
Current and Potential Future Distribution of Endemic Salvia ceratophylloides Ard. (Lamiaceae)

 ,
,  ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Species Occurrence Data
2.2. Environmental Variables
- SSP585: with an additional radiative forcing of 8.5 W/m2 by the year 2100;
- SSP370: with an additional radiative forcing of 7 W/m2 by the year 2100;
- SSP245: with an additional radiative forcer of 4.5 W/m2 by the year 2100;
- SSP126: with an additional radiative forcer of 2.6 W/m2 by the year 2100.
2.3. Model Construction
2.4. Distribution Maps: Visualisation and Analysis
3. Results
3.1. Natural Distribution Data
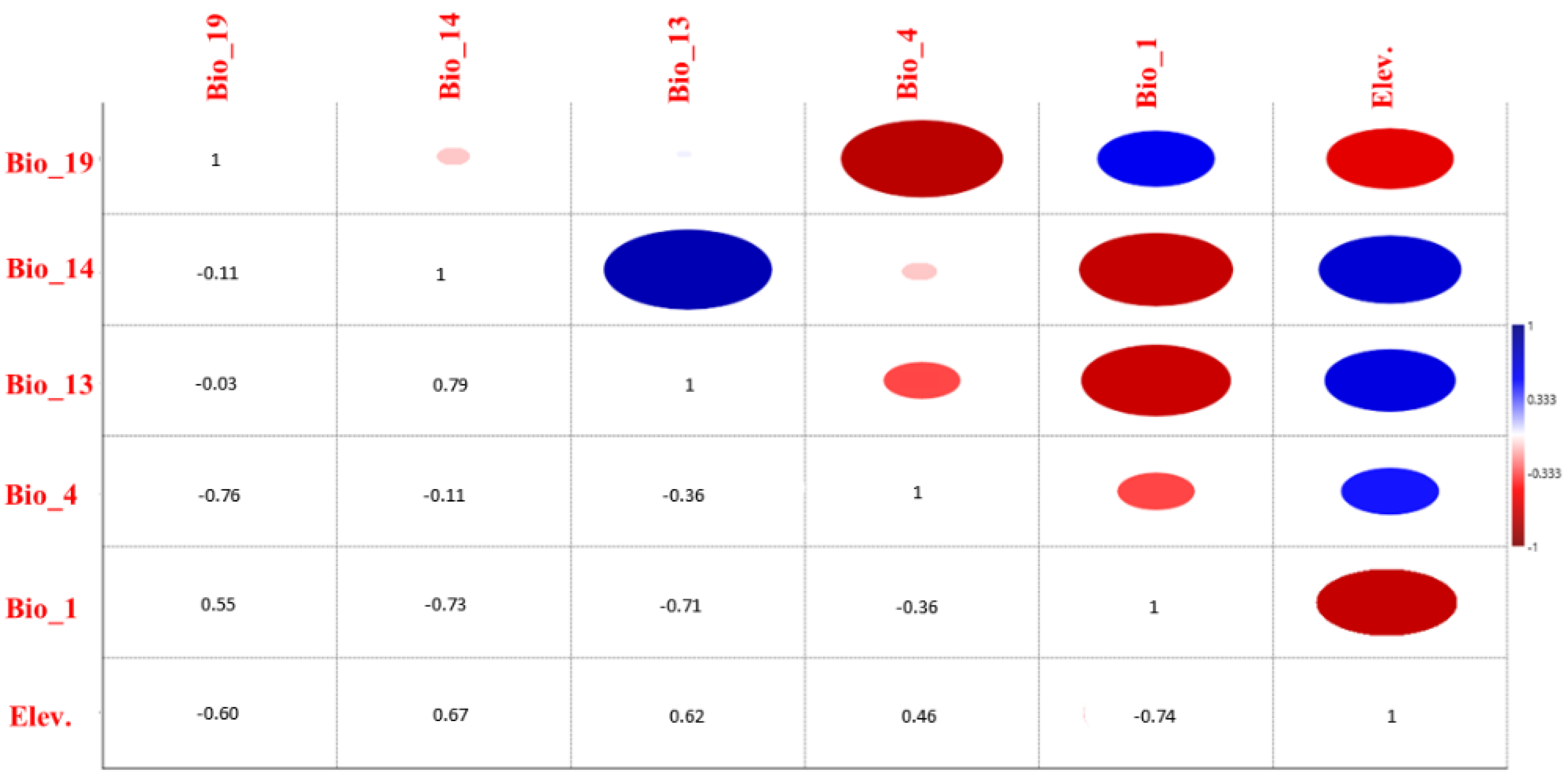
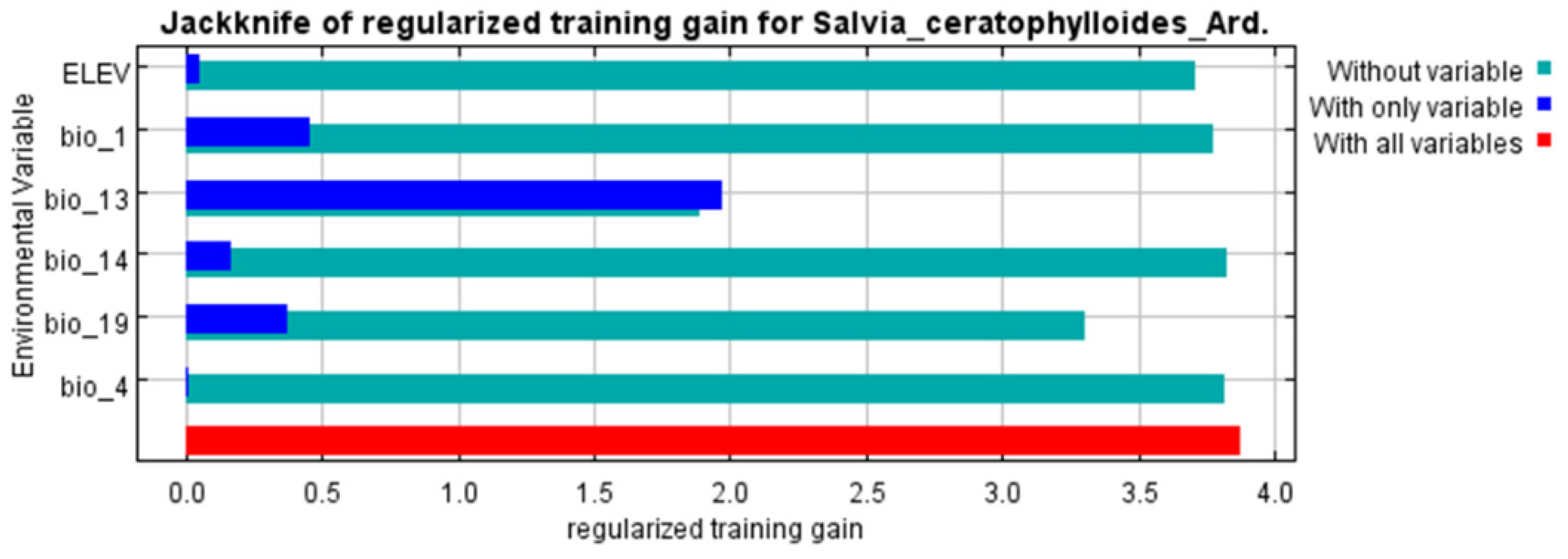
3.2. Analysis and Evaluation of Environmental Variables
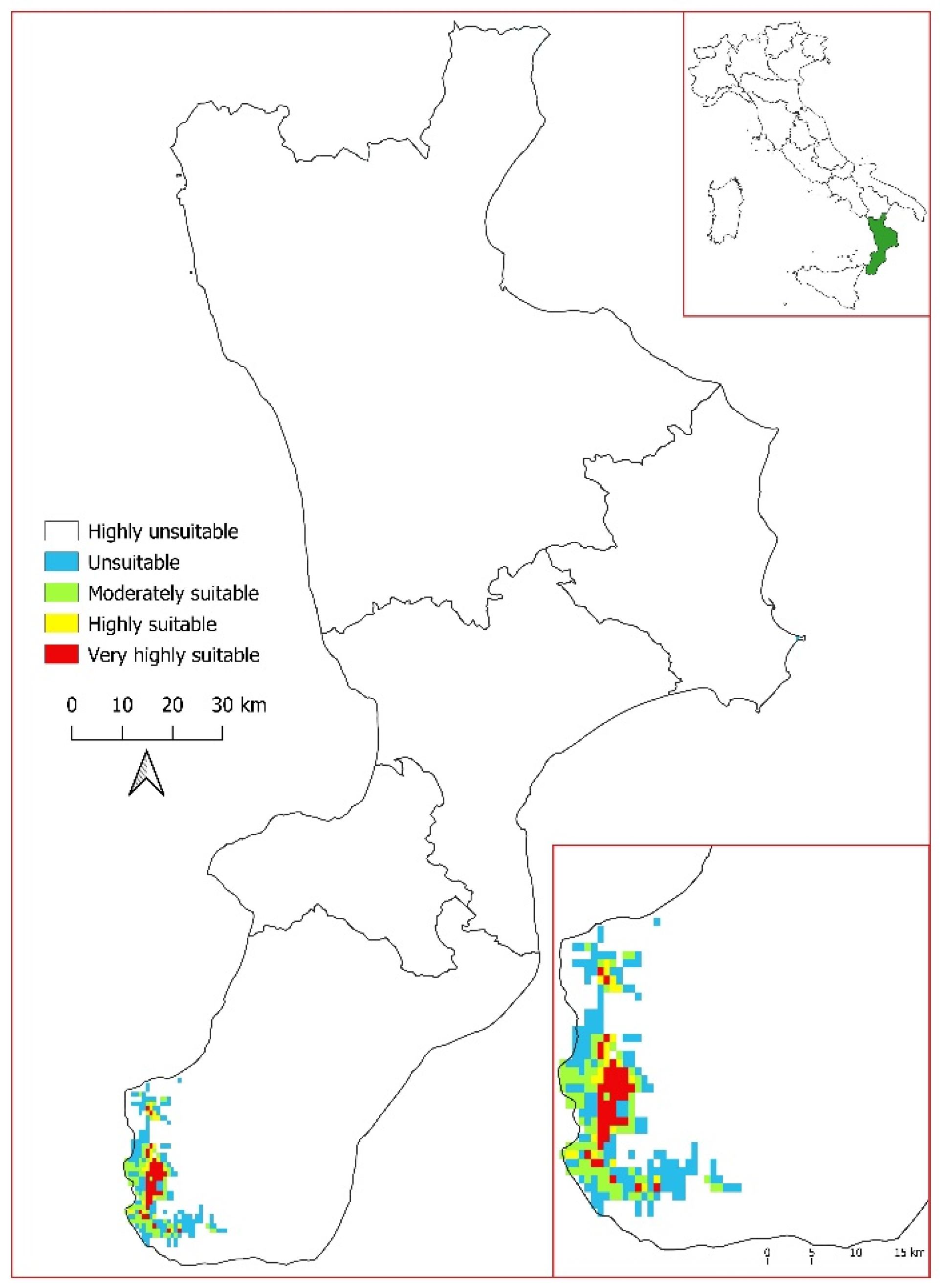
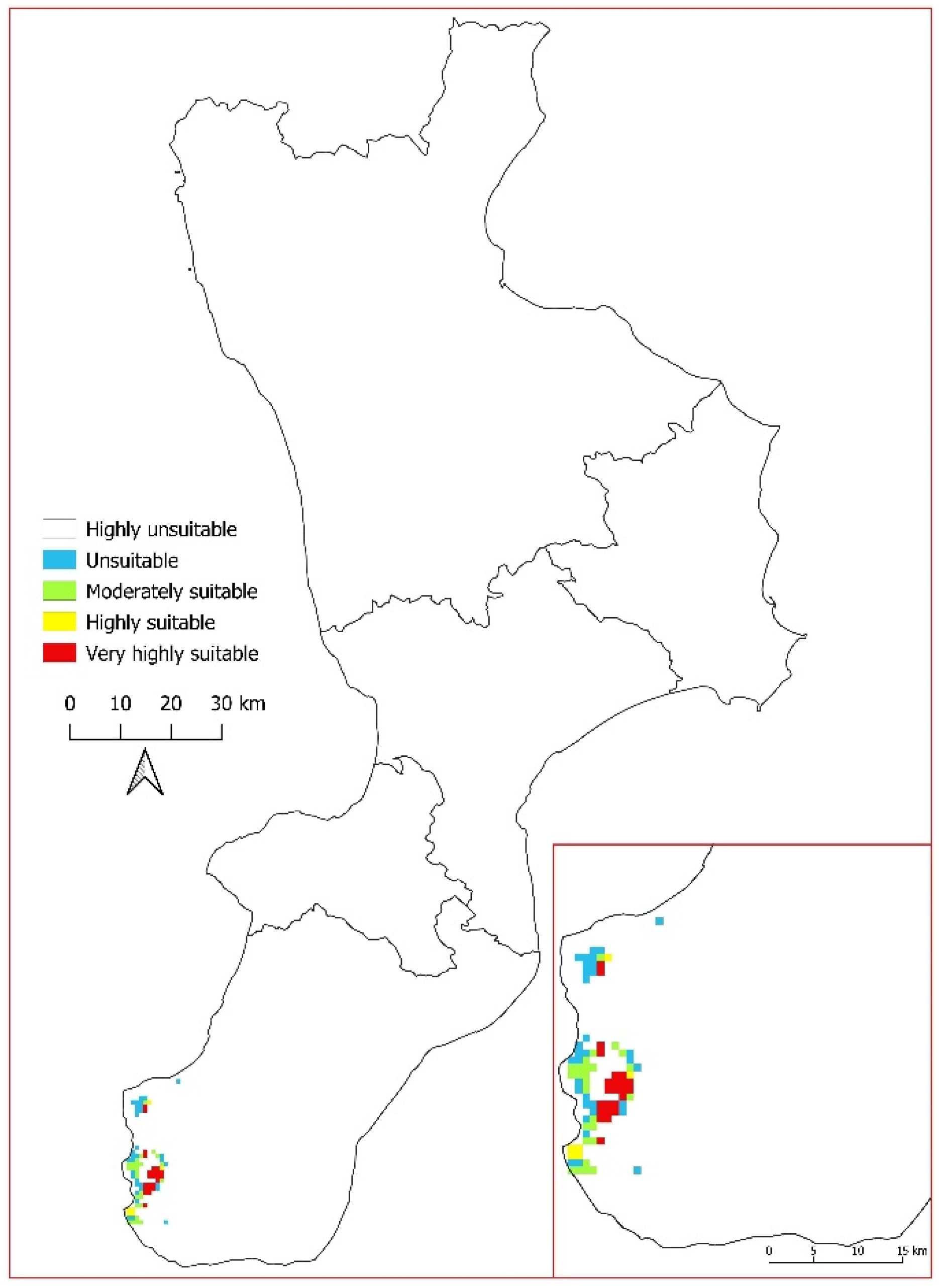
3.3. Current Potential Distribution of Salvia ceratophylloides
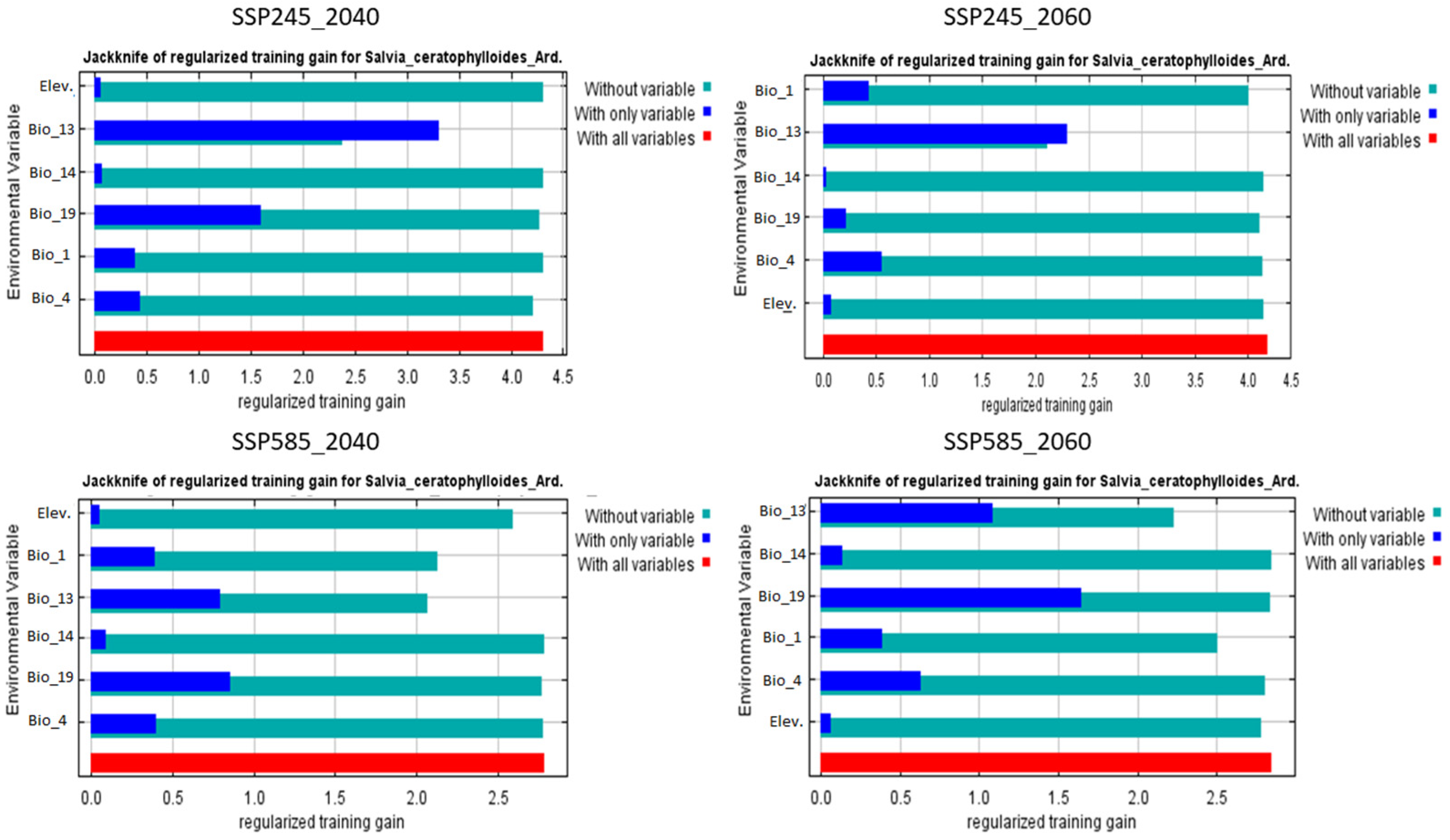
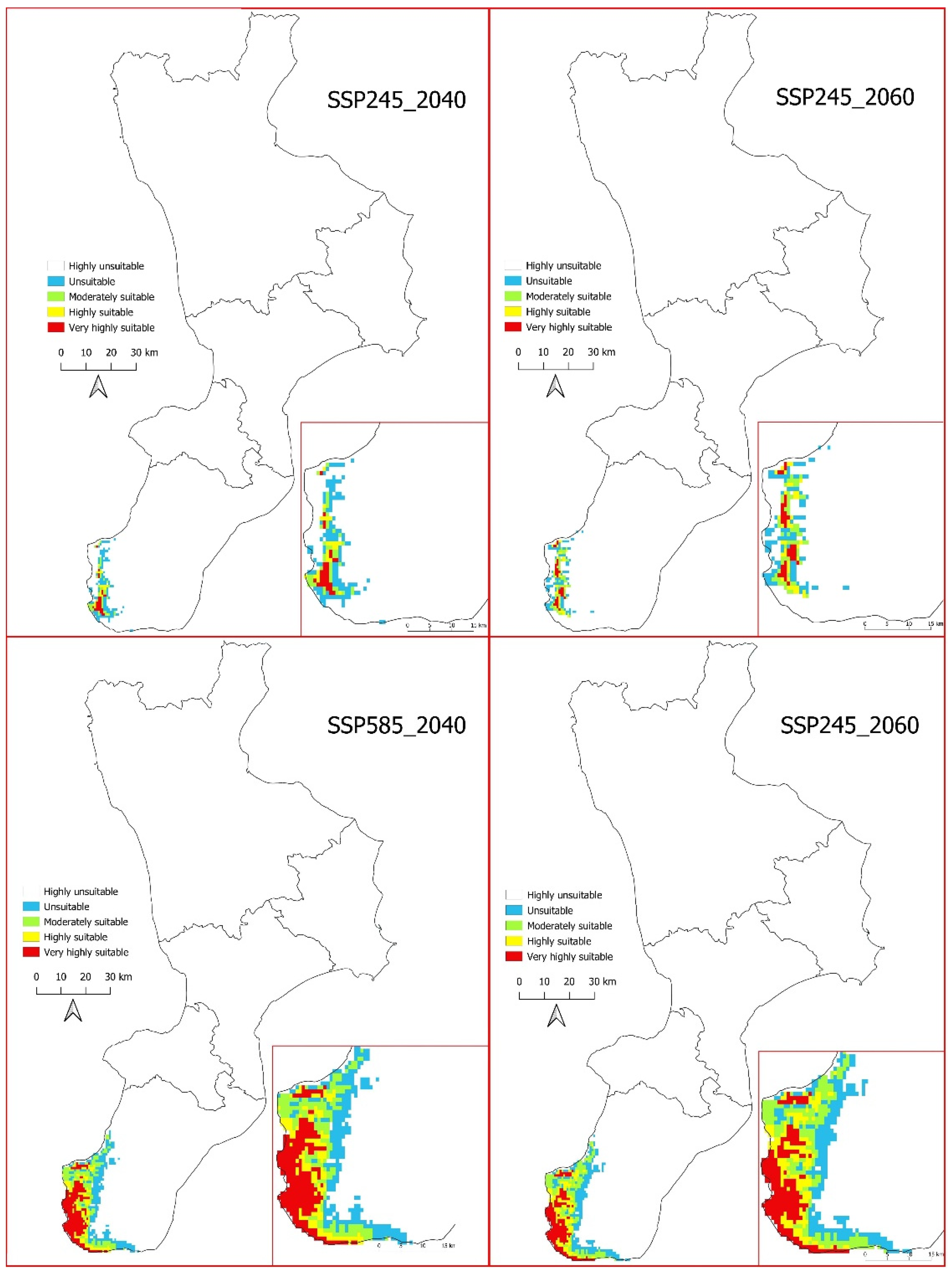
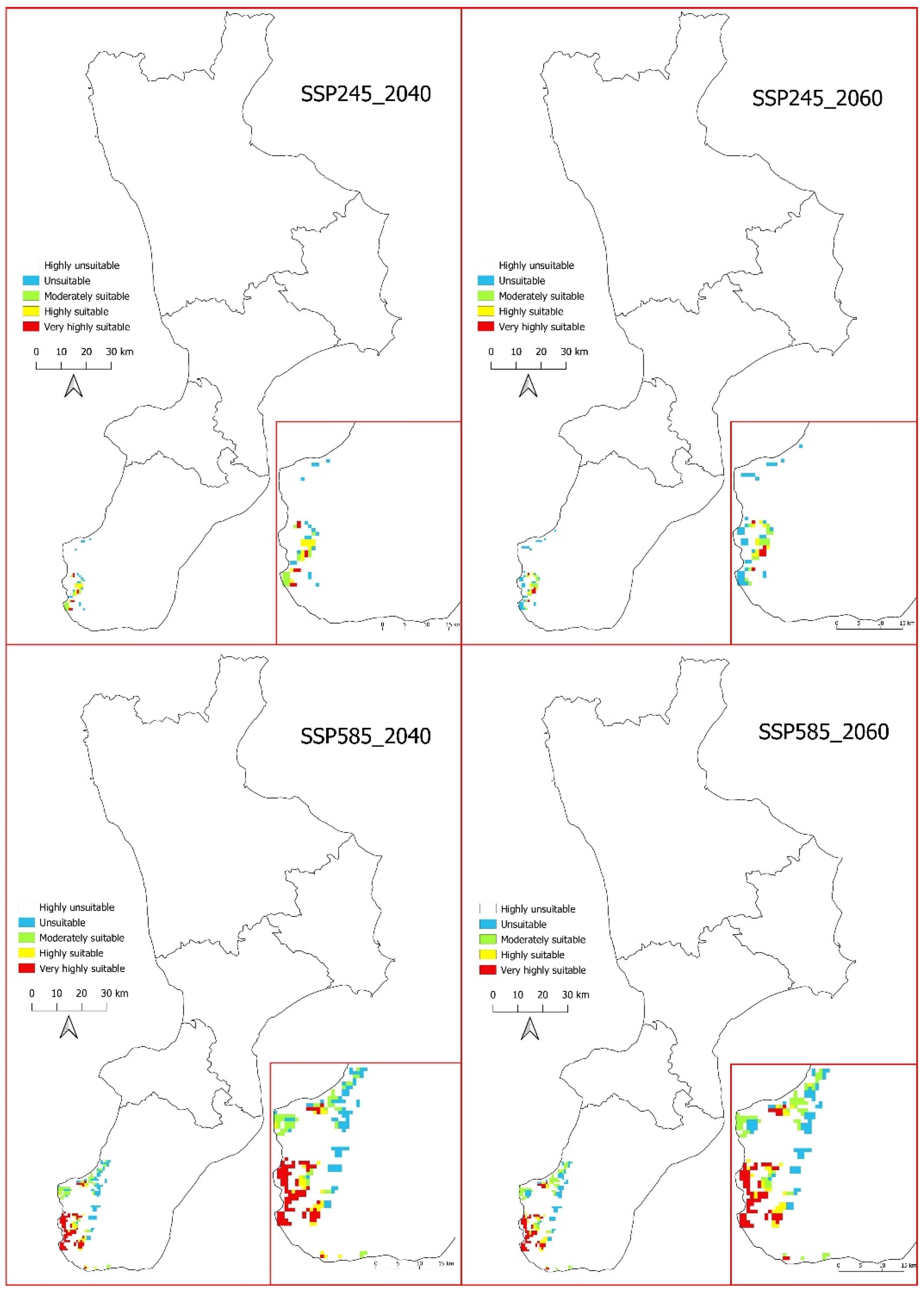
3.4. Future Potential Distribution of Salvia ceratophylloides
3.5. Real Distribution of Salvia ceratophylloides Analysed with Two Limiting Factors: Geology and Land Use
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Walther, G.R.; Post, E.; Convey, P.; Menzel, A.; Parmesan, C.; Beebee, T.J.; Fromentin, J.M.; Hoegh-Guldberg, O.; Bairlein, F. Ecological responses to recent climate change. Nature 2002, 416, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Thuiller, W.; Lavorel, S.; Araújo, M.B.; Sykes, M.T.; Prentice, I.C. Climate change threats to plant diversity in Europe. Proc. Natl. Acad. Sci. USA 2005, 102, 8245–8250. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Saiz, J.C.; Martínez García, F.; Gavilán, R.G. Plant Conservation in Spain: Strategies to halt the loss of plant diversity. Mediterr. Bot. 2018, 39, 65–66. [Google Scholar] [CrossRef]
- Wood, A.; Stedman-Edwards, P.; Mang, J. The Root Causes of Biodiversity Loss; Routledge: London, UK, 2000; p. 416. [Google Scholar]
- Del Río, S.; Canas, R.; Cano, E.; Cano-Ortiz, A.; Musarella, C.; Pinto-Gomes, C.; Penas, A. Modelling the impacts of climate change on habitat suitability and vulnerability in deciduous forests in Spain. Ecol. Indic. 2021, 131, 108202. [Google Scholar] [CrossRef]
- Dineva, S. Applying Artificial Intelligence (AI) for Mitigation Climate Change Consequences of the Natural Disasters. Res. J. Ecol. Environ. Sci. 2022, 1, 1–8. [Google Scholar] [CrossRef]
- Içik, K. Rare and endemic species: Why are they prone to extinction? Turk. J. Bot. 2011, 35, 411–417. [Google Scholar]
- Signorino, G.; Cannavò, S.; Crisafulli, A.; Musarella, C.M.; Spampinato, G. Fagonia cretica L. Inf. Bot. Ita. 2011, 43, 397–399. [Google Scholar]
- Spampinato, G.; Musarella, C.M.; Cano-Ortiz, A.; Signorino, G. Habitat, occurrence and conservation status of the Saharo-Macaronesian and Southern-Mediterranean element Fagonia cretica L. (Zygophyllaceae) in Italy. J. Arid. Land 2018, 10, 140–151. [Google Scholar] [CrossRef] [Green Version]
- Abdelaal, M.; Fois, M.; Fenu, G.; Bacchetta, G. Using MaxEnt modeling to predict the potential distribution of the endemic plant Rosa arabica Crép. in Egypt. Ecol. Inform. 2019, 50, 68–75. [Google Scholar] [CrossRef]
- Carmona, E.C.; Ortiz, A.C.; Musarella, C.M. Introductory Chapter: Endemism as a Basic Element for the Conservation of Species and Habitats. In Endemic Species [Internet]; Carmona, E.C., Musarella, C.M., Ortiz, A.C., Eds.; IntechOpen: London, UK, 2019; Available online: https://www.intechopen.com/chapters/65963 (accessed on 16 October 2022).
- Orsenigo, S.; Bernardo, L.; Cambria, S.; Gargano, D.; Laface, V.L.A.; Musarella, C.M.; Passalacqua, N.G.; Spampinato, G.; Tavilla, G.; Fenu, G. Global and Regional IUCN Red List Assessments: 9. Ital. Botanist. 2020, 9, 111–123. [Google Scholar] [CrossRef]
- Laface, V.L.A.; Musarella, C.M.; Sorgonà, A.; Spampinato, G. Analysis of the population structure and dynamic of endemic Salvia ceratophylloides Ard. (Lamiaceae). Sustainability 2022, 14, 10295. [Google Scholar] [CrossRef]
- Caruso, G. Anthyllis hermanniae L. subsp. brutia Brullo & Giusso (Fabaceae): Population survey and conservation tasks. Res. J. Ecol. Environ. Sci. 2022, 2, 92–102. [Google Scholar] [CrossRef]
- Jones, P.D.; Osborn, T.J.; Briffa, K.R. The evolution of climate over the last millennium. Science 2001, 292, 662–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xoplaki, E.; Gonzales-Rouco, F.J.; Luterbacher, J.; Wanner, H. Mediterranean summer air temperature variability and its connection to the large-scale atmospheric circulation and SSTs. Clim. Dyn. 2003, 20, 723–739. [Google Scholar] [CrossRef]
- Solomon, S.; Qin, D.; Manning, M.; Alley, R.B.; Berntsen, T.; Bindoff, N.L.; Chen, Z.; Chidthaisong, A.; Gregory, J.M.; Hegerl, G.C.; et al. Technical summary. In Climate Change 2007: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Solomon, S., Qin, D., Manning, M., Chen, Z., Marquis, M., Averyt, K., Tignor, M., Miller, H., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2007. [Google Scholar]
- Mariotti, A.; Pan, Y.; Zeng, N.; Alessandri, A. Long-term climate change in the Mediterranean region in the midst of decadal variability. Clim. Dyn. 2015, 44, 1437–1456. [Google Scholar] [CrossRef]
- Intergovernmental Panel on Climate Change (IPCC). Climate Change 2022: Mitigation of Climate Change. In Contribution of Working Group III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Shukla, P.R., Skea, J., Slade, R., al Khourdajie, A., van Diemen, R., McCollum, D., Pathak, M., Some, S., Vyas, P., Fradera, R., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2022. [Google Scholar]
- Laface, V.L.A.; Musarella, C.M.; Ortiz, A.C.; Canas, R.Q.; Cannavò, S.; Spampinato, G. Three New Alien Taxa for Europe and a Chorological Update on the Alien Vascular Flora of Calabria (Southern Italy). Plants 2020, 9, 1181. [Google Scholar] [CrossRef]
- Musarella, C.M.; Stinca, A.; Cano-Ortíz, A.; Laface, V.L.A.; Petrilli, R.; Esposito, A.; Spampinato, G. New data on the alien vascular flora of Calabria (Southern Italy). Ann. Bot. 2020, 10, 55–66. [Google Scholar] [CrossRef]
- Musarella, C.M. Solanum torvum Sw. (Solanaceae): A new alien species for Europe. Genet. Resour. Crop Evol. 2020, 67, 515–522. [Google Scholar] [CrossRef]
- Raposo, M.A.M.; Pinto Gomes, C.J.; Nunes, L.J.R. Evaluation of Species Invasiveness: A Case Study with Acacia dealbata Link. on the Slopes of Cabeça (Seia-Portugal). Sustainability 2021, 13, 11233. [Google Scholar] [CrossRef]
- Zhang, Y.; Tang, J.; Ren, G.; Zhao, K.; Wang, X. Global potential distribution prediction of Xanthium italicum based on Maxent model. Sci. Rep. 2021, 11, 16545. [Google Scholar] [CrossRef]
- de Carvalho, C.A.; Raposo, M.; Pinto-Gomes, C.; Matos, R. Native or Exotic: A Bibliographical Review of the Debate on Ecological Science Methodologies: Valuable Lessons for Urban Green Space Design. Land 2022, 11, 1201. [Google Scholar] [CrossRef]
- Spampinato, G.; Laface, V.L.A.; Posillipo, G.; Cano-Ortiz, A.; Quinto-Canas, R.; Musarella, C.M. Alien flora in Calabria (Southern Italy): An updated checklist. Biol. Invasions 2022, 24, 2323–2334. [Google Scholar] [CrossRef]
- Ighbareyeh, J.M.H.; Suliemieh, A.A.-R.A.; Sheqwarah, M.; Cano-Ortiz, A.; Carmona, E.C. Flora and Phytosociological of Plant in Al-Dawaimah of Palestine. Res. J. Ecol. Environ. Sci. 2022, 2, 58–91. [Google Scholar] [CrossRef]
- Molina-Venegas, R.; Aparicio, A.; Lavergne, S.; Arroyo, J. Climatic and topographical correlates of plant palaeo- and neoendemism in a Mediterranean biodiversity hotspot. Ann. Bot. 2017, 119, 229–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rundel, P.W.; Arroyo, M.T.K.; Cowling, R.M.; Keeley, J.E.; Lamont, B.B.; Vargas, P. Mediterranean biomes: Evolution of their vegetation, floras, and climate. Annu. Rev. Ecol. Evol. Syst. 2016, 47, 383–407. [Google Scholar] [CrossRef]
- Musarella, C.M.; Tripodi, G. La flora della rupe e dei ruderi di Pentidattilo (Reggio Calabria). Inform. Bot. Ital. 2004, 36, 3–12. [Google Scholar]
- Pecl, G.T.; Araújo, M.B.; Bell, J.D.; Blanchard, J.; Bonebrake, T.C.; Chen, I.C.; Clark, T.D.; Colwell, R.K.; Danielsen, F.; Evengård, B.; et al. Biodiversity redistribution under climate change: Impacts on ecosystems and human well-being. Science 2017, 355, eaai9214. [Google Scholar] [CrossRef]
- Qin, A.; Liu, B.; Guo, Q.; Bussmann, R.W.; Ma, F.; Jian, Z.; Xu, G.; Pei, S. Maxent modeling for predicting impacts of climate change on the potential distribution of Thuja sutchuenensis Franch., an extremely endangered conifer from southwestern China. Glob. Ecol. Conserv. 2017, 10, 139–146. [Google Scholar] [CrossRef]
- del Río, S.; Álvarez-Esteban, R.; Cano, E.; Pinto-Gomes, C.; Penas, Á. Potential Impacts of Climate Change on Habitat Suitability of Fagus sylvatica L. For. Spain Plant Biosyst. 2018, 152, 1205–1213. [Google Scholar] [CrossRef]
- Rus, J.D.; Ramírez-Rodríguez, R.; Amich, F.; Melendo-Luque, M. Habitat Distribution Modelling, under the Present Climatic Scenario, of the Threatened Endemic Iberian Delphinium fissum subsp. sordidum (Ranunculaceae) and Implications for Its Conservation. Plant Biosyst. 2018, 152, 891–900. [Google Scholar] [CrossRef]
- Anand, V.; Oinam, B.; Singh, H. Predicting the current and future potential spatial distribution of endangered Rucervus eldii eldii (Sangai) using MaxEnt model. Environ. Monit. Assess. 2021, 193, 147. [Google Scholar] [CrossRef]
- Rojo, J.; Fernández-González, F.; Lara, B.; Bouso, V.; Crespo, G.; Hernández-Palacios, G.; Rodríguez-Rojo, M.P.; Rodríguez-Torres, A.; Smith, M.; Pérez-Badia, R. The effects of climate change on the flowering phenology of alder trees in Southwestern Europe. Mediterr. Bot. 2021, 42, e67360. [Google Scholar] [CrossRef]
- Wilson, C.D.; Roberts, D.; Reid, N. Applying species distribution modelling to identify areas of high conservation value for endangered species: A case study using Margaritifera margaritifera (L.). Biol. Conserv. 2011, 144, 821–829. [Google Scholar] [CrossRef]
- Reich, D.; Flatscher, R.; Pellegrino, G.; Hülber, K.; Wessely, J.; Gattringer, A.; Greimler, J. Biogeography of amphi-adriatic Gentianella crispata (Gentianaceae): A northern refugium and recent trans-adriatic migration. Plant Biosyst. 2022, 156, 754–768. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of maxent for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Soilhi, Z.; Sayari, N.; Benalouache, N.; Mekki, M. Predicting current and future distributions of Mentha pulegium L. in Tunisia under climate change conditions, using the MaxEnt model. Ecol. Inform. 2022, 68, 101533. [Google Scholar] [CrossRef]
- Thakur, S.; Rai, I.D.; Singh, B.; Dutt, H.C.; Musarella, C.M. Predicting the Suitable Habitats of Elwendia persica in Indian Himalayan Region (IHR). Plant Biosyst. 2023, in press.
- Gebrewahid, Y.; Abrehe, S.; Meresa, E.; Eyasu, G.; Abay, K.; Gebreab, G.; Kidanemariam, K.; Adissu, G.; Abreha, G.; Darcha, G. Current and future predicting potential areas of Oxytenanthera abyssinica (A. Richard) using MaxEnt model under climate change in Northern Ethiopia. Ecol. Process. 2020, 9, 6. [Google Scholar] [CrossRef] [Green Version]
- Bhandari, M.S.; Shankhwar, R.; Maikhuri, S.; Pandey, S.; Meena, R.K.; Ginwal, H.S.; Kant, R.; Rawat, P.S.; Martins-Ferreira, M.A.; Silveira, L.H. Prediction of ecological and geological niches of Salvadora oleoides in arid zones of India: Causes and consequences of global warming. Arab. J. Geosci. 2021, 14, 524. [Google Scholar] [CrossRef]
- Dai, X.; Wu, W.; Ji, L.; Tian, S.; Yang, B.; Guan, B.; Wu, D. MaxEnt model-based prediction of potential distributions of Parnassia wightiana (Celastraceae) in China. Biodivers. Data J. 2022, 10, e81073. [Google Scholar] [CrossRef]
- Khan, A.M.; Li, Q.; Saqib, Z.; Khan, N.; Habib, T.; Khalid, N.; Majeed, M.; Tariq, A. MaxEnt Modelling and Impact of Climate Change on Habitat Suitability Variations of Economically Important Chilgoza Pine (Pinus gerardiana Wall.) in South Asia. Forests 2022, 13, 715. [Google Scholar] [CrossRef]
- Zhang, L.; Zhu, L.; Li, Y.; Zhu, W.; Chen, Y. Maxent Modelling Predicts a Shift in Suitable Habitats of a Subtropical Evergreen Tree (Cyclobalanopsis glauca (Thunberg) Oersted) under Climate Change Scenarios in China. Forests 2022, 13, 126. [Google Scholar] [CrossRef]
- Parveen, S.; Kaur, S.; Baishya, R.; Goel, S. Predicting the potential suitable habitats of genus Nymphaea in India using MaxEnt modeling. Environ. Monit. Assess. 2022, 194, 853. [Google Scholar] [CrossRef]
- Khajoei Nasab, F.; Mehrabian, A.; Mostafavi, H.; Neemati, A. The influence of climate change on the suitable habitats of Allium species endemic to Iran. Environ. Monit. Assess. 2022, 194, 169. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stohlgren, T.J. Maxent modeling for predicting suitable habitat for threatened and endangered tree Canacomyrica monticola in New Caledonia. J. Ecol. Nat. Environ. 2009, 1, 94–98. [Google Scholar]
- Marcer, A.; Sáez, L.; Molowny-Horas, R.; Pons, X.; Pino, J. Using species distribution modelling to disentangle realised versus potential distributions for rare species conservation. Biol. Conserv. 2013, 166, 221–230. [Google Scholar] [CrossRef]
- Shcheglovitova, M.; Anderson, R.P. Estimating optimal complexity for ecological niche models: A jackknife approach for species with small sample sizes. Ecol. Model. 2013, 269, 9–17. [Google Scholar] [CrossRef]
- Booth, T.H. Why understanding the pioneering and continuing contributions of BIOCLIM to species distribution modelling is important. Austral Ecol. 2018, 43, 852–860. [Google Scholar] [CrossRef] [Green Version]
- Heywood, V.H.; Richardson, I.B.K. Labiatae. In Flora Europaea; Tutin, T.G., Heywood, V.H., Burges, N.A., Moore, D.M., Valentine, D.H., Walters, S.M., Webb, D.A., Eds.; Cambridge at the University Press: Cambridge, UK, 1972; Volume 3, pp. 126–192. [Google Scholar]
- Kadereit, J.W. The Families and Genera of Vascular Plants; Lamiales: Berlin, Germany, 2004; Volume VII. [Google Scholar]
- Perrino, E.V.; Valerio, F.; Gannouchi, A.; Trani, A.; Mezzapesa, G. Ecological and Plant Community Implication on Essential Oils Composition in Useful Wild Officinal Species: A Pilot Case Study in Apulia (Italy). Plants 2021, 10, 574. [Google Scholar] [CrossRef]
- Valerio, F.; Mezzapesa, G.N.; Ghannouchi, A.; Mondelli, D.; Logrieco, A.F.; Perrino, E.V. Characterization and Antimicrobial Properties of Essential Oils from Four Wild Taxa of Lamiaceae Family Growing in Apulia. Agronomy 2021, 11, 1431. [Google Scholar] [CrossRef]
- Cianfaglione, K.; Bartolucci, F.; Ciaschetti, G.; Conti, F.; Pirone, G. Characterization of Thymus vulgaris subsp. vulgaris Community by Using a Multidisciplinary Approach: A Case Study from Central Italy. Sustainability 2022, 14, 3981. [Google Scholar] [CrossRef]
- Peruzzi, L.; Conti, F.; Bartolucci, F. An inventory of vascular plants endemic to Italy. Phytotaxa 2014, 168, 1–75. [Google Scholar] [CrossRef]
- Bartolucci, F.; Peruzzi, L.; Galasso, G.; Albano, A.; Alessandrini, A.; Ardenghi, N.M.G.; Astuti, G.; Bacchetta, G.; Ballelli, S.; Banfi, E.; et al. An updated checklist of the vascular flora native to Italy. Plant Biosyst. 2018, 152, 179–303. [Google Scholar] [CrossRef]
- Pignatti, S. Flora d’Italia; Edagricole: Bologna, Italy, 1982; Volume 2, pp. 502–507. [Google Scholar]
- Pignatti, S. Flora d’Italia, 2nd ed.; Edagricole: Bologna, Italy, 2018; Volume 3, pp. 301–310. [Google Scholar]
- Spampinato, G. Guida alla flora dell’Aspromonte, 2nd ed.; Laruffa Editore: Reggio Calabria, Italy, 2014; pp. 38, 244–245. [Google Scholar]
- Spampinato, G.; Laface, V.L.A.; Ortiz, A.C.; Canas, R.Q.; Musarella, C.M. Salvia ceratophylloides Ard. (Lamiaceae): A rare endemic species of Calabria (Southern Italy). In Endemic Species; Cano Carmona, E., Musarella, C.M., Cano Ortiz, A., Eds.; IntechOpen: London, UK, 2019. [Google Scholar]
- Salmeri, C. Karyological data of some plant species native to South Italy. Flora Mediterr. 2019, 29, 334–340. [Google Scholar] [CrossRef]
- Tenore, M. Sylloge Plantarum Vascularium Florae Neapolitanae Hucusque Detectarum; Ex Typographia Fibreni: Neapoli, Greece, 1831; p. 1. [Google Scholar]
- Macchiati, C. Catalogo delle piante raccolte nei dintorni di Reggio Calabria dal settembre 1881 al febbraio 1883. Nuovo Giorn. Bot. Ital. 1884, 16, 59–100. [Google Scholar]
- Lacaita, C. Addenda et emendanda ad floram italicam. Bull. Soc. Bot. Ital. 1921, 28, 18–19. [Google Scholar]
- Lacaita, C. Piante italiane critiche o rare: 67. Salvia ceratophylloides Arduino. Nuovo Giorn. Bot. Ital. 1921, 28, 144–147. [Google Scholar]
- Conti, F.; Manzi, A.; Pedrotti, F. Liste Rosse Regionali delle Piante d’Italia; WWF Italia, Società Botanica Italiana: Camerino, Italy, 1997. [Google Scholar]
- Del Carratore, F.; Garbari, F. Il Gen. Salvia Sect. Plethiosphace (Lamiaceae) in Italia. Arch. Geobot. 2001, 7, 41–62. [Google Scholar]
- Scoppola, A.; Spampinato, G. Atlante delle specie a rischio di estinzione. Versione 1.0. CD-Rom enclosed to the volume. In Stato Delle Conoscenze Sulla Flora Vascolare d’Italia; Scoppola, A., Blasi, C., Eds.; Palombi Editori: Roma, Italy, 2005. [Google Scholar]
- Spampinato, G.; Crisafulli, A. Struttura delle popolazioni e sinecologia di Salvia ceratophylloides (Lamiaceae) specie endemica minacciata di estinzione, 56. In Book of Abstracts of 103° S.B.I. Congress; Università Mediterranea di Reggio Calabria: Reggio Calabria, Italy, 2008; pp. 17–19. [Google Scholar]
- Crisafulli, A.; Cannavò, S.; Maiorca, G.; Musarella, C.M.; Signorino, G.; Spampinato, G. Aggiornamenti floristici per la Calabria. Inform. Bot. Ital. 2010, 42, 431–442. [Google Scholar]
- Spampinato, G.; Crisafulli, A.; Marino, A.; Signorino, G. Salvia ceratophylloides Ard. Inf. Bot. Ital. 2011, 43, 381–458. [Google Scholar]
- Laface, V.L.A.; Musarella, C.M.; Spampinato, G. Conservation status of the Aspromontana flora: Monitoring and new stations of Salvia ceratophylloides Ard. (Lamiaceae) endemic species in Reggio Calabria (southern Italy). In Abstracts Book of 113°Congresso della Società Botanica Italiana (V International Plant Science Conference (IPSC); Società Botanica Italiana: Fisciano, Italy, 2018; pp. 12–15. ISBN 978-88-85915-22-0. [Google Scholar]
- International Union for Conservation of Nature (IUCN). IUCN Red List Categories and Criteria: Version 3.1, 2nd ed.; IUCN: Gland, Switzerland; Cambridge, UK, 2012; pp. iv + 32. ISBN 978-2-8317-1435-6. [Google Scholar]
- Cassa per il Mezzogiorno—Cassa per Opere Straordinarie di Pubblico Interesse nell’Italia Meridionale, Carta Geologica della Calabria; Cassa per il Mezzogiorno: Rome, Italy, 1967–1972.
- Pesaresi, S.; Galdenzi, D.; Biondi, E.; Casavecchia, S. Bioclimate of Italy: Application of the worldwide bioclimatic classification system. J. Maps 2014, 10, 538–553. [Google Scholar] [CrossRef]
- Abate, E.; Azzarà, M.; Trifilò, P. When Water Availability Is Low, Two Mediterranean Salvia Species Rely on Root Hydraulics. Plants 2021, 10, 1888. [Google Scholar] [CrossRef] [PubMed]
- Abate, E.; Nardini, A.; Petruzzellis, F.; Trifilò, P. Too dry to survive: Leaf hydraulic failure in two Salvia species can be predicted on the basis of water content. Plant Physiol. Biochem. 2021, 166, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Vescio, R.; Abenavoli, M.R.; Araniti, F.; Musarella, C.M.; Sofo, A.; Laface, V.L.A.; Spampinato, G.; Sorgonà, A. The Assessment and the Within-Plant Variation of the Morpho-Physiological Traits and VOCs Profile in Endemic and Rare Salvia ceratophylloides Ard. (Lamiaceae). Plants 2021, 10, 474. [Google Scholar] [CrossRef]
- QGIS 2022. QGIS Geographic Information System. Open Source Geospatial Foundation Project. Available online: http://qgis.osgeo.org (accessed on 10 October 2022).
- WordClim. Available online: https://www.worldclim.org/ (accessed on 10 October 2022).
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; et al. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Saupe, E.E.; Barve, V.; Myers, C.E.; Soberón, J.; Barve, N.; Hensz, C.M.; Peterson, A.T.; Owens, H.L.; Lira-Noriega, A. Variation in niche and distribution model performance: The need for a priori assessment of key causal factors. Ecol. Model. 2012, 237–238, 11–22. [Google Scholar] [CrossRef]
- Pearson, K. Notes on regression and inheritance in the case of two parents. Proc. R. Soc. Lond. 1895, 58, 240–242. [Google Scholar]
- Hammer, Ø. Past Software. In Natural History Museum; University of Oslo: Oslo, Norway, 2022. [Google Scholar]
- Quenouille, M.H. Approximate Tests of Correlation in Time Series. J. R. Stat. Soc. 1949, 11, 68–84. [Google Scholar]
- Castellana, S.; Martin, M.Á.; Solla, A.; Alcaide, F.; Villani, F.; Cherubini, M.; Neale, D.; Mattioni, C. Signatures of local adaptation to climate in natural populations of sweet chestnut (Castanea sativa Mill.) from southern Europe. Ann. For. Sci. 2021, 78, 1–21. [Google Scholar] [CrossRef]
- Abou-Shaara, H.F.; Darwish, A.A. Expected prevalence of the facultative parasitoid Megaselia scalaris of honey bees in Africa and the Mediterranean region under climate change conditions. Int. J. Trop. Insect Sci. 2021, 41, 3137–3145. [Google Scholar] [CrossRef]
- Phillips, S.J. Transferability, sample selection bias and background data in presence-only modeling: A response to Peterson et al. and (2007). Ecography 2008, 31, 272–278. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudik, M.; Schapire, R.E. Maxent Software for Modeling Species Niches and Distributions (Version 3.4.4). Available online: http://biodiversityinformatics.amnh.org/open_source/maxent/ (accessed on 11 October 2022).
- Fielding, A.H.; Bell, J.F. A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ. Conserv. 1997, 24, 38–49. [Google Scholar] [CrossRef]
- Warren, D.L.; Seifert, S.N. Ecological niche modeling in Maxent: The importance of model complexity and the performance of model selection criteria. Ecol. Appl. 2001, 21, 335–342. [Google Scholar] [CrossRef] [Green Version]
- Carta di Uso del Territorio della Regione Calabria. ARSAC. Available online: http://93.51.147.138/corine_land_cover.html (accessed on 11 October 2022).
- Bossard, M.; Feranec, J.; Otahel, J. CORINE Land Cover Technical Guide: Addendum 2000; European Environment Agency: Copenhagen, Denmark, 2000; Volume 40.
- European Forest Fire Informationon System (EFFIS). Available online: https://effis.jrc.ec.europa.eu/ (accessed on 14 October 2022).
- Canturk, U.; Kulaç, Ş. The effects of climate change scenarios on Tilia ssp. in Turkey. Environ. Monit. Assess. 2021, 193, 771. [Google Scholar] [CrossRef]
- Ma, B.; Sun, J. Predicting the distribution of Stipa purpurea across the Tibetan Plateau via the MaxEnt model. BMC Ecol. 2018, 18, 10. [Google Scholar] [CrossRef] [Green Version]
- Bonsignore, C.P.; Laface, V.L.A.; Vono, G.; Marullo, R.; Musarella, C.M.; Spampinato, G. Threats Posed to the Rediscovered and Rare Salvia ceratophylloides Ard. (Lamiaceae) by Borer and Seed Feeder Insect Species. Diversity 2021, 13, 33. [Google Scholar] [CrossRef]
- Olszewski, P.; Dyderski, M.K.; Dylewski, Ł.; Bogusch, P.; Schmid-Egger, C.; Ljubomirov, T.; Zimmermann, D.; Le Divelec, R.; Wiśniowski, B.; Twerd, L.; et al. European beewolf (Philanthus triangulum) will expand its geographic range as a result of climate warming. Reg. Environ. Chang. 2022, 22, 129. [Google Scholar] [CrossRef]
- Puchałka, R.; Dyderski, M.K.; Vítková, M.; Sádlo, J.; Klisz, M.; Netsvetov, M.; Prokopuk, Y.; Matisons, R.; Mionskowski, M.; Wojda, T.; et al. Black locust (Robinia pseudoacacia L.) range contraction and expansion in Europe under changing climate. Glob. Chang. Biol. 2021, 27, 1587–1600. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Description | Unit |
|---|---|---|
| Bio 1 | Annual Mean Temperature | °C |
| Bio 2 | Mean Diurnal Range (Mean of monthly (max temp–min temp)) | °C |
| Bio 3 | Isothermality (BIO2/BIO7) (*100) | % |
| Bio 4 | Temperature Seasonality (standard deviation *100) | % |
| Bio 5 | Max Temperature of Warmest Month | °C |
| Bio 6 | Min Temperature of Coldest Month | °C |
| Bio 7 | Temperature Annual Range (BIO5-BIO6) | °C |
| Bio 8 | Mean Temperature of Wettest Quarter | °C |
| Bio 9 | Mean Temperature of Driest Quarter | °C |
| Bio 10 | Mean Temperature of Warmest Quarter | °C |
| Bio 11 | Mean Temperature of Coldest Quarter | °C |
| Bio 12 | Annual Precipitation | mm |
| Bio 13 | Precipitation of Wettest Month | mm |
| Bio 14 | Precipitation of Driest Month | mm |
| Bio 15 | Precipitation Seasonality (Coefficient of Variation) | % |
| Bio 16 | Precipitation of Wettest Quarter | mm |
| Bio 17 | Precipitation of Driest Quarter | mm |
| Bio 18 | Precipitation of Warmest Quarter | mm |
| Bio 19 | Precipitation of Coldest Quarter | mm |
| Elev. | Elevation | meter |
| Slope | Slope | degree |
| Aspe. | Aspect | degree |
| Time | SSPs | Variable | Bio 1 | Bio 4 | Bio 13 | Bio 14 | Bio 19 | Elev. |
|---|---|---|---|---|---|---|---|---|
| Present time | Percent contribution (%) | 7.8 | 7.3 | 69.3 | 1 | 11.4 | 3.2 | |
| Permutation importance (%) | 13.3 | 1 | 66.9 | 0.8 | 9 | 9 | ||
| 2021/2040 | 245 | Percent contribution (%) | 7 | 5.5 | 86.3 | 0.1 | 0.9 | 0.2 |
| Permutation importance (%) | 0.1 | 1.1 | 97.8 | 0.2 | 0.6 | 0. | ||
| 585 | Percent contribution (%) | 24.4 | 15.4 | 40.7 | 0 | 3 | 16.5 | |
| Permutation importance (%) | 52.4 | 0 | 25 | 0.2 | 21.7 | 0.7 | ||
| 2041/2060 | 245 | Percent contribution (%) | 11.9 | 12.1 | 73.4 | 1 | 1.3 | 0.3 |
| Permutation importance (%) | 13.7 | 0.6 | 78.7 | 0.8 | 5.4 | 0.7 | ||
| 585 | Percent contribution (%) | 16.5 | 14.2 | 25.6 | 0.1 | 42.6 | 1 | |
| Permutation importance (%) | 15.4 | 1.5 | 76 | 0.1 | 0.8 | 6.2 |
| Time | SSP | Unit | Area tot. | ≥0.80 | 0.61–0.80 | 0.41–0.60 | 0.21–0.40 | ≤0.20 |
|---|---|---|---|---|---|---|---|---|
| Present time | km2 | 237.321 | 30.440 | 20.962 | 59.434 | 126.485 | 14,813.597 | |
| % | 1.58 | 0.20 | 0.14 | 0.39 | 0.84 | 98.42 | ||
| 2021/2040 | 245 | km2 | 153.986 | 25.020 | 18.258 | 26.326 | 84.382 | 14,896.932 |
| % | 1.02 | 0.17 | 0.12 | 0.17 | 0.56 | 98.98 | ||
| % inc./dec. | −35.11 | −17.81 | −12.90 | −55.71 | −33.29 | +0.56 | ||
| 585 | km2 | 633.513 | 129.708 | 94.667 | 171.816 | 237.322 | 14,417.405 | |
| % | 4.21 | 0.86 | 0.63 | 1.14 | 1.58 | 95.79 | ||
| % inc./dec. | +166.94 | +326.11 | +351.61 | +189.09 | +87.63 | −2.67 | ||
| 2041/2060 | 245 | km2 | 171.414 | 29.071 | 22.984 | 41.241 | 78.118 | 14,879.504 |
| % | 1.14 | 0.19 | 0.15 | 0.27 | 0.52 | 98.86 | ||
| % inc./dec. | −27.77 | −4.50 | +9.65 | −30.61 | −38.24 | +0.44 | ||
| 585 | km2 | 643.814 | 145.446 | 122.362 | 150.514 | 225.492 | 14,407.104 | |
| % | 4.28 | 0.97 | 0.81 | 1.00 | 1.50 | 95.72 | ||
| % inc./dec. | +171.28 | +377.81 | +483.73 | +153.25 | +78.28 | −2.74 |
| Time | SSP | Unit | Area tot. | ≥0.80 | 0.61–0.80 | 0.41–0.60 | 0.21–0.40 | ≤0.20 |
|---|---|---|---|---|---|---|---|---|
| Present time | km2 | 62.427 | 16.285 | 4.071 | 19.000 | 23.071 | 2066.204 | |
| % sub. | 2.93 | 0.77 | 0.19 | 0.89 | 1.08 | 97.07 | ||
| % reg. ter. | 0.41 | 0.11 | 0.03 | 0.13 | 0.15 | 13.73 | ||
| 2021/2040 | 245 | km2 | 31.892 | 5.428 | 9.500 | 5.428 | 11.536 | 2096.739 |
| % sub. | 1.50 | 0.25 | 0.45 | 0.25 | 0.54 | 98.50 | ||
| % reg. ter. | 0.21 | 0.04 | 0.06 | 0.04 | 0.08 | 13.93 | ||
| 585 | km2 | 187.960 | 56.320 | 17.642 | 37.321 | 76.677 | 1940.671 | |
| % sub. | 8.83 | 2.65 | 0.83 | 1.75 | 3.60 | 91.17 | ||
| % reg. ter. | 1.25 | 0.37 | 0.12 | 0.25 | 0.51 | 12.89 | ||
| 2041/2060 | 245 | km2 | 50.213 | 4.750 | 6.107 | 10.857 | 28.499 | 2078.418 |
| % sub. | 2.36 | 0.22 | 0.29 | 0.51 | 1.34 | 97.64 | ||
| % reg. ter. | 0.33 | 0.03 | 0.04 | 0.07 | 0.19 | 13.81 | ||
| 585 | km2 | 192.031 | 50.892 | 19.678 | 48.856 | 72.605 | 1936.600 | |
| % sub. | 9.02 | 2.39 | 0.92 | 2.30 | 3.41 | 90.98 | ||
| % reg. ter. | 1.28 | 0.34 | 0.13 | 0.32 | 0.48 | 12.87 |
| Time | Unit | Area tot. | ≥0.80 | 0.61–0.80 | 0.41–0.60 | 0.21–0.40 |
|---|---|---|---|---|---|---|
| Present time | km2 | 237.321 | 30.440 | 20.962 | 59.434 | 126.485 |
| Present time– | km2 | 183.295 | 26.476 | 16.972 | 40.732 | 99.115 |
| land use | % | 23 | 13 | 19 | 31 | 22 |
| Present time– | km2 | 62.427 | 16.285 | 4.071 | 19.000 | 23.071 |
| geological substrate | % | 74 | 47 | 81 | 68 | 82 |
| Present time– geological substrate– land use | km2 | 41.392 | 14.928 | 2.714 | 10.857 | 12.893 |
| % | 83 | 51 | 87 | 82 | 90 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laface, V.L.A.; Musarella, C.M.; Tavilla, G.; Sorgonà, A.; Cano-Ortiz, A.; Quinto Canas, R.; Spampinato, G. Current and Potential Future Distribution of Endemic Salvia ceratophylloides Ard. (Lamiaceae). Land 2023, 12, 247. https://doi.org/10.3390/land12010247
Laface VLA, Musarella CM, Tavilla G, Sorgonà A, Cano-Ortiz A, Quinto Canas R, Spampinato G. Current and Potential Future Distribution of Endemic Salvia ceratophylloides Ard. (Lamiaceae). Land. 2023; 12(1):247. https://doi.org/10.3390/land12010247
Chicago/Turabian StyleLaface, Valentina Lucia Astrid, Carmelo Maria Musarella, Gianmarco Tavilla, Agostino Sorgonà, Ana Cano-Ortiz, Ricardo Quinto Canas, and Giovanni Spampinato. 2023. "Current and Potential Future Distribution of Endemic Salvia ceratophylloides Ard. (Lamiaceae)" Land 12, no. 1: 247. https://doi.org/10.3390/land12010247