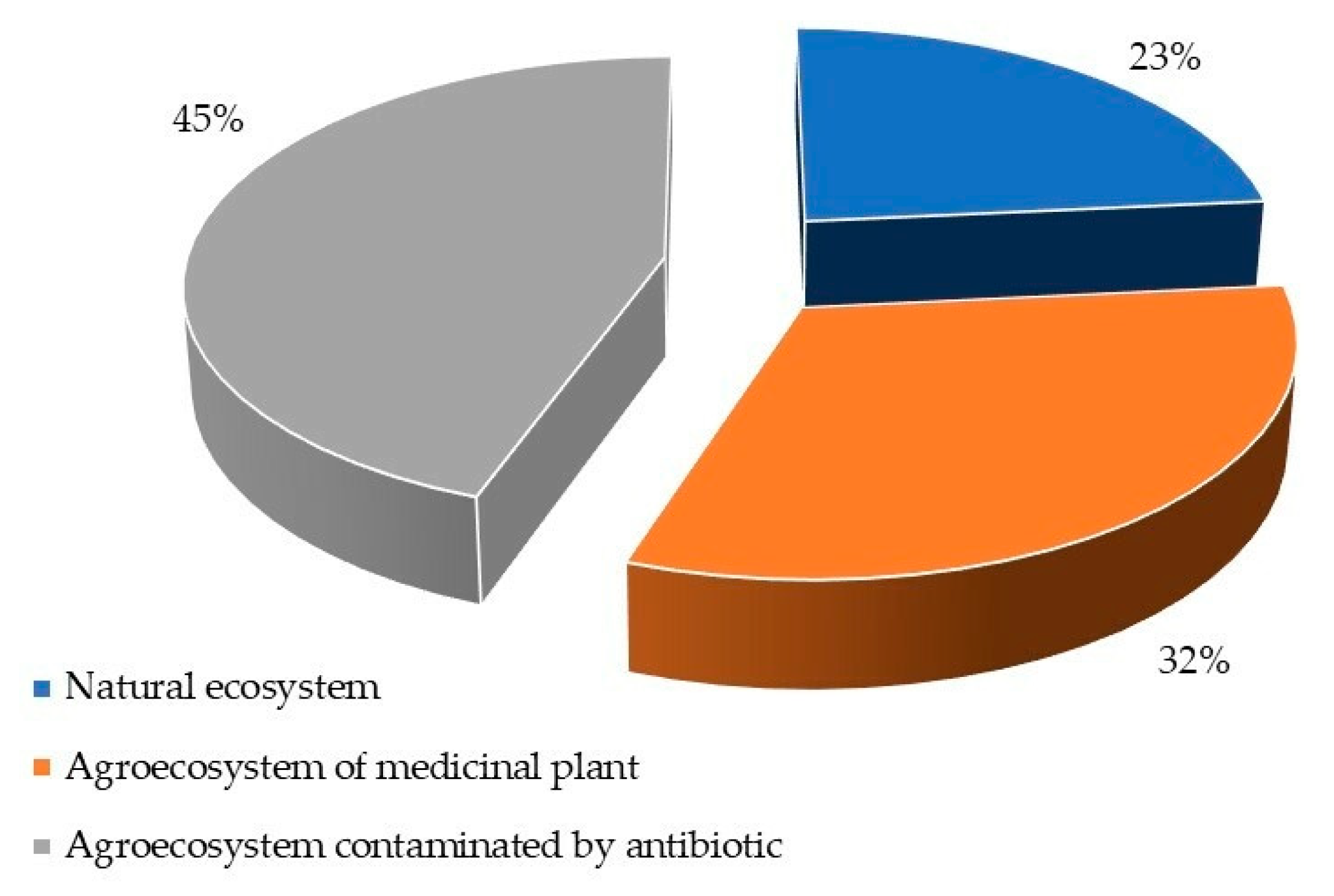
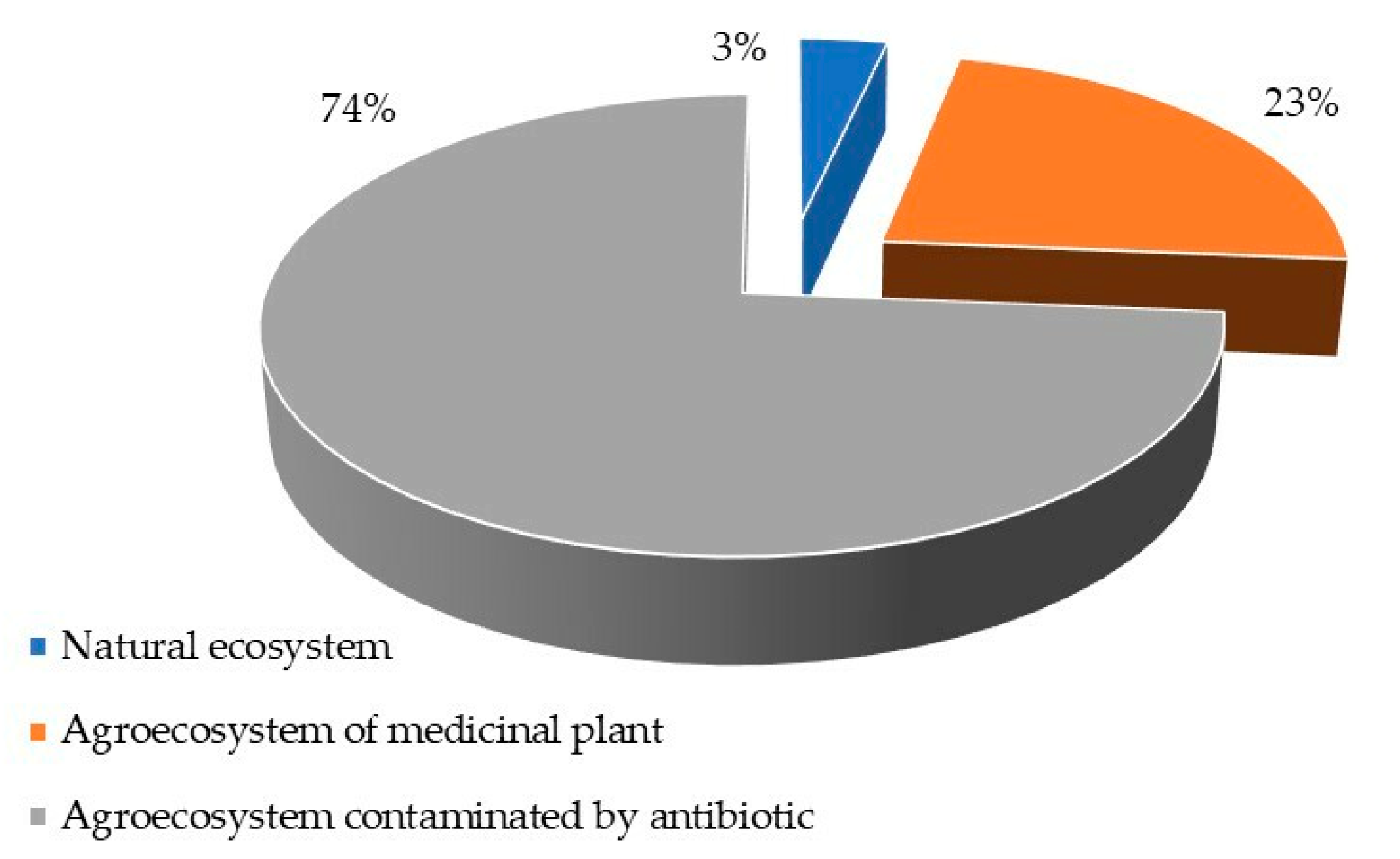
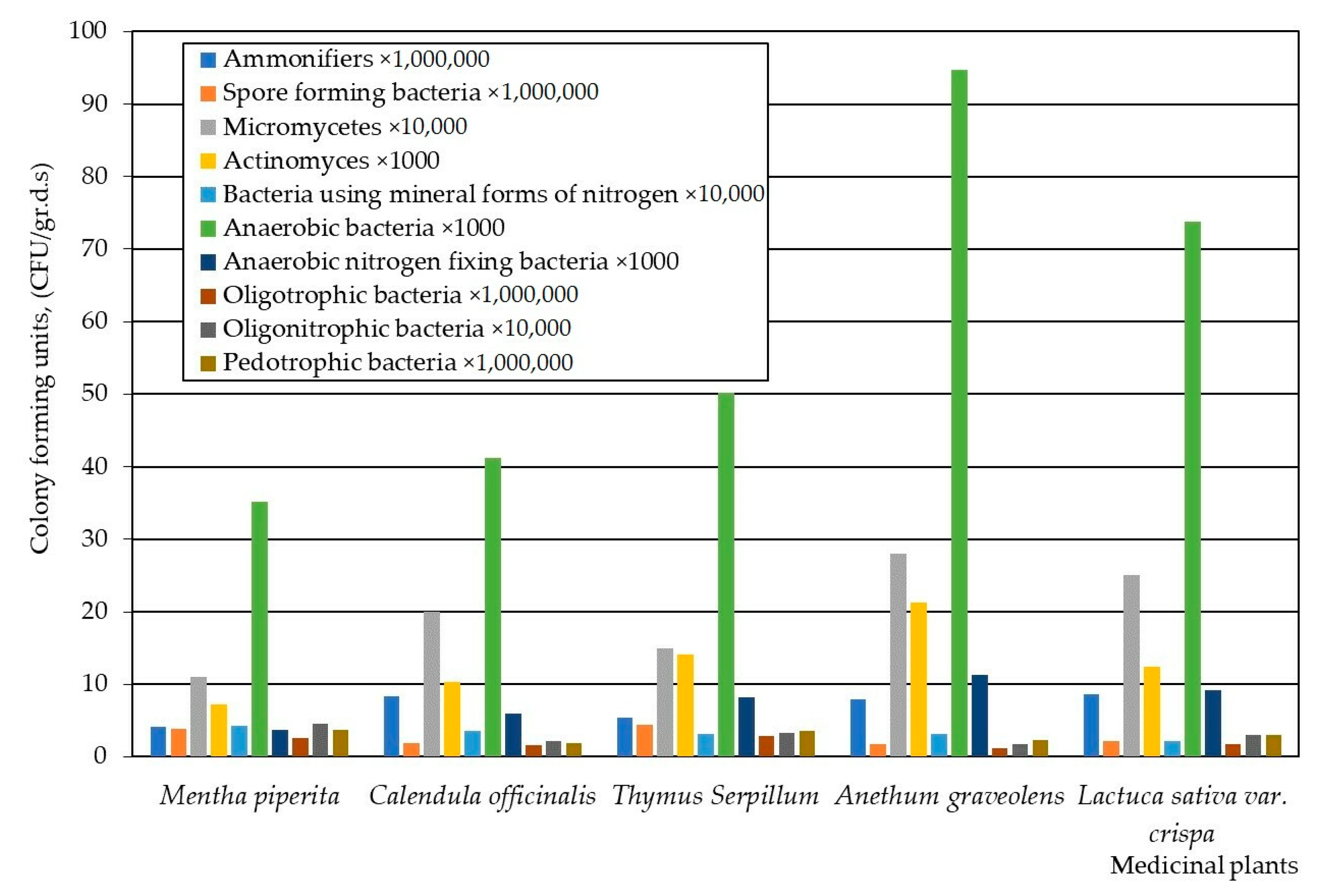
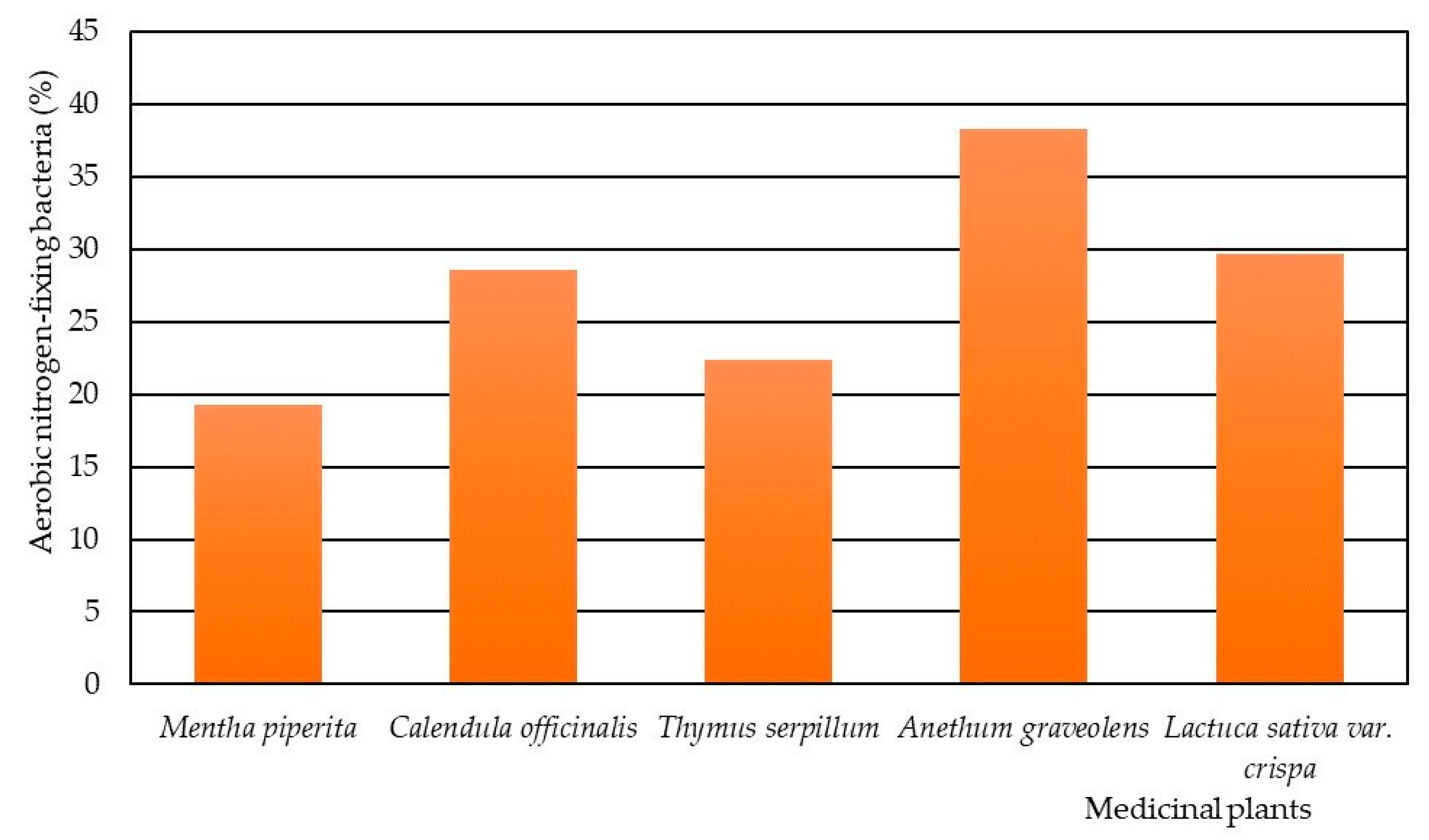
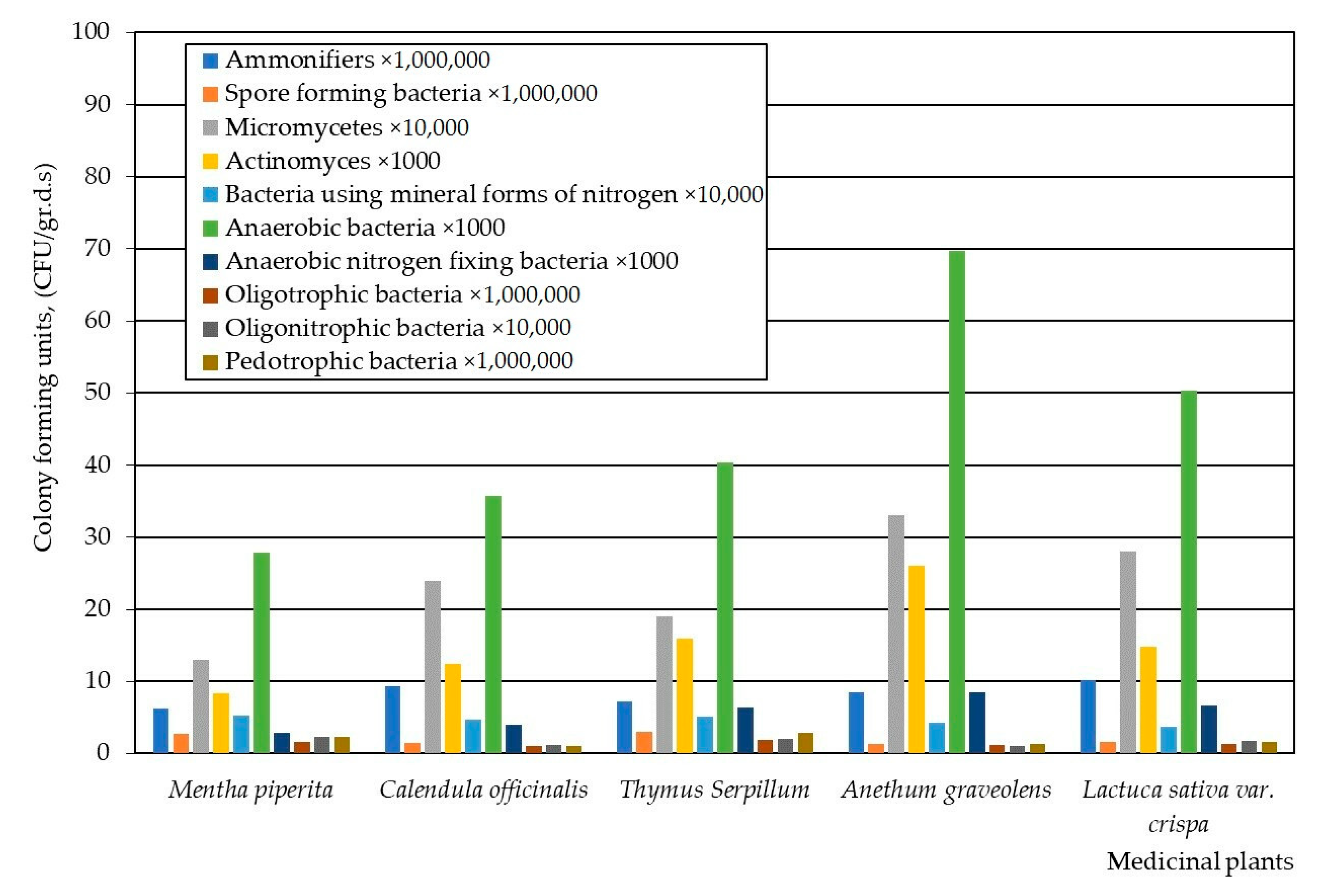
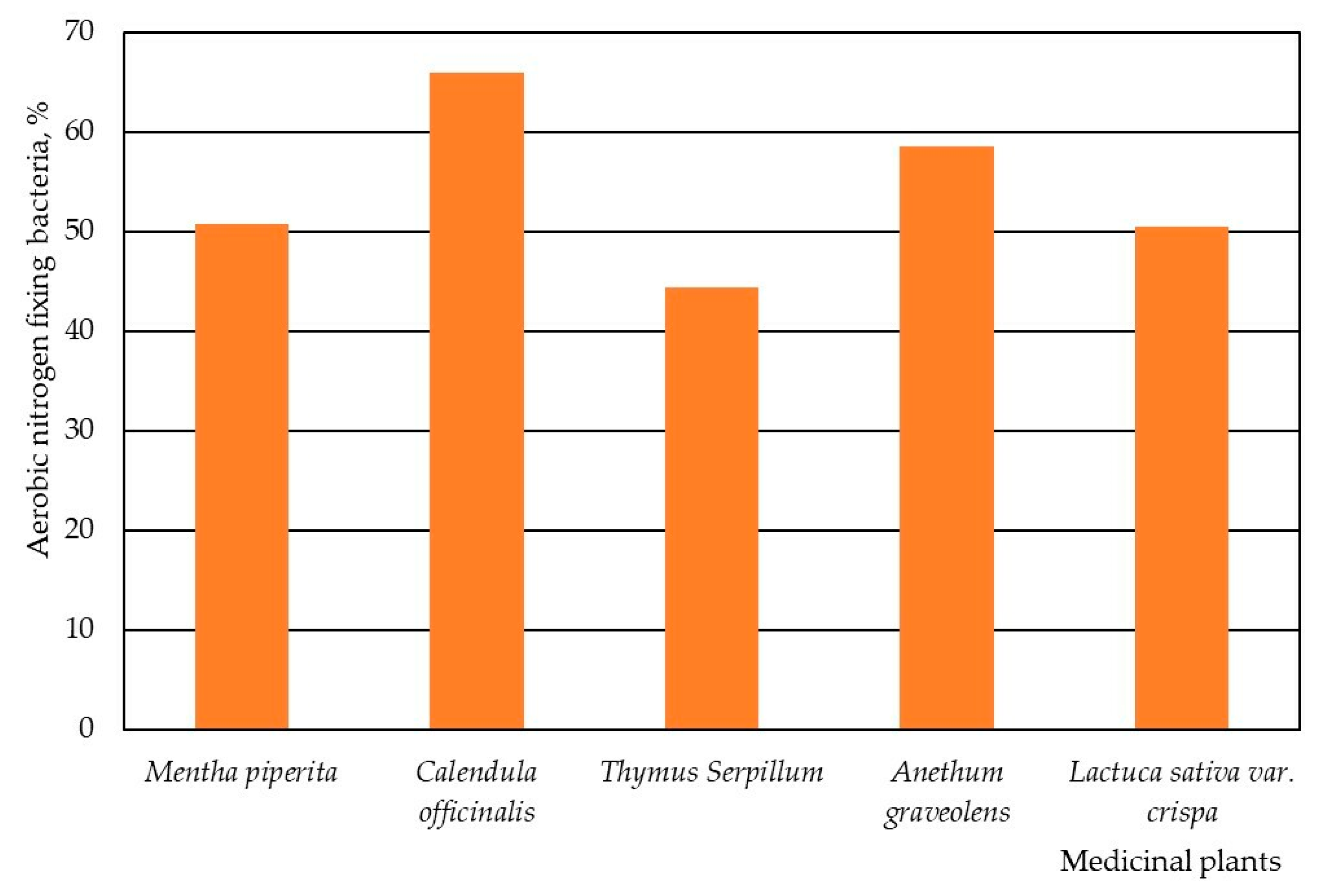
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bungau, S.; Suciu, R.; Bumbu, A.; Cioca, G.; Tit, D.M. Study on hospital waste management in medical rehabilitation clinical hospital, Baile Felix. J. Environ. Prot. Ecol. 2015, 16, 980–987. [Google Scholar]
- Bungau, S.; Tit, D.; Fodor, K.; Cioca, G.; Agop, M.; Iovan, C.; Cseppento, D.; Bumbu, A.; Bustea, C. Aspects Regarding the Pharmaceutical Waste Management in Romania. Sustainability 2018, 10, 2788. [Google Scholar] [CrossRef] [Green Version]
- Symochko, L. Microbiological Control of Soil-Borne Antibiotic Resistance Human Pathogens in Agroecosystems. Int. J. Ecosyst. Ecol. Sci. IJEES 2018, 8, 591–598. [Google Scholar] [CrossRef]
- Xiao, R.; Huang, D.; Du, L.; Song, B.; Yin, L.; Chen, Y.; Gao, L.; Li, R.; Huang, H.; Zeng, G. Antibiotic resistance in soil-plant systems: A review of the source, dissemination, influence factors, and potential exposure risks. Sci. Total Environ. 2023, 869, 161855. [Google Scholar] [CrossRef] [PubMed]
- Wright, G.D. The Antibiotic Resistome. Expert Opin. Drug Discov. 2010, 5, 779–788. [Google Scholar] [CrossRef]
- Symochko, L. Soil Microbiome: Diversity, Activity, Functional and Structural Successions. Int. J. Ecosyst. Ecol. Sci. IJEES 2020, 10, 277–284. [Google Scholar] [CrossRef]
- Patyka, V.P.; Symochko, L. Soil microbiological monitoring of natural and transformed ecosystems in the Trans-Carpathian region of Ukraine. Mikrobiolohichnyi Zhurnal 2013, 75, 21–31. [Google Scholar]
- Allen, H.K.; Donato, J.; Wang, H.H.; Cloud-Hansen, K.A.; Davies, J.; Handelsman, J. Call of the Wild: Antibiotic Resistance Genes in Natural Environments. Nat. Rev. Microbiol. 2010, 8, 251–259. [Google Scholar] [CrossRef]
- Martínez, J.L. Antibiotics and Antibiotic Resistance Genes in Natural Environments. Science 2008, 321, 365–367. [Google Scholar] [CrossRef]
- Schmitt, H.; van Beelen, P.; Tolls, J.; van Leeuwen, C.L. Pollution-Induced Community Tolerance of Soil Microbial Communities Caused by the Antibiotic Sulfachloropyridazine. Environ. Sci. Technol. 2004, 38, 1148–1153. [Google Scholar] [CrossRef]
- Boxall, A.B.A.; Johnson, P.; Smith, E.J.; Sinclair, C.J.; Stutt, E.; Levy, L.S. Uptake of Veterinary Medicines from Soils into Plants. J. Agric. Food Chem. 2006, 54, 2288–2297. [Google Scholar] [CrossRef] [PubMed]
- Christian, T.; Schneider, R.J.; Färber, H.A.; Skutlarek, D.; Meyer, M.T.; Goldbach, H.E. Determination of Antibiotic Residues in Manure, Soil, and Surface Waters. Acta Hydrochim. Hydrobiol. 2003, 31, 36–44. [Google Scholar] [CrossRef]
- Xiaoping, D.; Yingjian, S.; Zhenjun, S.; Jianzhong, S. Effects of Apramycin on microbial activity in different types of soil. Ecol. Environ. 2004, 13, 565–568. [Google Scholar]
- Díaz-Cruz, M.S.; López de Alda, M.J.; Barceló, D. Environmental Behavior and Analysis of Veterinary and Human Drugs in Soils, Sediments and Sludge. TrAC Trends Anal. Chem. 2003, 22, 340–351. [Google Scholar] [CrossRef]
- Wright, G.D. The Antibiotic Resistome: The Nexus of Chemical and Genetic Diversity. Nat. Rev. Microbiol. 2007, 5, 175–186. [Google Scholar] [CrossRef]
- Chee-Sanford, J.C.; Aminov, R.I.; Krapac, I.J.; Garrigues-Jeanjean, N.; Mackie, R.I. Occurrence and Diversity of Tetracycline Resistance Genes in Lagoons and Groundwater Underlying Two Swine Production Facilities. Appl. Environ. Microbiol. 2001, 67, 1494–1502. [Google Scholar] [CrossRef] [Green Version]
- Aminov, R.I.; Mackie, R.I. Evolution and Ecology of Antibiotic Resistance Genes. FEMS Microbiol. Lett. 2007, 271, 147–161. [Google Scholar] [CrossRef]
- Sanchez-Cid, C.; Guironnet, A.; Wiest, L.; Vulliet, E.; Vogel, T.M. Gentamicin Adsorption onto Soil Particles Prevents Overall Short-Term Effects on the Soil Microbiome and Resistome. Antibiotics 2021, 10, 191. [Google Scholar] [CrossRef]
- Meng, M.; Li, Y.; Yao, H. Plasmid-Mediated Transfer of Antibiotic Resistance Genes in Soil. Antibiotics 2022, 11, 525. [Google Scholar] [CrossRef]
- Symochko, L.; Patyka, V.; Symochko, V.; Kalinichenko, A. Soil Microbial Activity and Functional Diversity in Primeval Beech Forests. J. Earth Sci. Eng. 2015, 5, 363–371. [Google Scholar] [CrossRef] [Green Version]
- Cavigelli, M.A.; Robertson, G.P. The Functional Significance of Denitrifier Community Composition in a Terrestrial Ecosystem. Ecology 2000, 81, 1402–1414. [Google Scholar] [CrossRef]
- Ebert, I.; Bachmann, J.; Kühnen, U.; Küster, A.; Kussatz, C.; Maletzki, D.; Schlüter, C. Toxicity of the Fluoroquinolone Antibiotics Enrofloxacin and Ciprofloxacin to Photoautotrophic Aquatic Organisms. Environ. Toxicol. Chem. 2011, 30, 2786–2792. [Google Scholar] [CrossRef] [PubMed]
- Forsberg, K.J.; Reyes, A.; Wang, B.; Selleck, E.M.; Sommer, M.O.A.; Dantas, G. The Shared Antibiotic Resistome of Soil Bacteria and Human Pathogens. Science 2012, 337, 1107–1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammesfahr, U.; Heuer, H.; Manzke, B.; Smalla, K.; Thiele-Bruhn, S. Impact of the Antibiotic Sulfadiazine and Pig Manure on the Microbial Community Structure in Agricultural Soils. Soil Biol. Biochem. 2008, 40, 1583–1591. [Google Scholar] [CrossRef]
- Heuer, H.; Schmitt, H.; Smalla, K. Antibiotic Resistance Gene Spread Due to Manure Application on Agricultural Fields. Curr. Opin. Microbiol. 2011, 14, 236–243. [Google Scholar] [CrossRef]
- Kummerer, K. Resistance in the Environment. J. Antimicrob. Chemother. 2004, 54, 311–320. [Google Scholar] [CrossRef] [Green Version]
- Goldman, E.; Green, L.H. (Eds.) Practical Handbook of Microbiology, 3rd ed.; CRC Press: Boca Raton, FL, USA, 2015; ISBN 978-0-429-16893-2. [Google Scholar]
- Shyrobokov, V.P. Medical Microbiology, Virology, and Immunology; Nova Knyha: Vinnytsia, Ukraine, 2011; ISBN 978-966-382-325-6. [Google Scholar]
- Symochko, L.; Hamuda, H.B. Microbial Monitoring of Soil as Additional Tools for Conservation Biology. Obuda Univ. E-Bull. 2015, 5, 177–185. [Google Scholar]
- Heuer, H.; Smalla, K. Manure and Sulfadiazine Synergistically Increased Bacterial Antibiotic Resistance in Soil over at Least Two Months. Environ. Microbiol. 2007, 9, 657–666. [Google Scholar] [CrossRef]
- Symochko, L. Food Safety in Agroecosystems-Soil Resistome. In Proceedings of the 13th Paris International Conference (PACBES-19), Paris, France, 14–16 March 2019; pp. 99–102. [Google Scholar]
- Mikola, J.; Setälä, H. No Evidence of Trophic Cascades in an Experimental Microbial Based Soil Food Web. Ecology 1998, 79, 153–164. [Google Scholar] [CrossRef]
- Symochko, L.; Hoxha, E.; Bayoumi Hamuda, H. Mapping Hot Spots of Soil Microbiome Using GIS Technology. J. Agric. For. 2021, 67, 191–203. [Google Scholar] [CrossRef]
- Aljeldah, M.M. Antimicrobial Resistance and Its Spread Is a Global Threat. Antibiotics 2022, 11, 1082. [Google Scholar] [CrossRef] [PubMed]
- Mackenzie, J.S.; Jeggo, M. The One Health Approach—Why Is It So Important? Trop. Med. Infect. Dis. 2019, 4, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Neill, J. Tackling Drug-Resistant Infections Globally: Final Report and Recommendations; Government of the United Kingdom: London, UK, 2016; Available online: https://amr-review.org/sites/default/files/160518_Final%20paper_with%20cover.pdf (accessed on 19 May 2016).
- Serwecińska, L. Antimicrobials and Antibiotic-Resistant Bacteria: A Risk to the Environment and to Public Health. Water 2020, 12, 3313. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Hu, H.-W.; Maestre, F.T.; Guerra, C.A.; Eisenhauer, N.; Eldridge, D.J.; Zhu, Y.-G.; Chen, Q.-L.; Trivedi, P.; Du, S.; et al. The global distribution and environmental drivers of the soil antibiotic resistome. Microbiome 2022, 10, 219. [Google Scholar] [CrossRef] [PubMed]
- Flandroy, L.; Poutahidis, T.; Berg, G.; Clarke, G.; Dao, M.-C.; Decaestecker, E.; Furman, E.; Haahtela, T.; Massart, S.; Plovier, H.; et al. The impact of human activities and lifestyles on the interlinked microbiota and health of humans and of ecosystems. Sci. Total Environ. 2018, 627, 1018–1038. [Google Scholar] [CrossRef]
- Symochko, L. Assessment of Sorption and Toxicity of Fluoroquinolone Antibiotic in Agroecosystems. Agroecol. J. 2017, 3, 147–151. [Google Scholar] [CrossRef]
- Lladó, S.; López-Mondéjar, R.; Baldrian, P. Forest Soil Bacteria: Diversity, Involvement in Ecosystem Processes, and Response to Global Change. Microbiol. Mol. Biol. Rev. 2017, 81, e00063-16. [Google Scholar] [CrossRef] [Green Version]
- Symochko, L.; Mariychuk, R.; Demyanyuk, O.; Symochko, V. Enrofloxacin in Agroecosystems: Uptake by Plants and Phytotoxical Effect. In Proceedings of the 2019 International Council on Technologies of Environmental Protection (ICTEP), Starý Smokovec, Slovakia, 23–25 October 2019; pp. 250–253. [Google Scholar]
- Torsvik, V.; Øvreås, L. Microbial Diversity and Function in Soil: From Genes to Ecosystems. Curr. Opin. Microbiol. 2002, 5, 240–245. [Google Scholar] [CrossRef]
- van der Heijden, M.G.A.; Bardgett, R.D.; van Straalen, N.M. The Unseen Majority: Soil Microbes as Drivers of Plant Diversity and Productivity in Terrestrial Ecosystems. Ecol. Lett. 2008, 11, 296–310. [Google Scholar] [CrossRef]
- Demyanyuk, O.; Symochko, L.; Shatsman, D. Structure and Dynamics of Soil Microbial Communities of Natural and Transformed Ecosystems. Environ. Res. Eng. Manag. 2020, 76, 97–105. [Google Scholar] [CrossRef]
- Laconi, A.; Mughini-Gras, L.; Tolosi, R.; Grilli, G.; Trocino, A.; Carraro, L.; Di Cesare, F.; Cagnardi, P.; Piccirillo, A. Microbial Community Composition and Antimicrobial Resistance in Agricultural Soils Fertilized with Livestock Manure from Conventional Farming in Northern Italy. Sci. Total Environ. 2021, 760, 143404. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Snow, D.; Walia, H.; Li, X. Transmission Routes of the Microbiome and Resistome from Manure to Soil and Lettuce. Environ. Sci. Technol. 2021, 55, 11102–11112. [Google Scholar] [CrossRef]
- Subirats, J.; Murray, R.; Yin, X.; Zhang, T.; Topp, E. Impact of Chicken Litter Pre-Application Treatment on the Abundance, Field Persistence, and Transfer of Antibiotic Resistant Bacteria and Antibiotic Resistance Genes to Vegetables. Sci. Total Environ. 2021, 801, 149718. [Google Scholar] [CrossRef] [PubMed]
- Rasschaert, G.; Van Elst, D.; Colson, L.; Herman, L.; de Carvalho Ferreira, H.C.; Dewulf, J.; Decrop, J.; Meirlaen, J.; Heyndrickx, M.; Daeseleire, E. Antibiotic Residues and Antibiotic-Resistant Bacteria in Pig Slurry Used to Fertilize Agricultural Fields. Antibiotics 2020, 9, 34. [Google Scholar] [CrossRef] [Green Version]
- Stanton, I.C.; Tipper, H.J.; Chau, K.; Klümper, U.; Subirats, J.; Murray, A.K. Does Environmental Exposure to Pharmaceutical and Personal Care Product Residues Result in the Selection of Antimicrobial-Resistant Microorganisms, and Is This Important in Terms of Human Health Outcomes? Environ. Toxicol. Chem. 2022. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microorganisms | Ammonifiers | Spore Forming Bacteria | Micromycetes | Actinomyces | Bacteria Using Mineral Forms of Nitrogen | Anaerobic Bacteria | Anaerobic Nitrogen Fixing Bacteria | Oligotrophic Bacteria | Oligonitrophic Bacteria | Pedotrophic Bacteria | Aerobic Nitrogen Fixing Bacteria |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Ammonifiers | 1 | ||||||||||
| Spore-forming bacteria | −0.96 | 1 | |||||||||
| Micromycetes | −0.92 | 0.99 | 1 | ||||||||
| Actinomyces | −0.97 | 1.00 | 0.98 | 1 | |||||||
| Bacteria using mineral forms of nitrogen | −0.85 | 0.71 | 0.61 | 0.73 | 1 | ||||||
| Anaerobic bacteria | −0.90 | 0.99 | 1.00 | 0.98 | 0.58 | 1 | |||||
| Anaerobic nitrogen-fixing bacteria | −0.90 | 0.85 | 0.79 | 0.87 | 0.82 | 0.77 | 1 | ||||
| Oligotrophic bacteria | −0.97 | 0.92 | 0.87 | 0.94 | 0.85 | 0.86 | 0.98 | 1 | |||
| Oligonitrophic bacteria | 0.95 | −1.00 | −1.00 | −1.00 | −0.67 | −0.99 | −0.82 | −0.90 | 1 | ||
| Pedotrophic bacteria | −0.98 | 0.97 | 0.94 | 0.98 | 0.79 | 0.92 | 0.93 | 0.98 | −0.96 | 1 | |
| Aerobic nitrogen-fixing bacteria | 0.89 | −0.98 | −1.00 | −0.97 | −0.57 | −1.00 | −0.77 | −0.85 | 0.99 | −0.92 | 1 |
| Microorganisms | Ammonifiers | Spore Forming Bacteria | Micromycetes | Actinomyces | Bacteria Using Mineral Forms of Nitrogen | Anaerobic Bacteria | Anaerobic Nitrogen Fixing Bacteria | Oligotrophic Bacteria | Oligonitrophic Bacteria | Pedotrophic Bacteria | Aerobic Nitrogen Fixing Bacteria |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Ammonifiers | 1 | ||||||||||
| Spore-forming bacteria | −0.88 | 1 | |||||||||
| Micromycetes | 0.88 | −0.74 | 1 | ||||||||
| Actinomyces | 0.44 | −0.40 | 0.75 | 1 | |||||||
| Bacteria using mineral forms of nitrogen | −0.70 | 0.38 | −0.72 | −0.46 | 1 | ||||||
| Anaerobic bacteria | 0.59 | −0.57 | 0.89 | 0.89 | −0.67 | 1 | |||||
| Anaerobic nitrogen-fixing bacteria | 0.61 | −0.45 | 0.85 | 0.93 | −0.76 | 0.94 | 1 | ||||
| Oligotrophic bacteria | −0.87 | 0.99 | −0.90 | −0.52 | 0.41 | −0.68 | −0.56 | 1 | |||
| Oligonitrophic bacteria | −0.83 | 0.79 | −0.85 | −0.75 | 0.45 | −0.66 | −0.73 | 0.82 | 1 | ||
| Pedotrophic bacteria | −0.79 | 0.82 | −0.55 | −0.15 | 0.16 | −0.14 | −0.17 | 0.75 | 0.71 | 1 | |
| Aerobic nitrogen-fixing bacteria | 0.80 | −0.85 | 0.96 | 0.82 | −0.51 | 0.88 | 0.82 | −0.92 | −0.90 | −0.58 | 1 |
| Microorganisms | Ammonifiers | Spore Forming Bacteria | Micromycetes | Actinomyces | Bacteria Using Mineral Forms of Nitrogen | Anaerobic Bacteria | Anaerobic Nitrogen Fixing Bacteria | Oligotrophic Bacteria × | Oligonitrophic Bacteria | Pedotrophic Bacteria | Aerobic Nitrogen Fixing Bacteria |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Ammonifiers | 1 | ||||||||||
| Spore-forming bacteria | −0.79 | 1 | |||||||||
| Micromycetes | 0.77 | −0.83 | 1 | ||||||||
| Actinomyces | 0.30 | −0.47 | 0.83 | 1 | |||||||
| Bacteria using mineral forms of nitrogen | −0.87 | 0.67 | −0.79 | −0.43 | 1 | ||||||
| Anaerobic bacteria | 0.46 | −0.79 | 0.91 | 0.96 | −0.64 | 1 | |||||
| Anaerobic nitrogen-fixing bacteria | 0.42 | −0.36 | 0.82 | 0.93 | −0.61 | 0.94 | 1 | ||||
| Oligotrophic bacteria | −0.72 | 0.98 | −0.78 | −0.38 | 0.51 | −0.47 | −0.22 | 1 | |||
| Oligonitrophic bacteria | −0.64 | 0.88 | −0.82 | −0.64 | 0.42 | −0.63 | −0.46 | 0.91 | 1 | ||
| Pedotrophic bacteria | −0.75 | 0.98 | −0.81 | −0.31 | 0.56 | −0.42 | −0.17 | 0.99 | 0.85 | 1 | |
| Aerobic nitrogen-fixing bacteria | 0.45 | −0.80 | 0.43 | 0.15 | −0.13 | 0.16 | −0.11 | −0.91 | −0.83 | −0.89 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Symochko, L.; Demyanyuk, O.; Symochko, V.; Grulova, D.; Fejer, J.; Mariychuk, R. The Spreading of Antibiotic-Resistant Bacteria in Terrestrial Ecosystems and the Formation of Soil Resistome. Land 2023, 12, 769. https://doi.org/10.3390/land12040769
Symochko L, Demyanyuk O, Symochko V, Grulova D, Fejer J, Mariychuk R. The Spreading of Antibiotic-Resistant Bacteria in Terrestrial Ecosystems and the Formation of Soil Resistome. Land. 2023; 12(4):769. https://doi.org/10.3390/land12040769
Chicago/Turabian StyleSymochko, Lyudmyla, Olena Demyanyuk, Vitaliy Symochko, Daniela Grulova, Jozef Fejer, and Ruslan Mariychuk. 2023. "The Spreading of Antibiotic-Resistant Bacteria in Terrestrial Ecosystems and the Formation of Soil Resistome" Land 12, no. 4: 769. https://doi.org/10.3390/land12040769