
Updating Distribution, Ecology, and Hotspots for Three Amphibian Species to Set Conservation Priorities in a European Glacial Refugium

 , , and
, , and 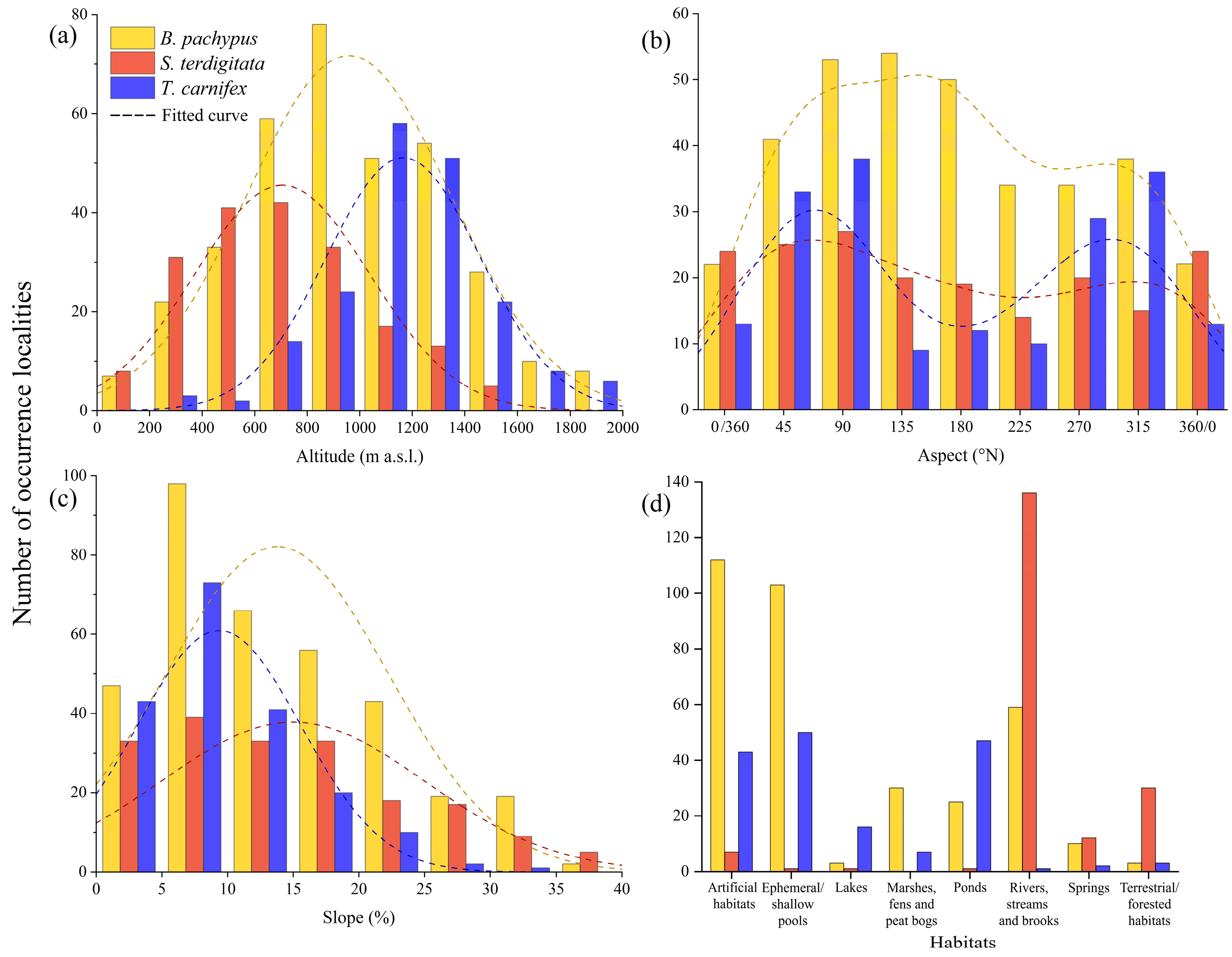{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Target Species and Study Area
2.2. Data Collection
2.3. Data Analysis
3. Results
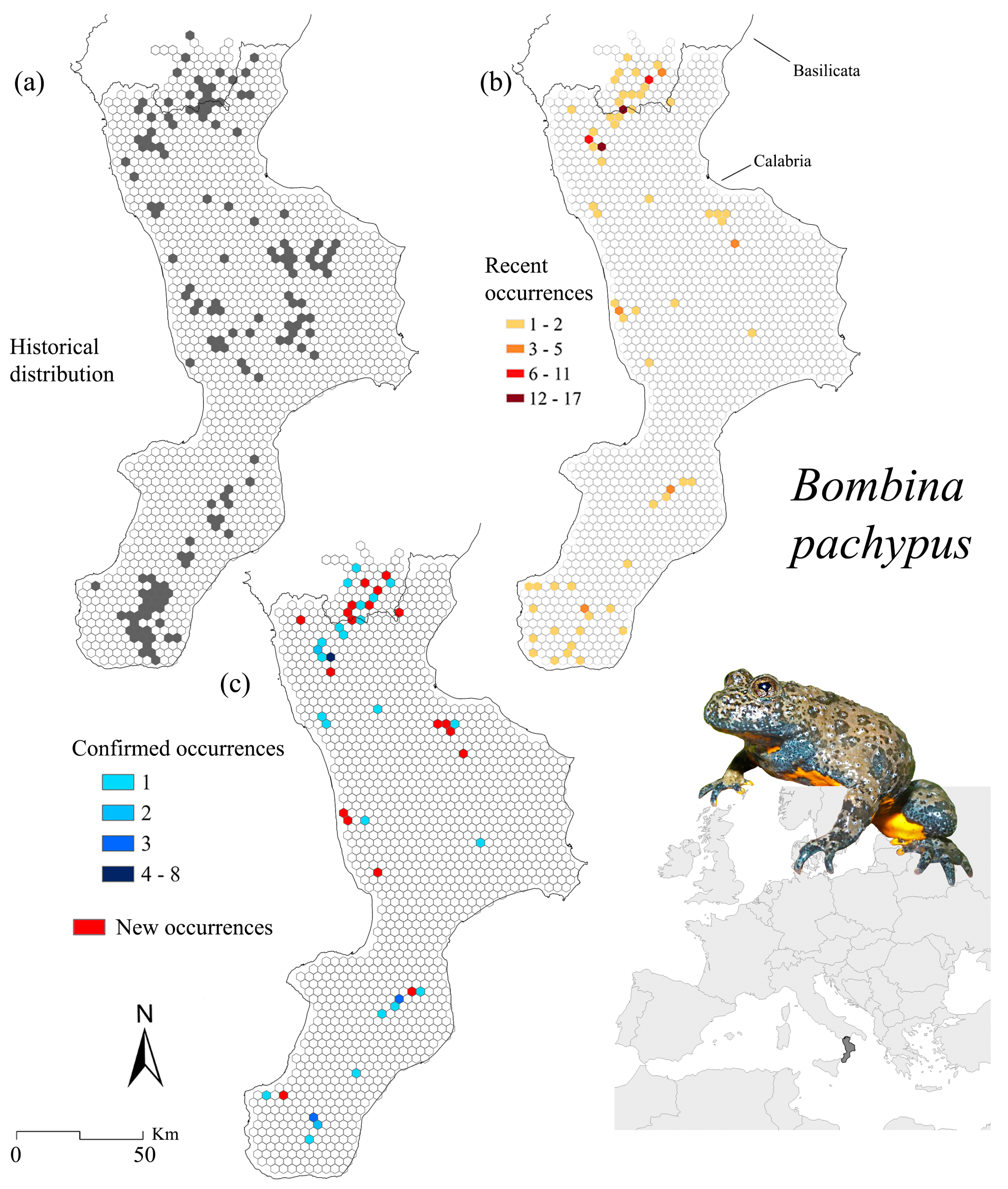
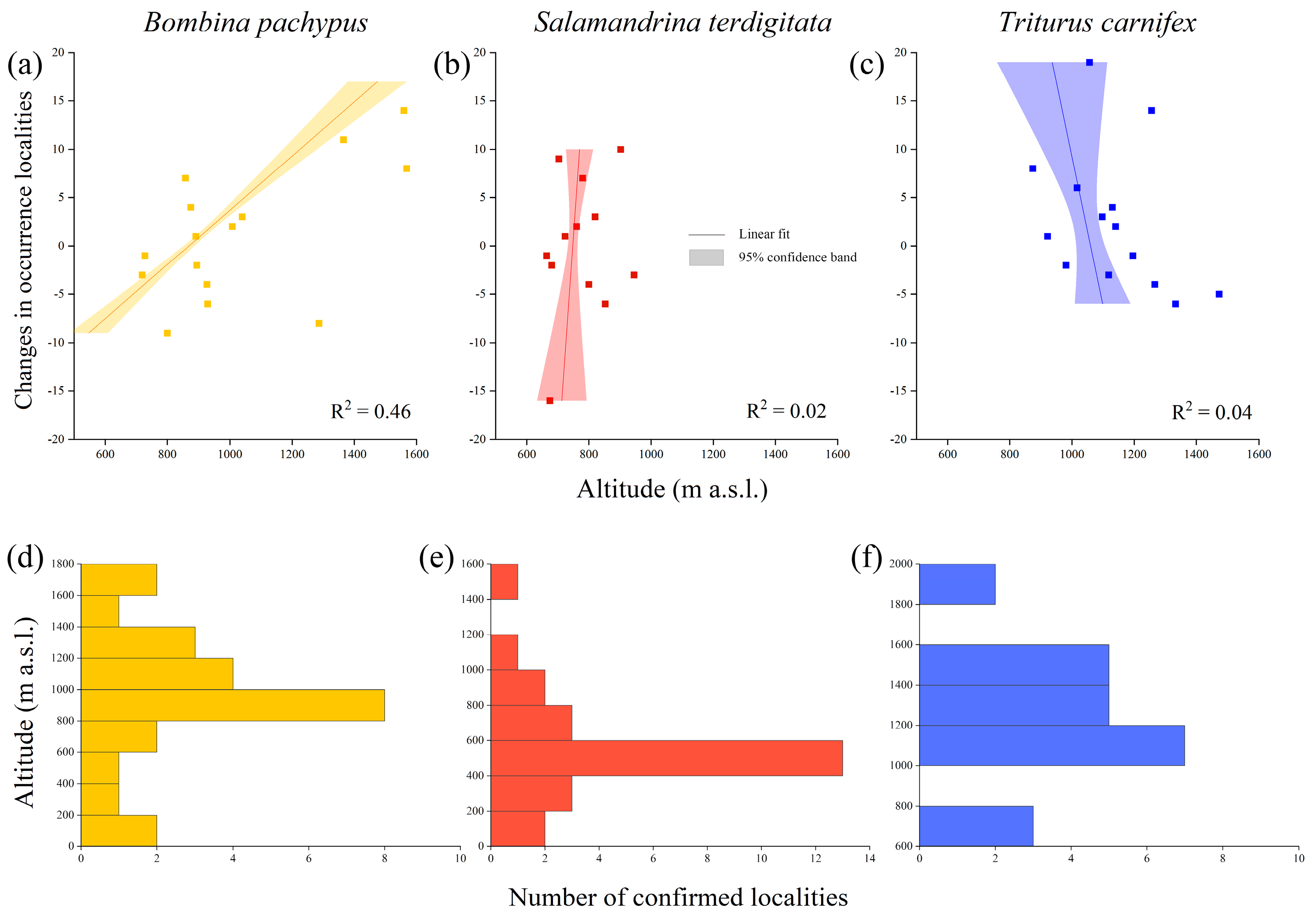
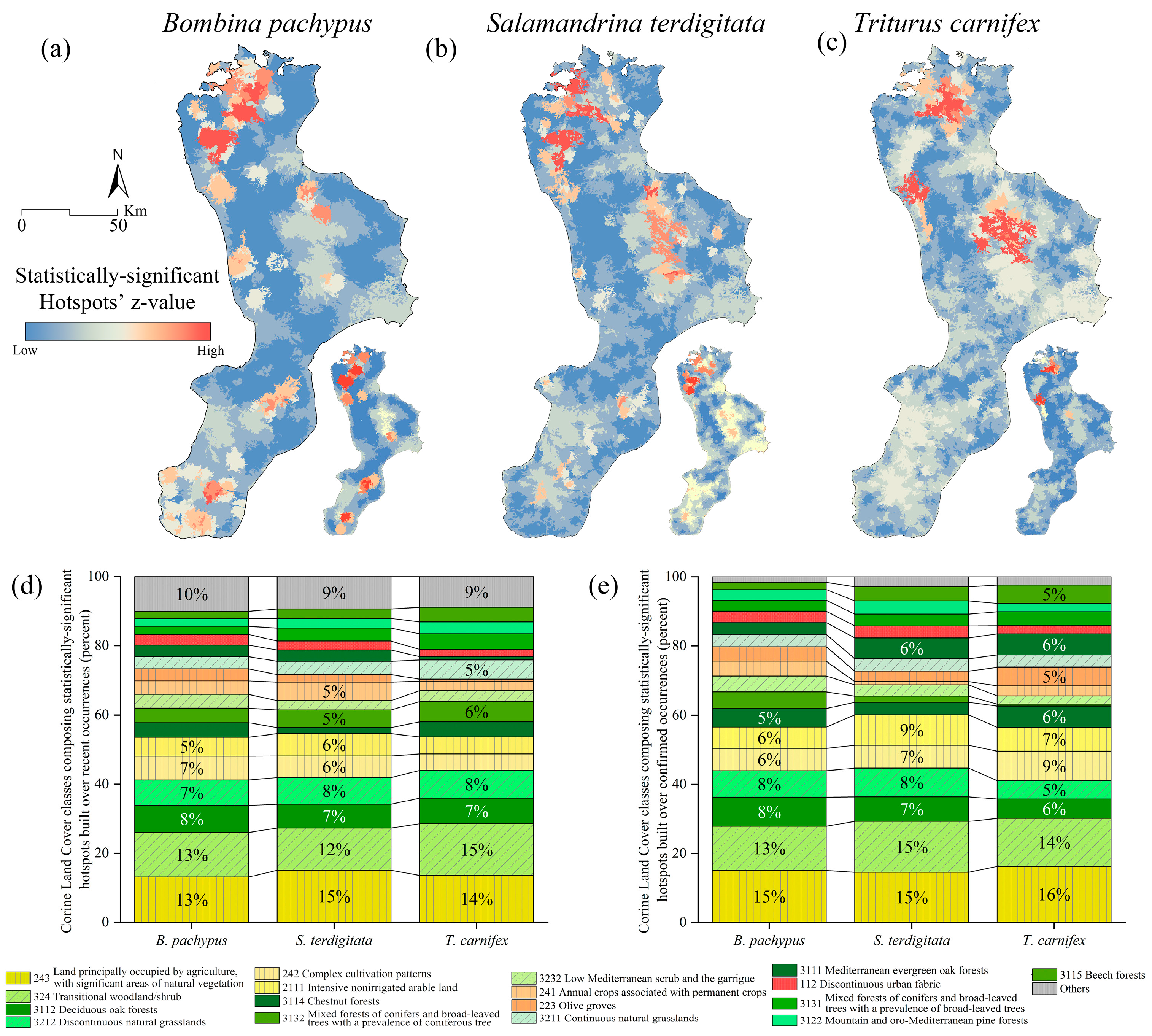
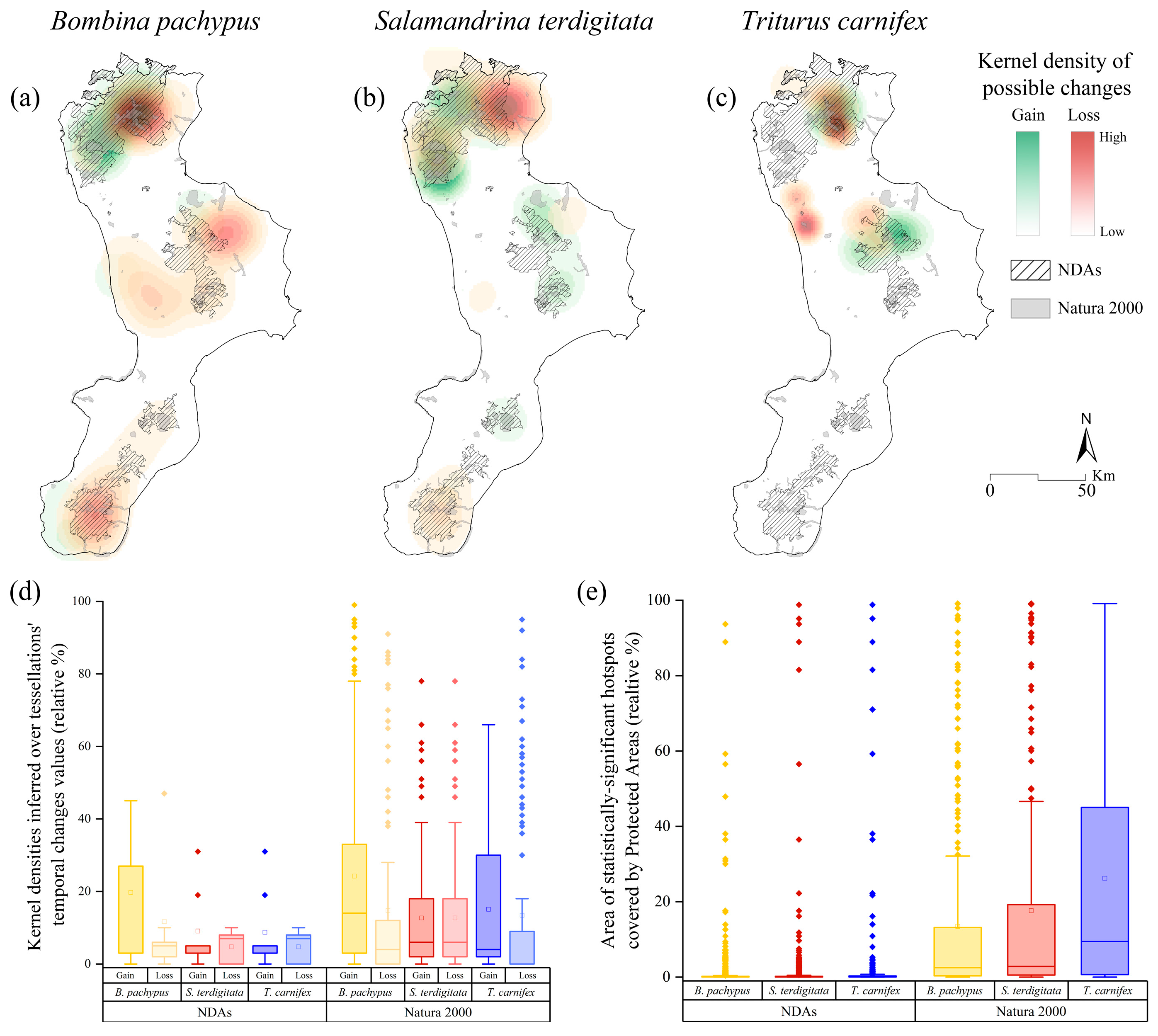
3.1. Bombina pachypus
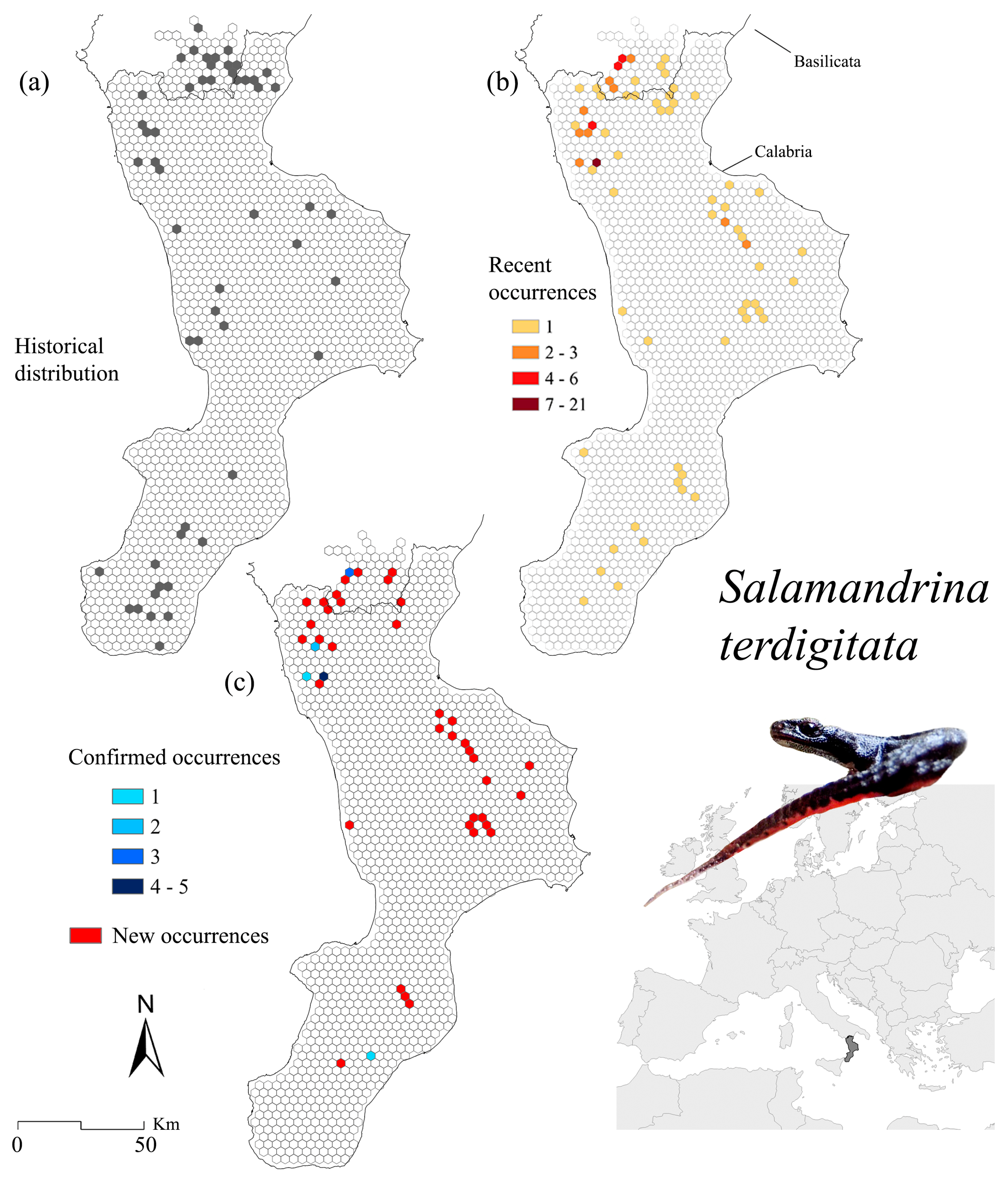
3.2. Salamandrina terdigitata
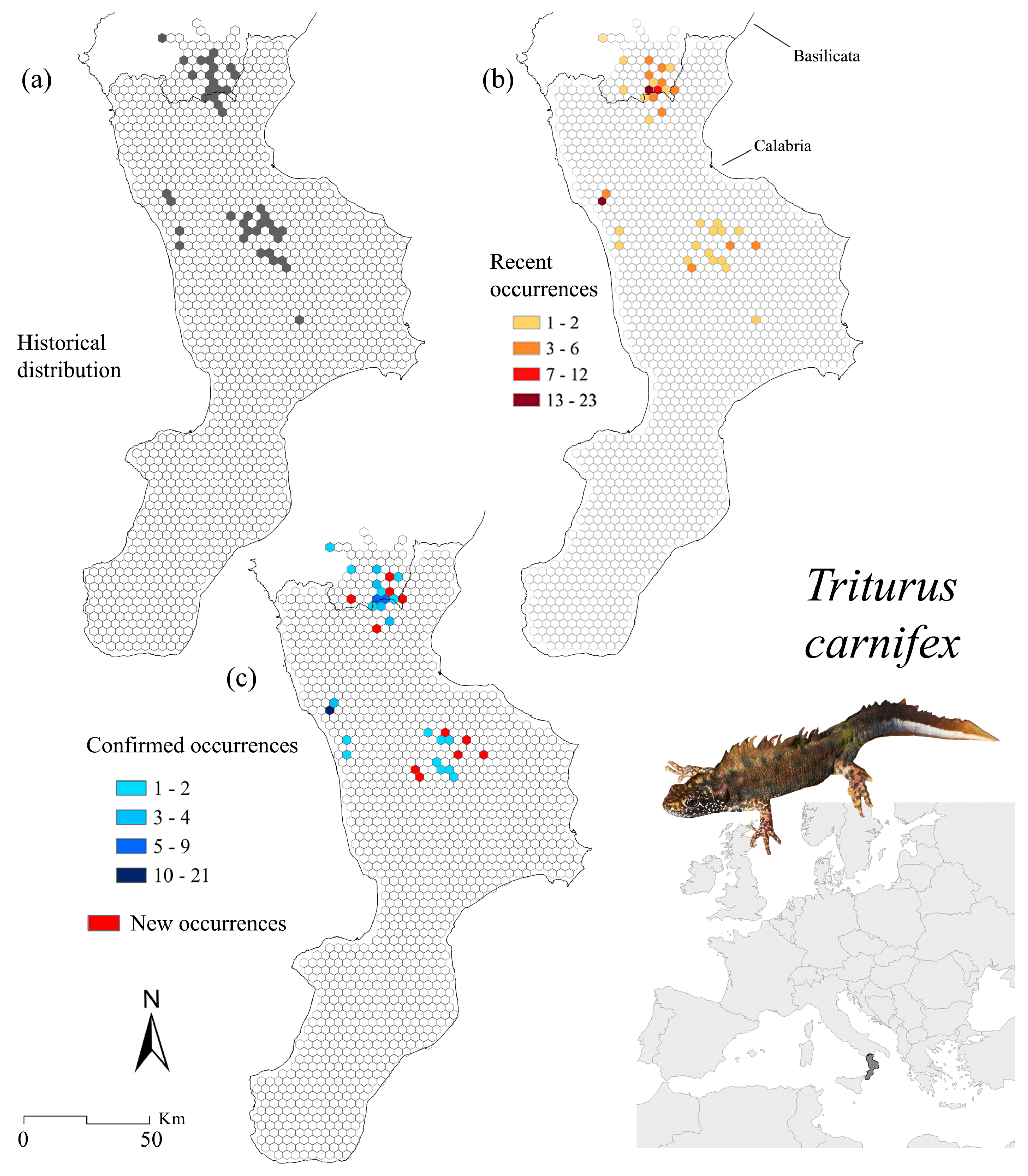
3.3. Triturus carnifex
4. Discussion
4.1. Bombina pachypus
4.2. Salamadrina terdigitata
4.3. Triturus carnifex
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hewitt, G. The genetic legacy of the quaternary ice ages. Nature 2000, 405, 907–913. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, T.; Fritz, U.; Delfino, M.; Ulrich, W.; Habel, J.C. Biogeography of Italy revisited: Genetic lineages confirm major phylogeographic patterns and a pre-pleistocene origin of its biota. Front. Zool. 2021, 18, 34. [Google Scholar] [CrossRef] [PubMed]
- Canestrelli, D.; Cimmaruta, R.; Costantini, V.; Nascetti, G. Genetic diversity and phylogeography of the apennine yellow-bellied toad Bombina pachypus, with implications for conservation. Mol. Ecol. 2006, 15, 3741–3754. [Google Scholar] [CrossRef]
- Canestrelli, D.; Aloise, G.; Cecchetti, S.; Nascetti, G. Birth of a hotspot of intraspecific genetic diversity: Notes from the underground. Mol. Ecol. 2010, 19, 5432–5451. [Google Scholar] [CrossRef]
- Canestrelli, D.; Sacco, F.; Nascetti, G. On glacial refugia, genetic diversity, and microevolutionary processes: Deep phylogeographical structure in the endemic newt Lissotriton italicus. Biol. J. Linn. Soc. 2012, 105, 42–55. [Google Scholar] [CrossRef]
- Iannella, M.; D’Alessandro, P.; Biondi, M. Evidences for a shared history for spectacled salamanders, haplotypes and climate. Sci. Rep. 2018, 8, 16507. [Google Scholar] [CrossRef]
- Schultze, N.; Spitzweg, C.; Corti, C.; Delaugerre, M.; Di Nicola, M.R.; Geniez, P.; Lapini, L.; Liuzzi, C.; Lunghi, E.; Novarini, N. Mitochondrial ghost lineages blur phylogeography and taxonomy of Natrix helvetica and N. natrix in Italy and Corsica. Zool. Scri. 2020, 49, 395–411. [Google Scholar] [CrossRef]
- Bisconti, R.; Aloise, G.; Siclari, A.; Fava, V.; Provenzano, M.; Arduino, P.; Chiocchio, A.; Nascetti, G.; Canestrelli, D. Forest dormouse (Dryomys nitedula) populations in southern Italy belong to a deeply divergent evolutionary lineage. Hystrix It. J. Mamm. 2018, 29, 75–79. [Google Scholar] [CrossRef]
- Chiocchio, A.; Colangelo, P.; Aloise, G.; Amori, G.; Bertolino, S.; Bisconti, R.; Castiglia, R.; Canestrelli, D. Population genetic structure of the bank vole Myodes glareolus within its glacial refugium in Peninsular Italy. J. Zool. Syst. Evol. Res. 2019, 57, 959–969. [Google Scholar] [CrossRef]
- Wauters, L.A.; Amori, G.; Aloise, G.; Gippoliti, S.; Agnelli, P.; Galimberti, A.; Casiraghi, M.; Preatoni, D.; Martinoli, A. New endemic mammal species for Europe: Sciurus meridionalis (Rodentia, Sciuridae). Hystrix 2017, 28, 1. [Google Scholar]
- Temple, H.; Cox, N. European Red List of Amphibians; Office for Official Publications of the European Communities: Luxembourg, 2009. [Google Scholar]
- Sillero, N.; Campos, J.; Bonardi, A.; Corti, C.; Creemers, R.; Crochet, P.-A.; Isailović, J.C.; Denoël, M.; Ficetola, G.F.; Gonçalves, J. Updated distribution and biogeography of amphibians and reptiles of Europe. Amphib. Reptil. 2014, 35, 1–31. [Google Scholar] [CrossRef]
- Rondinini, C.; Battistoni, A.; Peronace, V.; Teofili, C. Lista Rossa Dei Vertebrati Italiani; Comitato Italiano IUCN e Ministero dell’Ambiente e della Tutela del Territorio e del Mare: Rome, Italy, 2013. [Google Scholar]
- IUCN. The IUCN Red List of Threatened Species. Version 2021-3. Available online: http://www.iucnredlist.org (accessed on 10 April 2022).
- Hoffmann, M.; Hilton-Taylor, C.; Angulo, A.; Böhm, M.; Brooks, T.M.; Butchart, S.H.; Carpenter, K.E.; Chanson, J.; Collen, B.; Cox, N.A. The impact of conservation on the status of the world’s vertebrates. Science 2010, 330, 1503–1509. [Google Scholar] [CrossRef] [PubMed]
- Falaschi, M.; Muraro, M.; Gibertini, C.; Delle Monache, D.; Lo Parrino, E.; Faraci, F.; Belluardo, F.; Di Nicola, M.R.; Manenti, R.; Ficetola, G.F. Explaining declines of newt abundance in northern Italy. Freshw. Biol. 2022, 67, 1174–1187. [Google Scholar] [CrossRef]
- Canessa, S.; Oneto, F.; Ottonello, D.; Arillo, A.; Salvidio, S. Land abandonment may reduce disturbance and affect the breeding sites of an endangered amphibian in northern Italy. Oryx 2013, 47, 280–287. [Google Scholar] [CrossRef]
- Salvidio, S.; Andreone, F.; Angelini, J.; Bassu, L.; Bennati, R.; Bernini, F.; Bettiol, K.; Biggi, E.; Bionda, R.; Bocca, M.; et al. Assessing the status of amphibian breeding sites in Italy: A national survey. Acta Herpetol. 2011, 6, 119–126. [Google Scholar]
- Romano, A.; Salvidio, S.; Mongillo, D.; Olivari, S. Importance of a traditional irrigation system in amphibian conservation in the Cinque Terre National Park (NW Italy). J. Nat. Conserv. 2014, 22, 445–452. [Google Scholar] [CrossRef]
- Iannella, M.; Console, G.; D’Alessandro, P.; Cerasoli, F.; Mantoni, C.; Ruggieri, F.; Di Donato, F.; Biondi, M. Preliminary analysis of the diet of Triturus carnifex and pollution in Mountain Karst Ponds in Central Apennines. Water 2020, 12, 44. [Google Scholar] [CrossRef]
- Ficetola, G.F.; De Bernardi, F. Amphibians in a human-dominated landscape: The community structure is related to habitat features and isolation. Biol. Conserv. 2004, 119, 219–230. [Google Scholar] [CrossRef]
- D’Amen, M.; Bombi, P. Global warming and biodiversity: Evidence of climate-linked amphibian declines in Italy. Biol. Conserv. 2009, 142, 3060–3067. [Google Scholar] [CrossRef]
- Zampiglia, M.; Bisconti, R.; Maiorano, L.; Aloise, G.; Siclari, A.; Pellegrino, F.; Martino, G.; Pezzarossa, A.; Chiocchio, A.; Martino, C.; et al. Drilling down hotspots of intraspecific diversity to bring them into on-ground conservation of threatened species. Front. Ecol. Evol. 2019, 7, 205. [Google Scholar] [CrossRef]
- Macaluso, L.; Villa, A.; Carnevale, G.; Delfino, M. Past, present, and future climate space of the only endemic vertebrate genus of the Italian Peninsula. Sci. Rep. 2021, 11, 22139. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.; Dondero, L.; Allaria, G.; Morales Sanchez, B.N.; Rosa, G.; Salvidio, S.; Grasselli, E. Modelling the amphibian chytrid fungus spread by connectivity analysis: Towards a national monitoring network in Italy. Biodivers. Conserv. 2021, 30, 2807–2825. [Google Scholar] [CrossRef]
- European Environment Information and Observation Network (EIONET). Article 17 Web Tool.
- Stoch, F.; Grignetti, A. IV Report Direttiva Habitat: Specie Animali; Ercole, S., Angelini, P., Carnevali, L., Casella, L., Giacanelli, V., Grignetti, A., La Mesa, G., Nardelli, R., Serra, L., Stoch, F., et al., Eds.; Rapporti Direttive Natura (2013–2018); Sintesi Dello Stato Di Conservazione Delle Specie e Degli Habitat Di Interesse Comunitario e Delle Azioni Di Contrasto Alle Specie Esotiche Di Rilevanza Unionale in Italia; Serie Rapporti; ISPRA: Rome, Italy, 2021. [Google Scholar]
- Sindaco, R.; Doria, G.; Razzetti, E.; Bernini, F. Atlante Degli Anfibi e Dei Rettili d’Italia; Societas Herpetologica Italica, Edizioni Polistampa: Firenze, Italy, 2006. [Google Scholar]
- Hofman, S.; Spolsky, C.; Uzzell, T.; Cogălniceanu, D.; Babik, W.; Szymura, J.M. Phylogeography of the fire-bellied toads Bombina: Independent pleistocene histories inferred from mitochondrial genomes. Mol. Ecol. 2007, 16, 2301–2316. [Google Scholar] [CrossRef]
- Zheng, Y.; Fu, J.; Li, S. Toward understanding the distribution of Laurasian frogs: A test of savage’s biogeographical hypothesis using the genus Bombina. Mol. Phylogenet. Evol. 2009, 52, 70–83. [Google Scholar] [CrossRef]
- Fijarczyk, A.; Nadachowska, K.; Hofman, S.; Litvinchuk, S.N.; Babik, W.; Stuglik, M.; Gollmann, G.; Choleva, L.; Cogălniceanu, D.; Vukov, T. Nuclear and mitochondrial phylogeography of the European fire-bellied toads Bombina bombina and Bombina variegata supports their independent histories. Mol. Ecol. 2011, 20, 3381–3398. [Google Scholar] [CrossRef] [PubMed]
- Dufresnes, C.; Suchan, T.; Smirnov, N.A.; Denoël, M.; Rosanov, J.M.; Litvinchuk, S.N. Revisiting a speciation classic: Comparative analyses support sharp but leaky transitions between Bombina toads. J. Biogeogr. 2021, 48, 548–560. [Google Scholar] [CrossRef]
- Lanza, B.; Vanni, S. Notes on the biogeography of the Mediterranean Islands amphibians. Atti Convegni Lincei 1991, 85, 335–344. [Google Scholar]
- Talarico, E.; Sperone, E.; Tripepi, S. Amphibians of the Pollino National Park: Distribution and notes on conservation. Ital. J. Zool. 2004, 71, 203–208. [Google Scholar] [CrossRef]
- Lanza, B.; Andreone, F.; Bologna, M.A.; Corti, C.; Razzetti, E. Fauna d’Italia. Amphibia; Vol XLII. Amphibia; Edizioni Calderini: Bologna, Italy, 2007; Volume 24. [Google Scholar]
- Bernabò, I.; Biondi, M.; Cittadino, V.; Sperone, E.; Iannella, M. Addressing conservation measures through fine-tuned species distribution models for Bombina pachypus, an Italian endangered endemic anuran. Glob. Ecol. Conserv. 2022. accepted. [Google Scholar]
- Mattoccia, M.; Romano, A.; Sbordoni, V. Mitochondrial DNA sequence analysis of the spectacled salamander, Salamandrina terdigitata (Urodela: Salamandridae), supports the existence of two distinct species. Zootaxa 2005, 995, 19. [Google Scholar] [CrossRef]
- Canestrelli, D.; Zangari, F.; Nascetti, G. Genetic evidence for two distinct species within the Italian endemic Salamandrina terdigitata (Bonnaterre, 1789) (Amphibia: Urodela: Salamandridae). Herpetol. J. 2006, 16, 221–227. [Google Scholar]
- Romano, A.; Mattoccia, M.; Marta, S.; Bogaerts, S.; Pasmans, F.; Sbordoni, V. Distribution and morphological characterization of the endemic Italian salamanders Salamandrina perspicillata (Savi, 1821) and S. terdigitata (Bonnaterre, 1789) (Caudata: Salamandridae). Ital. J. Zool. 2009, 76, 422–432. [Google Scholar] [CrossRef]
- Angelini, C.; Vanni, S.; Vignoli, L. Salamandrina terdigitata (Bonnatterre, 1789), Salamandrina perspicillata (Savi, 1821). In Fauna d’Italia XLII: Amphibia; Calderini: Bologna, Italy, 2007; pp. 228–237. [Google Scholar]
- Romano, A.; Bartolomei, R.; Conte, A.; Fulco, E. Amphibians in southern Apennine: Distribution, ecology and conservation notes in the “Appennino Lucano, Val d’Agri e Lagonegrese” National Park (Southern Italy). Acta Herpetol. 2012, 7, 203–219. [Google Scholar]
- Sindaco, R.; Romano, A.; Mattoccia, M.; Sbordoni, V.; Andreone, F.; Corti, C. Salamandrina terdigitata. Available online: https://www.iucnredlist.org/species/59468/11945789 (accessed on 15 June 2022).
- Vanni, S.; Andreone, F.; Tripepi, S. Triturus carnifex (Laurenti, 1768). In Fauna d’Italia XLII: Amphibia; Calderini: Bologna, Italy, 2007; pp. 265–272. [Google Scholar]
- Andreone, F.; Corti, C.; Ficetola, F.; Razzetti, E.; Romano, A.; Sindaco, R. Triturus carnifex. Available online: http://www.iucn.it/scheda.php?id=1167401365 (accessed on 15 June 2022).
- Canestrelli, D.; Salvi, D.; Maura, M.; Bologna, M.A.; Nascetti, G. One species, three pleistocene evolutionary histories: Phylogeography of the Italian crested newt, Triturus carnifex. PLoS ONE 2012, 7, e41754. [Google Scholar] [CrossRef]
- Mattoccia, M.; Marta, S.; Romano, A.; Sbordoni, V. Phylogeography of an Italian endemic salamander (Genus Salamandrina): Glacial refugia, postglacial expansions, and secondary contact. Biol. J. Linn. Soc. 2011, 104, 903–992. [Google Scholar] [CrossRef]
- Hauswaldt, J.S.; Angelini, C.; Pollok, A.; Steinfartz, S. Hybridization of two ancient salamander lineages: Molecular evidence for endemic spectacled salamanders on the Apennine Peninsula. J. Zool. 2011, 284, 248–256. [Google Scholar] [CrossRef]
- Pignatti, S.; Cavalli, R.M.; Cuomo, V.; Fusilli, L.; Pascucci, S.; Poscolieri, M.; Santini, F. Evaluating hyperion capability for land cover mapping in a fragmented ecosystem: Pollino National Park, Italy. Remote Sens. Environ. 2009, 113, 622–634. [Google Scholar] [CrossRef]
- Rivas-Martínez, S.; Rivas-Sáenz, S.; Penas, A. Worldwide bioclimatic classification system. Glob. Geobot. 2011, 1, 1–634. [Google Scholar]
- Spampinato, G. Biodiversità delle foreste calabresi. In Foreste Di Calabria; Regione Calabria—Assessorato Alle Foreste e alla Forestazione, Protezione Civile, Pari Opportunità: Catanzaro, Italy, 2003. [Google Scholar]
- Spampinato, G.; Bernardo, L.; Passalaqua, N. La Vegetazione D’Italia, Con Carta Delle Serie Di Vegetazione in Scala 1:500,000; Blasi, C., Ed.; Palombi Editori: Roma, Italy, 2010. [Google Scholar]
- ISPRA Territorio. Processi e trasformazioni in Italia. Rapporti 296/2018. ISPRA Rapp. 2018, 296, 2018. [Google Scholar]
- Spampinato, G.; Crisafulli, A.; Cameriere, P. Trasformazione delle zone costiere della regione Calabria (Italia Meridionale) e conseguenze sulla flora degli ambienti umidi. Fitosociologia 2007, 44, 119–128. [Google Scholar]
- Spampinato, G.; Crisarà, R.; Cameriere, P.; Cano-Ortiz, A.; Musarella, C.M. Analysis of the forest landscape and its transformations through phytotoponyms: A case study in Calabria (Southern Italy). Land 2022, 11, 518. [Google Scholar] [CrossRef]
- Sperone, E.; Bonacci, A.; Brunelli, E.; Corapi, B.; Tripepi, S. Ecologia e conservazione dell’erpetofauna della catena costiera Calabra. Stud. Trentini Sci. Nat. Acta Biol. 2007, 83, 99–104. [Google Scholar]
- Sperone, E.; Bonacci, A.; Corapi, B.; Tripepi, S. Notes on the distribution and ecology of the Apennine yellow-bellied toad Bombina pachypus in Calabria and Lucania. In Proceedings of the 6° Congresso Nazionale della Societas Herpetologica Italica, Stilgrafica, Roma, Italy, 27 September–1 October 2006; Bologna, M.A., Capula, M., Carpaneto, G.M., Luiselli, L., Marangoni, C., Venchi, A., Eds.; 2006; pp. 33–34. [Google Scholar]
- Tripepi, S.; Sperone, E.; Talarico, E. Distribuzione, ecologia e conservazione del tritone crestato Italiano, “Triturus carnifex”. In Calabria ed in Lucania Meridionale, Proceedings of the V Congresso Nazionale Societas Herpetologica Italica, Calci (Pisa), Italy, 29 September–3 October 2004; Zuffi, M.A.L., Ed.; Firenze University Press: Firenze, Italy, 2006; pp. 261–267. [Google Scholar]
- Dodd, C.K. Amphibian Ecology and Conservation: A Handbook of Techniques; Oxford University Press: Oxford, UK, 2010; ISBN 0-19-954119-1. [Google Scholar]
- Stoch, F.; Genovesi, P. Manuali per il monitoraggio di specie e habitat di interesse comunitario (Direttiva 92/43/CEE) in Italia: Specie animali. ISPRA Ser. Man. Linee Guida 2016, 141, 1–292. [Google Scholar]
- ESRI Inc. ArcGIS Pro 2.9; ESRI Inc.: Redlands, CA, USA, 2022. [Google Scholar]
- Alexander, J.M.; Diez, J.M.; Levine, J.M. Novel competitors shape species’ responses to climate change. Nature 2015, 525, 515–518. [Google Scholar] [CrossRef]
- Iannella, M.; D’Alessandro, P.; Biondi, M. Forecasting the spread associated with climate change in eastern Europe of the invasive Asiatic Flea Beetle, Luperomorpha xanthodera (Coleoptera: Chrysomelidae). Eur. J. Entomol. 2020, 117, 130–138. [Google Scholar] [CrossRef]
- Iannella, M.; Console, G.; Cerasoli, F.; De Simone, W.; D’Alessandro, P.; Biondi, M. A step towards SDMs: A “couple-and-weigh” framework based on accessible data for biodiversity conservation and landscape planning. Divers. Distrib. 2021, 27, 2412–2427. [Google Scholar] [CrossRef]
- Shepard, I.D.; Wissinger, S.A.; Greig, H.S. Elevation alters outcome of competition between resident and range-shifting species. Glob. Chang. Biol. 2021, 27, 270–281. [Google Scholar] [CrossRef]
- Cerasoli, F.; Iannella, M.; Biondi, M. Between the hammer and the anvil: How the combined effect of global warming and the non-native common slider could threaten the European pond turtle. Manag. Biol. Invasions 2019, 10, 428–448. [Google Scholar] [CrossRef]
- Poloni, R.; Iannella, M.; Fusco, G.; Fattorini, S. Conservation biogeography of high-altitude longhorn beetles under climate change. Insect Conserv. Diver. 2022, 15, 429–444. [Google Scholar] [CrossRef]
- Di Musciano, M.; Carranza, M.; Frate, L.; Di Cecco, V.; Di Martino, L.; Frattaroli, A.; Stanisci, A. Distribution of plant species and dispersal traits along environmental gradients in central Mediterranean summits. Diversity 2018, 10, 58. [Google Scholar] [CrossRef]
- Di Musciano, M.; Ricci, L.; Di Cecco, V.; Bricca, A.; Di Martino, L.; Frattaroli, A.R. Elevational patterns of plant dispersal ability in southern Europe. Plant Biosyst. 2022, 1–9. [Google Scholar] [CrossRef]
- Sillero, N. Climate change in action: Local elevational shifts on Iberian amphibians and reptiles. Reg. Environ. Chang. 2021, 21, 101. [Google Scholar] [CrossRef]
- Getis, A.; Ord, J.K. The analysis of spatial association by use of distance statistics. Geogr. Anal. 1992, 24, 189–206. [Google Scholar] [CrossRef]
- Iannella, M.; D’Alessandro, P.; Biondi, M. Entomological knowledge in madagascar by GBIF datasets: Estimates on the coverage and possible biases (Insecta). Fragm. Entomol. 2019, 51, 1–10. [Google Scholar] [CrossRef]
- De Giglio, O.; Fasano, F.; Diella, G.; Lopuzzo, M.; Napoli, C.; Apollonio, F.; Brigida, S.; Calia, C.; Campanale, C.; Marzella, A. Legionella and legionellosis in touristic-recreational facilities: Influence of climate factors and geostatistical analysis in southern Italy (2001–2017). Environ. Res. 2019, 178, 108721. [Google Scholar] [CrossRef]
- Sánchez-Martín, J.-M.; Rengifo-Gallego, J.-I.; Blas-Morato, R. Hot spot analysis versus cluster and outlier analysis: An enquiry into the grouping of rural accommodation in Extremadura (Spain). ISPRS Int. J. Geo-Inf. 2019, 8, 176. [Google Scholar] [CrossRef]
- Tegegne, T.K.; Chojenta, C.; Getachew, T.; Smith, R.; Loxton, D. Service environment link and false discovery rate correction: Methodological considerations in population and health facility surveys. PLoS ONE 2019, 14, e0219860. [Google Scholar] [CrossRef]
- Gigante, D.; Foggi, B.; Venanzoni, R.; Viciani, D.; Buffa, G. Habitats on the grid: The spatial dimension does matter for red-listing. J. Nat. Conserv. 2016, 32, 1–9. [Google Scholar] [CrossRef]
- Iannella, M.; Fiasca, B.; Di Lorenzo, T.; Biondi, M.; Di Cicco, M.; Galassi, D.M.P. Jumping into the grids: Mapping biodiversity hotspots in groundwater habitat types across Europe. Ecography 2020, 43, 1825–1841. [Google Scholar] [CrossRef]
- Canestrelli, D.; Zampiglia, M.; Nascetti, G. Widespread occurrence of Batrachochytrium dendrobatidis in contemporary and historical samples of the endangered Bombina pachypus along the Italian Peninsula. PLoS ONE 2013, 8, e63349. [Google Scholar] [CrossRef]
- Tripepi, S.; Rossi, F.; Serroni, P.; Brunelli, E. Distribuzione altitudinale degli anfibi in Calabria. Stud. Trentini Sci. Nat. Acta Biol. 1996, 71, 97–101. [Google Scholar]
- Andreone, F.; Corti, C.; Ficetola, F.; Razzetti, E.; Romano, A.; Sindaco, R. Bombina pachypus. Available online: http://www.iucn.it/scheda.php?id=-291037272 (accessed on 15 June 2022).
- Barbieri, F.; Bernini, F.; Guarino, F.M.; Venchi, A. Distribution and conservation status of Bombina variegata in Italy (Amphibia, Bombinatoridae). Boll. Zool. 2004, 71, 83–90. [Google Scholar]
- Stagni, G.; Dall’olio, R.; Fusini, U.; Mazzotti, S.; Scoccianti, C.; Serra, A. Declining populations of apennine yellow-bellied toad Bombina pachypus in the northern Apennines (Italy): Is Batrachochytrium dendrobatidis the main cause? Ital. J. Zool. 2004, 71, 151–154. [Google Scholar] [CrossRef]
- Canestrelli, D.; Zampiglia, M.; Bisconti, R.; Nascetti, G. Proposta Di Intervento per La Conservazione Ed Il Recupero Delle Popolazioni Di Ululone Appenninico Bombina pachypus in Italia Peninsulare; Dipartimento DEB Università degli Studi della Tuscia e Ministero dell’Ambiente e della Tutela del Territorio e del Mare: Roma, Italy, 2014. [Google Scholar]
- Bernabò, I.; Crispino, F.; Gervasio, G.; Tripepi, S.; Marchianò, V. Aggiornamento su presenza e status di Bombina pachypus e Salamandrina terdigitata nel parco nazionale del pollino: Risultati preliminari. In Proceedings of the XII National Congress Societas Herpetologica Italica, Rende-Cosenza, Italy, 1–5 October 2018; Ventura, Ed.; 2021; pp. 169–174. [Google Scholar]
- Martino, G.; Chiocchio, A.; Siclari, A.; Canestrelli, D. Distribution and conservation status of endangered amphibians within the Aspromonte Mountain Region, a hotspot of Mediterranean biodiversity. ARPHA Prepr. 2022, 3, e86533. [Google Scholar]
- LUTRIA. Monitoraggio e Conservazione Delle Popolazioni Di Lontra (Lutra lutra) e Di Altre Specie Di Interesse Comunitario Di Ambiente Umido-Acquatico Nel Parco Nazionale Della Sila. Report 2. Monitoraggio Degli Anfibi e Valutazioni Gestionali, 2021.
- GREENWOOD. Attività Di Monitoraggio Della Batracofauna e Realizzazione Banca Dati Degli Habitat e Individuazione Dei Siti Di Monitoraggio Nel Perimetro Del Parco Nazionale Della Sila. Cap. 11400—E.F.2018. CIG: 5124A2333; Di Lavoro, G., Bernabò, I., Gervasio, G., Crispino, F., Campana, M., Eds.; Relazione Conclusiva Ottobre; 2019. [Google Scholar]
- Piazzini, S.; Martini, F. Monitoraggio Anfibi, Rettili e Pesci Nei Siti Della Rete Natura 2000 Del Parco Nazionale Della Sila. Relazione Finale. Campionamenti 2018-2019; 2019.
- Sperone, E.; Rizzo, P. Progetto Di Monitoraggio Del Lupo e Dell’ululone Nel Parco Nazionale Della Sila. CIG: Z471E31AE0—CAP. 1551 E.F. 2017 GESTIONE RR.PP. 2012/2013/2014/2015. Report Sul Monitoraggio Dell’Ululone Appenninico Bombina Pachypus—Relazione Tecnica Ottobre 2018; 2018.
- LIFE WetFlyAmphibia. LIFE14 NAT/IT/000759. Conservation of Amphibians and Butterflies of Open Wet Areas and Their Habitats at the Foreste Casentinesi National Park. 2019. Available online: http://www.lifewetflyamphibia.eu/ (accessed on 15 June 2022).
- Vignoli, L.; Macale, D.; Pieroni, A.; Bologna, M.A. Restocking of the Apennine yellow-bellied toad in Central Italy. In Global Conservation Translocation Perspectives: 2021. Case Studies from Around the Globe; IUCN: Gland, Switzerland, 2021; p. 52. [Google Scholar]
- Salvidio, S.; Oneto, F.; Ottonello, D.; Braida, L.; Ferravante, C.; Grasselli, E.; Vecchione, G.; Canessa, S.; Arillo, A.; Cardelli, M. Conservation of the Apennine yellow-bellied toad Bombina variegata pachypus in Liguria (NW Italy). Herpetol. Facts 2014, 1, 97–104. [Google Scholar]
- Brusco, A.; Marchianò, V.; Puntillo, M.; Cittadino, V.; Sperone, E.; Tripepi, S.; Bernabò, I. Progetto “ululone”: Dati preliminari sui primi tre anni di attività. In Proceedings of the XIII Congresso Nazionale SHI, Lipari, Italy, 22–26 September 2021. [Google Scholar]
- Angelini, C.; Vanni, S.; Vignoli, L. Anfibi d’Italia. In Quaderni di Conservazione della Natura; Lanza, B., Nistri, A., Vanni, S., Eds.; Ministero dell’Ambiente e della Tutela del Territorio e del Mare; Istituto Superiore per la protezione la ricerca ambientale: Rome, Italy, 2007; Volume 29, p. 456 + 1 CD bilingue. ISBN 1592-2901. [Google Scholar]
- Della Rocca, F.; Bottari, V.; Filippi, E.; Luiselli, L.; Utzeri, C. Modelling aspects of terrestrial ecology in an Italian endemic salamander, Salamandrina perspicillata. Rev. Écol. Terre Vie 2008, 63, 261–270. [Google Scholar]
- Caloiero, T.; Callegari, G.; Cantasano, N.; Coletta, V.; Pellicone, G.; Veltri, A. Bioclimatic analysis in a region of southern Italy (Calabria). Plant Biosyst. 2016, 150, 1282–1295. [Google Scholar] [CrossRef]
- Giacoma, C.; Picariello, O.; Puntillo, D.; Rossi, F.; Tripepi, S. The distribution and habitats of the newt (Triturus, Amphibia) in Calabria (southern Italy). Monit. Zool. Ital.-Ital. J. Zool. 1988, 22, 449–464. [Google Scholar]
- Seglie, D.; Sindaco, R. Segnalazioni faunistiche piemontesi e valdostane, V.(Amphibia, Reptilia, Mammalia). Riv. Piemont. Stor. Nat. 2011, 32, 419–438. [Google Scholar]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bernabò, I.; Cittadino, V.; Tripepi, S.; Marchianò, V.; Piazzini, S.; Biondi, M.; Iannella, M. Updating Distribution, Ecology, and Hotspots for Three Amphibian Species to Set Conservation Priorities in a European Glacial Refugium. Land 2022, 11, 1292. https://doi.org/10.3390/land11081292
Bernabò I, Cittadino V, Tripepi S, Marchianò V, Piazzini S, Biondi M, Iannella M. Updating Distribution, Ecology, and Hotspots for Three Amphibian Species to Set Conservation Priorities in a European Glacial Refugium. Land. 2022; 11(8):1292. https://doi.org/10.3390/land11081292
Chicago/Turabian StyleBernabò, Ilaria, Viviana Cittadino, Sandro Tripepi, Vittoria Marchianò, Sandro Piazzini, Maurizio Biondi, and Mattia Iannella. 2022. "Updating Distribution, Ecology, and Hotspots for Three Amphibian Species to Set Conservation Priorities in a European Glacial Refugium" Land 11, no. 8: 1292. https://doi.org/10.3390/land11081292