
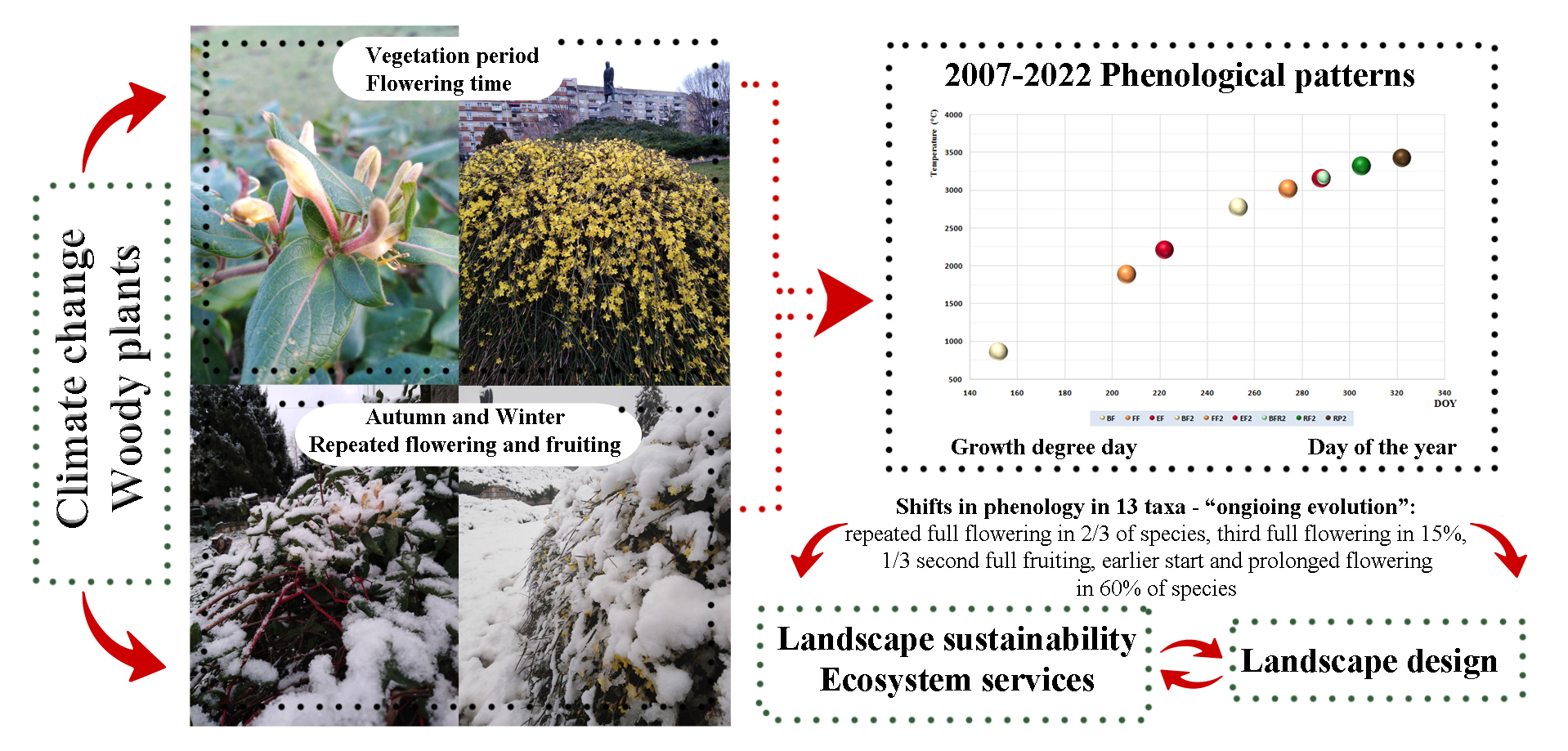
Phenological Flowering Patterns of Woody Plants in the Function of Landscape Design: Case Study Belgrade

Abstract
:
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Data Acquisition
2.3. Data Processing
3. Results
3.1. Chronology of Climate Data
Climate in Serbia in 2022
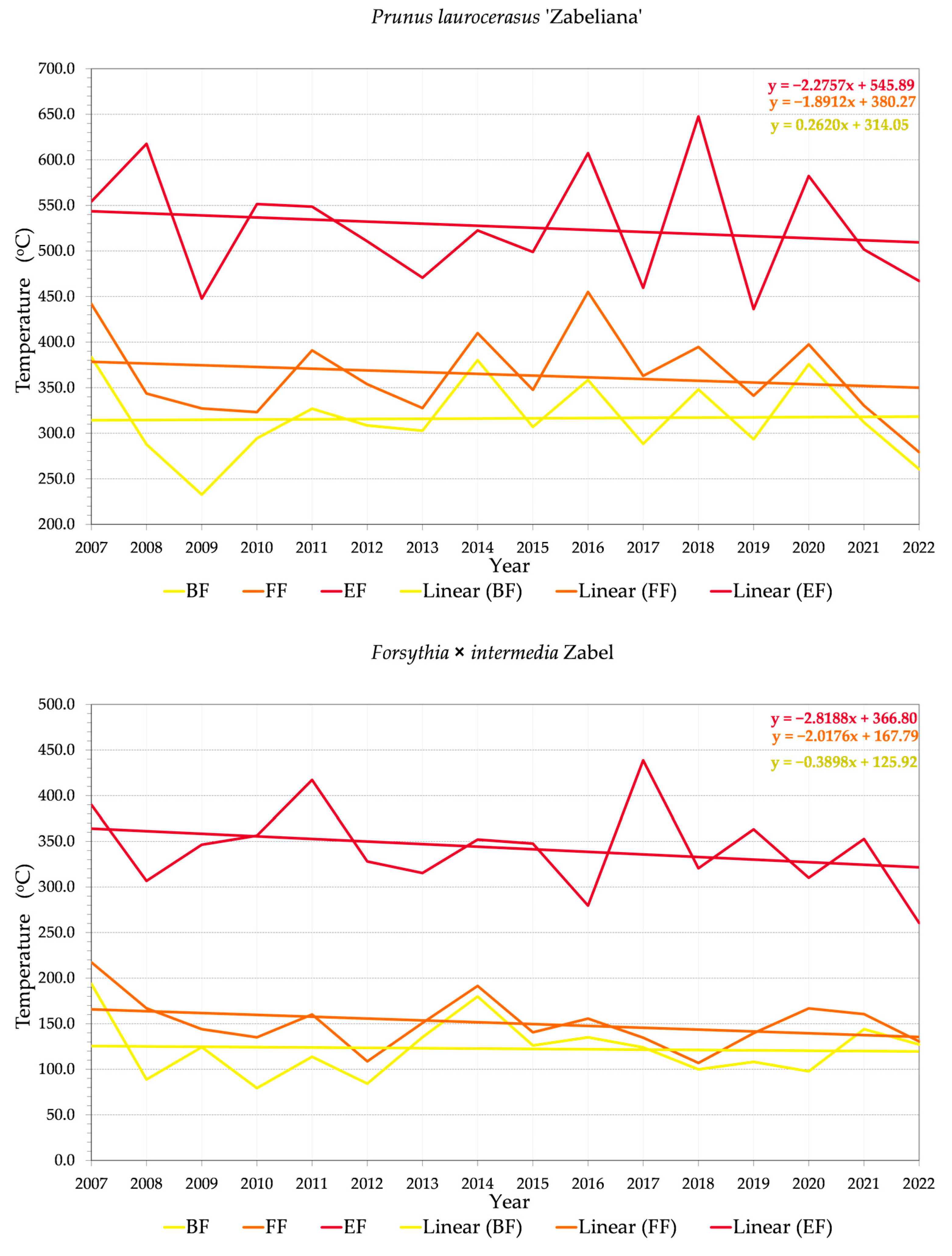
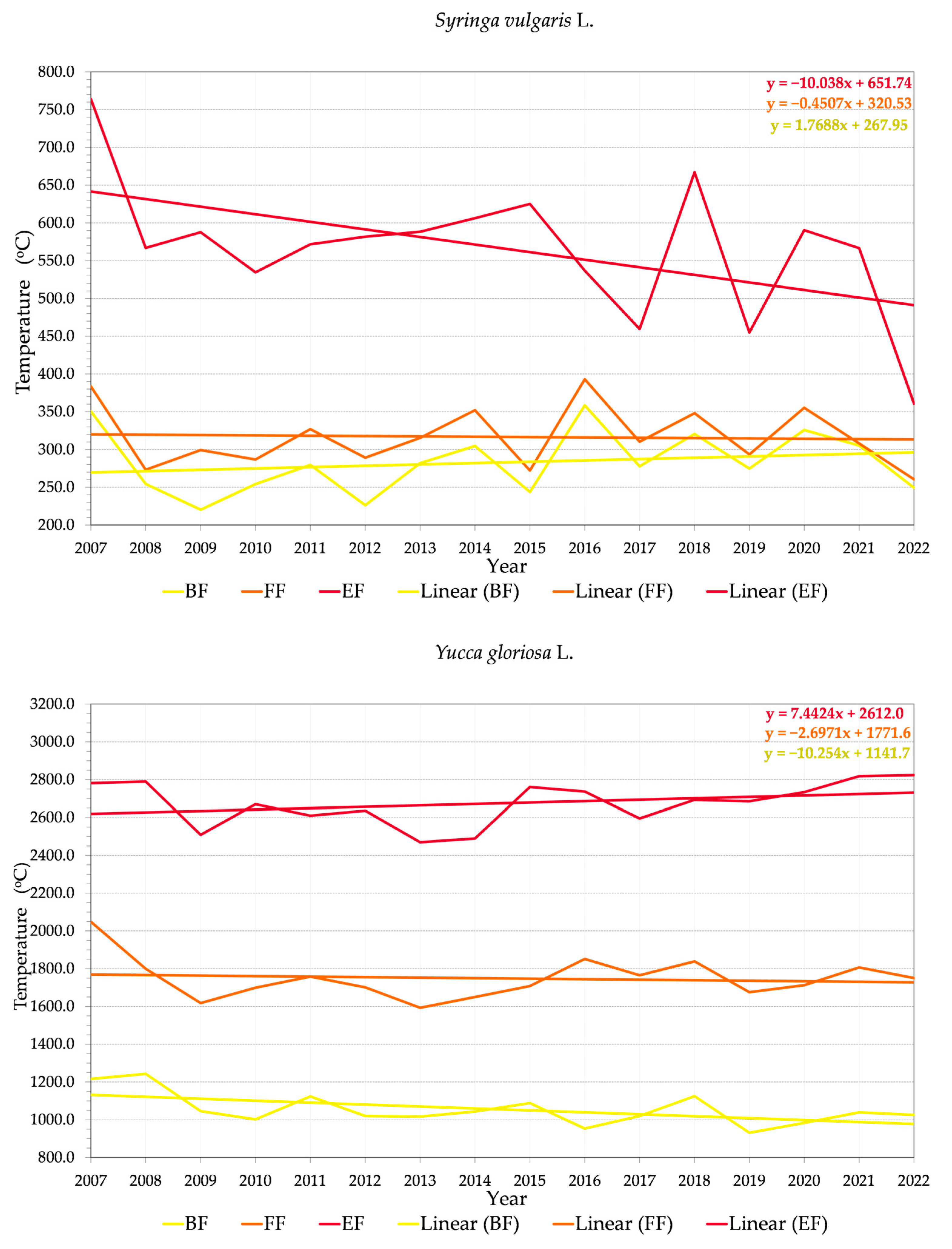
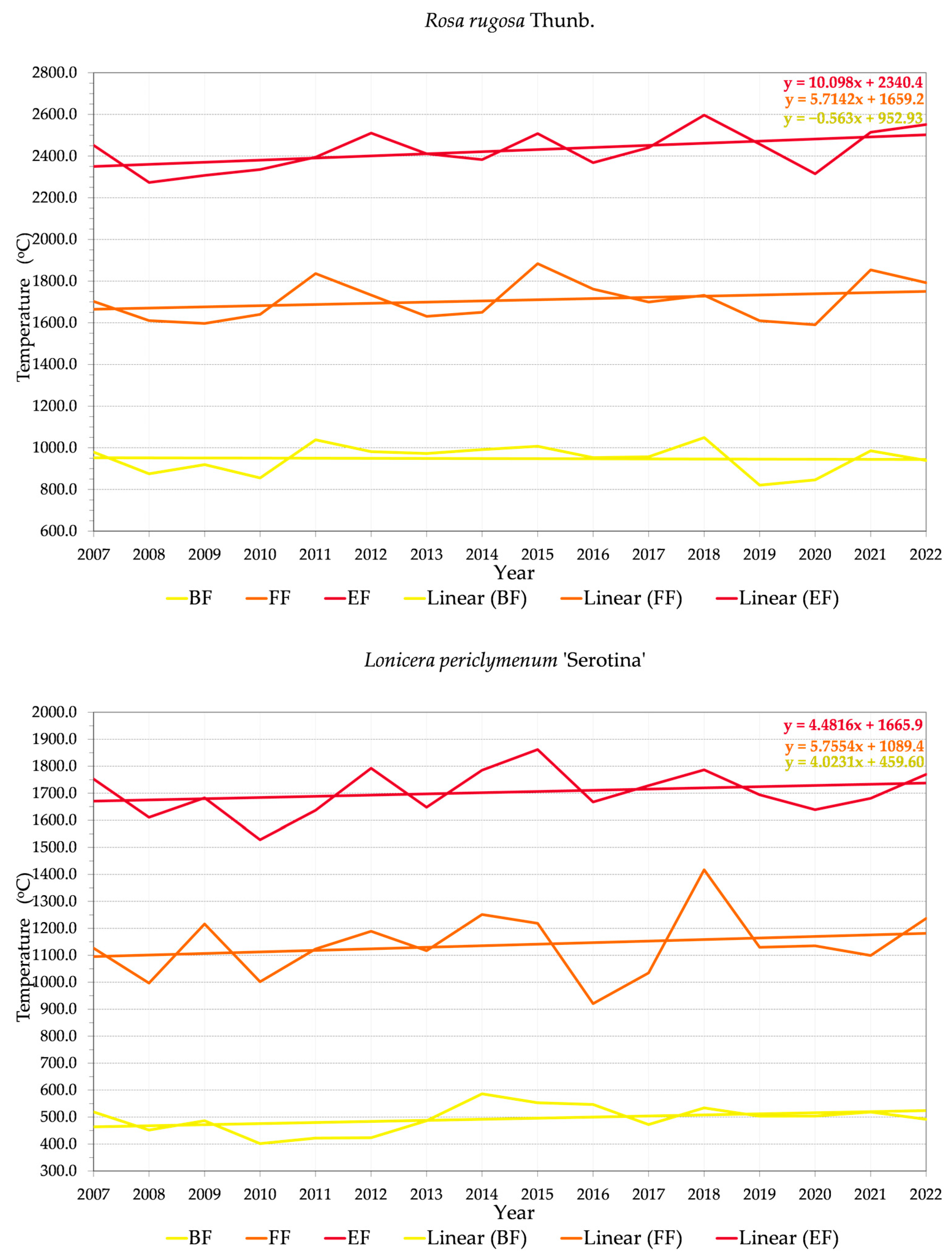
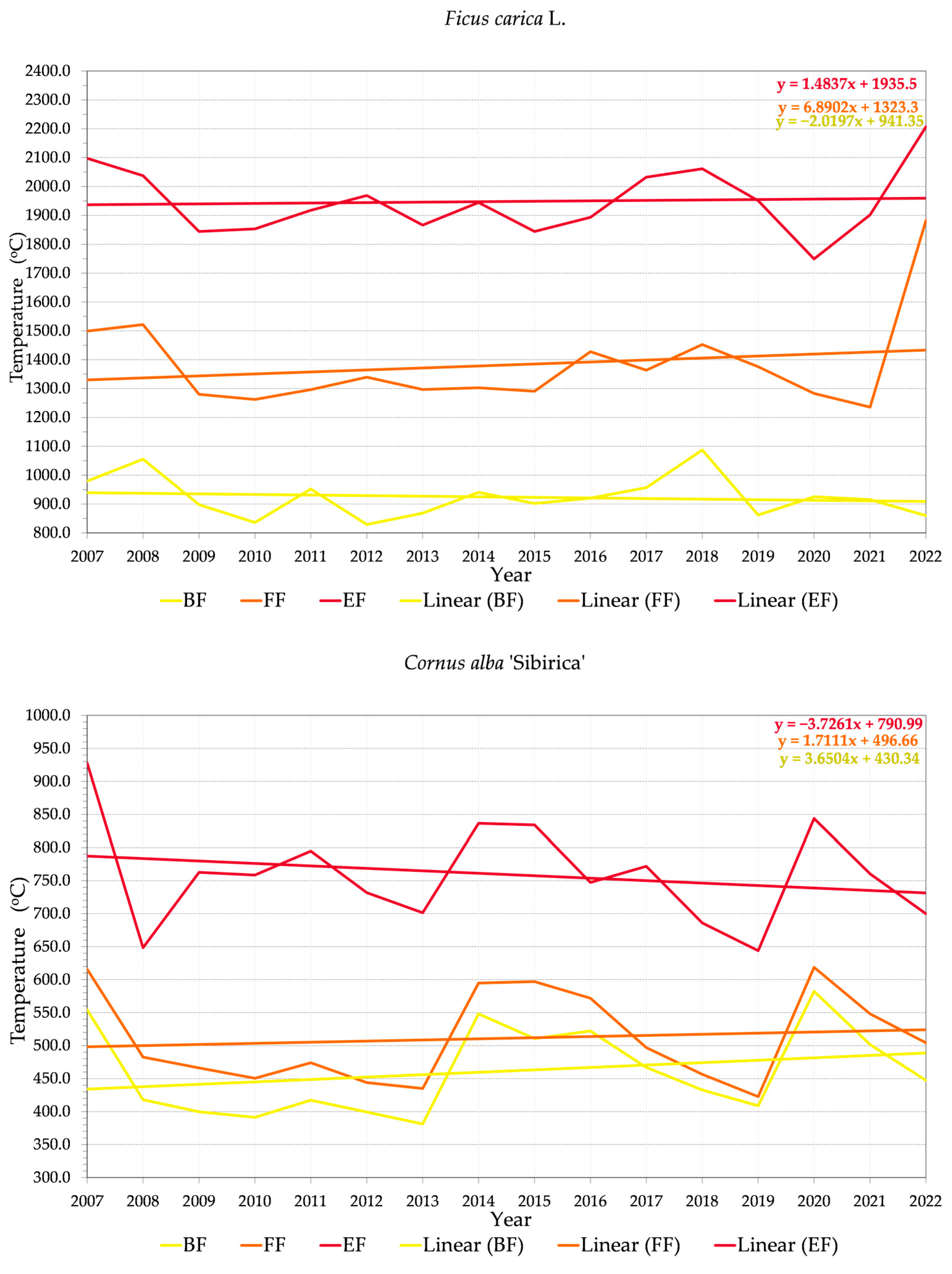
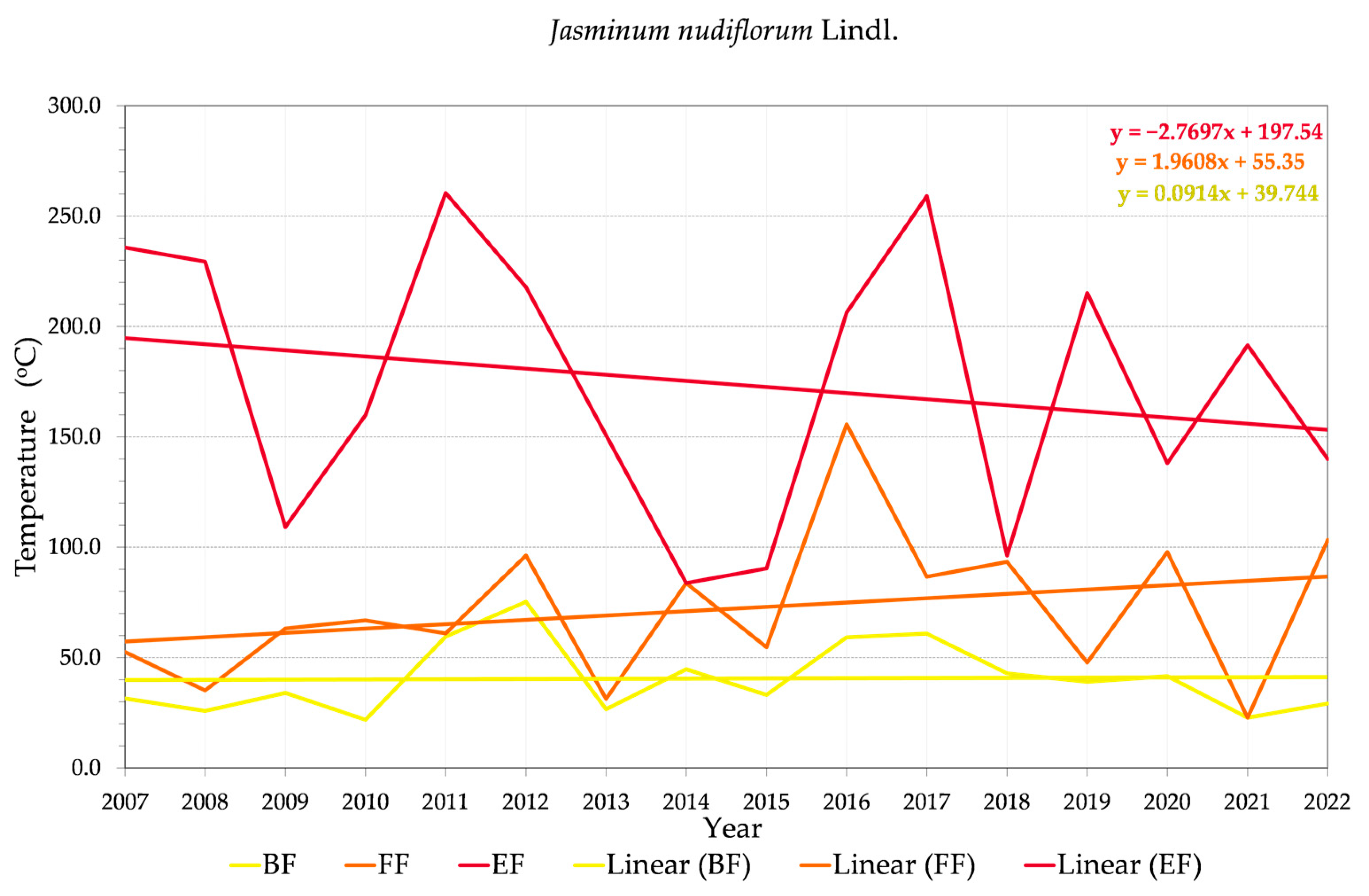
3.2. Phenology and Flowering Patterns
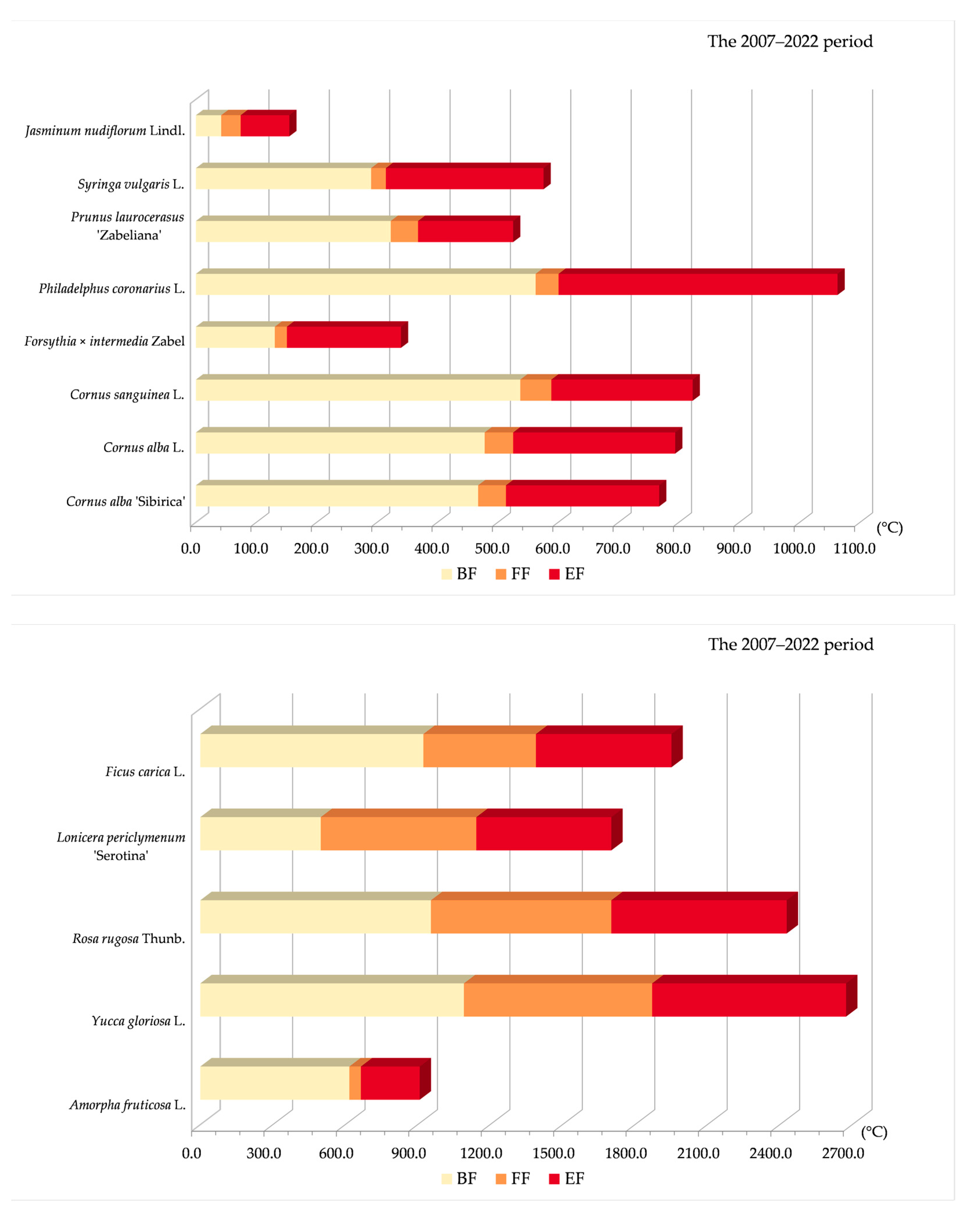
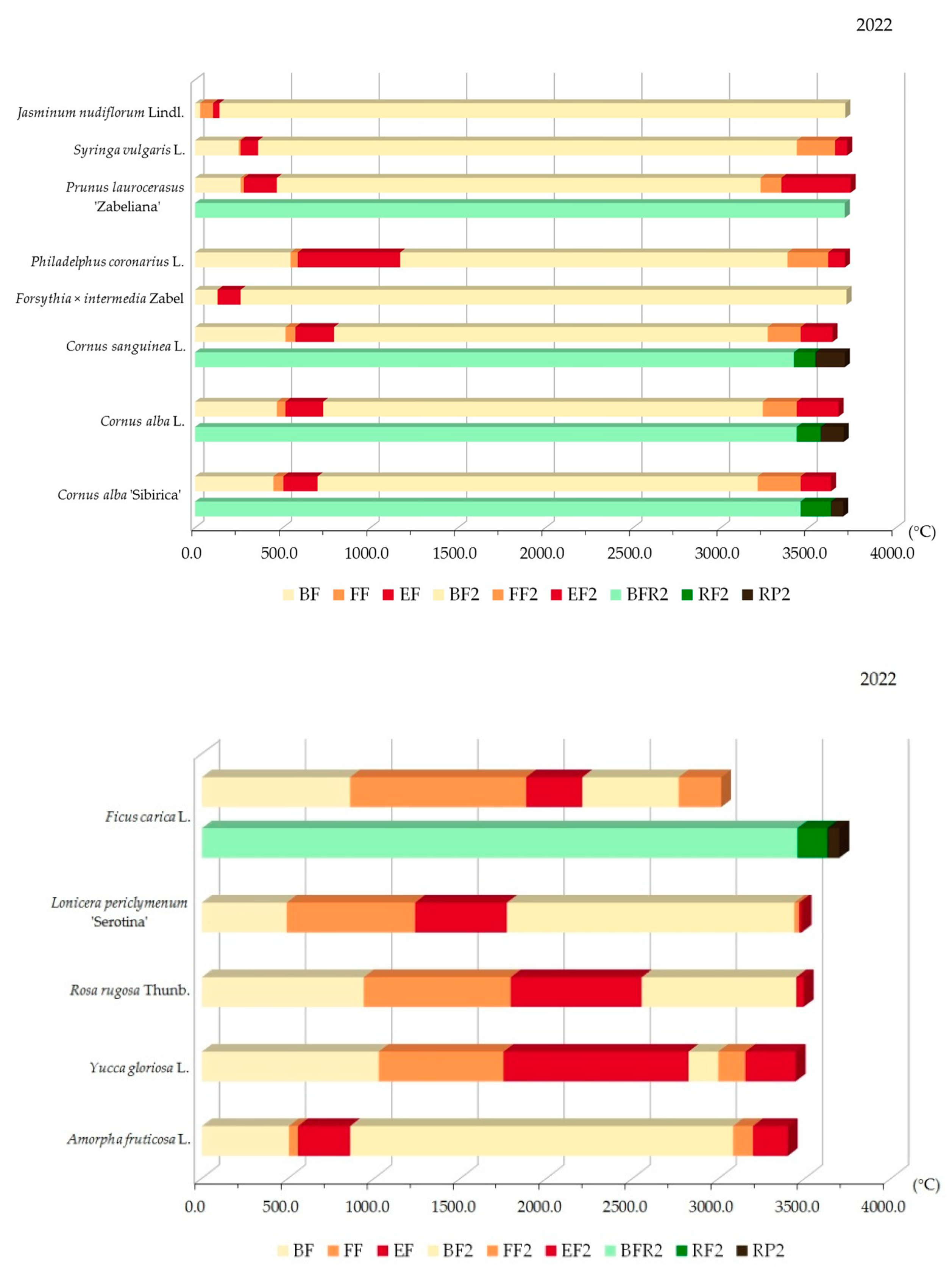
3.2.1. Variability of Relations between Flowering Phenophase and Sums of Temperatures (GDD)
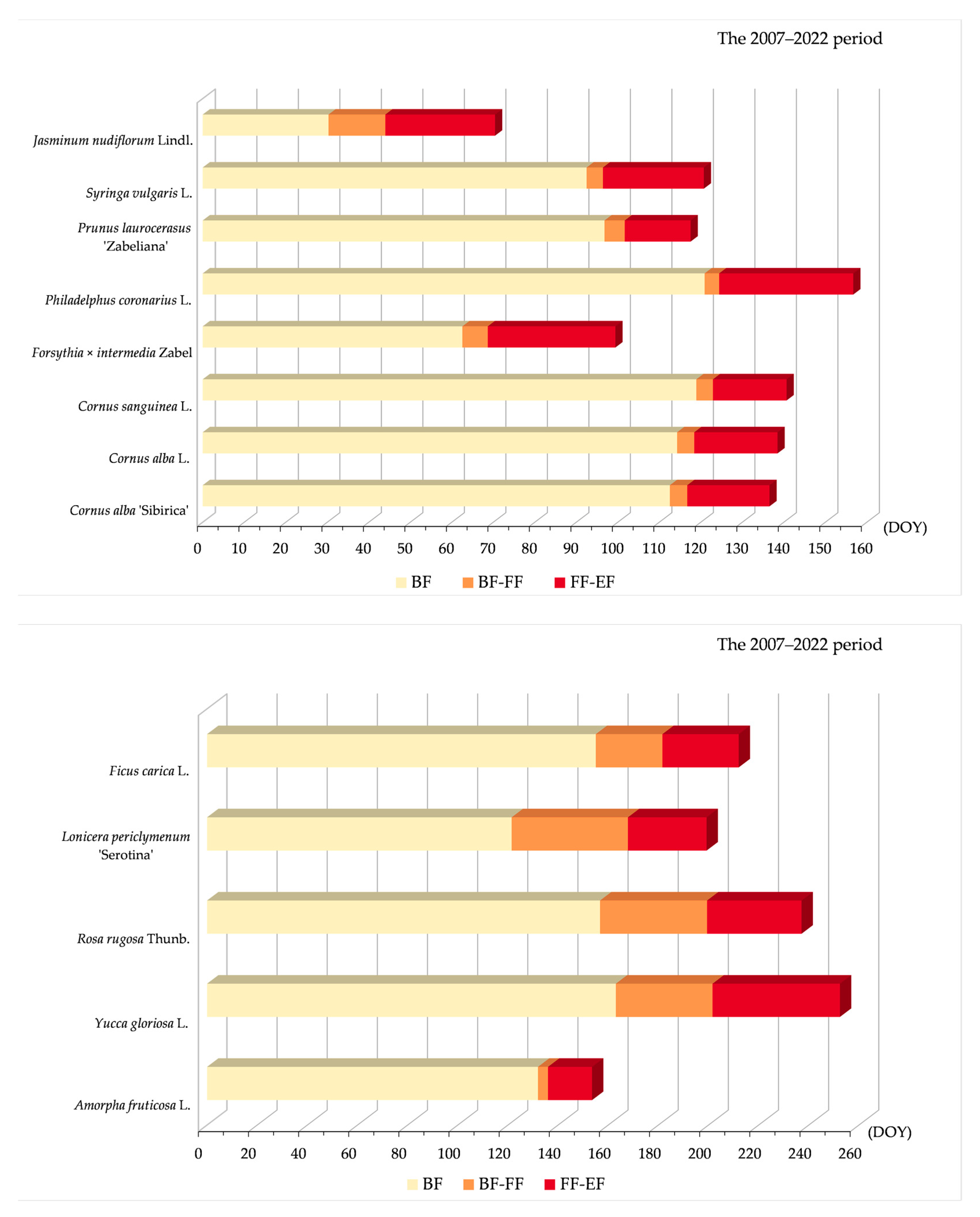
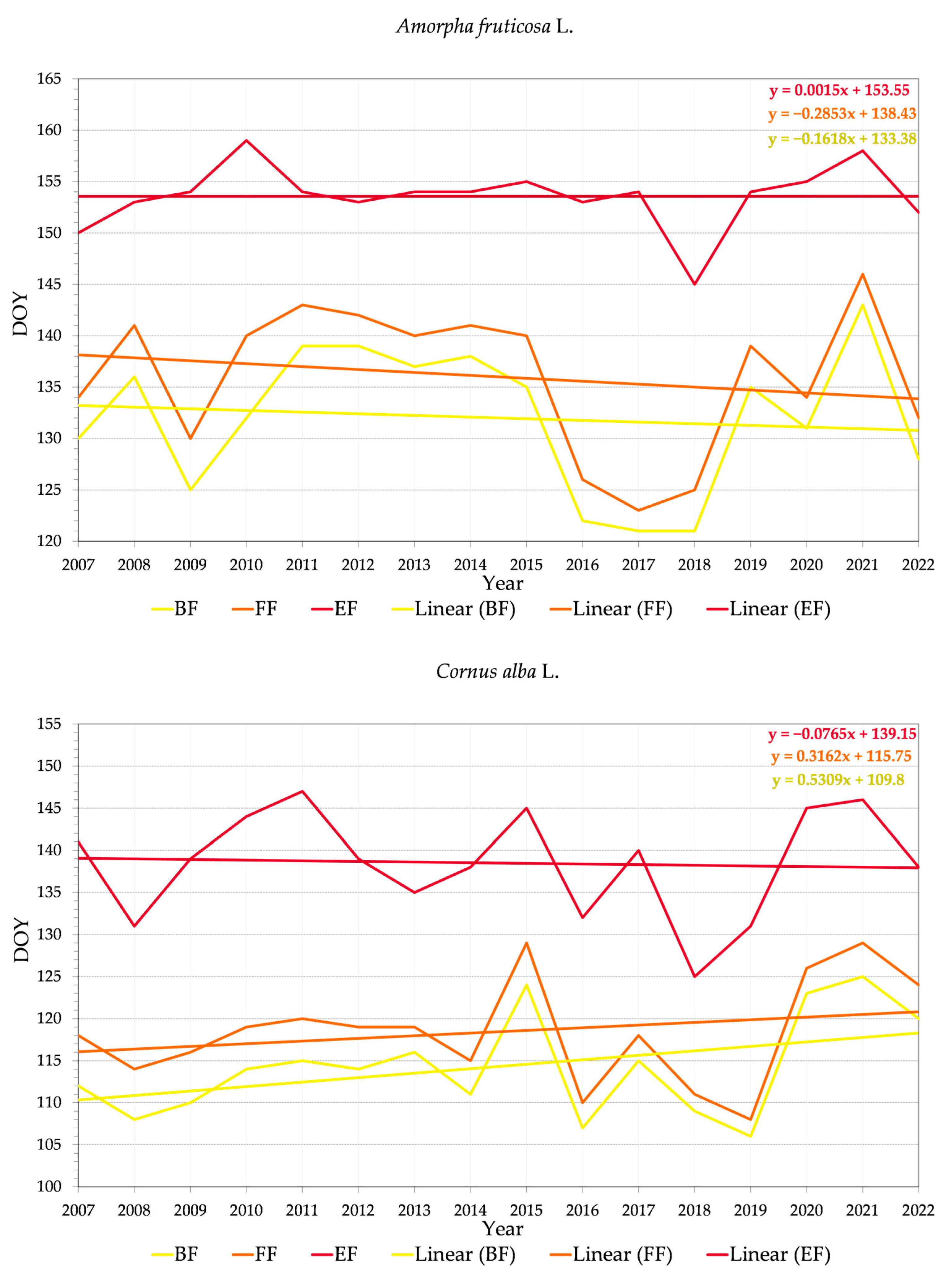
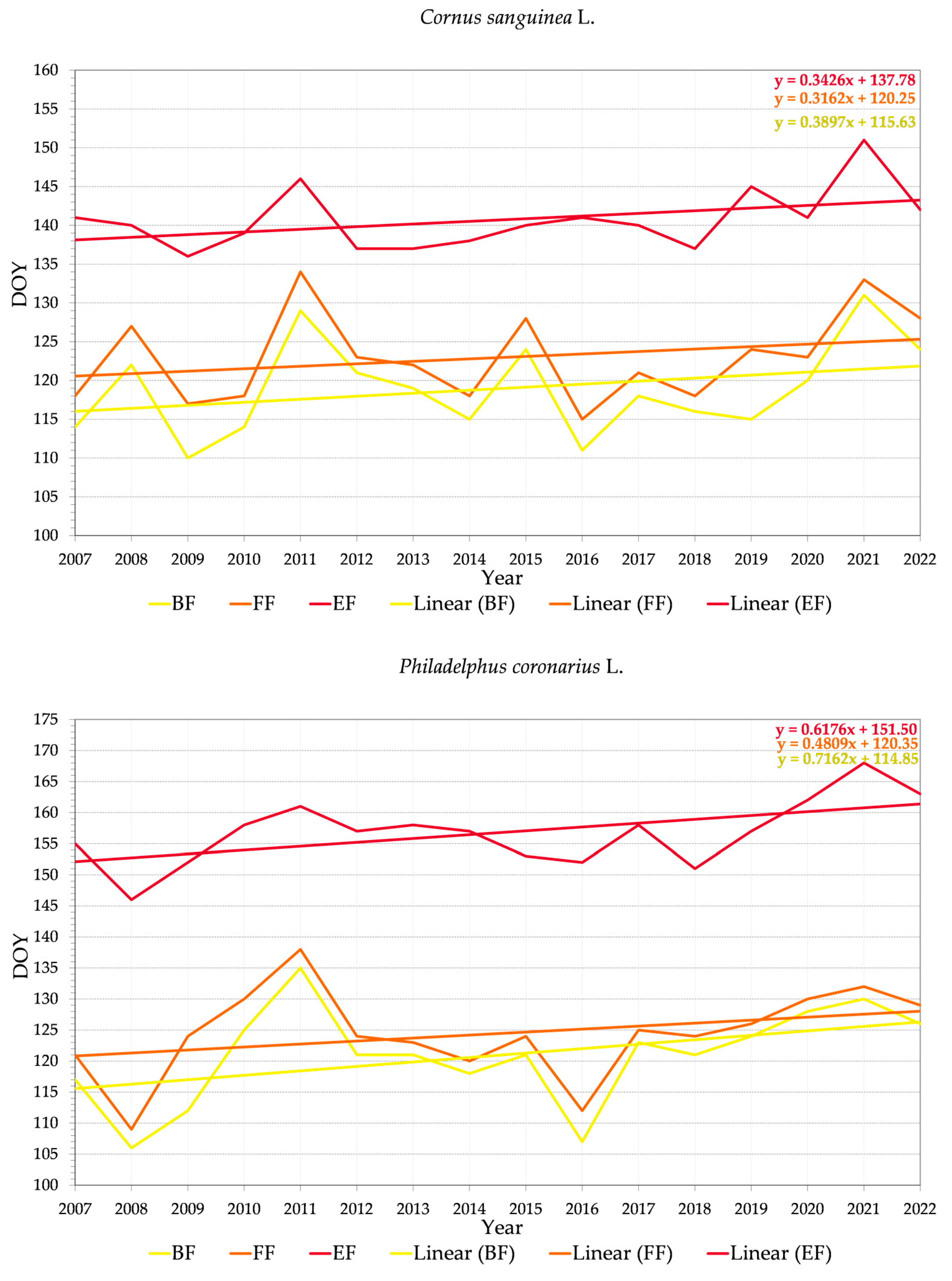
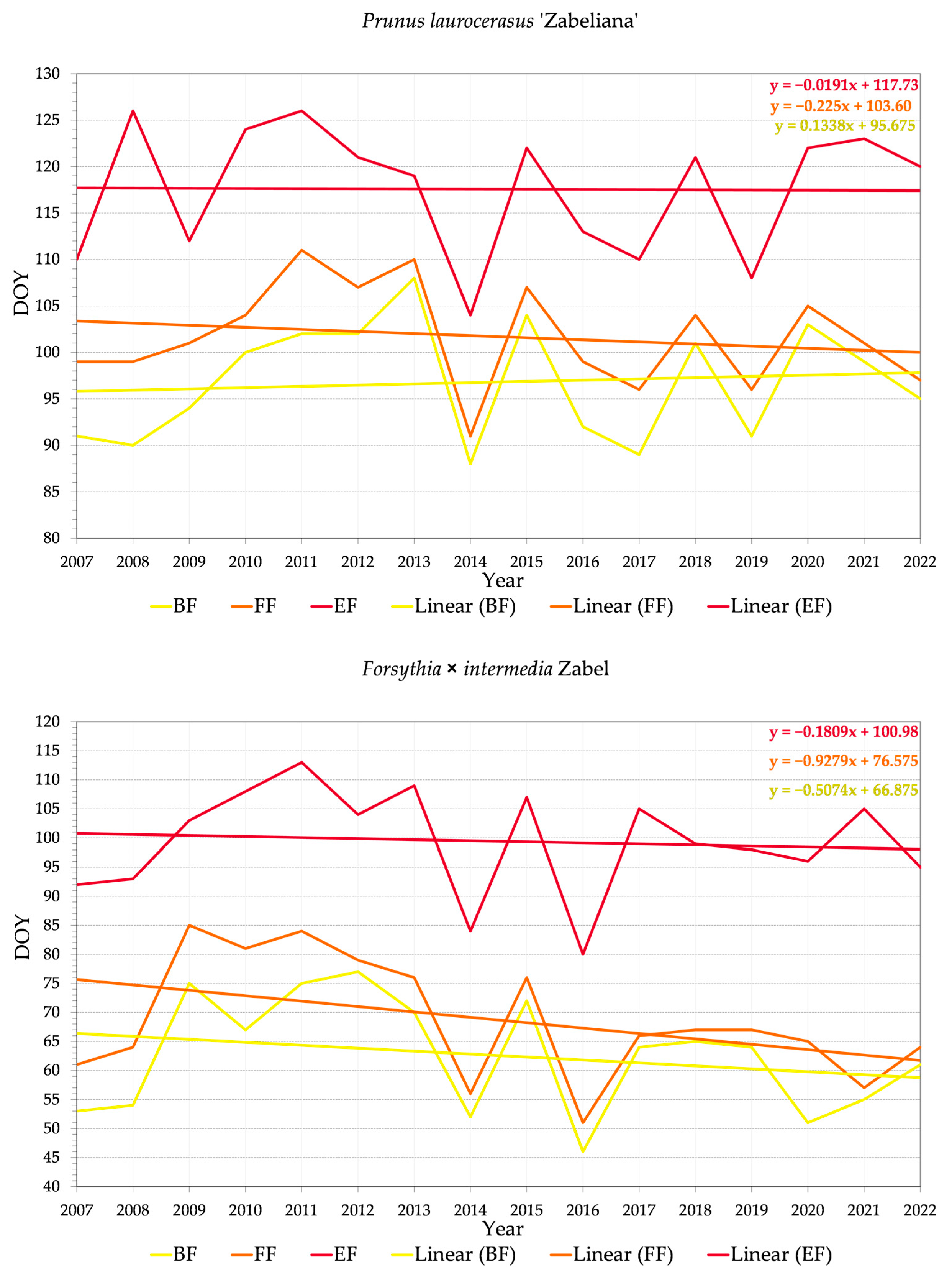
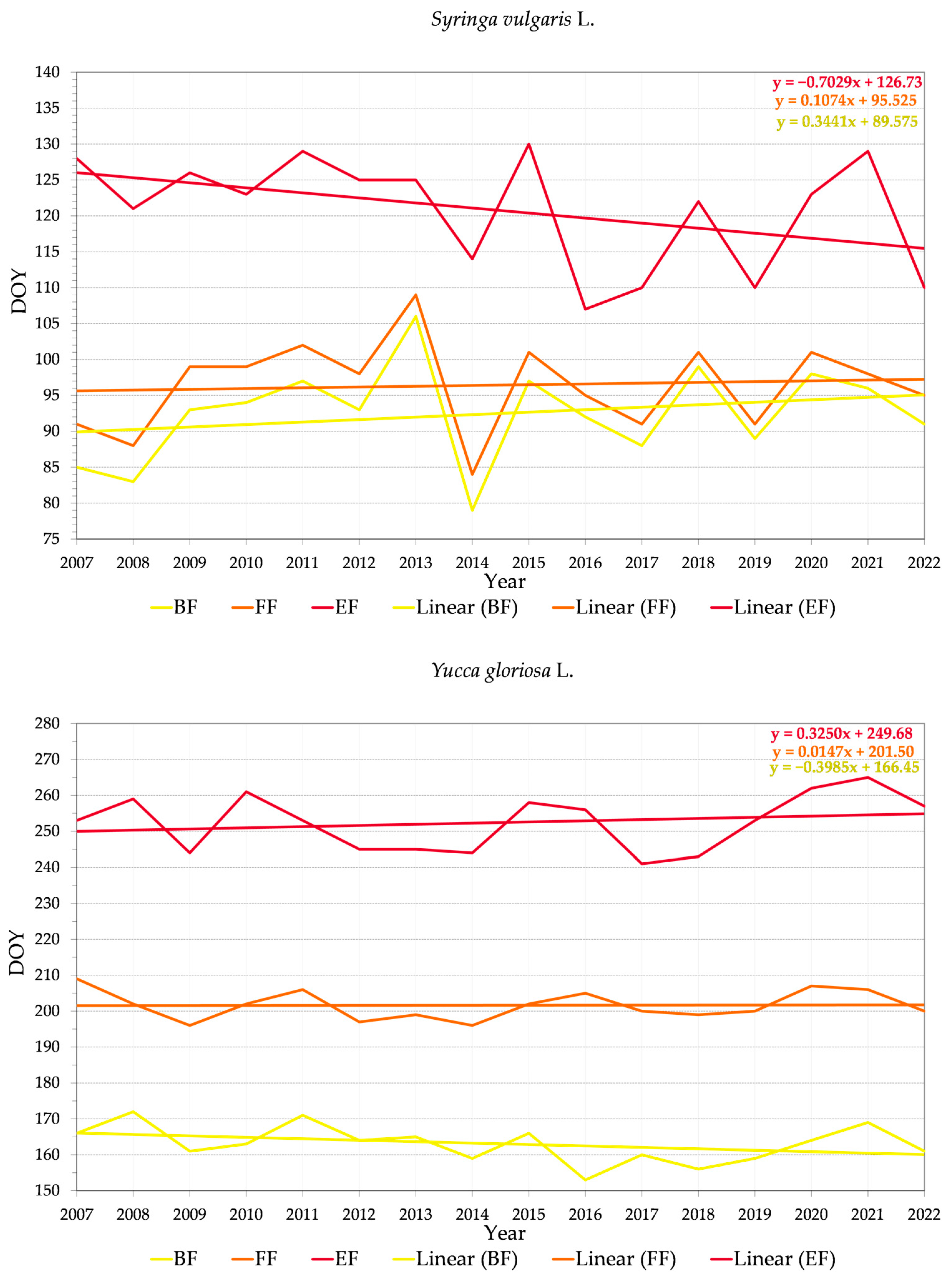
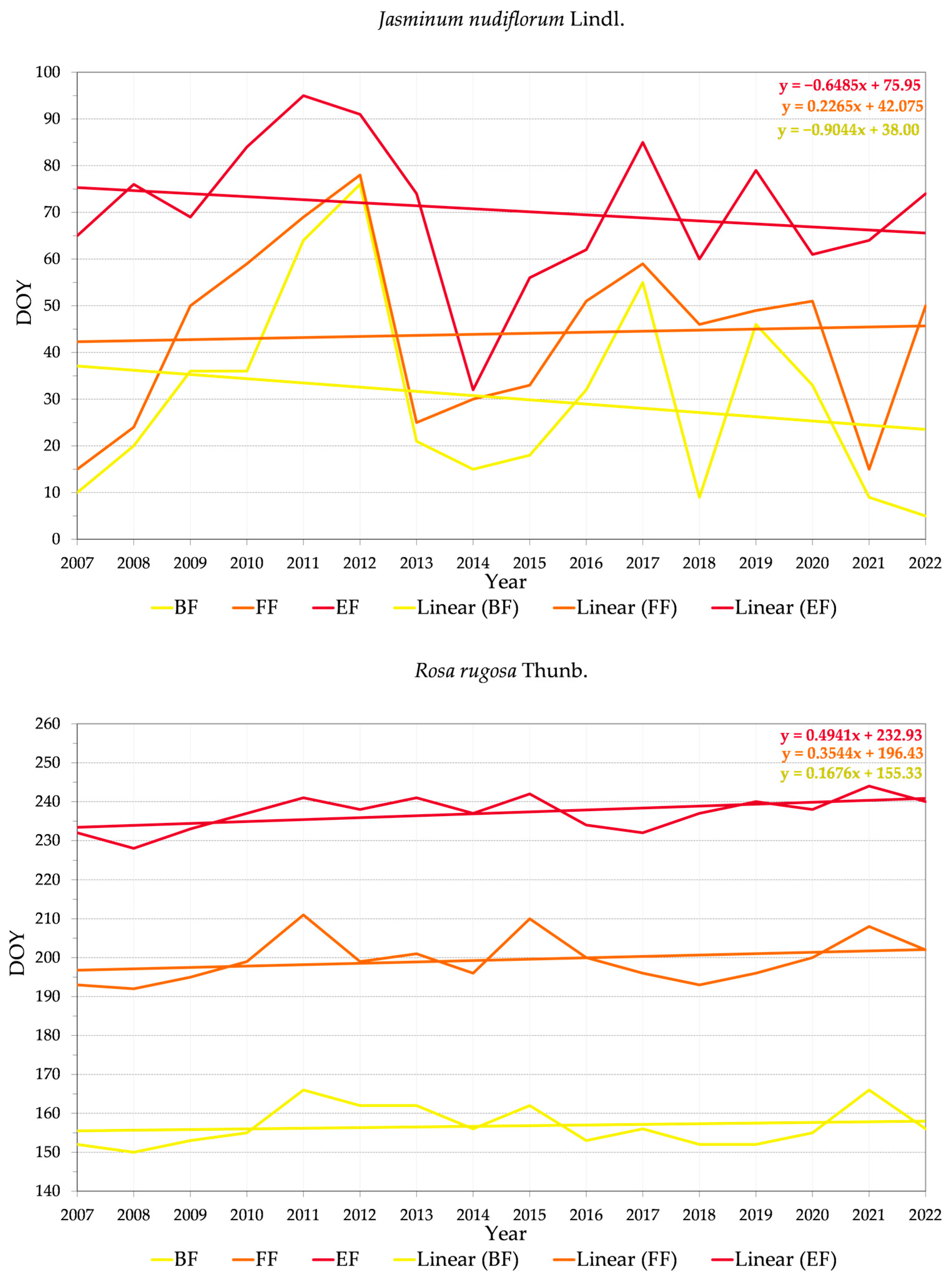
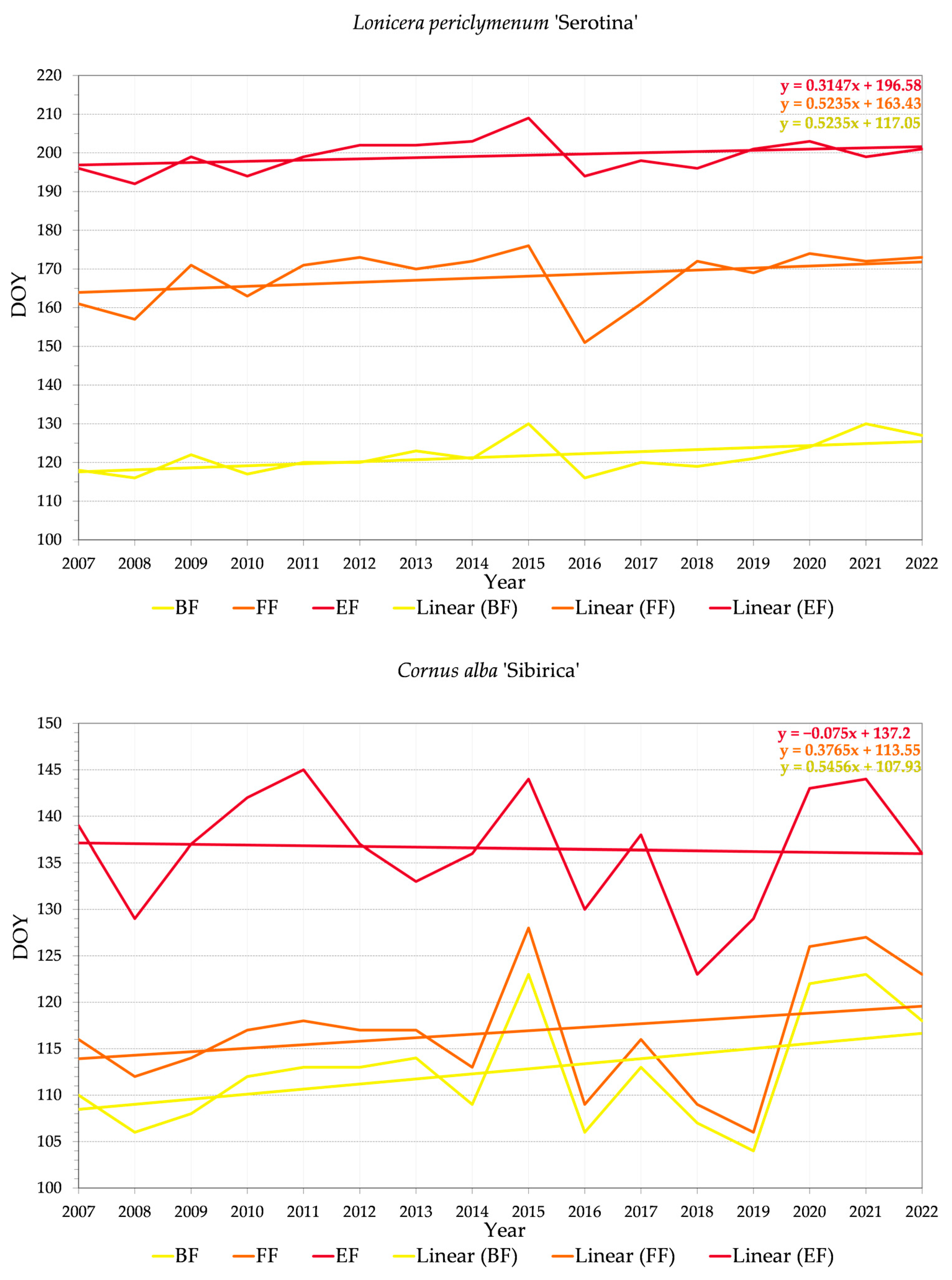
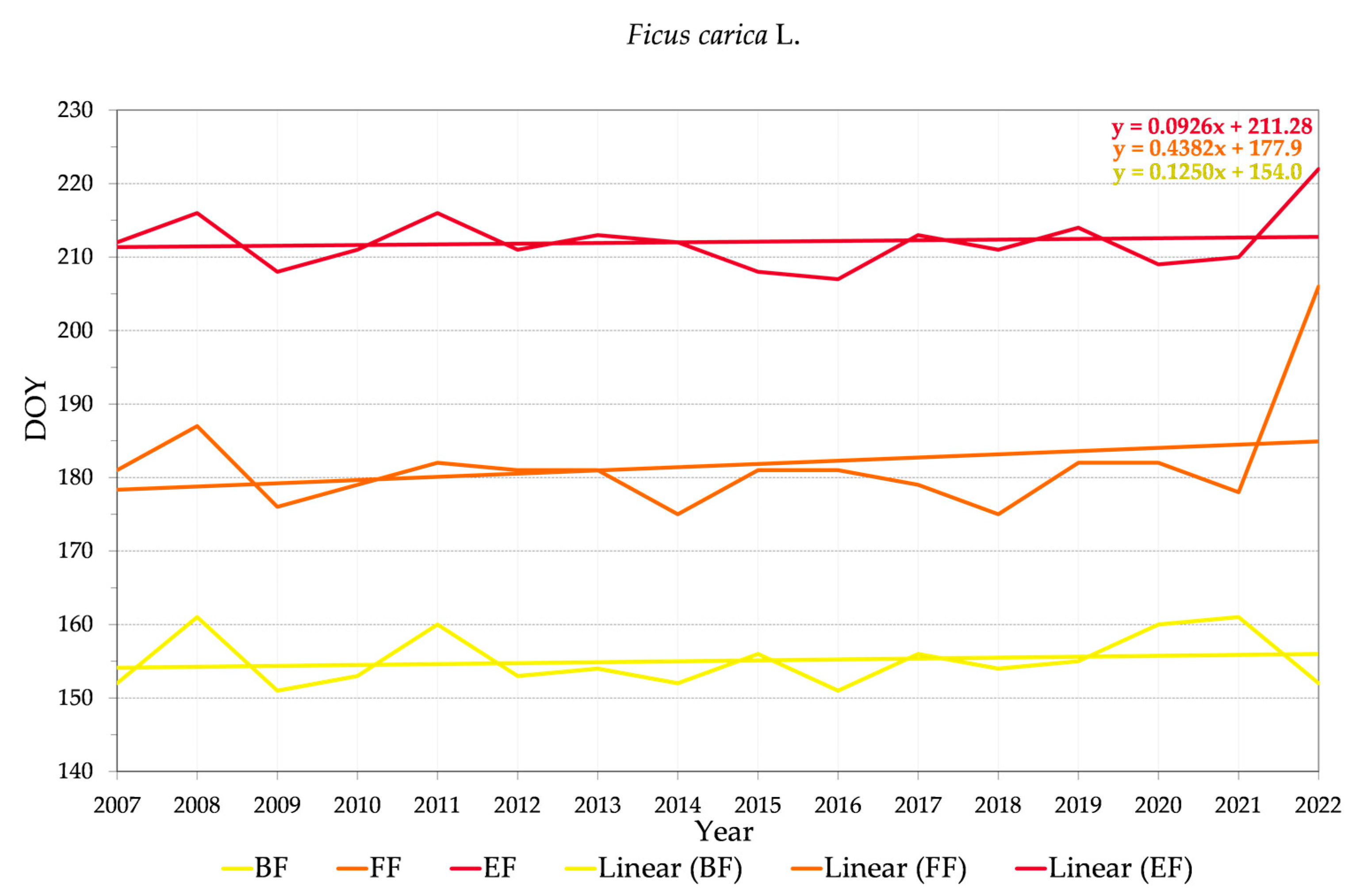
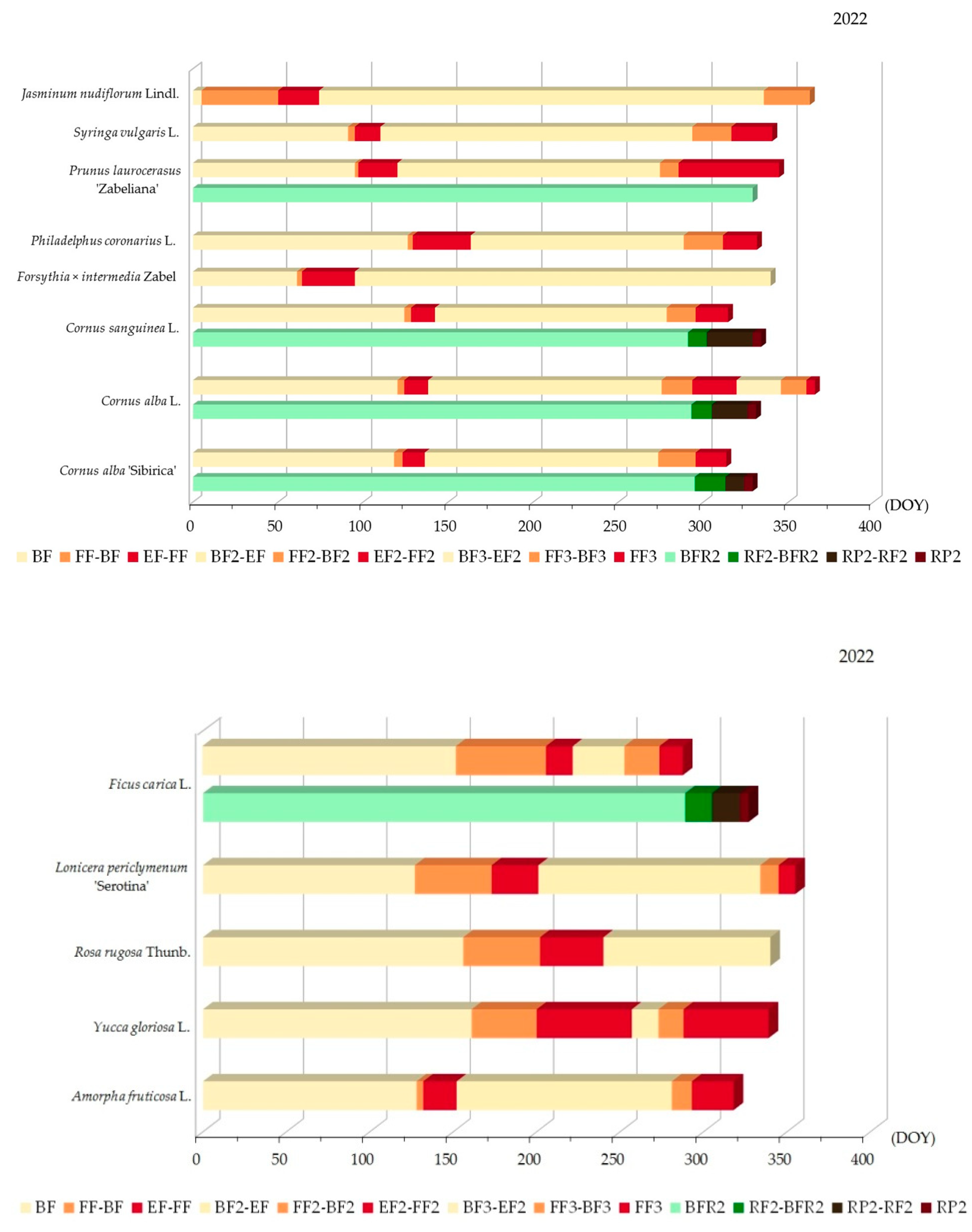
3.2.2. Variability of Relations between Flowering Phenophase and Day of the Year (DOY)
3.3. Relationships between Phenophases and Temperature during 2022
4. Discussion
4.1. Climate and Phenology
4.2. Phenological Relation to DOY and GDD
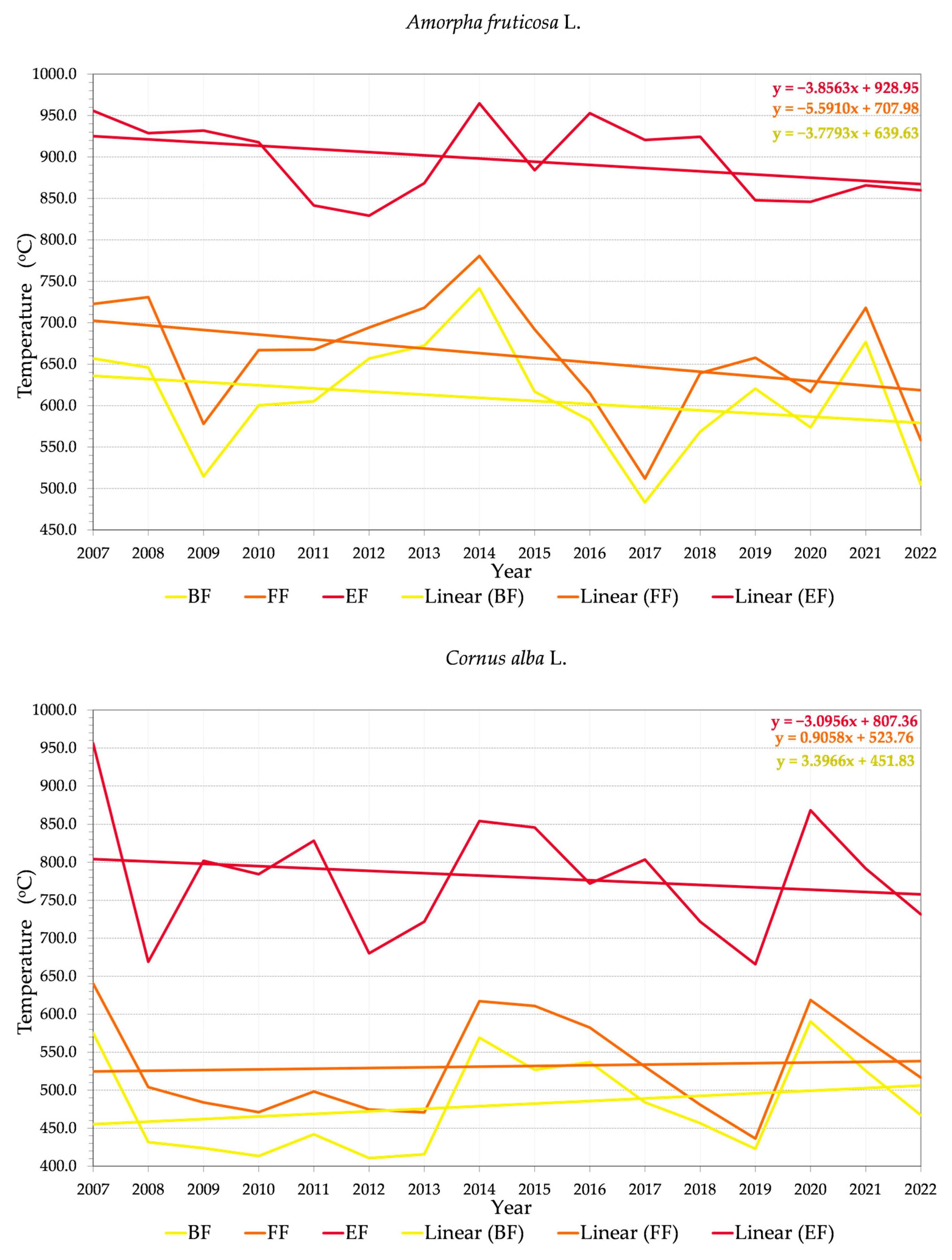
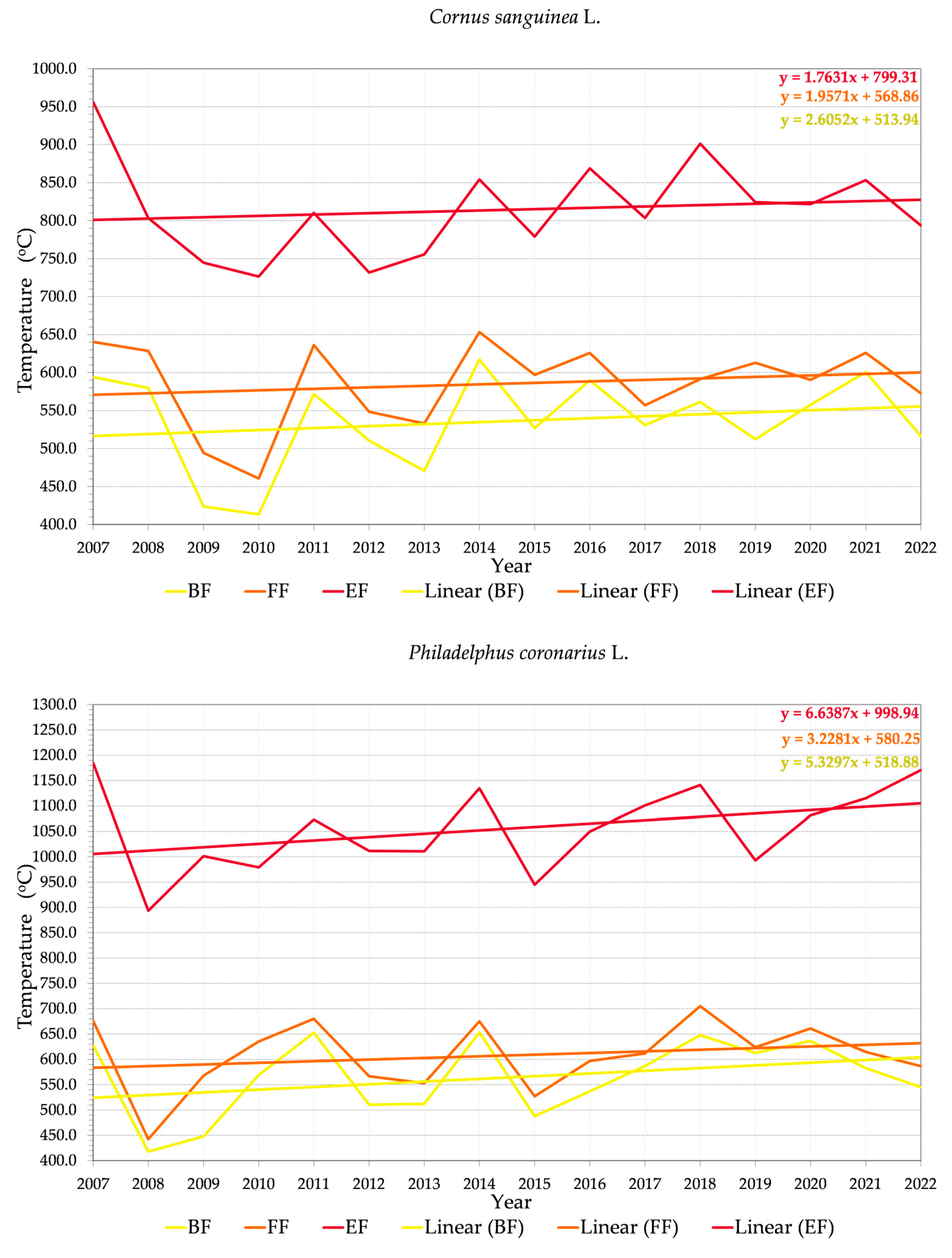
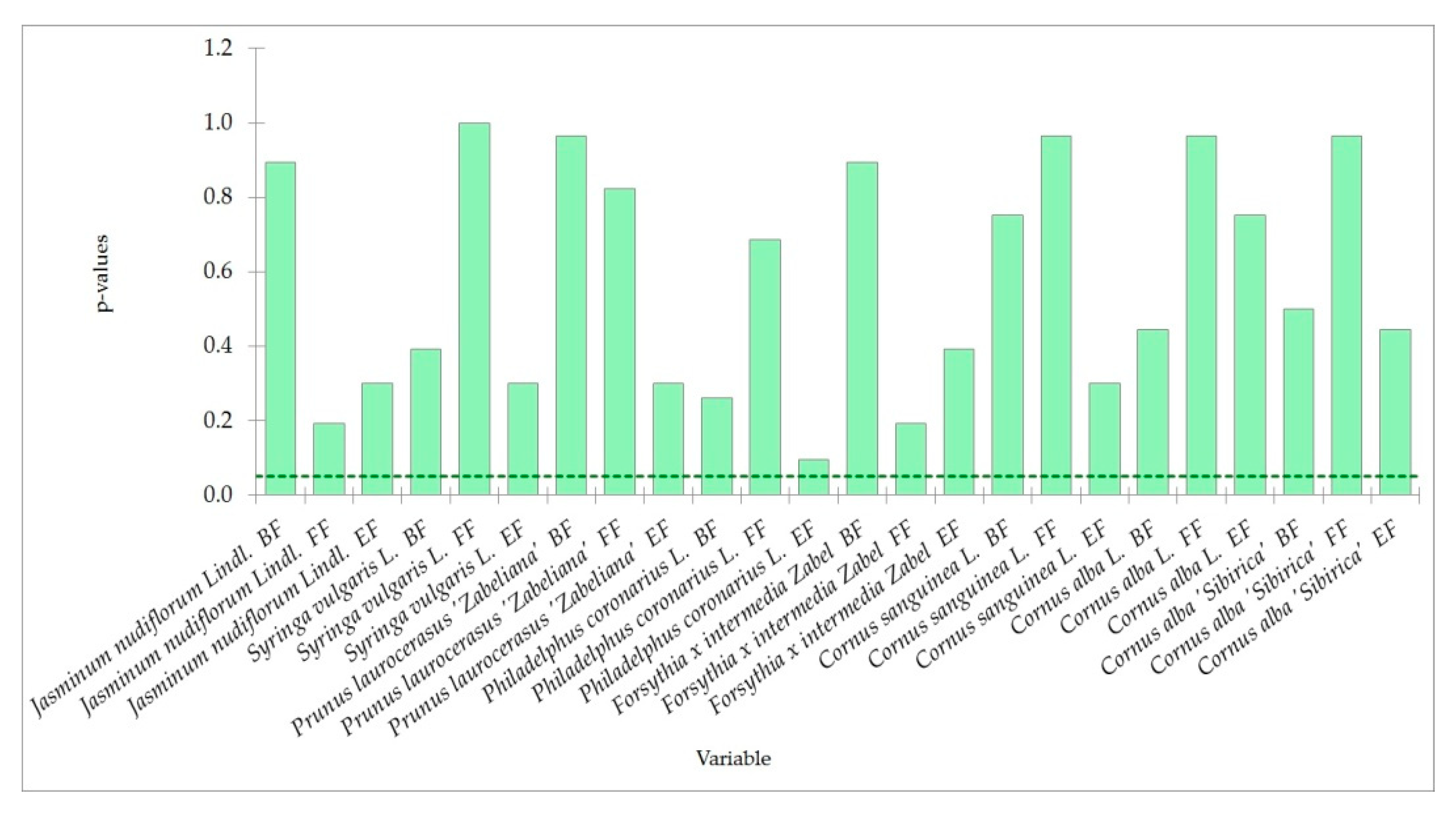
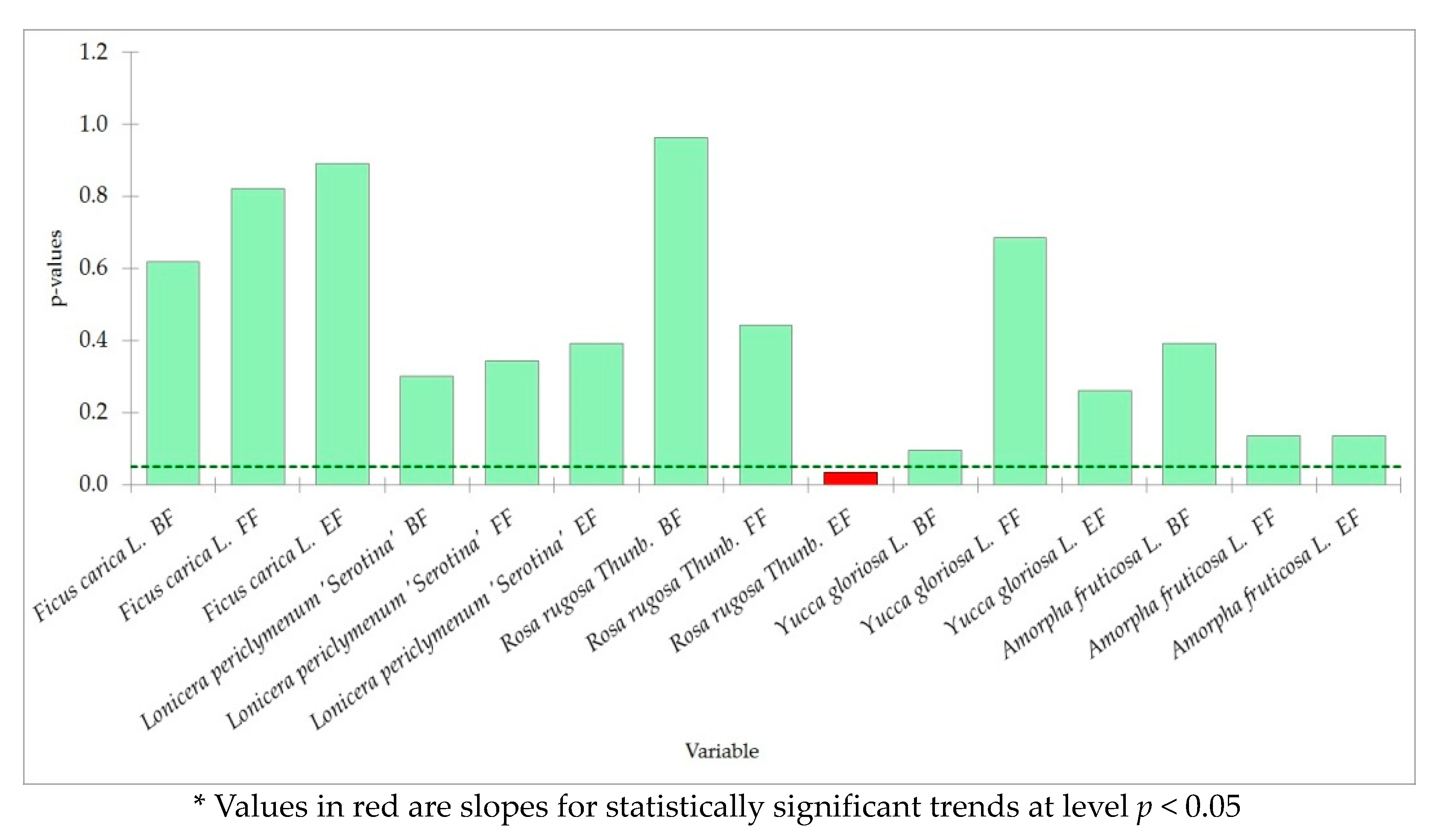
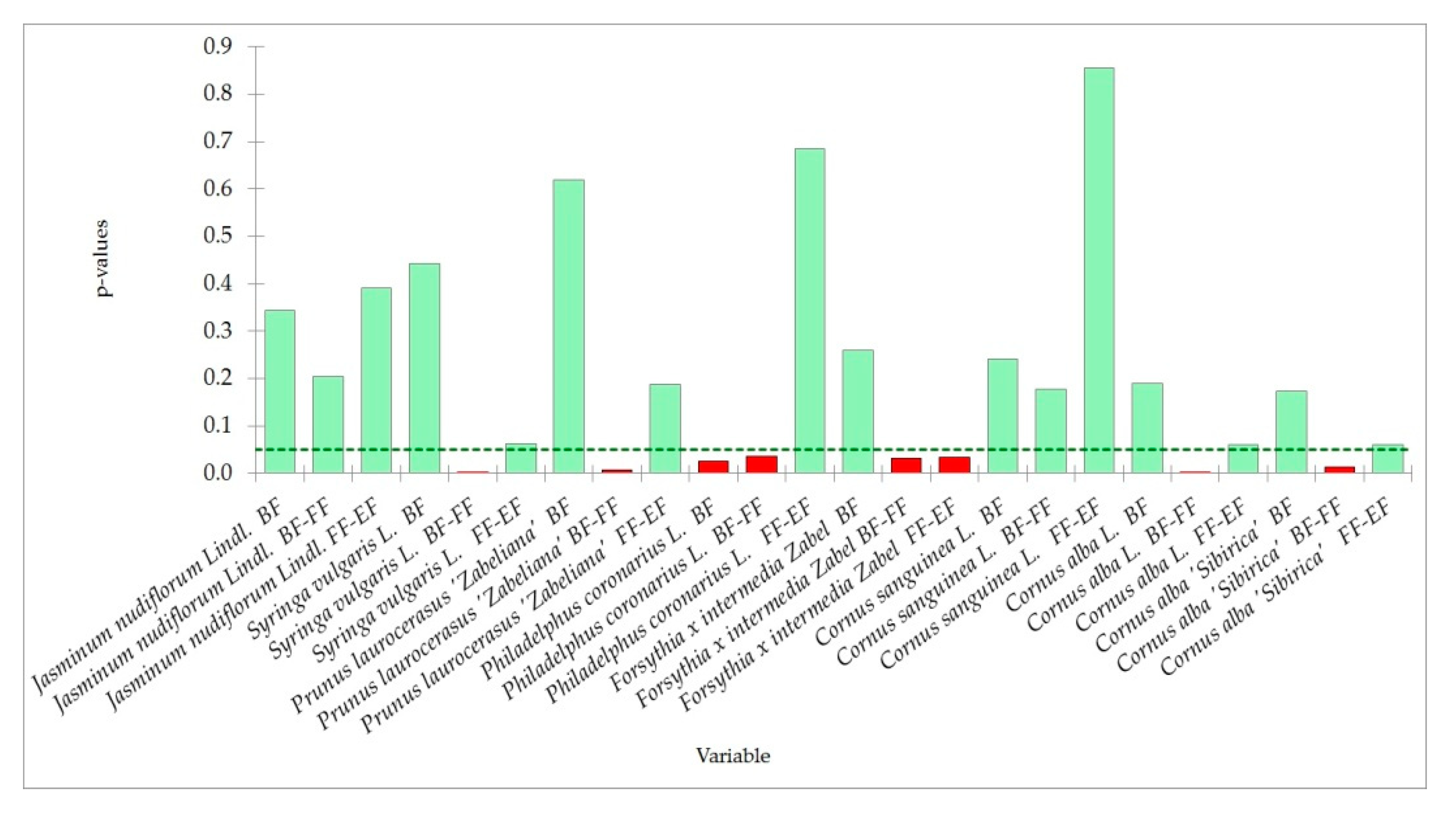
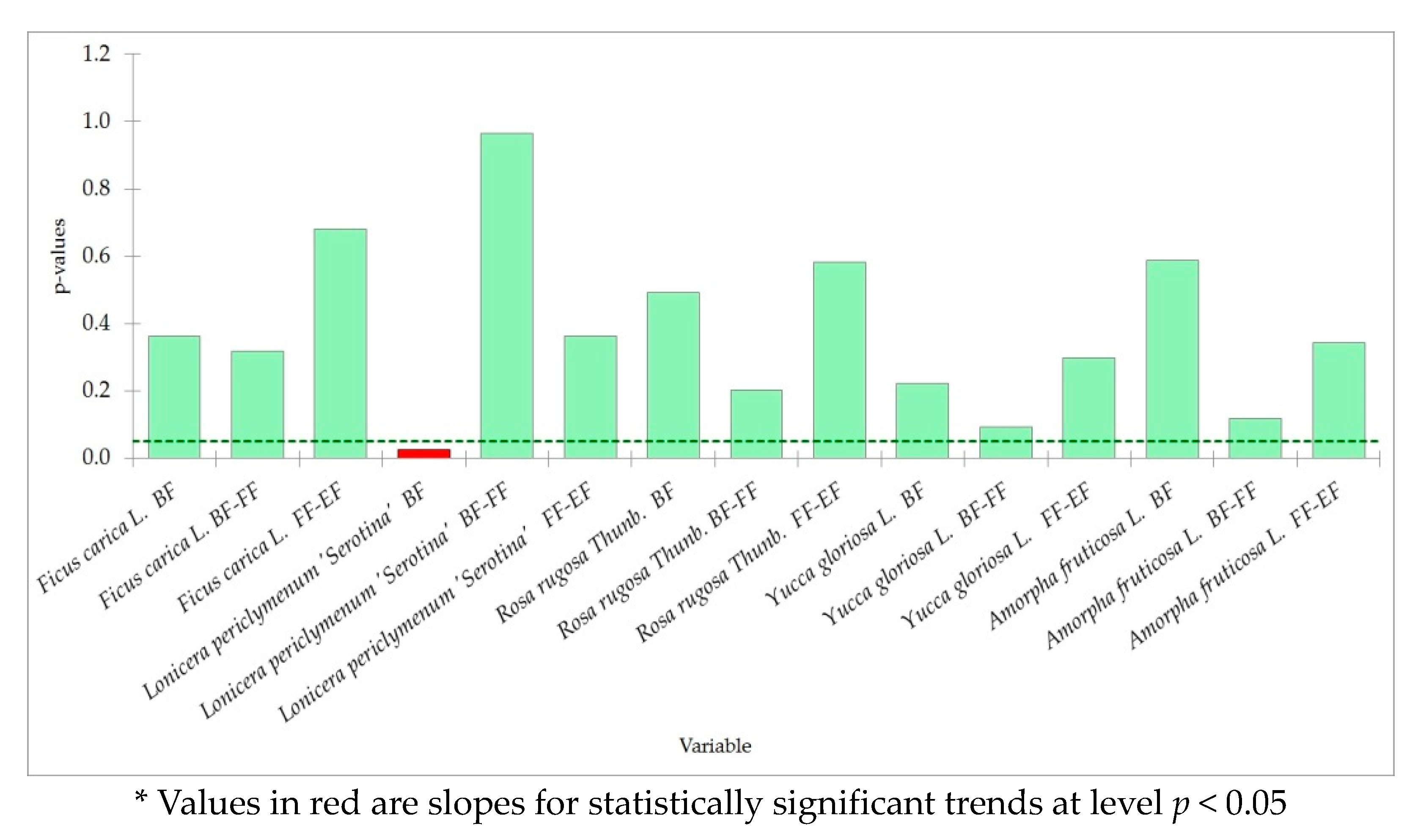
4.3. Trends in Phenology
4.4. Repeated Flowering (and Fruiting)
4.5. Third Flowering
4.6. Phenology and Landscape Design
4.7. Seasonal Shifts
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Forman, R.T.T. Urban Regions: Ecology and Planning Beyond the City; Cambridge University Press: Cambridge, UK, 2008; p. 478. [Google Scholar]
- Wu, J.G. Landscape sustainability science: Ecosystem services and human well-being in changing landscapes. Landsc. Ecol. 2013, 28, 999–1023. [Google Scholar] [CrossRef]
- Frazier, A.; Vadjunec, J.; Kedron, P.; Fagin, T. Linking landscape ecology and land system architecture for land system science: An introduction to the special issue. J. Land Use Sci. 2019, 14, 123–134. [Google Scholar] [CrossRef]
- Qiu, J.; Carpenter, S.R.; Booth, E.G.; Motew, M.; Zipper, S.C.; Kucharik, C.J.; Loheide, S.P., II; Turner, M.G. Understanding relationships among ecosystem services across spatial scales and over time. Environ. Res. Lett. 2018, 13, 054020. [Google Scholar] [CrossRef]
- Forman, R.T.T. Urban Ecology: Science of Cities; Cambridge University Press: Cambridge, UK, 2014; p. 478. [Google Scholar]
- de Vries, B.J.M. Sustainability Science; Cambridge University Press: Cambridge, UK, 2013; p. 605. [Google Scholar]
- Wu, J. Linking landscape, land system and design approaches to achieve sustainability. J. Land Use Sci. 2019, 14, 173–189. [Google Scholar] [CrossRef]
- Cleland, E.; Chuine, I.; Menzel, A.; Mooney, H.; Schwartz, M. Shifting plant phenology in response to global change. Trends Ecol. Evol. 2007, 22, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Matthews, E.R.; Mazer, S.J. Historical changes in flowering phenology are governed by temperature x precipitation interactions in a widespread perennial herb in western North America. New Phytol. 2016, 210, 157–167. [Google Scholar] [CrossRef] [Green Version]
- WMO. State of the Global Climate 2021: WMO Provisional Report; WMO No. 1290; World Meteorological Organization (WMO), World Climate Data and Monitoring Programme: Geneva, Switzerland, 2021; p. 57. Available online: https://library.wmo.int/doc_num.php?explnum_id=10859 (accessed on 7 December 2022).
- Vitasse, Y.; Porté, A.J.; Kremer, A.; Michalet, R.; Delzon, S. Responses of canopy duration to temperature changes in four temperate tree species: Relative contributions of spring and autumn leaf phenology. Oecologia 2009, 161, 187–198. [Google Scholar] [CrossRef]
- Schuster, C.; Estrella, N.; Menzel, A. Shifting and extension of phonological periods with increasing temperature along elevational transects in southern Bavaria. Plant Biol. 2014, 16, 332–344. [Google Scholar] [CrossRef]
- Liang, L.; Schwartz, M.D.; Zhang, X. Mapping temperate vegetation climate adaptation variability using normalized land surface phenology. Climate 2016, 4, 24. [Google Scholar] [CrossRef] [Green Version]
- Stojicic, D.J. The Impact of Environmental Factors on Flowering of Woody Species from Magnoliophyta group in Belgrade. Ph.D. Thesis, Univerzitet u Beogradu-Šumarski fakultet, Belgrade, Serbia, 2014; p. 243. (In Serbian). [Google Scholar]
- Rathcke, B.; Lacey, E.P. Phenological patterns of terrestrial plants. Annu. Rev. Ecol. Syst. 1985, 16, 179–214. [Google Scholar] [CrossRef]
- Lindholm, G. Land and Landscape; Linking Use, Experience and Property Development in Urban Areas. Land 2019, 8, 137. [Google Scholar] [CrossRef] [Green Version]
- Lalić, B.; Ejcinger, J.; Dalamarta, A.; Orlandini, S.; Firanj Sremac, A.; Paher, B. Meteorology and Climatology for Agronomists. [Meteorologija i klimatologija za agronome]; Univerzitet u Novom Sadu-Poljoprivredni Fakultet: Novi Sad, Serbia, 2021; p. 219. (In Serbian) [Google Scholar]
- Waser, N.M.; Real, L.A. Effective mutualism between sequentially flowering plant species. Nature 1979, 281, 670. [Google Scholar] [CrossRef]
- Memmott, J.; Craze, P.G.; Waser, N.M.; Price, M.V. Global warming and the disruption of plant–pollinator interactions. Ecol. Lett. 2007, 10, 710–717. [Google Scholar] [CrossRef] [PubMed]
- Fitter, A.; Fitter, R.; Harris, I.; Williamson, M. Relationships between first flowering date and temperature in the flora of a locality in central England. Funct. Ecol. 1995, 9, 55–60. [Google Scholar] [CrossRef]
- Bock, A.; Sparks, T.H.; Estrella, N.; Jee, N.; Casebow, A.; Schunk, C.; Leuchner, M.; Menzel, A. Changes in first flowering dates and flowering duration of 232 plant species on the island of Guernsey. Glob. Chang. Biol. 2014, 20, 3508–3519. [Google Scholar] [CrossRef]
- König, P.; Tautenhahn, S.; Cornelissen, J.H.C.; Kattge, J.; Bönisch, G.; Römermann, C. Advances in flowering phenology across the northern hemisphere are explained by functional traits. Glob. Ecol. Biogeogr. 2018, 27, 310–321. [Google Scholar] [CrossRef]
- Büntgen, U.; Piermattei, A.; Krusic, P.J.; Esper, J.; Sparks, T.; Crivellaro, A. Plants in the UK flower a month earlier under recent warming. Proc. Biol. Sci. 2022, 289, 20212456. [Google Scholar] [CrossRef]
- Trunschke, J.; Stöcklin, J. Plasticity of flower longevity in alpine plants is increased in populations from high elevation compared to low elevation populations. Alp. Bot. 2017, 127, 41–51. [Google Scholar] [CrossRef] [Green Version]
- Bucher, S.; Römermann, C. Flowering patterns change along elevational gradients and relate to life-history strategies in 29 herbaceous species. Alp. Bot. 2020, 130, 41–58. [Google Scholar] [CrossRef] [Green Version]
- Vázquez, D.P.; Vitale, N.; Dorado, J.; Amico, G.; Stevani, E.L. Phenological mismatches and the demography of solitary bees. Proc. R. Soc. B Biol. Sci. 2023, 290, 20221847. [Google Scholar] [CrossRef]
- Roulston, T.; Goodell, K. The Role of Resources and Risks in Regulating Wild Bee Populations. Annu. Rev. Entomol. 2011, 56, 293–312. [Google Scholar] [CrossRef] [Green Version]
- Goulson, D.; Nicholls, E.; Botías, C.; Rotheray, E.L. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 2015, 347, 1255957. [Google Scholar] [CrossRef]
- Belsky, J.; Joshi, N. Impact of biotic and abiotic stressors on managed and feral bees. Insects 2019, 10, 233. [Google Scholar] [CrossRef] [Green Version]
- Parreño, M.A.; Alaux, C.; Brunet, J.; Buydens, L.; Filipiak, M.; Henry, M.; Keller, A.; Klein, A.; Kuhlmann, M.; Leroy, C.; et al. Critical links between biodiversity and health in wild bee conservation. Trends Ecol. Evol. 2022, 37, 309–321. [Google Scholar] [CrossRef] [PubMed]
- Hobbie, S. Effects of plant species on nutrient cycling. Trends Ecol. Evol. 1992, 7, 336–339. [Google Scholar] [CrossRef] [PubMed]
- Morris, L.A. Soil Development and Properties | Nutrient Cycling. In Encyclopedia of Forest Sciences; Burley, J., Ed.; Elsevier: Amsterdam, The Netherlands, 2004; pp. 1227–1235. [Google Scholar]
- González de Andrés, E. Interactions between Climate and Nutrient Cycles on Forest Response to Global Change: The Role of Mixed Forests. Forests 2019, 10, 609. [Google Scholar] [CrossRef] [Green Version]
- Twining, C.W.; Shipley, J.R.; Matthews, B. Climate change creates nutritional phenological mismatches. Trends Ecol. Evol. 2022, 37, 736–739. [Google Scholar] [CrossRef]
- Patt, J.M.; Wainright, S.C.; Hamilton, G.C.; Whittinghill, D.; Bosley, K.; Dietrick, J.; Lashomb, J.H. Assimilation of carbon and nitrogen from pollen and nectar by a predaceous larva and its effects on growth and development. Ecol. Entomol. 2003, 28, 717–728. [Google Scholar] [CrossRef]
- Both, C.; van Asch, M.L.; Bijlsma, R.G.; van den Burg, A.B.; Visser, M.E. Climate change and unequal phenological changes across four trophic levels: Constraints or adaptations? J. Anim. Ecol. 2009, 78, 73–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Junker, R.R.; Blüthgen, N.; Brehm, T.; Binkenstein, J.; Paulus, J.; Martin Schaefer, H.; Stang, M. Specialization on traits as basis for the niche-breadth of flower visitors and as structuring mechanism of ecological networks. Funct. Ecol. 2013, 27, 329–341. [Google Scholar] [CrossRef]
- DeGrandi-Hoffman, G.; Corby-Harris, V.; Carroll, M.; Toth, A.L.; Gage, S.; Watkins de Jong, E.; Graham, H.; Chambers, M.; Meador, C.; Obernesser, B. The Importance of Time and Place: Nutrient Composition and Utilization of Seasonal Pollens by European Honey Bees (Apis mellifera L.). Insects 2021, 12, 235. [Google Scholar] [CrossRef] [PubMed]
- Miller-Rushing, A.J.; Inouye, D.W. Variation in the impact of climate change on flowering phenology and abundance: An examination of two pairs of closely related wildflower species. Am. J. Bot. 2009, 96, 1821–1829. [Google Scholar] [CrossRef] [PubMed]
- Forrest, J.; Inouye, D.W.; Thomson, J.D. Flowering phenology in subalpine meadows: Does climate variation influence community co-flowering patterns? Ecology 2010, 91, 431–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, J.G.; Sconiers, W.; Spasojevic, M.J.; Ashton, I.W.; Suding, K.N. Phenological Changes in Alpine Plants in Response to Increased Snowpack, Temperature, and Nitrogen. Arct. Antarct. Alp. Res. 2012, 44, 135–142. [Google Scholar] [CrossRef]
- Wray, J.; Elle, E. Flowering phenology and nesting resources influence pollinator community composition in a fragmented ecosystem. Landsc. Ecol. 2014, 30, 261–272. [Google Scholar] [CrossRef]
- Flo, V.; Bosch, J.; Arnan, X.; Primante, C.; Martín González, A.M.; Barril-Graells, H.; Rodrigo, A. Yearly fluctuations of flower landscape in a Mediterranean scrubland: Consequences for floral resource availability. PLoS ONE 2018, 13, e0191268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FHZ. Available online: https://www.fhmzbih.gov.ba/latinica/FENO/uputstvo.php (accessed on 12 October 2022).
- Koch, E.; Bruns, E.; Chmielewski, F.M.; Defila, C.; Lipa, W.; Menzel, A. Guidelines for Plant Phenological Observations; WMO Technical Commission for Climatology, Open Program Area Group on Monitoring and Analysis of Climate Variability and Change (OPAG2): Geneva, Switzerland, 2007; p. 39. [Google Scholar]
- Meier, U. BBCH-Monograph. Growth Stages of Plants. [Entwicklungsstadien von Pflanzen. Estadios de las Plantas. Stades De Développement des Plantes]; Blackwell Wissenschafts-Verla: Berlin, Germany; Vienna, Austria, 1997; p. 622. [Google Scholar]
- Meier, U. Growth Stages of Mono and Dicotyledonous Plants, BBCH Monograph, 2nd ed.; Federal Biological Research Centre for Agriculture and Forestry: Bonn, Germany, 2001; p. 158. [Google Scholar]
- Buttler, K.P.; Schmid, W. Guideline for the Phenological Observers of the German Meteorological Service (BAPH), 3rd ed.; Anleitung fur die phanologischen Beobachter des Deutschen Wetterdienstes, 3. Aufl; Deutscher Wetterdienst: Offenbach am Main, Germany, 1991; p. 191. (In German) [Google Scholar]
- Meier, U. Phenological growth stages. In Phenology: An Integrative Environmental Science; Schwartz, M.D., Ed.; Springer: Dordrecht, The Netherlands, 2003; Volume 39, pp. 269–283. [Google Scholar]
- Vučetić, V. Secular trend analysis of growing degree-days in Croatia. Idojаrаs 2009, 113, 39–46. [Google Scholar]
- McMaster, G.S.; Wilhelm, W.W. Growing degree-days one equation, two interpretations. Agric. For. Meteorol. 1997, 87, 291–300. [Google Scholar] [CrossRef] [Green Version]
- Vučetić, V.; Vučetić, M. Variations of phenological stages of olive-trees along the Adriatic coast. Period. Biol. 2005, 107, 335–340. [Google Scholar]
- Milovanović, B. Natural Causes of Climate Variability in Serbia. [Прирoдни фактoри кoлебања климе у Србији]; GI SASA–Special Editions 93; Geographical Institute Jovan Cvijić SASA: Belgrade, Serbia, 2017; p. 152. (In Serbian) [Google Scholar]
- Diress, S.A.; Bedada, T.B. Precipitation and Temperature trend analysis by Mann Kendall test: The case of Addis Ababa methodological station, Addis Ababa, Ethiopia. Afr. J. Land Policy Geospat. Sci. 2021, 4, 517–526. [Google Scholar]
- Rustum, R.; Adeloye, A.J.; Mwale, F. Spatial and temporal Trend Analysis of Long Term rainfall records in data-poor catchments with missing data, a case study of Lower Shire floodplain in Malawi for the Period 1953–2010. Hydrol. Earth Syst. Sci. Discuss. 2017, preprint. [Google Scholar] [CrossRef] [Green Version]
- Quade, D. Nonparametric Partial Correlation. In Measurement in the Social Sciences; Blalock, H.M., Ed.; North Carolina State University, Department of Statistics, Aldine: Chicago, IL, USA, 1974; pp. 369–399. [Google Scholar]
- Horvat, J.; Mijoč, J. Basic Statistics. [Osnove Statistike]; Naklada Ljevak: Zagreb, Croatia, 2012; p. 482. (In Croatian) [Google Scholar]
- Nassauer, J.I.; Opdam, P. Design in science: Extending the landscape ecology paradigm. Landsc. Ecol. 2008, 23, 633–644. [Google Scholar] [CrossRef]
- Liao, C.; Qiu, J.; Chen, B.; Chen, D.; Fu, B.; Georgescu, M.; He, C.; Jenerette, G.D.; Li, X.; Li, X.; et al. Advancing landscape sustainability science: Theoretical foundation and synergies with innovations in methodology, design, and application. Landsc. Ecol. 2020, 35, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Borgogno-Mondino, E.; Fissore, V. Reading Greenness in Urban Areas: Possible Roles of Phenological Metrics from the Copernicus HR-VPP Dataset. Remote Sens. 2022, 14, 4517. [Google Scholar] [CrossRef]
- Penzar, I.; Penzar, B. Agrometeorology. [Agrometeorologija]; Školska knjiga: Zagreb, Croatia, 2000; p. 228. (In Croatian) [Google Scholar]
- IPCC. Global Warming of 1.5 °C. An IPCC Special Report on the Impacts of Global Warming of 1.5 °C above Pre-Industrial Levels and Related Global Greenhouse Gas Emission Pathways, in the Context of Strengthening the Global Response to the Threat of Climate Change, Sustainable Development, and Efforts to Eradicate Poverty; Masson-Delmotte, V., Zhai, P., Pörtner, H.-O., Roberts, D., Skea, J., Shukla, P., Pirani, A., Moufouma-Okia, W., Péan, C., Pidcock, R., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2018; p. 616. [Google Scholar]
- IPCC. Climate Change and Land: An IPCC Special Report on Climate Change, Desertification, Land Degradation, Sustainable Land Management, Food Security, and Greenhouse Gas Fluxes in Terrestrial Ecosystems; Shukla, P.R., Skea, J., Buendia, E.C., Masson-Delmotte, V., Pörtner, H.-O., Roberts, D., Zhai, P., Slade, R., Connors, S., van Diemen, R., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2019; p. 874. [Google Scholar]
- Vujović, D.; Todorović, N. Urban-rural fog differences in Belgrade area, Serbia. Theor. Appl. Climatol. 2018, 131, 889–898. [Google Scholar] [CrossRef]
- Reich, P. Phenology of tropical forests: Patterns, causes, and consequences. Can. J. Bot. 1995, 73, 164–174. [Google Scholar] [CrossRef]
- Morellato, C. South America. In Phenology: An Integrative Environmental Science; Schwartz, M.D., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2003; pp. 75–92. [Google Scholar]
- Sanchez-Azofeifa, A.; Kalacska, M.E.; Quesada, M.; Stoner, K.E.; Lobo, J.A.; Arroyo-Mora, P. Tropical dry climates. In Phenology: An Integrative Environmental Science; Schwartz, M.D., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2003; pp. 121–138. [Google Scholar]
- Ocokoljić, M.; Petrov, D.J. Decorative Dendrology. [Dekorativna dendrologija]; Univerzitet u Beogradu-Šumarski Fakultet: Belgrade, Serbia, 2022; p. 409. (In Serbian) [Google Scholar]
- Cvjetićanin, R.; Brujić, J.; Perović, M.; Stupar, V. Dendrology. [Dendrologija]; Univerzitet u Beogradu-Šumarski Fakultet: Belgrade, Serbia, 2016; p. 557. (In Serbian) [Google Scholar]
- De Réaumur, R.A.F. Observations of the Thermometer, Made in Paris during the Year 1735 Compared with Those Which Have Been Made under the Line at Ile de France, at Algiers and in Some of Our American Islands. [Observations du Thermomètre, Faites à Paris Pendant l´Année 1735 Comparées avec Celles qui ont été Faites sous la ligne à l′Ile de France, à Alger et en Quelques-unes de Nosîles de l′Amérique]; Mémoires de l’Academie Royale des Sciences de Paris: Paris, France, 1735; pp. 545–576. (In French) [Google Scholar]
- Menzel, A. Trends in phenological phases in Europe between 1951 and 1996. Int. J. Biometeorol. 2000, 44, 76–81. [Google Scholar] [CrossRef]
- Piao, S.; Liu, Q.; Chen, A.; Janssens, I.A.; Fu, Y.; Dai, J.; Liu, L.; Lian, X.; Shen, M.; Zhu, X. Plant phenology and global climate change: Current progresses and challenges. Glob. Chang. Biol. 2019, 25, 1922–1940. [Google Scholar] [CrossRef] [PubMed]
- Schüepp, W. Phänometrische Experimente über die Winter—(x) ruhe einiger. Pflanzen 1950, 2, 205–211. [Google Scholar]
- Stemkovski, M.; Pearse, W.D.; Griffin, S.R.; Pardee, G.L.; Gibbs, J.; Griswold, T.; Neff, J.L.; Oram, R.; Rightmyer, M.G.; Sheffield, C.S.; et al. Bee phenology is predicted by climatic variation and functional traits. Ecol. Lett. 2020, 23, 1589–1598. [Google Scholar] [CrossRef]
- Filipiak, Z.M.; Denisow, B.; Stawiarz, E.; Filipiak, M. Unravelling the dependence of a wild bee on floral diversity and composition using a feeding experiment. Sci. Total Environ. 2022, 820, 153326. [Google Scholar] [CrossRef]
- Vučetić, V. Does climate change influence on beginning of common lilac flowering? Croat. Meteorol. J. 2011, 46, 45–53. [Google Scholar]
- Cosmovision. Available online: http://www.cosmovisions.com/laurierscerises.htm (accessed on 15 December 2022).
- Moisescu, E.; Antoce, A.O. Figs (Ficus carica L.) used as raw material for obtaining alcoholic fermented beverages. Beverages 2022, 8, 60. [Google Scholar] [CrossRef]
- Kikuchi, M.; Yamauchi, Y.; Takahashi, Y.; Sugiyama, M. Studies on the constituents of syringa spp viii. isolation and structures of phenylpropanoid glycosides from the leaves of syringa reticulata blume hara. Yakugaku Zasshi 1989, 109, 366–371. [Google Scholar] [CrossRef]
- Washington State University, PNW Plants. Available online: http://pnwplants.wsu.edu/PlantDisplay.aspx?PlantID=191 (accessed on 15 December 2022).
- Brickell, C.D.; Alexander, C.; Cubey, J.J.; David, J.C.; Hoffman, M.H.A.; Leslie, A.C.; Malécot, V.; Hetterscheid, W.L.A.; Jin, X. International Code of Nomenclature for Cultivated Plants (ICNCP), 9th ed.; Scripta Horticulturae 18; International Society of Horticultural Science: Leuven, Belgium, 2016; p. 190. [Google Scholar]
- Kollmann, J.; Grub, P.J. Biological Flora of Central Europe: Cornus sanguinea L. Flora 2001, 196, 161–179. [Google Scholar] [CrossRef]
- Boonnour, K.; Wainwright, H.; Hicks, R.G.T. The micropropagation of Lonicera periclymenum L. (Hony-suckle). Acta Hortic. 1988, 226, 183–190. [Google Scholar] [CrossRef]
- Dirr, M.A. Dirr’s Hardy Trees and Shrubs: An Illustrated Encyclopedia; Timber Press: Portland, OR, USA, 1997; p. 494. [Google Scholar]
- Bean, W.J. Trees & Shrubs Hardy in the British Isles, 8th ed.; John Murray: London, UK, 1976; Volume 4, pp. 127–146. [Google Scholar]
- Höchstatter, F. Yucca II (Agavaceae). Indehiscent-Fruited Species in the Southwest, Midwest and East of the USA; Fritz Hochstatter: Mannheim, Germany, 2002; p. 340. [Google Scholar]
- Ribotta, S.; Liccari, F.; Muggia, L.; Pallavicini, A.; Bagnolini, F.; Tordoni, E.; Bacaro, G. Invasion at the Edge: The Case of Rosa rugosa (Rosaceae) in Italy. Diversity 2021, 13, 645. [Google Scholar] [CrossRef]
- Chiesura, A. The role of urban parks for the sustainable city. Landsc. Urban Plan. 2003, 68, 129–138. [Google Scholar] [CrossRef]
- Wookey, P.A.; Parsons, A.N.; Welker, J.M.; Potter, J.A.; Callaghan, T.V.; Lee, A.; Press, M.C. Comparative responses of phenology and reproductive development to simulated environmental change in sub-arctic and high arctic plants. Oikos 1993, 67, 490–502. [Google Scholar] [CrossRef]
- Chapin, F.S.; Shaver, G.R. Physiological and growth responses of arctic plants to a field experiment simulating climatic change. Ecology 1996, 77, 822–840. [Google Scholar] [CrossRef]
- Stenseth, N.C.; Mysterud, A. Climate, changing phenology, and other life history traits: Nonlinearity and match-mismatch to the environment. Proc. Natl. Acad. Sci. USA 2002, 99, 13379–13381. [Google Scholar] [CrossRef] [Green Version]
- Visser, M.E.; Both, C. Shifts in phenology due to global climate change: The need for a yardstick. Proc. R. Soc. Lond. Biol. Sci. 2005, 272, 2561–2569. [Google Scholar] [CrossRef] [PubMed]
- Hooper, D.U. The role of complementarity and competition in ecosystem responses to variation in plant diversity. Ecology 1998, 79, 704–719. [Google Scholar] [CrossRef]
- Church, G. Trees and Shrubs for Flowers, (The Woody Plant); A Fireflay Book: Richmond Hill, ON, Canada, 2020; p. 160. [Google Scholar]
- Parmesan, C.; Yohe, G. A globally coherent fingerprint of climate change impacts across natural systems. Nature 2003, 421, 37–424. [Google Scholar] [CrossRef] [PubMed]
- Root, T.L.; Price, J.T.; Hall, K.R.; Schneider, S.H.; Rosenzweig, C.; Pounds, J.A. Fingerprints of global warming on wild animals and plants. Nature 2003, 421, 57–60. [Google Scholar] [CrossRef]
- Hoyle, H.; Hitchmough, J.; Jorgensen, A. All about the ‘wow factor’? The relationships between aesthetics, restorative effect and perceived biodiversity in designed urban planting. Landsc. Urban Plan. 2017, 164, 109–123. [Google Scholar] [CrossRef]
- Clark, N.E.; Lovell, R.; Wheeler, B.W.; Higgins, S.L.; Depledge, M.H.; Norris, K. Biodiversity, cultural pathways, and human health: A framework. Trends Ecol. Evol. 2014, 29, 198–204. [Google Scholar] [CrossRef] [Green Version]
- Hoyle, H.; Norton, B.; Dunnett, N.; Richards, P.; Russell, J.; Warren, P. Plant species or flower colour diversity? Identifying the drivers of public and invertebrate response to designed annual meadows. Landsc. Urban. Plan. 2018, 180, 103–113. [Google Scholar] [CrossRef]
- Menzel, A. Plant phenological anomalies in Germany and their relation to air temperature and NAO. Clim. Chang. 2003, 57, 243–263. [Google Scholar] [CrossRef]
- Fitter, A.H.; Fitter, R.S.R. Rapid changes in flowering time in British plants. Science 2002, 296, 1689–1691. [Google Scholar] [CrossRef]
- Sparks, T.H.; Menzel, A. Observed changes in seasons: An overview. Int. J. Climatol. 2002, 22, 1715–1725. [Google Scholar] [CrossRef]
- Dunne, J.; Harte, J.; Taylor, K. Subalpine meadow flowering phenology responses to climate change: Integrating experimental and gradient methods. Ecol. Monogr. 2003, 73, 69–86. [Google Scholar] [CrossRef]
- Arft, A.M.; Walker, M.; Gurevitch, J.; Alatalo, J.; Bret-Harte, M.; Dale, M.; Diemer, M.; Gugerli, F.; Henry, G.; Jones, M.; et al. Responses of tundra plants to experimental warming: Meta-analysis of the international tundra experiment. Ecol. Monogr. 1999, 69, 491–511. [Google Scholar] [CrossRef]
- Price, M.V.; Waser, N.M. Effects of experimental warming on plant reproductive phenology in a subalpine meadow. Ecology 1998, 79, 1261–1271. [Google Scholar] [CrossRef]
- Menzel, A.; Sparks, T.; Estrella, N.; Koch, E.; Aasa, A.; Ahas, R.; Alm-Kübler, K.; Bissolli, P.; Braslavska, O.; Briede, A.; et al. European phenological response to climate change matches the warming pattern. Glob. Chang. Biol. 2006, 12, 1969–1976. [Google Scholar] [CrossRef]
- Meehl, G.A.; Stocker, T.; Collins, W.; Friedlingstein, P.; Gaye, A.; Gregory, J.M.; Kitoh, A.; Knutti, R.; Murphy, J.M.; Noda, A.; et al. Global climate projections. In Climate Change 2007: The Physical Science Basis; Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Solomon, S., Qin, D., Manning, M., Chen, Z., Marquis, M., Averyt, K.B., Tignor, M., Miller, H.L., Eds.; Cambridge University Press: Cambridge, UK, 2007; pp. 747–846. [Google Scholar]
- Kerby, J.T.; Wilmers, C.C.; Post, E. Climate change, phenology and the nature of consumer–resource interactions: Advancing the match/mismatch hypothesis. In Trait-Mediated Indirect Interactions: Ecological and Evolutionary Perspectives; Ohgushi, T., Schmitz, O.J., Holt, R.D., Eds.; Cambridge University Press: Cambridge, UK, 2012; pp. 508–525. [Google Scholar]
- Thackeray, S.J.; Henrys, P.A.; Hemming, D.; Bell, J.R.; Botham, M.S.; Burthe, S.; Helaouet, P.; Johns, D.G.; Jones, I.D.; Leech, D.I.; et al. Phenological sensitivity to climate across taxa and trophic levels. Nature 2016, 535, 241–245. [Google Scholar] [CrossRef] [Green Version]
- Donoso, I.; Stefanescu, C.; Martínez-Abraín, A.; Traveset, A. Phenological asynchrony in plant-butterfly interactions associated with climate: A communitywide perspective. Oikos 2016, 125, 1434–1444. [Google Scholar] [CrossRef] [Green Version]
- Iler, A.M.; CaraDonna, P.J.; Forrest, J.R.K.; Post, E. Demographic consequences of phenological shifts in response to climate change. Ann. Rev. Ecol. Evol. Syst. 2021, 52, 221–245. [Google Scholar] [CrossRef]
- Vitale, N.; Torretta, J.P.; Durante, S.; Basilio, A.; Vázquez, D.P. Similarities and differences in the realized niche of two allopatric populations of a solitary bee under environmental variability. Apidologie 2020, 51, 439–454. [Google Scholar] [CrossRef]
- Davis, M.A.; Chew, M.K.; Hobbs, R.J.; Lugo, A.E.; John, J.; Ewel, J.J.; Vermeij, G.; Brown, J.; Rosenzweig, M.; Gardener, M.; et al. Don’t judge species on their origins. Nature 2011, 474, 153–154. [Google Scholar] [CrossRef] [PubMed]
- Hitchmough, J.D. Exotic plants and plantings in the sustainable, designed landscape. Landsc. Urban Plan. 2011, 100, 380–382. [Google Scholar] [CrossRef]
- Salisbury, A.; Armitage, J.; Bostock, H.; Perry, J.; Tatchell, M.; Thompson, K. Enhancing gardens as habitats for flower-visiting aerial insects (pollinators): Should we plant native or exotic species? J. Appl. Ecol. 2015, 52, 1156–1164. [Google Scholar] [CrossRef]






























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Scientific Name | Common Name | Class * | n ** |
|---|---|---|---|---|
| 1. Univerzijade Street 24, Novi Beograd, Ledine | Ficus carica L. | Edible Fig | D | 4.608 |
| 2. Ušće Street, Ušće shopping mall | Yucca gloriosa L. | Spanish Dagger | M | 5.424 |
| 3. Nikola Pašić Square, Pionirski Park | Cornus alba L. | Tartarian Dogwood | D | 5.840 |
| 4. Beogradska Street, Tašmajdan Park | Cornus alba L. Cornus alba ‘Sibirica’ | Tartarian Dogwood Siberian Dogwood | D D | 5.840 5.008 |
| 5. Savska Street, Gazela Park | Philadelphus coronarius L. Forsythia × intermedia Zabel Syringa vulgaris L. | European Mock-Orange Border Forsythia Common Lilac | D D D | 5.296 5.424 5.440 |
| 6. Vojvode Mišića Boulevard, Topčider Park | Philadelphus coronarius L. | European Mock-Orange | D | 5.296 |
| 7. Mirosavljeva Street, Banovo brdo | Cornus alba ‘Sibirica’ Prunus laurocerasus ‘Zabeliana’ | Siberian Dogwood Zabel’s Laurel | D D | 5.008 5.840 |
| 8. Vinogradski venac, Cerak | Cornus sanguinea L. | Common Dogwood | D | 5.024 |
| 9. Obrenovački Road, Ada Ciganlija | Yucca gloriosa L. | Spanish Dagger | D | 5.424 |
| 10. Savska Street, Ostružnica | Amorpha fruticosa L. | Indigo Bush | D | 5.088 |
| 11. Ive Lole Ribara Street, Ostružnica | Lonicera periclymenum ‘Serotina’ Rosa rugosa Thunb. | Honeysuckle Rugosa Rose | D D | 5.068 5.440 |
| 12. Nebojšina Street, Park of St. Sava’s Temple | Jasminum nudiflorum Lindl. | Weeping Winter Јasmine | D | 5.792 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ocokoljić, M.; Petrov, D.; Galečić, N.; Skočajić, D.; Košanin, O.; Simović, I. Phenological Flowering Patterns of Woody Plants in the Function of Landscape Design: Case Study Belgrade. Land 2023, 12, 706. https://doi.org/10.3390/land12030706
Ocokoljić M, Petrov D, Galečić N, Skočajić D, Košanin O, Simović I. Phenological Flowering Patterns of Woody Plants in the Function of Landscape Design: Case Study Belgrade. Land. 2023; 12(3):706. https://doi.org/10.3390/land12030706
Chicago/Turabian StyleOcokoljić, Mirjana, Djurdja Petrov, Nevenka Galečić, Dejan Skočajić, Olivera Košanin, and Isidora Simović. 2023. "Phenological Flowering Patterns of Woody Plants in the Function of Landscape Design: Case Study Belgrade" Land 12, no. 3: 706. https://doi.org/10.3390/land12030706