
Four Years Continuous Monitoring Reveals Different Effects of Urban Constructed Wetlands on Bats

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Bat Monitoring and Acoustic Analysis
2.3. Statistical Analysis
3. Results
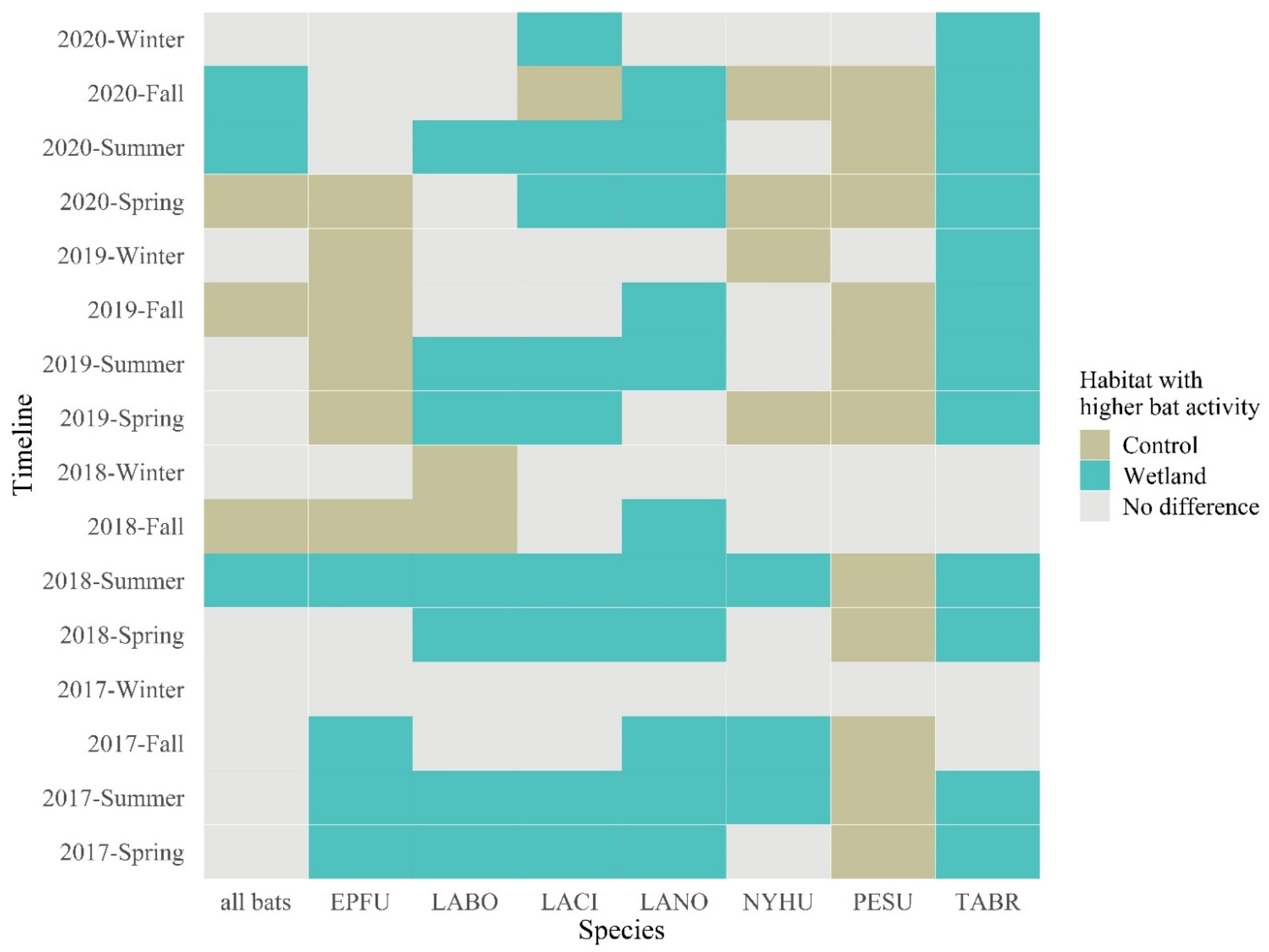
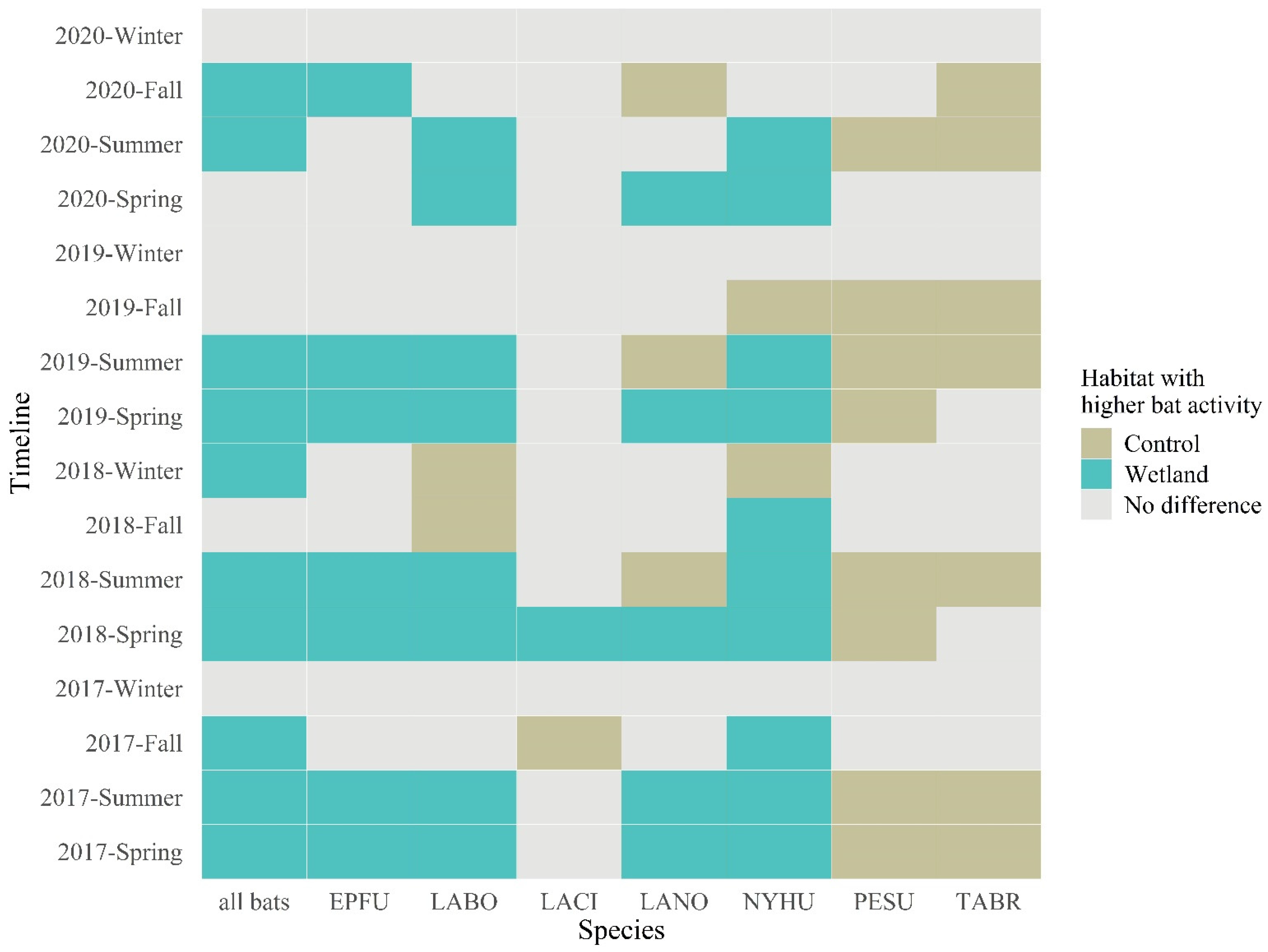
3.1. Wetland vs. Control Bat Activity Comparison by Wilcoxon Tests
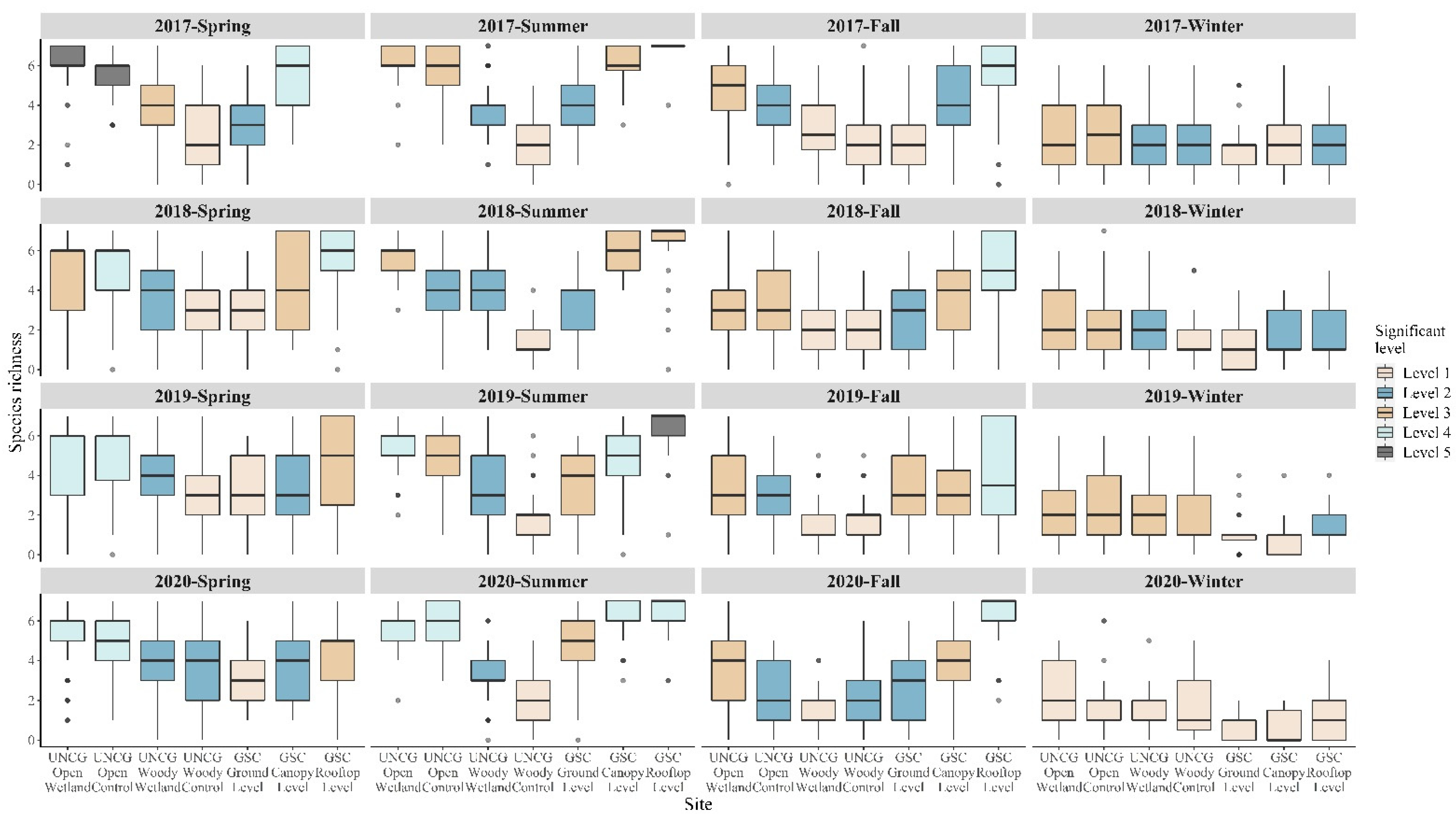
3.2. Species Richness Comparison by Generalized Linear Models
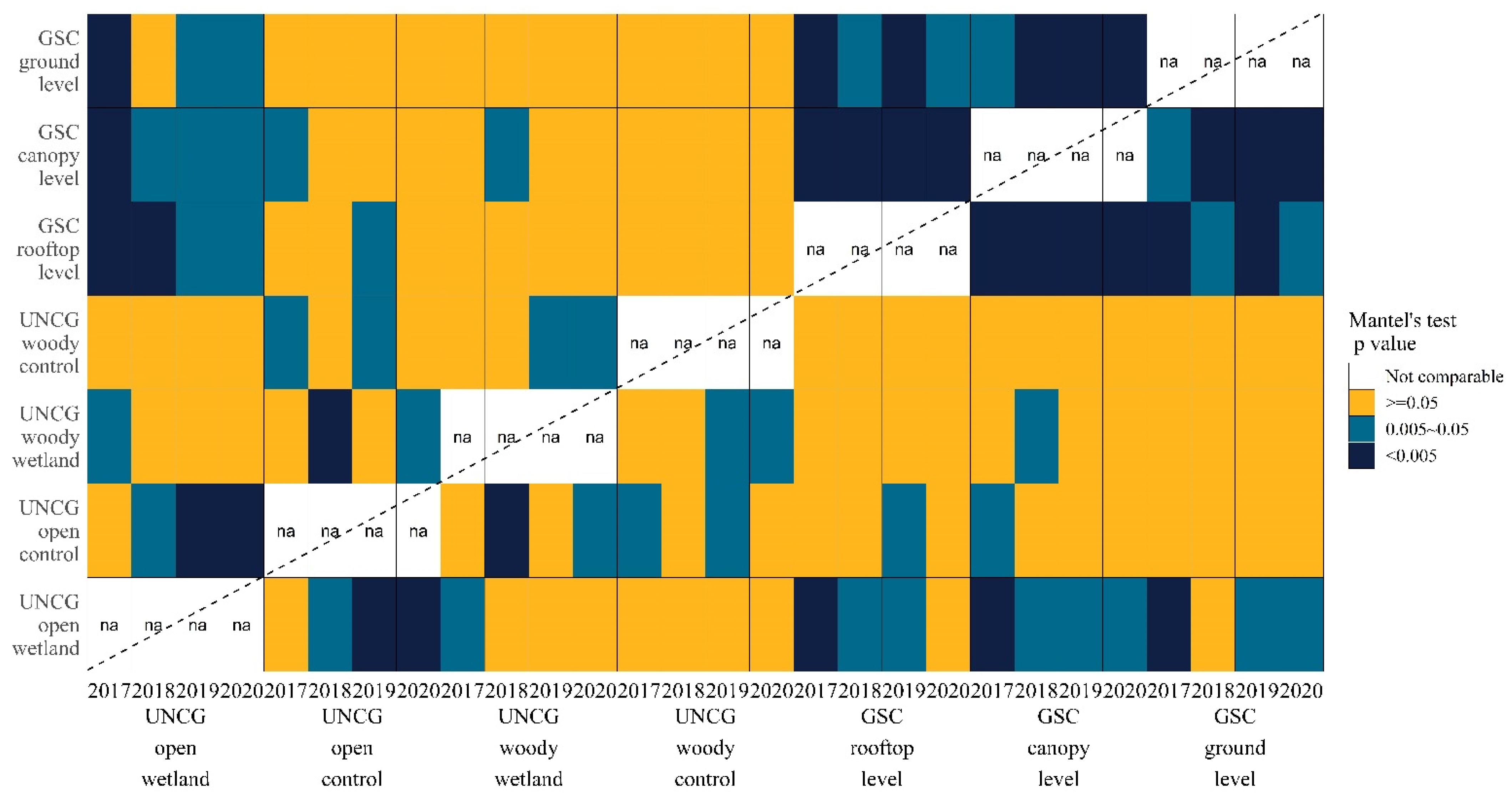
3.3. Community Composition Comparison by Mantel’s Tests
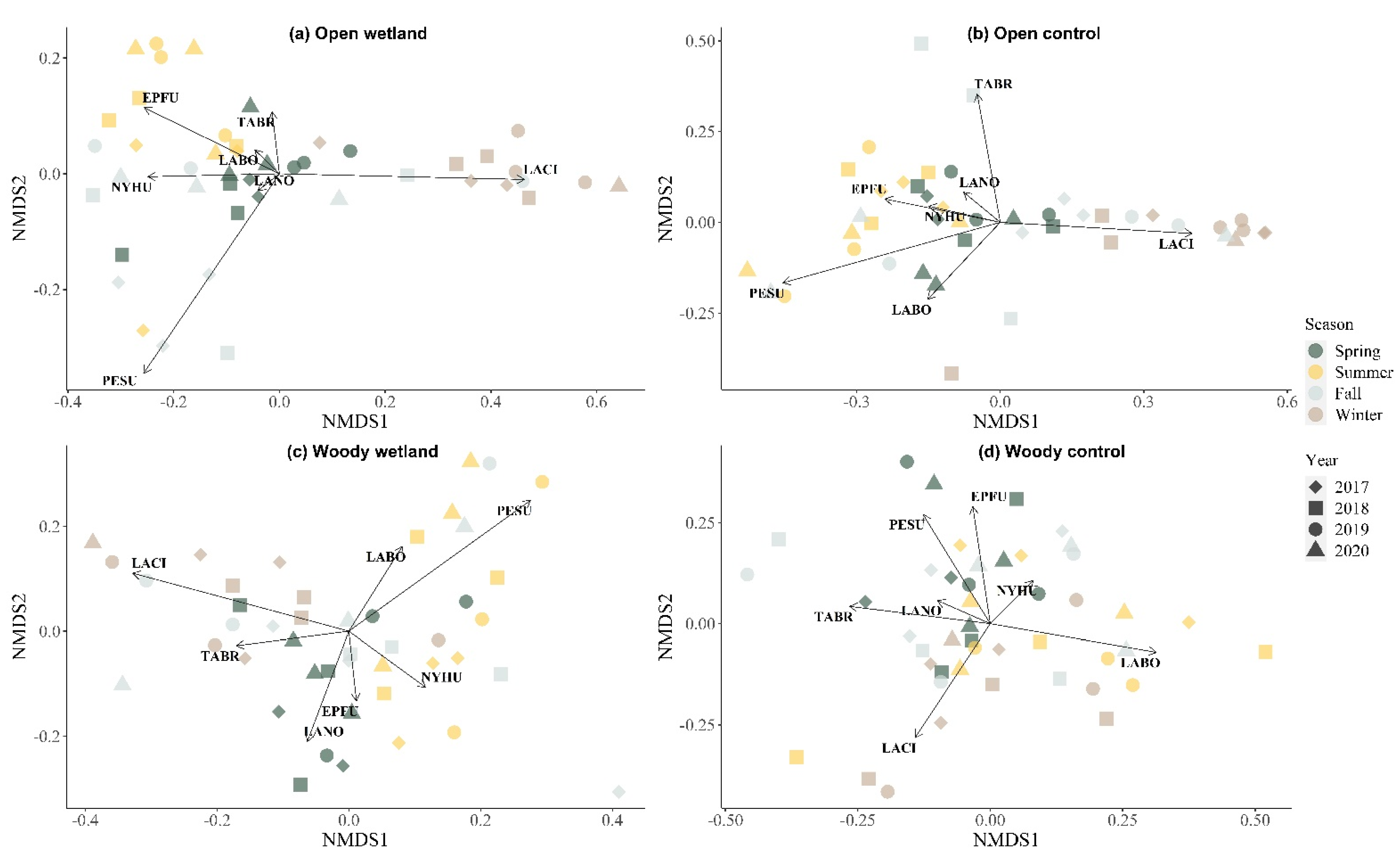
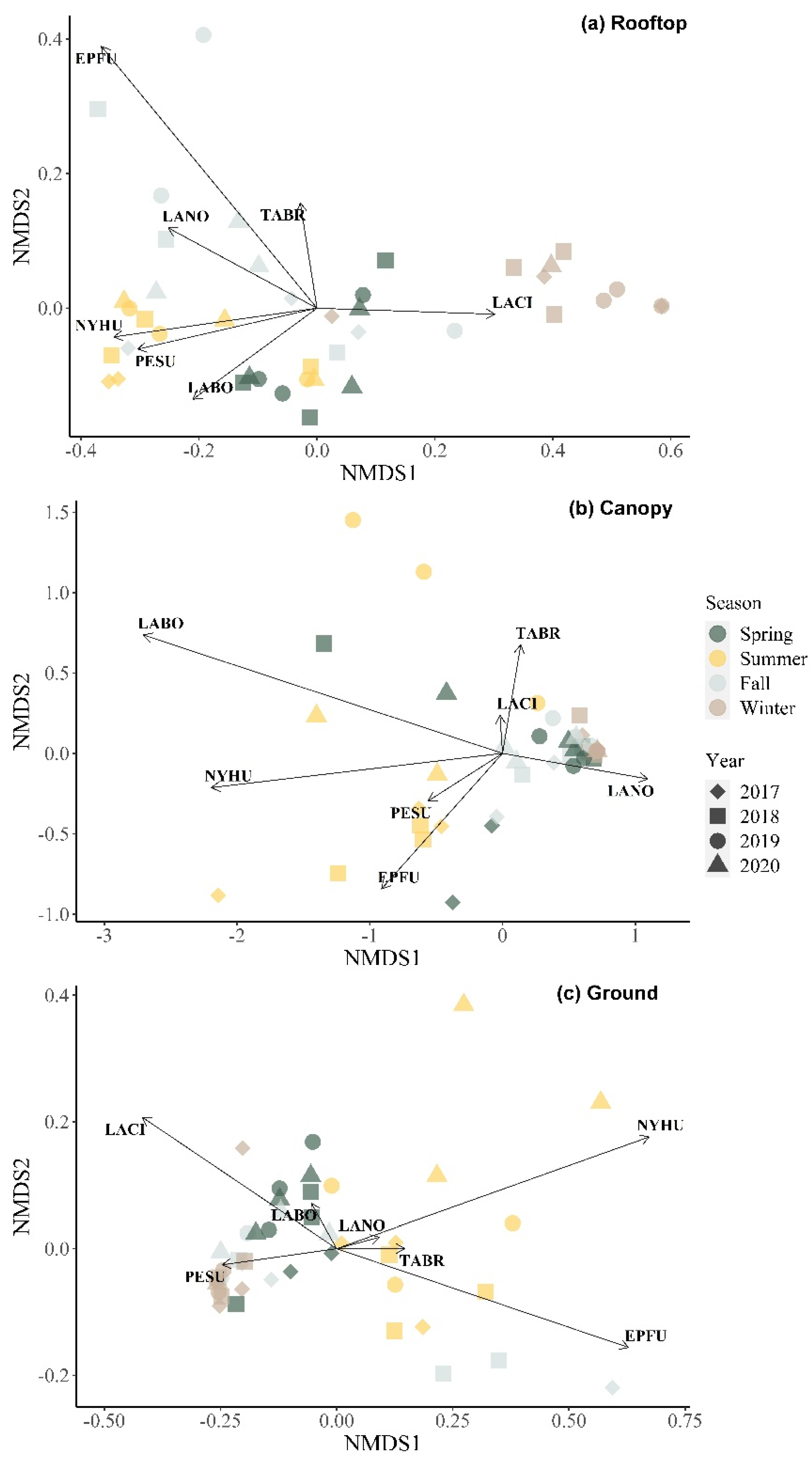
3.4. Community Composition Changes over Time by NMDS
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brinson, M.M. A Hydrogeomorphic Clasification for Wetlands; U.S. Environmental Laboratory: Vicksburg, MS, USA; Engineer Research and Development Center: Vicksburg, MS, USA, 1993. Available online: https://hdl.handle.net/11681/6483 (accessed on 10 September 2021).
- Davidson, N.C. How much wetland has the world lost? Long-term and recent trends in global wetland area. Mar. Freshw. Res. 2014, 65, 934–941. [Google Scholar] [CrossRef]
- Gardner, R.C.; Finlayson, C. Global Wetland Outlook: State of the World’s Wetlands and Their Services to People; Social Science Research Network: Rochester, NY, USA, 2018; Available online: https://ssrn.com/abstract=3261606 (accessed on 10 September 2021).
- Finlayson, C.M.; Davidson, N.C.; Spiers, A.G.; Stevenson, N.J. Global wetland inventory—Current status and future priorities. Mar. Freshw. Res. 1999, 50, 717–727. [Google Scholar] [CrossRef]
- Verhoeven, J.T.A.; Setter, T.L. Agricultural use of wetlands: Opportunities and limitations. Ann. Bot. 2010, 105, 155–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ingraham, M.W.; Foster, S.G. The value of ecosystem services provided by the U.S. National Wildlife Refuge System in the contiguous U.S. Ecol. Econ. 2008, 67, 608–618. [Google Scholar] [CrossRef]
- Woodward, R.T.; Wui, Y.-S. The economic value of wetland services: A meta-analysis. Ecol. Econ. 2001, 37, 257–270. [Google Scholar] [CrossRef]
- Mitsch, W.J.; Gosselink, J.G. The value of wetlands: Importance of scale and landscape setting. Ecol. Econ. 2000, 35, 25–33. [Google Scholar] [CrossRef]
- Russi, D.; Brink ten, P.; Farmer, A.; Badura, T.; Coates, D.; Forster, J.; Kumar, R.; Davidson, N. The Economics of Ecosystems and Biodiversity for Water and Wetlands. 2013. Available online: http://www.teebweb.org/wp-content/uploads/2013/04/TEEB_WaterWetlands_Report_2013.pdf (accessed on 10 September 2021).
- Zhang, C.; Wen, L.; Wang, Y.; Liu, C.; Zhou, Y.; Lei, G. Can Constructed Wetlands be Wildlife Refuges? A Review of Their Potential Biodiversity Conservation Value. Sustainability 2020, 12, 1442. [Google Scholar] [CrossRef] [Green Version]
- Kunz, T.H.; De Torrez, E.B.; Bauer, D.; Lobova, T.; Fleming, T.H. Ecosystem services provided by bats. Ann. N. Y. Acad. Sci. 2011, 1223, 1–38. [Google Scholar] [CrossRef]
- Boyles, J.G.; Cryan, P.M.; McCracken, G.F.; Kunz, T.H. Economic Importance of Bats in Agriculture. Science 2011, 332, 41–42. [Google Scholar] [CrossRef]
- Rodríguez-San Pedro, A.; Allendes, J.L.; Beltrán, C.A.; Chaperon, P.N.; Saldarriaga-Córdoba, M.M.; Silva, A.X.; Grez, A.A. Quantifying ecological and economic value of pest control services provided by bats in a vineyard landscape of central Chile. Agric. Ecosyst. Environ. 2020, 302, 107063. [Google Scholar] [CrossRef]
- de Jesús Crespo, R.; Méndez Lázaro, P.; Yee, S.H. Linking Wetland Ecosystem Services to Vector-borne Disease: Dengue Fever in the San Juan Bay Estuary, Puerto Rico. Wetlands 2019, 39, 1281–1293. [Google Scholar] [CrossRef]
- Tremlett, C.J.; Moore, M.; Chapman, M.A.; Zamora-Gutierrez, V.; Peh, K.S.-H. Pollination by bats enhances both quality and yield of a major cash crop in Mexico. J. Appl. Ecol. 2020, 57, 450–459. [Google Scholar] [CrossRef] [Green Version]
- Medellin, R.A.; Wiederholt, R.; Lopez-Hoffman, L. Conservation relevance of bat caves for biodiversity and ecosystem services. Biol. Conserv. 2017, 211, 45–50. [Google Scholar] [CrossRef]
- Jones, G.; Jacobs, D.; Kunz, T.; Willig, M.; Racey, P. Carpe noctem: The importance of bats as bioindicators. Endanger. Species Res. 2009, 8, 93–115. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Kalcounis-Rueppell, M. Separating the effects of water quality and urbanization on temperate insectivorous bats at the landscape scale. Ecol. Evol. 2018, 8, 667–678. [Google Scholar] [CrossRef]
- Russo, D.; Salinas-Ramos, V.B.; Cistrone, L.; Smeraldo, S.; Bosso, L.; Ancillotto, L. Do We Need to Use Bats as Bioindicators? Biology 2021, 10, 693. [Google Scholar] [CrossRef] [PubMed]
- Mickleburgh, S.P.; Hutson, A.M.; Racey, P.A. A review of the global conservation status of bats. Oryx 2002, 36, 18–34. [Google Scholar] [CrossRef] [Green Version]
- Stahlschmidt, P.; Pätzold, A.; Ressl, L.; Schulz, R.; Brühl, C.A. Constructed wetlands support bats in agricultural landscapes. Basic Appl. Ecol. 2012, 13, 196–203. [Google Scholar] [CrossRef]
- Sirami, C.; Jacobs, D.S.; Cumming, G.S. Artificial wetlands and surrounding habitats provide important foraging habitat for bats in agricultural landscapes in the Western Cape, South Africa. Biol. Conserv. 2013, 164, 30–38. [Google Scholar] [CrossRef]
- Fukui, D.; Murakami, M.; Nakano, S.; Aoi, T. Effect of emergent aquatic insects on bat foraging in a riparian forest. J. Anim. Ecol. 2006, 75, 1252–1258. [Google Scholar] [CrossRef] [PubMed]
- Salvarina, I. Bats and aquatic habitats: A review of habitat use and anthropogenic impacts. Mammal Rev. 2016, 46, 131–143. [Google Scholar] [CrossRef]
- Blakey, R.V.; Law, B.S.; Straka, T.M.; Kingsford, R.T.; Milne, D.J. Importance of wetlands to bats on a dry continent: A review and meta-analysis. Hystrix Ital. J. Mammal. 2018, 12, 41–52. [Google Scholar] [CrossRef]
- Seibold, S.; Buchner, J.; Bässler, C.; Muller, J. Ponds in acidic mountains are more important for bats in providing drinking water than insect prey. J. Zool. 2013, 290, 302–308. [Google Scholar] [CrossRef]
- Lintott, P.R.; Bunnefeld, N.; Park, K.J. Opportunities for improving the foraging potential of urban waterways for bats. Biol. Conserv. 2015, 191, 224–233. [Google Scholar] [CrossRef]
- Ober, H.K.; Hayes, J.P. Influence of Vegetation on Bat Use of Riparian Areas at Multiple Spatial Scales. J. Wildl. Manag. 2008, 72, 396–404. [Google Scholar] [CrossRef]
- Sofaer, H.R.; Skagen, S.K.; Barsugli, J.J.; Rashford, B.S.; Reese, G.C.; Hoeting, J.A.; Wood, A.W.; Noon, B.R. Projected Wetland Densities under Climate Change: Habitat Loss but Little Geographic Shift in Conservation Strategy. Ecol. Appl. 2016, 26, 1677–1692. [Google Scholar] [CrossRef]
- Straka, T.M.; Lentini, P.E.; Lumsden, L.F.; Wintle, B.A.; van der Ree, R. Urban bat communities are affected by wetland size, quality, and pollution levels. Ecol. Evol. 2016, 6, 4761–4774. [Google Scholar] [CrossRef] [Green Version]
- Naidoo, S.; Vosloo, D.; Schoeman, M.C. Foraging at Wastewater Treatment Works Increases the Potential for Metal Accumulation in an Urban Adapter, the Banana Bat (Neoromicia nana). Afr. Zool. 2013, 48, 39–55. [Google Scholar] [CrossRef]
- Mendes, E.S.; Pereira, M.J.R.; Marques, S.F.; Fonseca, C. A Mosaic of Opportunities? Spatio-Temporal Patterns of Bat Diversity and Activity in a Strongly Humanized Mediterranean Wetland. Eur. J. Wildl. Res. 2014, 60, 651–664. [Google Scholar] [CrossRef]
- Lookingbill, T.R.; Elmore, A.J.; Engelhardt, K.A.; Churchill, J.B.; Gates, J.E.; Johnson, J.B. Influence of wetland networks on bat activity in mixed-use landscapes. Biol. Conserv. 2010, 143, 974–983. [Google Scholar] [CrossRef]
- Parker, K.A.; Springall, B.T.; Garshong, R.A.; Malachi, A.N.; Dorn, L.E.; Costa-Terryll, A.; Mathis, R.A.; Lewis, A.N.; MacCheyne, C.L.; Davis, T.T.; et al. Rapid Increases in Bat Activity and Diversity after Wetland Construction in an Urban Ecosystem. Wetlands 2019, 39, 717–727. [Google Scholar] [CrossRef]
- Giosa, E.; Mammides, C.; Zotos, S. The importance of artificial wetlands for birds: A case study from Cyprus. PLoS ONE 2018, 13, e0197286. [Google Scholar] [CrossRef] [PubMed]
- Menzel, J.M.; Menzel, M.A.; Kilgo, J.C.; Ford, W.M.; Edwards, J.W. Bat response to Carolina bays and wetland restoration in the southeastern U.S. Coastal Plain. Wetlands 2005, 25, 542–550. [Google Scholar] [CrossRef]
- Whigham, D.F. Ecological issues related to wetland preservation, restoration, creation and assessment. Sci. Total Environ. 1999, 240, 31–40. [Google Scholar] [CrossRef]
- Gopal, B. Natural and Constructed Wetlands for Wastewater Treatment: Potentials and Problems. Water Sci. Technol. 1999, 40, 27–35. [Google Scholar] [CrossRef]
- Wiegleb, G.; Dahms, H.-U.; Byeon, W.; Choi, G. To What Extent Can Constructed Wetlands Enhance Biodiversity? Int. J. Environ. Sci. Technol. 2017, 8, 561–569. [Google Scholar] [CrossRef] [Green Version]
- Sievers, M.; Parris, K.M.; Swearer, S.E.; Hale, R. Stormwater wetlands can function as ecological traps for urban frogs. Ecol. Appl. 2018, 28, 1106–1115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haberl, R.; Perfler, R.; Mayer, H. Constructed Wetlands in Europe. Water Water Sci. Technol. 1995, 32, 305–315. [Google Scholar] [CrossRef]
- Sievers, M.; Hale, R.; Parris, K.M.; Swearer, S.E. Impacts of Human-Induced Environmental Change in Wetlands on Aquatic Animals. Biol. Rev. Camb. Philos. Soc. 2018, 93, 529–554. [Google Scholar] [CrossRef] [Green Version]
- Robertson, B.A.; Hutto, R.L. A Framework for Understanding Ecological Traps and an Evaluation of Existing Evidence. Ecology 2006, 87, 1075–1085. [Google Scholar] [CrossRef] [Green Version]
- Hale, R.; Swearer, S.E. Ecological traps: Current evidence and future directions. Proc. Biol. Sci. 2016, 283, 20152647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Severns, P.M. Habitat restoration facilitates an ecological trap for a locally rare, wetland-restricted butterfly. Insect Conserv. Divers. 2011, 4, 184–191. [Google Scholar] [CrossRef]
- Sievers, M.; Hale, R.; Swearer, S.E.; Parris, K.M. Contaminant mixtures interact to impair predator-avoidance behaviours and survival in a larval amphibian. Ecotoxicol. Environ. Saf. 2018, 161, 482–488. [Google Scholar] [CrossRef]
- Clarke-Wood, B.K.; Jenkins, K.M.; Law, B.S.; Blakey, R.V. The ecological response of insectivorous bats to coastal lagoon degradation. Biol. Conserv. 2016, 202, 10–19. [Google Scholar] [CrossRef]
- Laverty, T.M.; Berger, J. Do bats seek clean water? A perspective on biodiversity from the Namib Desert. Biol. Conserv. 2020, 248, 108686. [Google Scholar] [CrossRef]
- Sharley, D.J.; Sharp, S.M.; Marshall, S.; Jeppe, K.; Pettigrove, V.J. Linking urban land use to pollutants in constructed wetlands: Implications for stormwater and urban planning. Landsc. Urban Plan. 2017, 162, 80–91. [Google Scholar] [CrossRef]
- Paing, J.; Guilbert, A.; Gagnon, V.; Chazarenc, F. Effect of climate, wastewater composition, loading rates, system age and design on performances of French vertical flow constructed wetlands: A survey based on 169 full scale systems. Ecol. Eng. 2015, 80, 46–52. [Google Scholar] [CrossRef]
- Hijosa-Valsero, M.; Sidrach-Cardona, R.; Becares, E. Comparison of interannual removal variation of various constructed wetland types. Sci. Total Environ. 2012, 430, 174–183. [Google Scholar] [CrossRef]
- Saggaï, M.M.; Ainouche, A.; Nelson, M.; Cattin, F.; El Amrani, A. Long-term investigation of constructed wetland wastewater treatment and reuse: Selection of adapted plant species for metaremediation. J. Environ. Manag. 2017, 201, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Reinartz, J.A.; Warne, E.L. Development of vegetation in small created wetlands in southeastern Wisconsin. Wetlands 1993, 13, 153–164. [Google Scholar] [CrossRef]
- Spieles, D.J.; Coneybeer, M.; Horn, J. Community Structure and Quality after 10 Years in Two Central Ohio Mitigation Bank Wetlands. Environ. Manag. 2006, 38, 837–852. [Google Scholar] [CrossRef] [PubMed]
- Denton, R.D.; Richter, S.C. Amphibian communities in natural and constructed ridge top wetlands with implications for wetland construction. J. Wildl. Manag. 2013, 77, 886–896. [Google Scholar] [CrossRef]
- Schad, A.N.; Kennedy, J.H.; Dick, G.O.; Dodd, L. Aquatic macroinvertebrate richness and diversity associated with native submerged aquatic vegetation plantings increases in longer-managed and wetland-channeled effluent constructed urban wetlands. Wetl. Ecol. Manag. 2020, 28, 461–477. [Google Scholar] [CrossRef]
- Silva De Araújo, M.L.V.; Bernard, E. Green remnants are hotspots for bat activity in a large Brazilian urban area. Urban Ecosyst. 2016, 19, 287–296. [Google Scholar] [CrossRef]
- Tena, E.; Fandos, G.; De Paz, Ó.; De La Peña, R.; Tellería, J.L. Size does matter: Passive sampling in urban parks of a regional bat assemblage. Urban Ecosyst. 2020, 23, 227–234. [Google Scholar] [CrossRef]
- Krauel, J.J.; Lebuhn, G. Patterns of Bat Distribution and Foraging Activity in a Highly Urbanized Temperate Environment. PLoS ONE 2016, 11, e0168927. [Google Scholar] [CrossRef]
- Kalcounis, M.C.; Hobson, K.A.; Brigham, R.M.; Hecker, K.R. Bat Activity in the Boreal Forest: Importance of Stand Type and Vertical Strata. J. Mammal. 1999, 80, 673–682. [Google Scholar] [CrossRef] [Green Version]
- Springall, B.T.; Li, H.; Kalcounis-Rueppell, M.C. The In-Flight Social Calls of Insectivorous Bats: Species Specific Behaviors and Contexts of Social Call Production. Front. Ecol. Evol. 2019, 7, 441. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Crihfield, C.; Feng, Y.; Gaje, G.; Guzman, E.; Heckman, T.; Mellis, A.; Moore, L.; Romo Bechara, N.; Sanchez, S.; et al. The Weekend Effect on Urban Bat Activity Suggests Fine Scale Human-Induced Bat Movements. Animals 2020, 10, 1636. [Google Scholar] [CrossRef]
- Loeb, S.C.; Post, C.J.; Hall, S.T. Relationship between urbanization and bat community structure in national parks of the southeastern U.S. Urban Ecosyst. 2009, 12, 197–214. [Google Scholar] [CrossRef]
- Schimpp, S.A.; Li, H.; Kalcounis-Rueppell, M.C. Determining species specific nightly bat activity in sites with varying urban intensity. Urban Ecosyst. 2018, 21, 541–550. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; ISBN 3-900051-07-0. Available online: https://www.r-project.org/ (accessed on 10 September 2021).
- Thieurmel, B.; Elmarhraoui, A. Suncalc: Compute Sun Position, Sunlight Phases, Moon Position and Lunar Phase. 2019. Available online: https://cran.r-project.org/web/packages/suncalc/index.html (accessed on 10 September 2021).
- Reichert, B.E.; Bayless, M.; Cheng, T.L.; Coleman, J.T.H.; Francis, C.M.; Frick, W.F.; Gotthold, B.S.; Irvine, K.M.; Lausen, C.; Li, H.; et al. NABat: A top-down, bottom-up solution to collaborative continental-scale monitoring. Ambio 2021, 50, 901–913. [Google Scholar] [CrossRef]
- Loeb, S.C.; Hines, B.A.; Armstrong, M.P.; Zarnoch, S.J. Effects of Omnidirectional Microphone Placement and Survey Period on Bat Echolocation Call Quality and Detection Probabilities. Acta Chiropterol. 2020, 21, 453–464. [Google Scholar] [CrossRef]
- Findlay, S.V.; Barclay, R.M.R. Acoustic Surveys for Bats are Improved by Taking Habitat Type into Account. Wildl. Soc. Bull. 2020, 44, 86–93. [Google Scholar] [CrossRef]
- Quinn, G.P.; Keough, M.J. Experimental Design and Data Analysis for Biologists; Cambridge University Press U.S.: New York, NY, USA, 2002; ISBN 0-521-00976-6. [Google Scholar]
- Wickham, H.; Chang, W. Ggplot2: An Implementation of the Grammar of Graphics. R Package Version 0.7. Available online: http://CRAN.+R-project.+org/package=+ggplot2+2008 (accessed on 10 September 2021).
- Parker, K.A., Jr.; Li, H.; Kalcounis-Rueppell, M.C. Species-Specific Environmental Conditions for Winter Bat Acoustic Activity in North Carolina, United States. J. Mammal. 2020, 101, 1502–1512. [Google Scholar] [CrossRef]
- McCune, B.; Grace, J.B.; Urban, D.L. Analysis of Ecological Communities; MJM Software Design: Gleneden Beach, OR, USA, 2002; ISBN 978-0-9721290-0-8. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology (R Package). 2018. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 10 September 2021).
- Vindigni, M.A.; Morris, A.D.; Miller, D.A.; Kalcounis-Rueppell, M.C. Use of modified water sources by bats in a managed pine landscape. For. Ecol. Manag. 2009, 258, 2056–2061. [Google Scholar] [CrossRef]
- Kalcounis-Rueppell, M.C.; Payne, V.H.; Huff, S.R.; Boyko, A.L. Effects of wastewater treatment plant effluent on bat foraging ecology in an urban stream system. Biol. Conserv. 2007, 138, 120–130. [Google Scholar] [CrossRef]
- Li, H.; Parker, K.A.; Kalcounis-Rueppell, M.C. The luxury effect beyond cities: Bats respond to socioeconomic variation across landscapes. BMC Ecol. 2019, 19, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Wilkins, K.T. Patch or mosaic: Bat activity responds to fine-scale urban heterogeneity in a medium-sized city in the United States. Urban Ecosyst. 2014, 17, 1013–1031. [Google Scholar] [CrossRef]
- Li, H.; Wilkins, K.T. Selection of Building Roosts by Mexican Free-Tailed Bats (Tadarida brasiliensis) in an Urban Area. Acta Chiropterol. 2015, 17, 321–330. [Google Scholar] [CrossRef]
- Lewanzik, D.; Sundaramurthy, A.K.; Goerlitz, H.R.; Derryberry, E. Insectivorous bats integrate social information about species identity, conspecific activity and prey abundance to estimate cost–benefit ratio of interactions. J. Anim. Ecol. 2019, 88, 1462–1473. [Google Scholar] [CrossRef]
- Culina, A.; Garroway, C.J. Bats use social information within and across species. J. Anim. Ecol. 2019, 88, 1444–1446. [Google Scholar] [CrossRef] [PubMed]
- Neubaum, D.J.; O’Shea, T.J.; Wilson, K.R. Autumn Migration and Selection of Rock Crevices as Hibernacula by Big Brown Bats in Colorado. J. Mammal. 2006, 87, 470–479. [Google Scholar] [CrossRef]
- Duchamp, J.E.; Sparks, D.W.; Whitaker, J.O. Foraging-habitat selection by bats at an urban–rural interface: Comparison between a successful and a less successful species. Can. J. Zool. 2004, 82, 1157–1164. [Google Scholar] [CrossRef] [Green Version]
- Lehrer, E.W.; Gallo, T.; Fidino, M.; Kilgour, R.J.; Wolff, P.J.; Magle, S.B. Urban bat occupancy is highly influenced by noise and the location of water: Considerations for nature-based urban planning. Landsc. Urban Plan. 2021, 210, 104063. [Google Scholar] [CrossRef]
- Moretto, L.; Francis, C.M. What factors limit bat abundance and diversity in temperate, North American urban environments? J. Urban Ecol. 2017, 3, jux016. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| UNCG Open Wetland | UNCG Open Control | UNCG Woody Wetland | UNCG Woody Control | GSC Rooftop Level | GSC Canopy Level | GSC Ground Level | |
|---|---|---|---|---|---|---|---|
| 2017 Spring | 22,481 | 23,379 | 8824 | 3559 | N/A | 1394 | 262 |
| 2017 Summer | 36,428 | 36,284 | 5565 | 1131 | 8792 | 7725 | 1148 |
| 2017 Fall | 7917 | 5938 | 2152 | 2054 | 5514 | 2514 | 1635 |
| 2017 Winter | 4794 | 2445 | 1526 | 1561 | 309 | 281 | 280 |
| 2018 Spring | 19,952 | 19,578 | 9588 | 3964 | 4765 | 1909 | 497 |
| 2018 Summer | 35,358 | 19,219 | 5879 | 398 | 11,350 | 4972 | 1482 |
| 2018 Fall | 2621 | 8898 | 1481 | 1898 | 7554 | 1104 | 909 |
| 2018 Winter | 2728 | 1460 | 844 | 546 | 131 | 205 | 115 |
| 2019 Spring | 18,481 | 33,980 | 10,098 | 5217 | 2783 | 852 | 824 |
| 2019 Summer | 32,177 | 30,544 | 3906 | 873 | 13,172 | 4789 | 738 |
| 2019 Fall | 3662 | 5744 | 558 | 1436 | 11,869 | 1045 | 1183 |
| 2019 Winter | 2154 | 1806 | 812 | 887 | 225 | 153 | 117 |
| 2020 Spring | 20,712 | 28,819 | 12,160 | 10,979 | 1235 | 1177 | 498 |
| 2020 Summer | 35,598 | 26,851 | 2512 | 1227 | 10,072 | 8586 | 2274 |
| 2020 Fall | 6775 | 3020 | 1858 | 435 | 9066 | 3122 | 756 |
| 2020 Winter | 417 | 271 | 81 | 83 | 242 | 19 | 59 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.; Petric, R.; Alazzawi, Z.; Kauzlarich, J.; Mahmoud, R.H.; McFadden, R.; Perslow, N.; Rodriguez Flores, A.; Soufi, H.; Morales, K.; et al. Four Years Continuous Monitoring Reveals Different Effects of Urban Constructed Wetlands on Bats. Land 2021, 10, 1087. https://doi.org/10.3390/land10101087
Li H, Petric R, Alazzawi Z, Kauzlarich J, Mahmoud RH, McFadden R, Perslow N, Rodriguez Flores A, Soufi H, Morales K, et al. Four Years Continuous Monitoring Reveals Different Effects of Urban Constructed Wetlands on Bats. Land. 2021; 10(10):1087. https://doi.org/10.3390/land10101087
Chicago/Turabian StyleLi, Han, Radmila Petric, Zinah Alazzawi, Jake Kauzlarich, Rania H. Mahmoud, Rasheed McFadden, Niklas Perslow, Andrea Rodriguez Flores, Hadi Soufi, Kristina Morales, and et al. 2021. "Four Years Continuous Monitoring Reveals Different Effects of Urban Constructed Wetlands on Bats" Land 10, no. 10: 1087. https://doi.org/10.3390/land10101087