
Assessing Ecological Disturbance in Neotropical Forest Landscapes Using High-Level Diversity and High-Level Functionality: Surprising Outcomes from a Case Study with Spider Assemblages

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
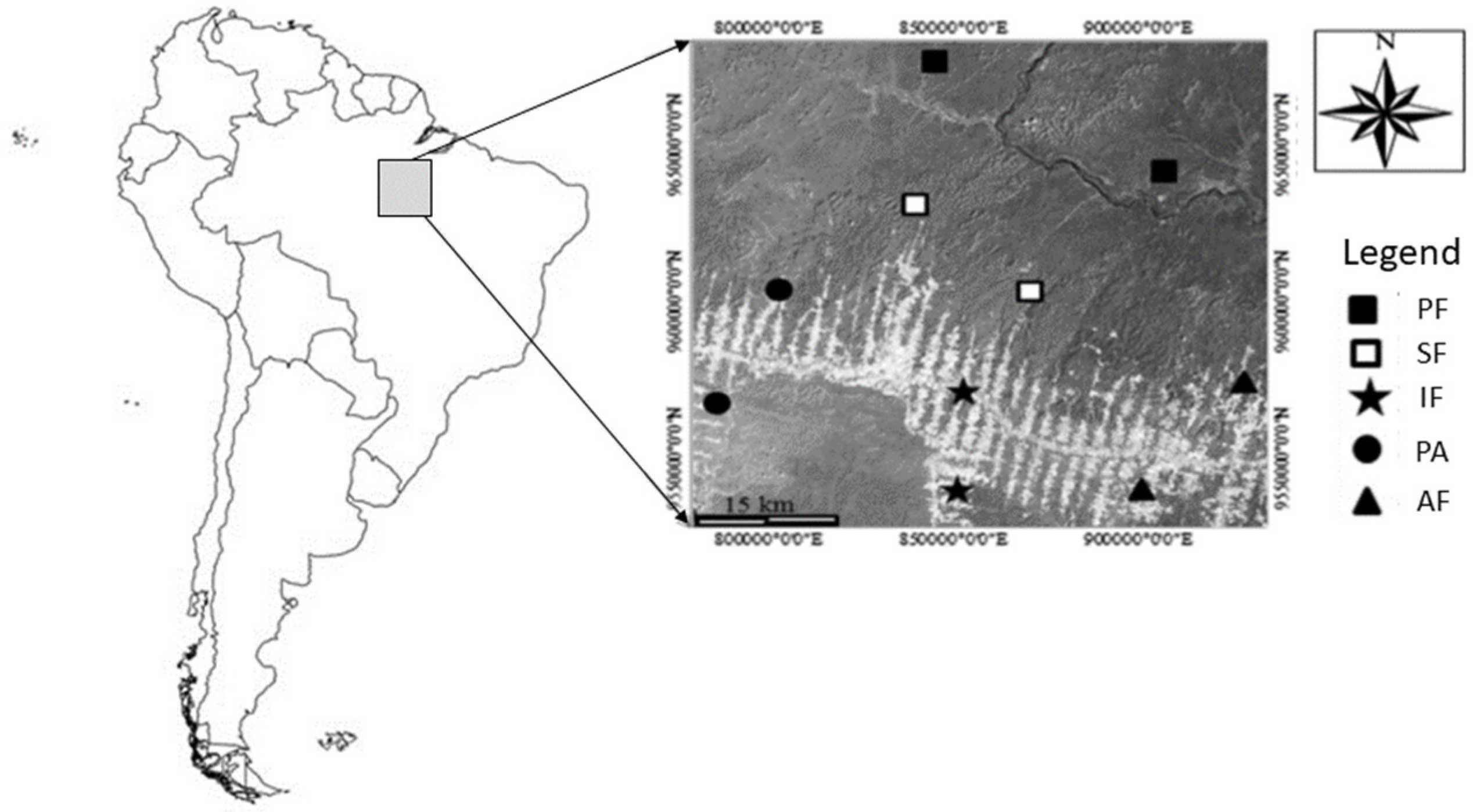
2.1. Study Sites
2.2. Data Collection and Identification
2.3. Environmental Factors and Variables
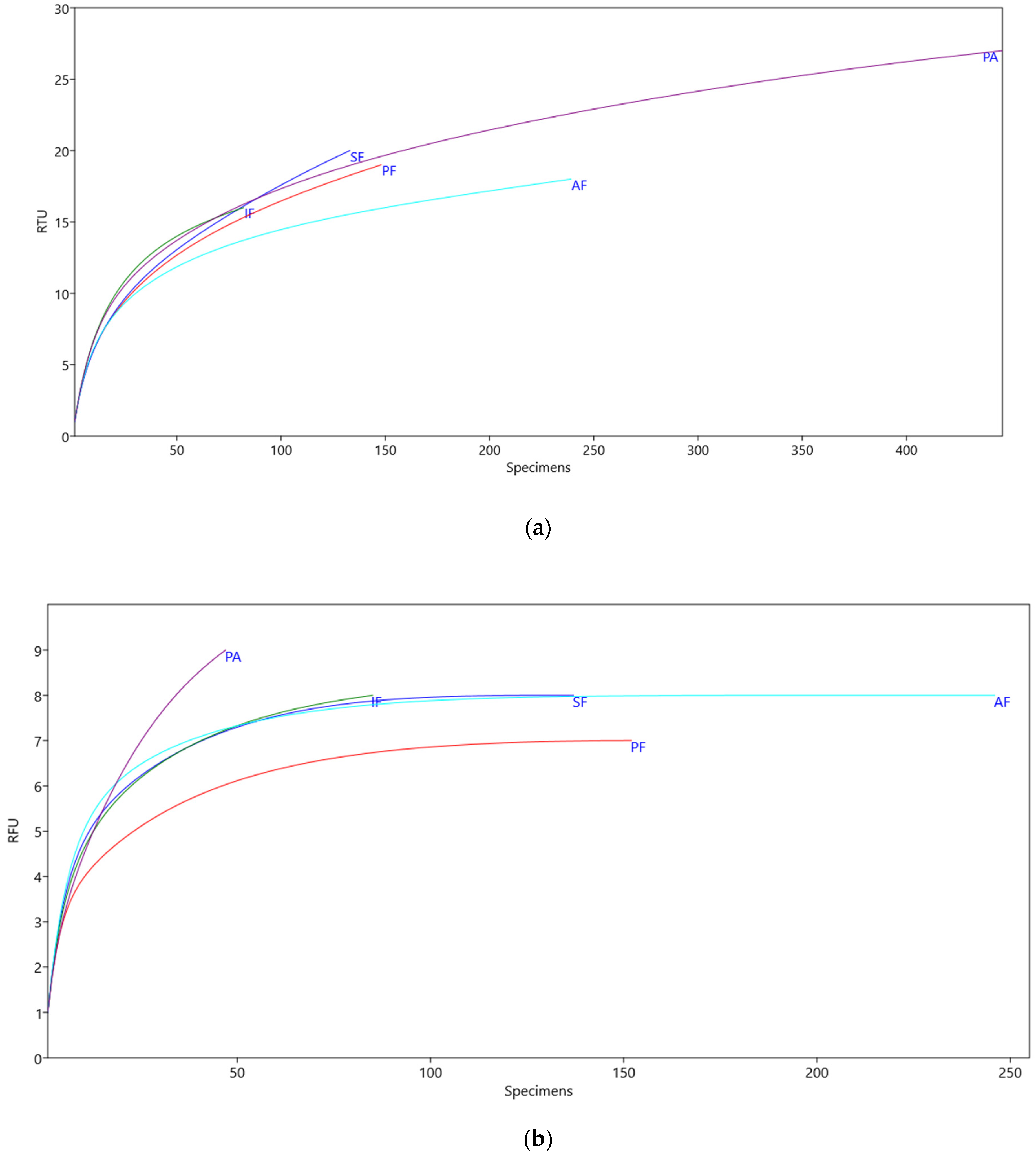
2.4. Sampling Completeness within Ecosystems
2.5. Taxonomic and Functional Diversity
2.6. Assemblage Analysis
Taxonomic Diversity and Functional Diversity Indicators’ Responses to Different Land Use
3. Results
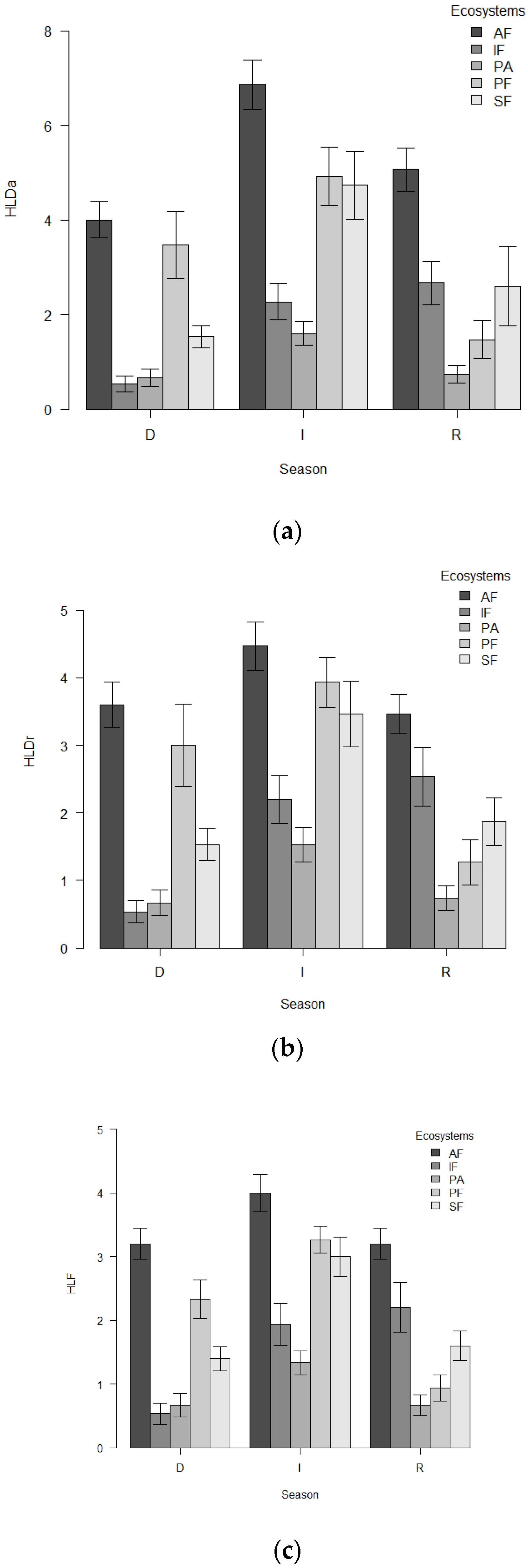
3.1. Taxonomic and Functional Diversity Responses to Disturbance
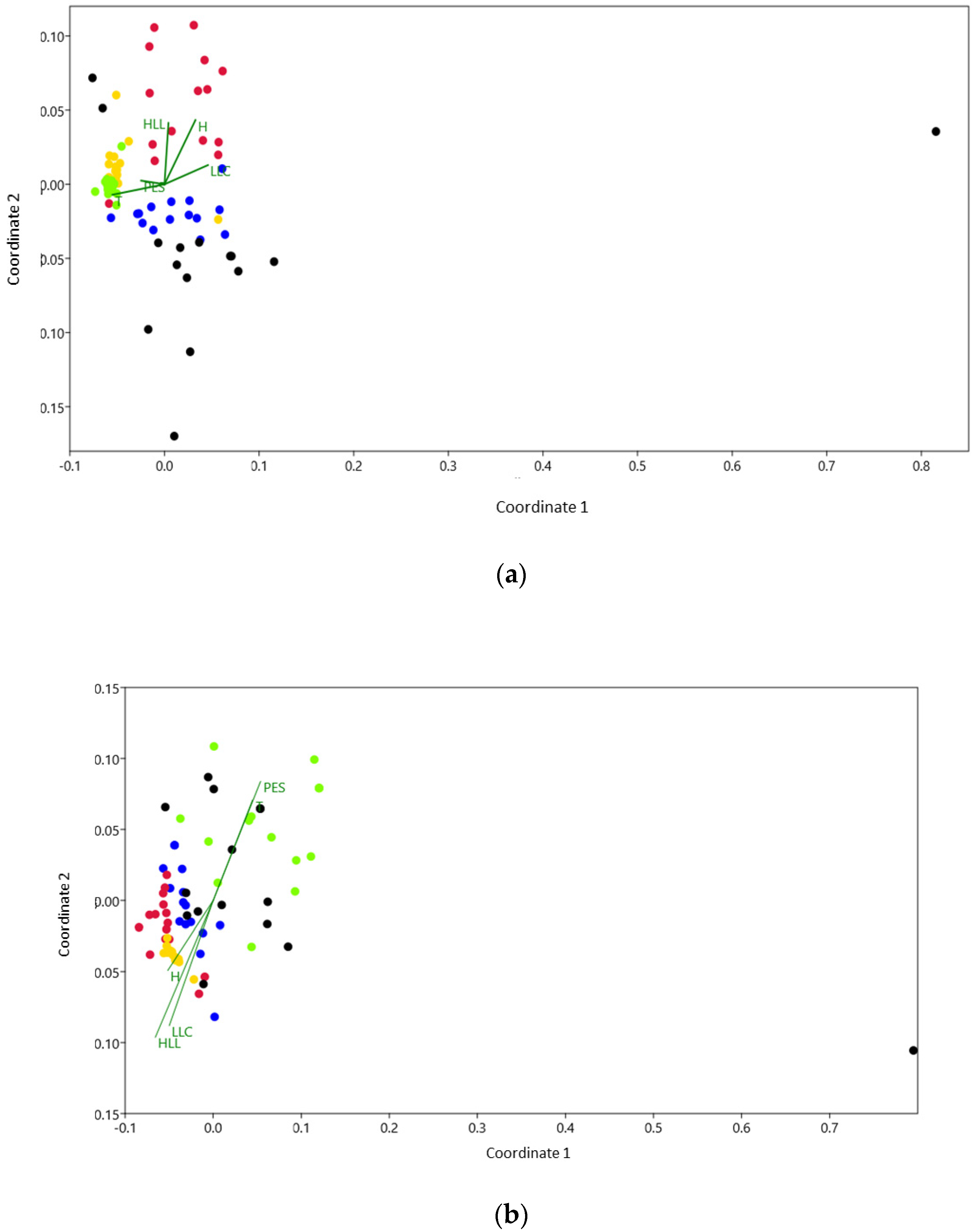
3.2. Taxonomic Composition and Functional Composition Responses to the Disturbance Gradient
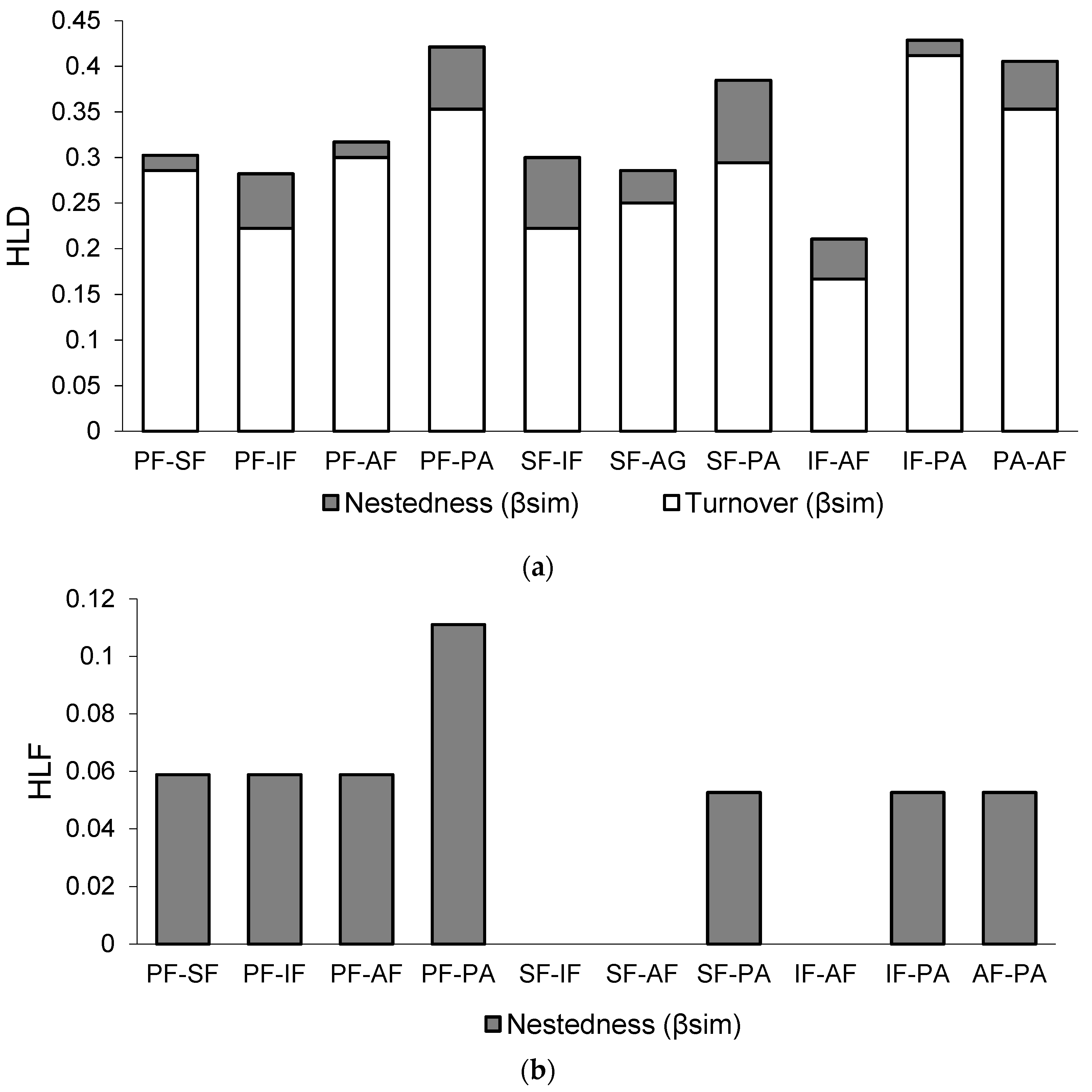
3.3. Comparing Taxonomic β-Diversity Indicators with Functional β-Diversity Indicators
3.4. Taxonomic and Functional Dissimilarity within the Disturbance Gradient
3.5. Families and Guilds as Disturbance Indicators
4. Discussion
4.1. Taxonomic Diversity and Functional Diversity Responses to the Disturbance Gradient
4.2. Taxonomic Composition and Functional Composition Responses to the Disturbance Gradient
4.3. Taxonomic β-Diversity and Functional β-Diversity Responses to the Disturbance Gradient
4.4. Taxonomic and Functional Dissimilarity within the Disturbance Gradient
4.5. Methodological Advantages and Drawbacks
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Calvo-Rodriguez, S.; Sanchez-Azofeifa, A.G.; Duran, S.M.; Espírito-Santo, M.M. Assessing ecosystem services in Neotropical dry forests: A systematic review. Environ. Conserv. 2016, 44, 34–43. [Google Scholar] [CrossRef]
- Brockerhoff, E.G.; Barbaro, L.; Castagneyrol, B.; Forrester, D.I.; Gardiner, B.; González-Olabarria, J.R.; Lyver, P.O.; Meurisse, N.; Oxbrough, A.; Taki, H.; et al. Forest biodiversity, eco-system functioning and the provision of ecosystem services. Biodivers. Conserv. 2017, 26, 3005–3035. [Google Scholar] [CrossRef] [Green Version]
- Baruch, Z.; Nozawa, S.; Johnson, E.; Yerena, E. Ecosystem dynamics and services of a paired Neotropical montane forest and pine plantation. Rev. Biol. Trop. 2019, 67, 24–35. [Google Scholar] [CrossRef]
- Wani, A.M.; Sahoo, G. Forest Ecosystem Services and Biodiversity. In Spatial Modeling in Forest Resources Management: Rural Livelihood and Sustainable Development; Shit, P.K., Pourghasemi, H.R., Das, P., Bhunia, G.S., Eds.; Springer Inter-National Publishing: Cham, Switzerland, 2021; pp. 529–552. [Google Scholar]
- Cortés-Calderón, S.; Mora, F.; Arreola-Villa, F.; Balvanera, P. Ecosystem services supply and interactions along secondary tropical dry forests succession. For. Ecol. Manag. 2021, 482, 118858. [Google Scholar] [CrossRef]
- Fearnside, P. Deforestation in Brazilian Amazonia: History, Rates, and Consequences. Conserv. Biol. 2005, 19, 680–688. [Google Scholar] [CrossRef]
- Barlow, J.; Lennox, G.D.; Ferreira, J.N.; Berenguer, E.; Lees, A.C.; Mac Nally, R.; Thomson, J.R.; Ferraz, S.F.D.B.; Louzada, J.N.; Oliveira, V.H.F.; et al. Anthropogenic disturbance in tropical forests can double biodiversity loss from deforestation. Nature 2016, 535, 144–147. [Google Scholar] [CrossRef] [Green Version]
- Barlow, J.; França, F.; Gardner, T.A.; Hicks, C.; Lennox, G.D.; Berenguer, E.; Castello, L.; Economo, E.P.; Ferreira, J.; Guénard, B.; et al. The future of hyperdiverse tropical ecosystems. Nat. Cell Biol. 2018, 559, 517–526. [Google Scholar] [CrossRef]
- Isbell, F.; Calcagno, V.; Hector, A.; Connolly, J.; Harpole, W.S.; Reich, P.B.; Scherer-Lorenzen, M.; Schmid, B.; Tilman, D.; Van Ruijven, J.; et al. High plant diversity is needed to maintain ecosystem services. Nature 2011, 477, 199–202. [Google Scholar] [CrossRef]
- Gamfeldt, L.; Snäll, T.; Bagchi, R.; Jonsson, M.; Gustafsson, L.; Kjellander, P.; Ruiz-Jaen, M.C.; Fröberg, M.; Stendahl, J.; Philipson, C.D.; et al. Higher levels of multiple ecosystem services are found in forests with more tree species. Nat. Commun. 2013, 4, 1340. [Google Scholar] [CrossRef]
- Dehling, J.M.; Dehling, D.M. Conserving ecological functions of frog communities in Borneo requires diverse forest landscapes. Glob. Ecol. Conserv. 2021, 26, e01481. [Google Scholar] [CrossRef]
- Cajaiba, R.L.; Périco, E.; Caron, E.; Dalzochio, M.S.; Silva, W.B.; Santos, M. Are disturbance gradients in neotropical ecosystems detected using rove beetles? A case study in the Brazilian Amazon. For. Ecol. Manag. 2017, 405, 319–327. [Google Scholar] [CrossRef]
- Cardoso, P.; Pekar, S.; Jocqué, R.; Coddington, J.A. Global Patterns of Guild Composition and Functional Diversity of Spiders. PLoS ONE 2011, 6, e21710. [Google Scholar] [CrossRef] [Green Version]
- Ometto, J.; Aguiar, A.P.; Martinelli, L. Amazon deforestation in Brazil: Effects, drivers and challenges. Carbon Manag. 2011, 2, 575–585. [Google Scholar] [CrossRef]
- Ochoa-Quintero, J.M.; Gardner, T.A.; Rosa, I.; de Barros Ferraz, S.F.; Sutherland, W.J. Thresholds of species loss in Amazonian de-forestation frontier landscapes. Conserv. Biol. 2015, 29, 440–451. [Google Scholar] [CrossRef]
- van Oudenhoven, A.P.E.; Schröter, M.; Drakou, E.G.; Geijzendorffer, I.R.; Jacobs, S.; van Bodegom, P.M.; Chazee, L.; Czúcz, B.; Grunewald, K.; Lillebø, A.I.; et al. Key criteria for developing ecosystem service indicators to inform decision making. Ecol. Indic. 2018, 95, 417–426. [Google Scholar] [CrossRef]
- Haines-Young, R.; Potschin, M. The links between biodiversity, ecosystem services and human well-being. In Ecosystem Ecology: A New Synthesis; Frid, C.L.J., Raffaelli, D.G., Eds.; Cambridge University Press: Cambridge, UK, 2010; pp. 110–139. [Google Scholar]
- Albert, C.; Galler, C.; Hermes, J.; Neuendorf, F.; von Haaren, C.; Lovett, A. Applying ecosystem services indicators in landscape planning and management: The ES-in-Planning framework. Ecol. Indic. 2016, 61, 100–113. [Google Scholar] [CrossRef]
- Birkhofer, K.; Rusch, A.; Andersson, G.; Bommarco, R.; Dänhardt, J.; Ekbom, B.; Jönsson, A.; Lindborg, R.; Olsson, O.; Rader, R.; et al. A framework to identify indicator species for ecosystem services in agricultural landscapes. Ecol. Indic. 2018, 91, 278–286. [Google Scholar] [CrossRef]
- Taylor, R.J.; Doran, N. Use of Terrestrial Invertebrates as Indicators of the Ecological Sustainability of Forest Management under the Montreal Process. J. Insect Conserv. 2001, 5, 221–231. [Google Scholar] [CrossRef]
- Gerlach, J.; Samways, M.J.; Pryke, J. Terrestrial invertebrates as bioindicators: An overview of available taxonomic groups. J. Insect Conserv. 2013, 17, 831–850. [Google Scholar] [CrossRef]
- Scott, A.G.; Oxford, G.S.; Selden, P.A. Epigeic spiders as ecological indicators of conservation value for peat bogs. Biol. Conserv. 2006, 127, 420–428. [Google Scholar] [CrossRef]
- Corcos, D.; Lami, F.; Nardi, D.; Boscutti, F.; Sigura, M.; Giannone, F.; Pantini, P.; Tagliapietra, A.; Busato, F.; Sibella, R.; et al. Cross-taxon congruence between predatory arthropods and plants across Mediterranean agricultural landscapes. Ecol. Indic. 2021, 123, 107366. [Google Scholar] [CrossRef]
- Argañaraz, C.I.; Rubio, G.D.; Rubio, M.; Castellarini, F. Ground-dwelling spiders in agroecosystems of the Dry Chaco: A rapid assessment of community shifts in response to land use changes. Biodiversity 2020, 21, 125–135. [Google Scholar] [CrossRef]
- Coddington, J.A.; Levi, H.W. Systematics and Evolution of Spiders (Araneae). Annu. Rev. Ecol. Syst. 1991, 22, 565–592. [Google Scholar] [CrossRef]
- Platnick, N.I. Dimensions of Biodiversity: Targeting Megadiverse Groups. The Living Planet in Crisis: Biodiversity Science and Policy; Columbia University Press: New York, NY, USA, 1999; p. 19. [Google Scholar]
- Clausen, I.H.S. The use of spiders (Araneae) as ecological indicators. Bull. Br. Arachnol. Soc. 1986, 7, 4. [Google Scholar]
- Maelfait, J.-P.; Hendrickx, F. Spiders as bio-indicators of anthropogenic stress in natural and semi-natural habitats in Flanders (Belgium): Some recent developments. In Proceedings of the 17th European Colloquium Arachnology, Edinburgh, UK, 1 September 1998; p. 8. [Google Scholar]
- Feest, A.; Cardoso, P. The comparison of site spider “biodiversity quality” in Portuguese protected areas. Ecol. Indic. 2012, 14, 229–235. [Google Scholar] [CrossRef]
- Štokmane, M.; Spungis, V. Diversity of grass-dwelling spiders (Arachnida: Araneae) in calcareous fens of the Coastal Lowland, Latvia. J. Insect Conserv. 2014, 18, 757–769. [Google Scholar] [CrossRef]
- Zografou, K.; Adamidis, G.C.; Komnenov, M.; Kati, V.; Sotirakopoulos, P.; Pitta, E.; Chatzaki, M. Diversity of spiders and orthopterans respond to intra-seasonal and spatial environmental changes. J. Insect Conserv. 2017, 21, 531–543. [Google Scholar] [CrossRef]
- Leonela, S.; Ana Elena de, V.; Fernando Pérez, M. Spiders as potential bioindicators of mountain grasslands health: The Argentine tarantula Grammostola vachoni (Araneae, Theraphosidae). Wildl. Res. 2018, 45, 64–71. [Google Scholar]
- Mahan, C.G.; Young, J.; Miller, B.J.; Saunders, M.C. Using Ecological Indicators and a Decision Support System for Integrated Ecological Assessment at Two National Park Units in the Mid-Atlantic Region, USA. Environ. Manag. 2015, 55, 508–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, C.; Xu, Y.; Bundy, A.; Grüss, A.; Coll, M.; Heymans, J.J.; Fulton, E.A.; Shannon, L.; Halouani, G.; Velez, L.; et al. Making ecological indicators management ready: Assessing the specificity, sensitivity, and threshold response of ecological indicators. Ecol. Indic. 2019, 105, 16–28. [Google Scholar] [CrossRef]
- Oliveira, U.; Brescovit, A.D.; Santos, A. Sampling effort and species richness assessment: A case study on Brazilian spiders. Biodivers. Conserv. 2017, 26, 1481–1493. [Google Scholar] [CrossRef]
- Cajaiba, R.L.; Périco, E.; Da Silva, W.B.; Vieira, T.B.; Dalzochio, M.S.; Bastos, R.; Cabral, J.A.; Santos, M. How informative is the response of Ground Beetles’ (Coleoptera: Carabidae) assemblages to anthropogenic land use changes? Insights for ecological status assessments from a case study in the Neotropics. Sci. Total Environ. 2018, 636, 1219–1227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cajaiba, R.L.; Cabral, J.A.; Santos, M. A Minimal Invasive Method to Forecast the Effects of Anthropogenic Disturbance on Tropical Cave Beetle Communities. Neotrop. Èntomol. 2015, 45, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.R.; Matthews, I.M. A review of extensive variation in the design of pitfall traps and a proposal for a standard pitfall trap design for monitoring ground-active arthropod biodiversity. Ecol. Evol. 2016, 6, 3953–3964. [Google Scholar] [CrossRef] [Green Version]
- Dale, V.H.; Beyeler, S.C. Challenges in the development and use of ecological indicators. Ecol. Indic. 2001, 1, 3–10. [Google Scholar] [CrossRef] [Green Version]
- Santos, M.; Cabral, J.A. Development of a stochastic dynamic model for ecological indicators’ prediction in changed Mediterra-nean agroecosystems of north-eastern Portugal. Ecol. Indic. 2004, 3, 285–303. [Google Scholar] [CrossRef]
- Oliver, I.B.; Andrew, J. A possible method for the rapid assessment of biodiversity. Conserv. Biol. 1993, 7, 562–568. [Google Scholar] [CrossRef]
- Sarkar, S.; Margules, C. Operationalizing biodiversity for conservation planning. J. Biosci. 2002, 27, 299–308. [Google Scholar] [CrossRef] [Green Version]
- Krell, F.-T. Parataxonomy vs. taxonomy in biodiversity studies—Pitfalls and applicability of ‘morphospecies’ sorting. Biodivers. Conserv. 2004, 13, 795–812. [Google Scholar] [CrossRef]
- Hortal, J.; Borges, P.; Gaspar, C. Evaluating the performance of species richness estimators: Sensitivity to sample grain size. J. Anim. Ecol. 2006, 75, 274–287. [Google Scholar] [CrossRef] [Green Version]
- Catalog, W.S. World Spider Catalog Version. Available online: https://wsc.nmbe.ch/archive (accessed on 22 January 2021).
- Swart, R.C.; Pryke, J.; Roets, F. Arthropod assemblages deep in natural forests show different responses to surrounding land use. Biodivers. Conserv. 2017, 27, 583–606. [Google Scholar] [CrossRef]
- Hammer, Ø.H.; David, A.T.; Ryan, P.D. Past: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Lowe, E.C.; Wolff, J.O.; Aceves-Aparicio, A.; Birkhofer, K.; Branco, V.V.; Cardoso, P.; Chichorro, F.; Fukushima, C.S.; Gonçalves-Souza, T.; Haddad, C.R.; et al. Towards establishment of a centralized spider traits database. J. Arachnol. 2020, 48, 103–109. [Google Scholar] [CrossRef]
- Carvalho, J.C.; Cardoso, P. Drivers of beta diversity in Macaronesian spiders in relation to dispersal ability. J. Biogeogr. 2014, 41, 1859–1870. [Google Scholar] [CrossRef]
- Carvalho, J.C.; Malumbres-Olarte, J.; Arnedo, M.A.; Crespo, L.C.; Domenech, M.; Cardoso, P. Taxonomic divergence and functional convergence in Iberian spider forest communities: Insights from beta diversity partitioning. J. Biogeogr. 2019, 47, 288–300. [Google Scholar] [CrossRef]
- Kanda, Y. Investigation of the freely available easy-to-use software ‘EZR’ for medical statistics. Bone Marrow Transpl. 2013, 48, 452–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graham, M.H. Confronting Multicollinearity in Ecological Multiple Regression. Ecology 2003, 84, 2809–2815. [Google Scholar] [CrossRef] [Green Version]
- Baselga, A. Partitioning the turnover and nestedness components of beta diversity. Glob. Ecol. Biogeogr. 2009, 19, 134–143. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Dufrêne, M.; Legendre, P. Species Assemblages and Indicator Species: The Need for a Flexible Asymmet-Rical Approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar]
- Cáceres, M.D.; Legendre, P. Associations between species and groups of sites: Indices and statistical inference. Ecology 2015, 90, 3566–3574. [Google Scholar] [CrossRef] [PubMed]
- Leote, P.; Cajaiba, R.L.; Cabral, J.; Brescovit, A.D.; Santos, M. Are data-mining techniques useful for selecting ecological indicators in biodiverse regions? Bridges between market basket analysis and indicator value analysis from a case study in the neotropics. Ecol. Indic. 2020, 109, 105833. [Google Scholar] [CrossRef]
- Cajaiba, R.L.; Périco, E.; Da Silva, W.B.; Leote, P.; Santos, M. Are Small Dung Beetles (Aphodiinae) useful for monitoring neotropical forests’ ecological status? Lessons from a preliminary case study in the Brazilian Amazon. For. Ecol. Manag. 2018, 429, 115–123. [Google Scholar] [CrossRef] [Green Version]
- Gotelli, N.J.; Ellison, A.M.; Dunn, R.R.; Sanders, N.J. Counting ants (Hymenoptera: Formicidae): Biodiversity sampling and statistical analysis for myrmecologists. Myrmecol. News 2011, 15, 7. [Google Scholar]
- Halaj, J.; Ross, D.; Moldenke, A. Habitat Structure and Prey Availability as Predictors of the Abundance and Community Organization of Spiders in Western Oregon Forest Canopies. J. Arachnol. 1998, 26, 203–220. [Google Scholar]
- Spears, L.R.; MacMahon, J.A. An experimental study of spiders in a shrub-steppe ecosystem: The effects of prey availability and shrub architecture. J. Arachnol. 2012, 40, 218–227. [Google Scholar] [CrossRef]
- Podgaiski, L.R.; Joner, F.; Lavorel, S.; Moretti, M.; Ibanez, S.; Mendonça, M.D.S.; Pillar, V.D. Spider Trait Assembly Patterns and Resilience under Fire-Induced Vegetation Change in South Brazilian Grasslands. PLoS ONE 2013, 8, e60207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Battirola, L.D.; Batistella, D.A.; Rosado-Neto, G.H.; Brescovit, A.D.; Marques, M.I. Spider assemblage (Arachnida: Araneae) associated with canopies of Vochysia divergens (Vochysiaceae) in the northern region of the Brazilian Pantanal. Zoologia 2016, 33. [Google Scholar] [CrossRef] [Green Version]
- Gómez, J.E.; Lohmiller, J.; Joern, A. Importance of vegetation structure to the assembly of an aerial web-building spider community in North American open grassland. J. Arachnol. 2016, 44, 28–35. [Google Scholar] [CrossRef]
- Rosas-Ramos, N.; Baños-Picón, L.; Tormos, J.; Asís, J.D. Farming system shapes traits and composition of spider assemblages in Mediterranean cherry orchards. PeerJ 2020, 8, e8856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hazzi, N.A.; Petrosky, A.; Karandikar, H.; Henderson, D.; Jiménez-Conejo, N. Effect of forest succession and microenvironmental variables on the abundance of two wandering spider species (Araneae: Ctenidae) in a montane tropical forest. J. Arachnol. 2020, 48, 140–145. [Google Scholar] [CrossRef]
- Santos, P.Z.F.; Crouzeilles, R.; Sansevero, J.B.B. Can agroforestry systems enhance biodiversity and ecosystem service provision in agricultural landscapes? A meta-analysis for the Brazilian Atlantic Forest. For. Ecol. Manag. 2019, 433, 140–145. [Google Scholar] [CrossRef]
- Baah-Acheamfour, M.; Carlyle, C.; Bork, E.W.; Chang, S.X. Trees increase soil carbon and its stability in three agroforestry systems in central Alberta, Canada. For. Ecol. Manag. 2014, 328, 131–139. [Google Scholar] [CrossRef]
- Navas, R.; Silva, R.J. Ecological Restoration Indicators in Agroforestry Systems in the Atlantic Forest. Ciência E Nat. 2016, 38, 656–664. [Google Scholar] [CrossRef] [Green Version]
- Baig, M.B.; Burgess, P.J.; Fike, J.H. Agroforestry for healthy ecosystems: Constraints, improvement strategies and extension in Paki-stan. Agrofor. Syst. 2020, 95, 995–1013. [Google Scholar] [CrossRef]
- Yolanda Bizuet-Flores, M.; Luisa Jiménez-Jiménez, M.; Zavala-Hurtado, A.; Corcuera, P. Diversity patterns of ground dwelling spiders (Arachnida: Araneae) in five prevailing plant communities of the Cuatro Ciénegas Basin, Coahuila, Mexico. Rev. Mex. Biodivers. 2015, 86, 153–163. [Google Scholar] [CrossRef]
- Hajian-Forooshani, Z.; Gonthier, D.J.; Marin, L.; Iverson, A.L.; Perfecto, I. Changes in species diversity of arboreal spiders in Mexican coffee agroecosystems: Untangling the web of local and landscape influences driving diversity. PeerJ 2014, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schroth, G.; Harvey, C.A. Biodiversity conservation in cocoa production landscapes: An overview. Biodivers. Conserv. 2007, 16, 2237–2244. [Google Scholar] [CrossRef]
- Rice, R.A.; Greenberg, R. Cacao Cultivation and the Conservation of Biological Diversity. AMBIO J. Hum. Environ. 2000, 29, 167–173. [Google Scholar] [CrossRef]
- Zschokke, S.; Hénaut, Y.; Benjamin, S.; García-Ballinas, J.A. Prey-capture strategies in sympatric web-building spiders. Can. J. Zool. 2006, 84, 964–973. [Google Scholar] [CrossRef]
- Sørensen, L.L. Composition and diversity of the spider fauna in the canopy of a montane forest in Tanzania. Biodivers. Conserv. 2004, 13, 437–452. [Google Scholar] [CrossRef]
- Prinzing, A.J. Use of Shifting Microclimatic Mosaics by Arthropods on Exposed Tree Trunks. Ann. Èntomol. Soc. Am. 2001, 94, 210–218. [Google Scholar] [CrossRef] [Green Version]
- Stenchly, K.; Clough, Y.; Buchori, D.; Tscharntke, T. Spider web guilds in cacao agroforestry—Comparing tree, plot and landscape-scale management. Divers. Distrib. 2011, 17, 748–756. [Google Scholar] [CrossRef]
- Biere, J.M.; Uetz, G.W. Web Orientation in the Spider Micrathena Gracilis (Araneae: Araneidae). Ecology 1981, 62, 336–344. [Google Scholar] [CrossRef]
- De Souza, A.L.T.; Martins, R.P. Foliage Density of Branches and Distribution of Plant-Dwelling Spiders1. Biotropica 2005, 37, 416–420. [Google Scholar] [CrossRef]
- Kaur, H.; Torma, A.; Gallé-Szpisjak, N.; Šeat, J.; Lőrinczi, G.; Módra, G.; Gallé, R. Road verges are important secondary habitats for grassland arthropods. J. Insect Conserv. 2019, 23, 899–907. [Google Scholar] [CrossRef] [Green Version]
- Oxbrough, A.G.; Gittings, T.; O’Halloran, J.; Giller, P.S.; Smith, G.F. Structural indicators of spider communities across the forest plantation cycle. For. Ecol. Manag. 2005, 212, 171–183. [Google Scholar] [CrossRef] [Green Version]
- Erdős, L.; Tölgyesi, C.; Horzse, M.; Tolnay, D.; Hurton, Á.; Schulcz, N.; Kormoczi, L.; Lengyel, A.; Batori, Z. Habitat complexity of the Pannonian forest-steppe zone and its nature conservation implications. Ecol. Complex 2014, 17, 107–118. [Google Scholar] [CrossRef]
- Cardoso, P.; Gaspar, C.; Pereira, L.C.; Silva, I.; Henriques, S.S.; da Silva, R.R.; Sousa, P. Assessing spider species richness and composition in Mediterranean cork oak forests. Acta Oecol. 2008, 33, 114–127. [Google Scholar] [CrossRef]
- Cardoso, P.; Henriques, S.S.; Gaspar, C.; Crespo, L.C.; Carvalho, R.; Schmidt, J.B.; Sousa, P.; Szuts, T. Species richness and composition assessment of spiders in a Mediterranean scrubland. J. Insect Conserv. 2009, 13, 45–55. [Google Scholar] [CrossRef] [Green Version]
- Delgado de la Flor, Y.A.; Perry, K.I.; Turo, K.J.; Parker, D.M.; Thompson, J.L.; Gardiner, M.M. Local and landscape-scale environmental filters drive the functional diversity and taxonomic composition of spiders across urban greenspaces. J. Appl. Ecol. 2020, 57, 1570–1580. [Google Scholar] [CrossRef]
- Cattin, M.-F.; Blandenier, G.; Banašek-Richter, C.; Bersier, L.-F. The impact of mowing as a management strategy for wet meadows on spider (Araneae) communities. Biol. Conserv. 2003, 113, 179–188. [Google Scholar] [CrossRef] [Green Version]
- Mouillot, D.; Graham, N.; Villéger, S.; Mason, N.W.; Bellwood, D.R. A functional approach reveals community responses to disturbances. Trends Ecol. Evol. 2013, 28, 167–177. [Google Scholar] [CrossRef]
- Petchey, O.L. On the statistical significance of functional diversity effects. Funct. Ecol. 2004, 18, 297–303. [Google Scholar] [CrossRef]
- Tilman, D.; Knops, J.; Wedin, D.; Reich, P.; Ritchie, M.; Siemann, E. The Influence of Functional Diversity and Composition on Eco-system Processes. Science 1997, 277, 1300. [Google Scholar] [CrossRef] [Green Version]
- Loreau, M.; Naeem, S.N.; Inchausti, P.; Bengtsson, J.; Grime, J.P.; Hector, A.; Hooper, D.U.; Huston, M.A.; Raffaelli, D.G.; Schmid, B.; et al. Biodiversity and Ecosystem Functioning: Current Knowledge and Future Challenges. Science 2001, 294, 804–808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barsoum, N.; Fuller, L.; Ashwood, F.; Reed, K.; Bonnet-Lebrun, A.-S.; Leung, F. Ground-dwelling spider (Araneae) and carabid beetle (Coleoptera: Carabidae) community assemblages in mixed and monoculture stands of oak (Quercus robur L./Quercus petraea (Matt.) Liebl.) and Scots pine (Pinus sylvestris L.). For. Ecol. Manag. 2014, 321, 29–41. [Google Scholar] [CrossRef]
- Schmidt, M.H.; Roschewitz, I.; Thies, C.; Tscharntke, T. Differential effects of landscape and management on diversity and density of ground-dwelling farmland spiders. J. Appl. Ecol. 2005, 42, 281–287. [Google Scholar] [CrossRef]
- Oxbrough, A.; French, V.; Irwin, S.; Kelly, T.C.; Smiddy, P.; O’Halloran, J. Can mixed species stands enhance arthropod diversity in plantation forests? For. Ecol. Manag. 2012, 270, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Peng, M.-H.; Hung, Y.-C.; Liu, K.-L.; Neoh, K.-B. Landscape configuration and habitat complexity shape arthropod assemblage in urban parks. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kostylev, V.E.; Erlandsson, J.; Ming, M.Y.; Williams, G.A. The relative importance of habitat complexity and surface area in assessing biodiversity: Fractal application on rocky shores. Ecol. Complex 2005, 2, 272–286. [Google Scholar] [CrossRef]
- LaRue, E.A.; Hardiman, B.S.; Elliott, J.M.; Fei, S. Structural diversity as a predictor of ecosystem function. Environ. Res. Lett. 2019, 14, 114011. [Google Scholar] [CrossRef]
- Winter, S.; Möller, G.C. Microhabitats in lowland beech forests as monitoring tool for nature conservation. For. Ecol. Manag. 2008, 255, 1251–1261. [Google Scholar] [CrossRef]
- Maseko, M.S.T.; Zungu, M.M.; Ehlers Smith, D.A.; Ehlers Smith, Y.C.; Downs, C.T. High microhabitat heterogeneity drives high func-tional traits in forest birds in five protected forest areas in the urban mosaic of Durban, South Africa. Glob. Ecol. Conserv. 2019, 18, e00645. [Google Scholar] [CrossRef]
- Bogoni, J.A.; Bogoni, T.C.; Graipel, M.E.; Marinho, J.R. The Influence of Landscape and Microhabitat on the Diversity of Large- and Medium-Sized Mammals in Atlantic Forest Remnants in a Matrix of Agroecosystem and Silviculture. ISRN For. 2013, 2013, 1–13. [Google Scholar] [CrossRef]
- Mammola, S.; Cardoso, P.; Angyal, D.; Balázs, G.; Blick, T.; Brustel, H.; Carter, J.; Ćurčić, S.; Danflous, S.; Dányi, L.; et al. Local- versus broad-scale environmental drivers of continental β-diversity patterns in subterranean spider communities across Europe. Proc. R. Soc. B Biol. Sci. 2019, 286, 20191579. [Google Scholar] [CrossRef] [Green Version]
- Oxbrough, A.R.; Paul, M. Páramo spiders of Volcán Chiles. In The Ecology of Volcán Chiles: High-Altitude Ecosystems on the Ecuador-Columbia Border; Ramsay, P.M., Ed.; Pebble & Shell: Plymouth, UK, 2001; p. 9. [Google Scholar]
- Lapinski, W.; Tschapka, M. Habitat use in an assemblage of Central American wandering spiders. J. Arachnol. 2013, 41, 151–159. [Google Scholar] [CrossRef]
- Luiz Pereira Souza, J.; Oliveira Fernandes, I. Do spatial and temporal scales affect the efficiency of surrogates in ant monitoring on the hydroelectric power-plant area in Brazilian Amazon? Ecol. Indic. 2021, 121, 107158. [Google Scholar] [CrossRef]
- García-Martínez, M.A.; Martínez-Tlapa, D.L.; Pérez-Toledo, G.R.; Quiroz-Robledo, L.N.; Castaño-Meneses, G.; Laborde, J.; Valenzuela-González, J.E. Taxonomic, Species and Functional Group Diversity of Ants in a Tropical Anthropogenic Landscape. Trop. Conserv. Sci. 2015, 8, 1017–1032. [Google Scholar] [CrossRef]
- Van Rijn, I.; Neeson, T.M.; Mandelik, Y. Reliability and refinement of the higher taxa approach for bee richness and composition assessments. Ecol. Appl. 2015, 25, 88–98. [Google Scholar] [CrossRef]
- Cardoso, P.; Silva, I.; De Oliveira, N.G.; Serrano, A.R. Higher taxa surrogates of spider (Araneae) diversity and their efficiency in conservation. Biol. Conserv. 2004, 117, 453–459. [Google Scholar] [CrossRef]
- Gaston, K.J. Biodiversity—Measurement. Prog. Phys. Geogr. Earth Environ. 1994, 18, 565–574. [Google Scholar] [CrossRef]
- de Oliveira, S.S.; Ortega, J.C.G.; Ribas, L.G.d.S.; Lopes, V.G.; Bini, L.M. Higher taxa are sufficient to represent biodiversity patterns. Ecol. Indic. 2020, 111, 105994. [Google Scholar] [CrossRef]
- Fernandes, C.; Cabral, J.A.; Crespí, A.L.; Hughes, S.J.; Santos, M. Converting simple vegetation surveys in functional dynamics. Acta Oecol. 2013, 48, 37–46. [Google Scholar] [CrossRef]
- Standen, V. The adequacy of collecting techniques for estimating species richness of grassland invertebrates. J. Appl. Ecol. 2000, 37, 884–893. [Google Scholar] [CrossRef] [Green Version]
- Privet, K.; Vedel, V.; Fortunel, C.; Orivel, J.; Martinez, Q.; Cerdan, A.; Baraloto, C.; Pétillon, J. Relative Efficiency of Pitfall Trapping vs. Nocturnal Hand Collecting in Assessing Soil-Dwelling Spider Diversity along A Structural Gradient of Neotropical Habitats. Diversity 2020, 12, 81. [Google Scholar] [CrossRef] [Green Version]
- Tourinho, A.L.; Lo-Man-Hung, N. Standardized Sampling Methods and Protocols for Harvestman and Spider Assemblages. In Measuring Arthropod Biodiversity: A Handbook of Sampling Methods; Santos, J.C., Fernandes, G.W., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 365–400. [Google Scholar]
- Dias, S.C.; Carvalho, L.S.; Bonaldo, A.; Brescovit, A.D. Refining the establishment of guilds in Neotropical spiders (Arachnida: Araneae). J. Nat. Hist. 2009, 44, 219–239. [Google Scholar] [CrossRef]
- Benítez-Malvido, J.; Martínez-Falcón, A.P.; Durán-Barrón, C.G. Diversity metrics of spider communities associated with an un-derstorey plant in tropical rain forest fragments. J. Trop. Ecol. 2020, 36, 47–55. [Google Scholar] [CrossRef]
- Munévar, A.; Cardoso, P.; Espejo, Y.M.P.; Zurita, G.A. Spiders (Arachnida: Araneae) in the semideciduous Atlantic Forest: An ecological and morphological trait dataset for functional studies. Biodivers. Data J. 2020, 8, e49889. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ecosystems | PF | SF | IF | AF |
|---|---|---|---|---|
| PF | 0 | - | - | - |
| SF | 9.37 *** | 0 | - | - |
| IF | 7.32 *** | 11.88 *** | 0 | - |
| AF | 14.39 *** | 12.39 *** | 14.80 *** | 0 |
| PA | 18.16 *** | 17.51 *** | 15.67 *** | 7.96 *** |
| Ecosystems | PF | SF | IF | AF |
|---|---|---|---|---|
| PF | 0 | - | - | - |
| SF | 8.93 *** | 0 | - | - |
| IF | 4.34 *** | 9.78 *** | 0 | - |
| AF | 13.39 *** | 9.88 *** | 12.34 *** | 0 |
| PA | 12.46 *** | 15.21 *** | 2.54 * | 23.97 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gonzalez, D.C.; Cajaiba, R.L.; Périco, E.; da Silva, W.B.; Brescovite, A.D.; Crespi, A.M.L.; Santos, M. Assessing Ecological Disturbance in Neotropical Forest Landscapes Using High-Level Diversity and High-Level Functionality: Surprising Outcomes from a Case Study with Spider Assemblages. Land 2021, 10, 758. https://doi.org/10.3390/land10070758
Gonzalez DC, Cajaiba RL, Périco E, da Silva WB, Brescovite AD, Crespi AML, Santos M. Assessing Ecological Disturbance in Neotropical Forest Landscapes Using High-Level Diversity and High-Level Functionality: Surprising Outcomes from a Case Study with Spider Assemblages. Land. 2021; 10(7):758. https://doi.org/10.3390/land10070758
Chicago/Turabian StyleGonzalez, Darinka Costa, Reinaldo Lucas Cajaiba, Eduardo Périco, Wully Barreto da Silva, Antônio Domingos Brescovite, António Maria Luis Crespi, and Mário Santos. 2021. "Assessing Ecological Disturbance in Neotropical Forest Landscapes Using High-Level Diversity and High-Level Functionality: Surprising Outcomes from a Case Study with Spider Assemblages" Land 10, no. 7: 758. https://doi.org/10.3390/land10070758