
Macrozoobenthos Structure and Dynamics in a Mediterranean Hypersaline Ecosystem with Implications for Wetland Conservation

1
Dipartimento di Biologia, Università di Roma “Tor Vergata”, 00133 Rome, Italy
2
Dipartimento di Biologia e Biotecnologie “Charles Darwin”, Università di Roma “La Sapienza”, 00185 Rome, Italy
3
CoNISMa (Consorzio Nazionale Interuniversitario per le Scienze del Mare), 00196 Rome, Italy
4
Dipartimento di Biologia Ambientale, Università di Roma “La Sapienza”, 00185 Rome, Italy
*
Author to whom correspondence should be addressed.
Water 2023, 15(7), 1411; https://doi.org/10.3390/w15071411
Submission received: 8 March 2023
/
Revised: 20 March 2023
/
Accepted: 3 April 2023
/
Published: 5 April 2023
(This article belongs to the Special Issue Research on the Aquatic Species Biodiversity and Morphology)
Abstract
:Within the framework of hypersaline environments, the Saline di Tarquinia salterns (Latium, Italy) represent an interesting case study from ecological, naturalistic, and conservationist perspectives. We studied macrozoobenthic community zonation related to the main environmental parameters, with particular emphasis on the beetle species Nebrioporus ceresyi (Aubé, 1836), one of the few Dytiscidae among Italian fauna particularly adapted to brackish and hypersaline waters. In this study, we report the structure and seasonal dynamics of the macrozoobenthic community, the spatial and temporal distribution of both larvae and adults of N. ceresyi, and the size–frequency distribution of the larvae. The results add new information on the macrozoobenthic assemblage of the Saline di Tarquinia salterns and the life cycle of this water beetle in the hypersaline habitat. A possible hypothesis about the discontinuous geographical distribution of the species of Dytiscidae investigated is also provided. Finally, N. ceresyi is proposed as an indicator species useful in characterizing the environmental zonation of hypersaline habitats.
1. Introduction
Hypersaline ecosystems are patchily distributed around the world and, despite the similar extreme conditions of their environments, they are heterogeneous in terms of origin, chemical, and biological characteristics [1]. In terms of their origin, natural hypersaline environments include marine environments such as solar salt ponds, sabkhas, and non-marine salt lakes. In addition, humans have created artificial hypersaline environments constructed to produce salt, i.e., solar salterns and saltworks. Along the Mediterranean coast, hypersaline habitats are widely distributed and include artificial solar salterns (saltworks), salt marshes, natural sabkhas (southern Mediterranean coast), and salt lakes such as the Jordan Dead Sea [2]. Notwithstanding their different origins, the ecological processes and dynamics of both natural and artificial hypersaline systems have a considerable degree of commonality, with salterns and sabkhas constituting the hypersaline pole of the variability gradient of transitional aquatic ecosystems. Solar salterns and hypersaline environments have received little attention in ecological studies on transitional waters compared to brackish and lagoon systems [3,4,5]. Salterns have productive, economic, and social importance equivalent to these other ecosystems, and therefore deserve similar attention. In contrast to the focus on biological components such as bacteria [6], algae [7], and plankton [8] that dominate hypersaline environments, there is still little knowledge about macrobenthic invertebrates, in particular entomofauna, in hypersaline waters. In particular, beetle fauna characterizes the biotopes of lentic and lotic freshwater in the circum-Mediterranean area. However, only a few species of two families are particularly adapted to brackish and hypersaline waters: the Hydraenidae (suborder Polyphaga) of the genus Ochthebius, and the Dytiscidae (suborder Adephaga) of the genus Nebrioporus [9]. The latter includes Nebrioporus ceresyi (Aubé, 1836), a species of Italian fauna that lives in lentic waters at sea level environments [10].
Within the framework of hypersaline environments, the Saline di Tarquinia salterns (Latium, Italy) represent an interesting case study from ecological, naturalistic, and conservationist perspectives. We have recently found numerous specimens of N. ceresyi, both in the adult and in the larval stages, in this location. This finding is of considerable interest from various points of view: faunal because it is the first report in the Saline di Tarquinia salterns and a confirmation for Latium fauna; biological, for the contribution to knowledge about the life cycle of this species; and conservationist, for possible useful indications for management purposes and for assessing the quality of the habitat.
The present work aims to: (i) describe the macrozoobenthic community along the salinity gradient in this system, (ii) report the first record of N. ceresyi in the hypersaline environments of Latium and infer the possible mechanisms of dispersion, (iii) reconstruct the dynamics of the biological cycle of the species, and (iv) propose useful indications for monitoring activities in the management of protected areas.
2. Materials and Methods
2.1. Study Area and Sampling Sites
The Saline di Tarquinia salterns extend over 193 ha along the Northern Latium coast and consist of about a hundred interconnected ponds several tens of centimetres deep [11]. Seawater enters through the western mouth into the northernmost pond, where it is artificially pumped and circulates through the sequence of ponds from the northern to the southern ones. This results in a marked salinity gradient in the ponds, ranging from sea salinity values (37–38%) to progressively increasing hypersalinity values exceeding 200%. The very shallow depth of the ponds causes the water mass to be strongly affected by weather phenomena and, therefore, extremely variable in physical and chemical parameters. Although the salterns are artificial habitats, due to their structures and dynamics, they consistently reflect the ecological patterns of transitional-water ecosystems, namely, drastic variability in environmental conditions, restricted area and marked separation from the sea, strong selection of fauna and flora, and a low number of invertebrate species in the biotic community, which in turn supports a high abundance of individuals due to the high availability of trophic resources. These constitute abundant food resources for an equally rich range of vertebrates, particularly fish and birds. Moreover, the salterns are a suitable habitat for rare halophile flora and fauna adapted to extreme conditions. For these reasons, the salterns have a special ecological value and have been recognized as wetlands of international importance under the Convention of Ramsar (1971). They have also been declared a Nature Reserve of Animal Repopulation under the decree of 25 January 1980 of the Ministry of Agricultural and Forests, a Site of Community Importance in the European Commission Habitats Directive (92/43/EEC), and a Special Protection Zone under the terms of the Birds Directive (2009/147/EC) and are included in the Natura 2000 Network. At the national level, the Saline di Tarquinia salterns are included in the network of 130 Nature State Reserves managed by the Nucleo Carabinieri Biodiversità delle Saline di Tarquinia attached to the Raggruppamento Carabinieri Biodiversità of Rome (Italy).
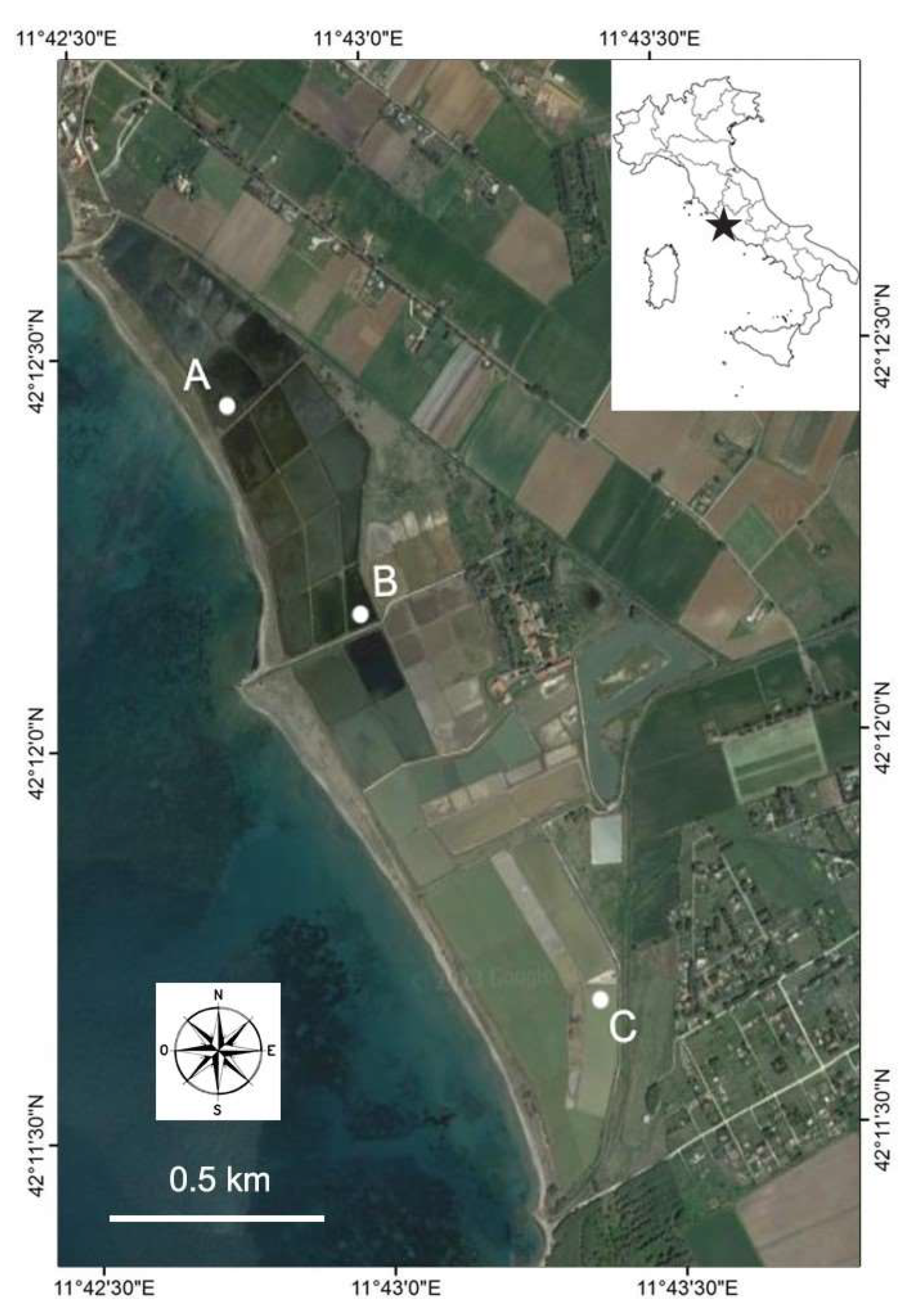
For this study, three sites (A, B and C) were selected in different ponds along the salinity gradient (Figure 1). Site A was located in the northern sector of the salterns, in a pond directly connected to the sea via an artificial canal, site B was located in the intermediate sector of the saltern system, and site C was located in a crystallization pond, where the depth is greatest and sodium chloride precipitation occurs.
2.2. Sampling and Laboratory Analysis
Samplings were conducted monthly in the following periods: September 2015 to May 2016, June 2017 to November 2017, and April 2021 to July 2021. Dissolved oxygen (DO; mg/L) and temperature (T; °C) were measured using a multiparameter probe (YK-22DO, Lutron, Coopersburg, PA, USA). The pH was measured using a pH meter (Testr PC5, XS Instrument, Hyderabad, India). Salinity (psu) was measured using a refractometer. Macrobenthos faunal samples were collected using the leaf bag technique [12]. Each leaf bag was prepared with 3 g of Phragmites australis dried leaves. The bags were placed in three sampling stations located in three ponds with progressively increasing salinity from seawater to hypersaline values (three replicates in each station). The bags were retrieved after thirty days of submersion, placed in plastic bags, and brought into the laboratory, where the leaves were gently washed, and the invertebrate fauna was sorted. Beetle specimens were collected for identification and kept in the authors’ collection.
2.3. Data Analysis
A two-way Permutational Analysis of Variance (PERMANOVA) based on Bray-Curtis (dis)similarity measures [13] was used to test significant differences between sites and seasons based on abiotic parameters.
As descriptors of the macrozoobenthic community structure, synthetic biotic parameters, i.e., species richness, abundance (number of individuals), diversity (Shannon index, based on the natural logarithm), and evenness (Pielou index) were computed by grouping the monthly samplings for each season (spring: April, May and June; summer: July, August and September; autumn: October, November and December; winter: January, February and March). The abundance data relative to macrozoobenthic assemblages were log-transformed and used as an input matrix for the non-metric multidimensional scaling (nMDS) ordination technique. A Bray-Curtis index was used for the (dis)similarity measure. The top-contributing taxa in discrimination between and within sites were identified by employing the Similarity Percentage (SIMPER) test [14]. Analysis of similarities (ANOSIM) was performed to evaluate significant differences between sites based on macrozoobenthic community structures.
3. Results
3.1. Environmental Variables
Mean values of the measured environmental variables varied between periods and sites. Water salinity varied from 34.3 psu (February, site A) to 123.9 psu (May, site C), DO from 6.4 mg/L (July, site B) to 13.2 mg/L (December, site B), temperature from 9 °C (December, site A) to 36.1 °C (August, site C), and pH from 7.7 (January and May, site C) to 8.7 (October, site A). Salinity increased along the corresponding gradient from site A to site C, the hypersaline site. Salinity was found to be more stable in sites A and B, while fluctuating more in site C. Temperature showed a similar trend, consistently rising from site A to site C and following expected seasonal fluctuations. DO and pH values exhibited significant within- and between-site variation and were generally lower at site C than at sites A and B (Figure 2).
The two-way PERMANOVA carried out on each environmental parameter indicated highly significant differences between sites for salinity, DO and pH. Salinity and temperature also showed significant differences between periods (Table 1).
3.2. Macrozoobenthic Community
A total of 27,642 individuals belonging to 18 taxa were collected (Table S1; Figure S1). Most species were crustaceans (38.9%), followed by molluscs (16.7%), insects (both larvae and adults; 16.7%), polychaetes (16.7%), nemerteans (5.5%) and bryozoans (5.5%).
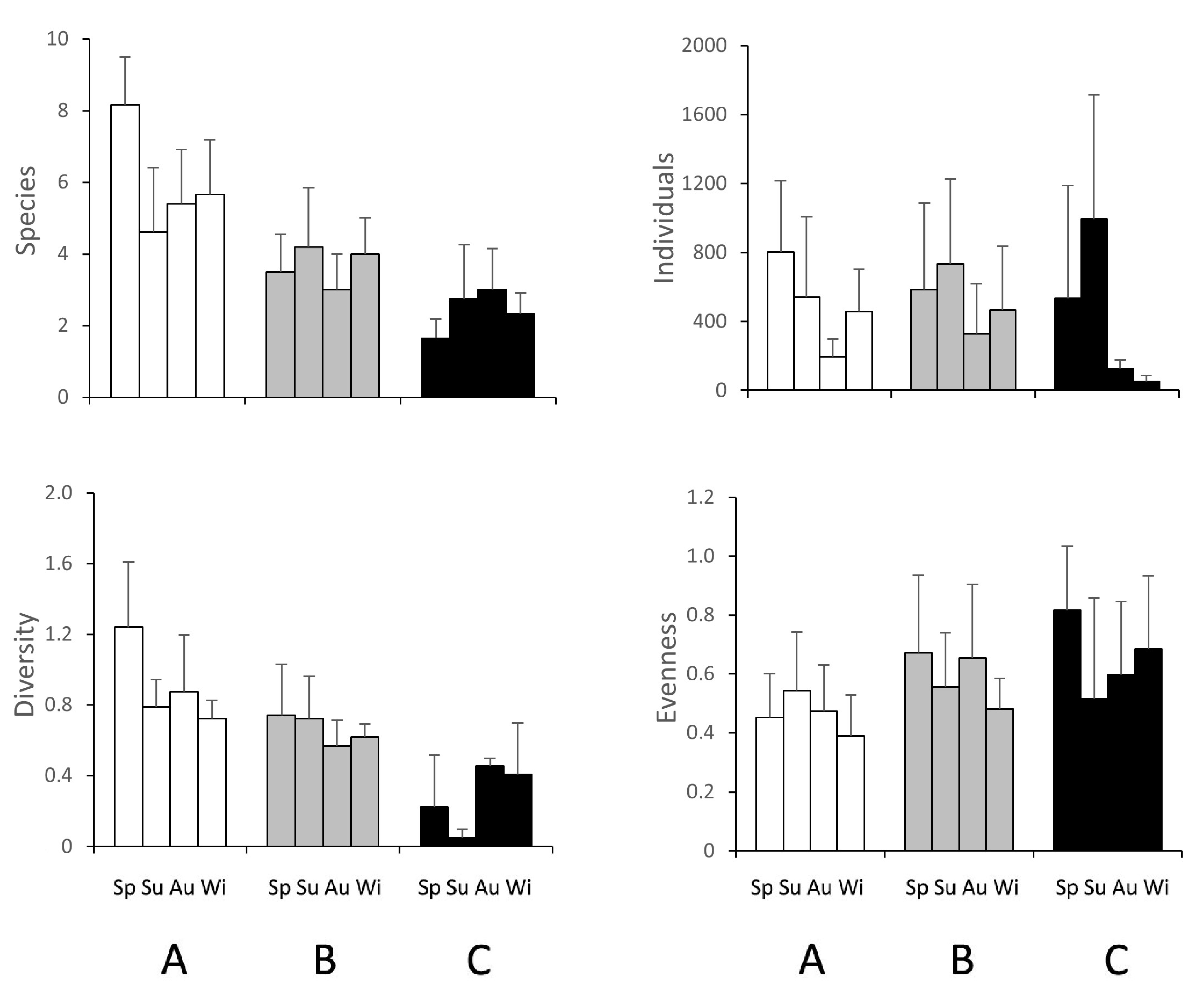
Variation in macrofaunal abundance is shown in Figure 3. The number of taxa decreased along the salinity gradient from site A to site C. Site C showed the highest abundance and the lowest diversity in summer, when the highest occurrence of chironomids larvae was found. Conversely, the euryhaline site A, which was closest to the sea, showed the highest diversity and the lowest evenness. Intermediate values were found in site B (Figure 3).
The ANOSIM test indicated that the three sites were significantly different in terms of their benthic macrofaunal composition (global R = 0.5; mean rank within = 492.6; mean rank between = 861.9; p = 0.0001).
The nMDS plot of the structure of the assemblage highlighted three major groups corresponding to the three sites (stress = 0.12), with the hypersaline site C being the most separated and sites A and B showing a partial overlap. The graph shows that the environmental factors mainly responsible for the separation between groups were salinity and pH, while temperature contributed most to the arrangement within groups (Figure 4).
The SIMPER analysis showed the taxa with the highest contribution to the (dis)similarity within and between sites. The top discriminating taxa were Gammarus aequicauda, Chironomidae, Hydrobia acuta and Cerastoderma glaucum for sites A and B; H. acuta, G. aequicauda, Chironomidae and N. ceresyi for sites A and C; and H. acuta, Chironomidae, N. ceresyi and C. glaucum for sites B and C (Table 2).
3.3. Nebrioporus Ceresyi Population
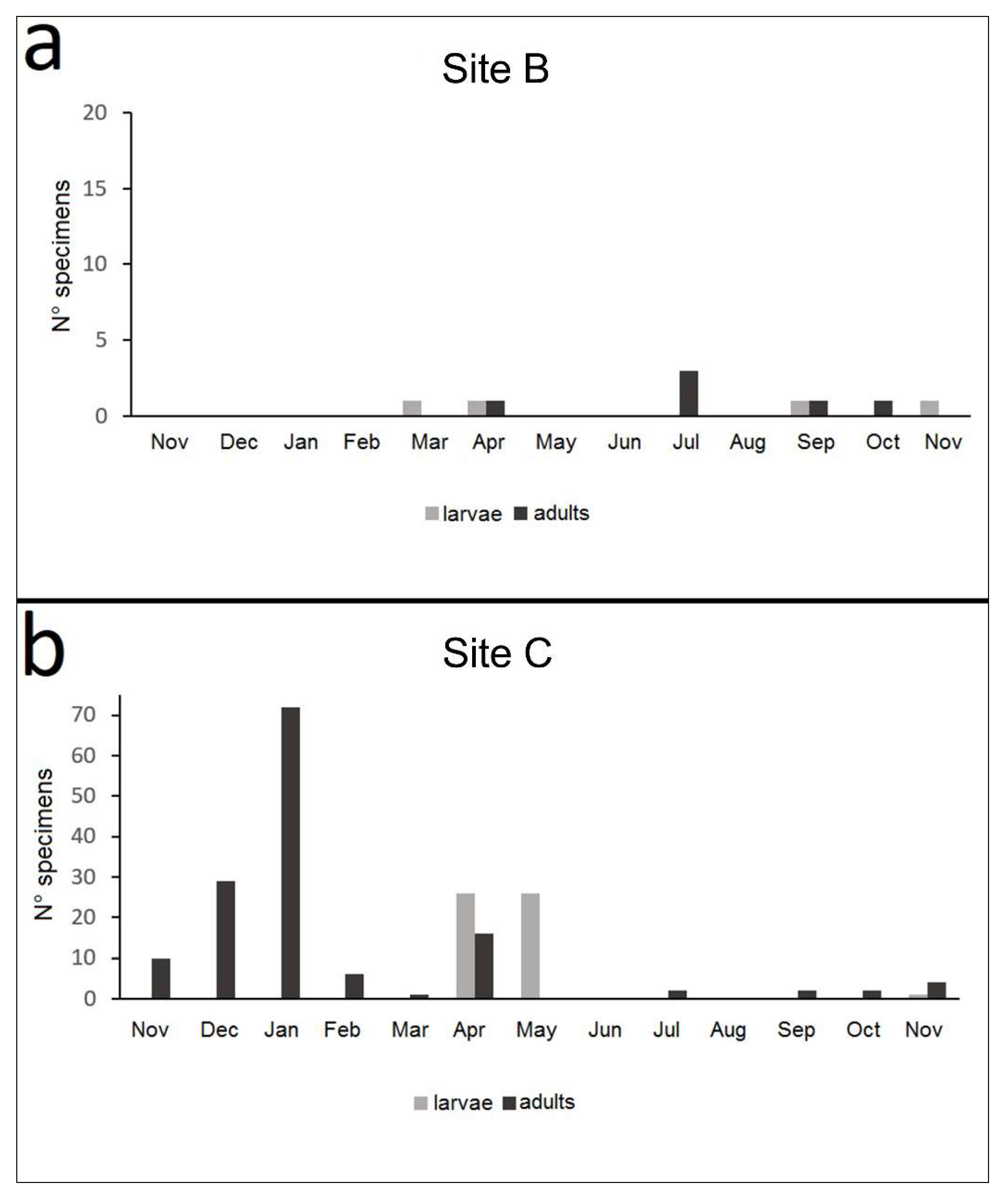
A total of 202 specimens of N. ceresyi were collected, comprising 145 adults and 57 larvae, all found in sites B and C (Figure 5). All the individuals were recorded in the period from September to July, showing a clear alternation between adult individuals and larvae, partially overlapping only in September and November. The specimens were identified according to the diagnostic characters available in the taxonomic literature, with particular reference to Franciscolo [9] and Toledo [10].
3.3.1. Brief Description of Larvae
The body is subcylindrical with a length of 1.5–8.0 mm, with a yellow to light brown colour pattern. As in other Dytiscidae species, the larvae are campodeiform with prognathous mouthparts and distinctly sclerotized head capsule. The cephalic capsule is longer than it is wide and has the typical “pear” shape with evident lateral sinuosity and occipital suture. The thorax typically consists of three segments, pro-, meso- and meta-thorax, with a sclerotized dorsal surface, each bearing slender articulated yellow legs equipped with swimming setae. The abdomen consists of eight segments with terga bordered with brown lateral margins and two round dark spots on each side, more marked in the first segments and less marked in the last ones, the eighth segment lacking spiniform protuberance, and having cerci with setae throughout their length (Figure 5a).
3.3.2. Brief Description of Adults
The body is elongated with a length of 4.0–5.0 mm and dorsally convex, a general yellowish-orange colour, and an elytra pattern with five parallel longitudinal dark striae including an outermost stria with variable thickness and an irregular edge (Figure 5b). The base of the pronotum is as wide as the base of the elytra with convergent lateral margins and posterior angles that are acute or sub-rectus and not rounded at the apex (Figure 5c). The elytra lack carinae or elevations but are covered with fine and dense punctuation (Figure 5d); subapical denticles and angularities are absent on the elytral apex (Figure 5f). The protarsi and mesotarsi are pseudotetrameric, with the fourth tarsomere being minute and included in the marginalization of the third (Figure 5e). The surface of the sternites is covered with dense and fine punctuation having the appearance of emery paper, particularly on the meta-sternites and meta-coxae.
3.3.3. Life Cycle
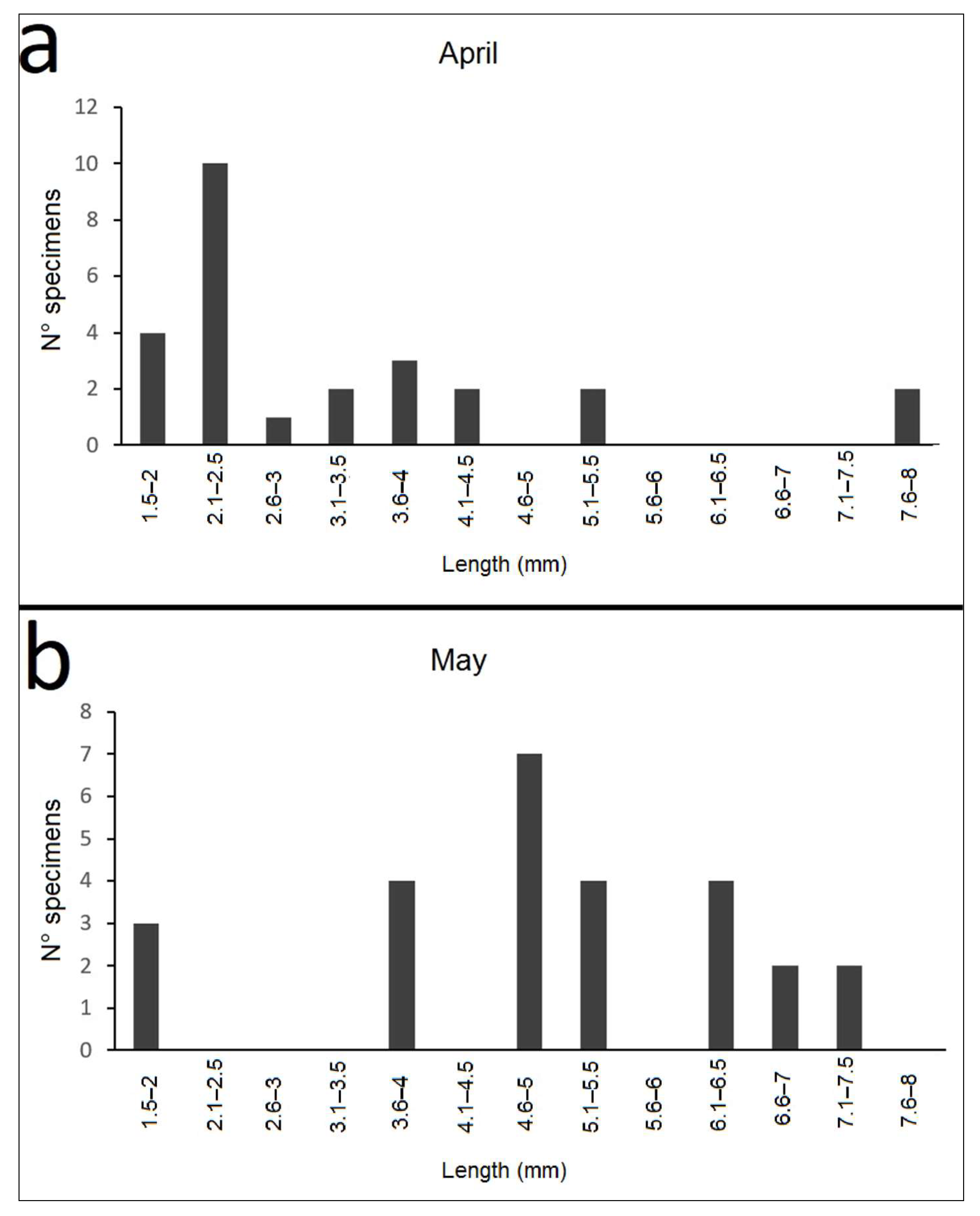
Adults were mainly recorded in autumn and throughout the winter until April; no adults occurred in May, June, or August, and only a few specimens were found in July. Larvae were first recorded on March 31; the highest number of larval specimens occurred in April and May. Only one and two larvae were found in September and November, respectively (Figure 6). The larval life span was just over one month, during which the larvae reached lengths varying from 1.8 to 8.0 mm (Figure 7).
4. Discussion
4.1. Macrozoobenthic Community Structure and Dynamics
The present study proved that the Saline di Tarquinia salterns constitute a typical example of the hypersaline environment associated with solar saltern systems. The investigated sites were found to be significantly differentiated based on environmental parameters, as the PERMANOVA highlighted, and were distributed along the typical salinity gradient extending from seawater to very hypersaline values in sites A and C, respectively. Accordingly, the lowest values of DO and pH recorded at the latter site can be explained by the strict dependence of these parameters on salinity [1]. The temperature was a minor factor in discriminating between sampling sites since a similar seasonal trend was found in every site. This is consistent with the results obtained in ecological studies of macroinvertebrate communities of lagoonal environments [5,15]. In this study, the slightly higher temperatures at site C are attributable to this station’s very low water depth.
Salinity strongly affected the macrozoobenthic community, as the synthetic biotic parameters demonstrated. The number of species decreased according to the increase in salinity, as did diversity, highlighting that the latter parameter was mainly affected by species richness rather than by the even distribution of individuals. Conversely, the highest values of evenness in site C were due to the dominance of very few taxa in this site, pointing out the unbalance between the scarcity of taxa and the high abundance of individuals in the hypersaline site.
The multivariate analysis confirmed these results, emphasizing major differences between sites based on the macrozoobenthic community structure. The habitat heterogeneity between sites was consistent with the salinity gradient, which proved to be the main constraining factor in the selection of benthic species and appeared to be persistent through time. A similar role for the salinity gradient is known for other transitional water ecosystems [16,17,18]. Temperature was mainly responsible for the heterogeneity within each site, according to seasonal variations, which were similar in the various sites. Salinity distinguished a marine pole at site A, where the influence of marine waters enhanced a more rich and more diverse assemblage. Here the most discriminating species (according to SIMPER analysis) was G. aequicauda, an amphipod commonly inhabiting sheltered coastal areas; other marine species, e.g., C. vulgatum, Monocorophium sextonae, Neodexiospira pseudocorrugata, as well as some typical components of brackish water zoocoenosis, e.g., Idotea chelipes and Monocorophium insidiosum [5], also characterized this site. The central sector of the saltern, site B, appeared suitable only for a smaller number of species, such as the halolimnobic molluscs C. glaucum and H. acuta [5] and the opportunistic chironomid larvae, which most distinguished this site from the others. In contrast to site A, in site C, which was most separated from the outlet to the sea, the environmental conditions allowed very few species to survive the extremely high salinity values. Together with C. glaucum, H. acuta and chironomids, the water beetle N. ceresyi most characterized this site, and was found for the first time to be a typical element of the macrozoobenthic community in hypersaline ecosystems. For these reasons, this species deserves special mention.
4.2. Nebrioporus Ceresyi Life Cycle and Distribution
The discovery of a large population of N. ceresyi represents the first record for the Saline di Tarquinia salterns and new data for the brackish environments of Lazio. This species was not reported among the entomofauna of the Saline di Tarquinia salterns by Zapparoli et al. [19]. Its presence in Pratica di Mare, in the province of Rome [20], requires validation, given the intense urbanization that this area has undergone in recent decades, which has led to the almost total disappearance of wetlands (Gravina, personal communication). The same is true for the dubious report of its presence in the Castel Porziano estate [21]. Furthermore, the discovery of numerous specimens of N. ceresyi in the adult and larval stages is of particular interest both for reconstructing its biological cycle and for plausible interpretations regarding its geographical distribution.
The biological cycle of N. ceresyi, articulated as described above, is only partially in agreement with what was reported for other species of Dytiscidae by Franciscolo [9], who identifies the autumn as the season of greatest abundance of adults and April and May as the period of development of larvae. Indeed, in the Saline di Tarquinia salterns, in addition to the typical autumn-winter generation with reproduction in spring, N. ceresyi has shown, albeit for only a small number of adults, the ability to survive the summer season and also to generate larvae in autumn. Its biological cycle extended throughout the year, with reproductive periods at the beginning of spring and at the end of summer, highlighting this species’ particular adaptation to the environment’s hypersalinity conditions.
Unlike the larvae, which are completely adapted to the aquatic environment thanks to their cutaneous respiration and the presence of swimming bristles on the limbs, the pupae develop in small cells arranged on the ground. These are usually found along the banks of the basin at a distance from the water surface such that they can be traversed, in one direction, by larvae that are late to reach land and, in the other direction, by newly emerged adults that must re-enter the water quickly [9]. The pupal stage, which is little known for these beetles, has been carefully described based on specimens including some adults and a larva collected in the Bagno dell’Acqua Lake, an endorheic saline lake within a calderic depression on Pantelleria [22].
N. ceresyi is a holo-Mediterranean species with a wide geographical distribution, having been observed in numerous countries, such as Algeria, Bulgaria, the Canary Islands, Corsica, Crete, Crimea, Croatia, Cyprus, Egypt, southern France, Greece, Israel, Iran, Iraq, Italy, Libya, Malta, Morocco, Portugal, Spain, Syria, Tunisia, Turkey and Turkmenistan [10,23,24].
However, as for most other aquatic beetles, the local distribution of N. ceresyi is very discontinuous, being linked to very specific ecological conditions. Along the Italian coast, the species has been reported in various brackish environments, including: Liguria, Oneglia [25,26]; Marche, Civitanova Marche and Cupra Marittima [20]; Tuscany, Orbetello Lagoon [9]; Molise, Portocannone [20]; Apulia, in brackish environments in the localities of Torre Colimena, and a residual pond pool behind dunes with a clayey bottom [27]; Porto Cesareo, in a small pond along the rocky coast of the Ionian Sea [27]; Alimini lakes on the Adriatic coast [9]; Basilicata, Bosco di Policoro, in a stretch of a canal with earth banks and in the temporary Pantano stream [27]; Campania, Naples [25,26]; Calabria, in a clay pool in Sibari [28]; Sardinia, specimens from Pula, Cagliari [26] preserved in the Museum of Verona, from the Quartu Sant’Elena-Stagno di Molentargius pond [9,20]; Casaraccio pond [9,29]; Saline Tonnara-Stintino, in brackish pools behind the dunes [9]; Sassari, behind the dunes of Platamona, in a freshwater marsh with Phragmites australis cover [9]; Sicily, Castelvetrano, Termini Imerese, Lago Biviere di Lentini [25,26,30,31,32]; in ponds of Passo Martino in Catania [9] (currently Passo Martino is a Catania city district and no reference to existing ponds was found); Aeolian Islands, Salina di Punta Lingua [33]; and Pantelleria, Lago Bagno dell’Acqua [22,26,30,31,33] (Figure S2). Of these, reports relating to the first half of the last century and no longer reconfirmed by more recent collections need confirmation regarding the current presence of N. ceresyi, such as, for example, the reports in Liguria, Naples and Pula, where the landscape has undergone drastic rearrangement. Franciscolo [9] expresses his doubts about the conservation of aquatic beetle fauna in environments profoundly modified by anthropogenic pressure, referring in particular to the striking case of the Biviere di Lentini, which was completely drained in the 1930s and then reconstituted in the 1970s. Similarly, the presence of N. ceresyi in other Sicilian records would require confirmation, such as in the cases of Castelvetrano, Termini Imerese and Passo Martino, where the habitat has been substantially modified by human intervention. A particularly marked transformation of anthropic origin occurred in the wetland area of Policoro in Basilicata, where the presence of this species is considered unlikely. In contrast, in Bagno dell’Acqua in Pantelleria, the presence of N. ceresyi has been reconfirmed by recent research [22]. In other Sardinian and Apulian localities and in the pond of Punta Lingua in Salina (Aeolian Islands), the presence of the species can be considered likely, due to the good conservation status of the habitat in these localities.
Regarding the scattered geographical distribution of N. ceresyi, it is worth pointing out that the members of the family Dytiscidae, although capable of flying, are capable of only limited movement. For example, for stream-inhabiting species, Franciscolo [9] explains the extremely localized distribution of the species by the fact that these beetles scarcely move along the course of the same stream or from one body of water to another. Indeed, N. ceresyi, although recorded in the Orbetello lagoon along the Tyrrhenian coast and in some brackish ponds on the northern coast of Sardinia, was not found on the island of Elba, which is located at an intermediate position [34]. In light of these considerations, the scattered and localized distribution of N. ceresyi could be explained by a mechanism of passive dispersion, likely due to the transport of some specimens embedded in mud clods trapped between the toes of migratory birds. This hypothesis could be confirmed by the fact that many brackish and hypersaline biotopes where this species has been reported are found along the migration routes followed by various bird species, such as flamingos [35]. Further evidence supporting this hypothesis comes from the presence of N. ceresyi on the island of Pantelleria, located in the Strait of Sicily halfway between Sicily and Tunisia.
4.3. Implications for Conservation
During the monitoring activities carried out in the Saline di Tarquinia salterns [36,37,38], N. ceresyi was found to be a species closely associated with stations characterized by particularly high salinity values, where it completes its life cycle. In such environments, this species appeared to play a key role in the trophic chain, which is extremely short due to the very low number of species living in such extreme conditions. Furthermore, considering its highly localized geographical distribution, the presence of N. ceresyi designates the Saline di Tarquinia salterns as a significant site within the Natura 2000 network, in accordance with Directive 92/43/EEC.
In addition, its presence in the hypersaline ponds testifies to the efficiency of the hydraulic regime management, which guarantees the good state of conservation of the habitat and is optimal for the survival of the very few species adapted to it. Among these, the Saline di Tarquinia salterns host the Mediterranean banded killifish, Aphanius fasciatus, a euryhaline bony fish belonging to the Cyprinodontidae family, which is listed in Directive 92/43/EEC among the “animal and plant species of community interest whose conservation requires the designation of special areas of conservation” (Annex II). Furthermore, in the Red List of indigenous freshwater fish in Italy, the Mediterranean banded killifish is considered “vulnerable” because of its discontinuous presence among the suitable habitats and its significant decline in some sites [39].
Therefore, it is reasonable to conclude that N. cereysi can be considered an “indicator species” of the hypersaline pole of the “coastal lagoons” habitat in the Saline di Tarquinia salterns. Its presence and abundance could be useful in characterizing the environmental zonation of protected areas that include hypersaline habitats, such as the “Riserva Naturale Statale” Saline di Tarquinia, an essential site for the preservation and conservation of both avifauna and protected fish species like the Mediterranean banded killifish.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/w15071411/s1, Table S1: Presence of the recorded taxa at sites A, B and C in spring (Sp), summer (Su), autumn (Au) and winter (Wi). Figure S1: Pie graphs showing the relative abundances of the macrozoobenthic taxa in each sampling site. Figure S2: Location of the historical records for the presence of N. ceresyi in Italy.
Author Contributions
Conceptualization, A.B., S.G., M.F.G. and D.V.; data curation, A.B., S.G. and M.F.G.; methodology, M.F.G. and D.V.; investigation, A.B., S.G., M.F.G. and D.V.; validation, A.B., S.G., M.F.G. and D.V.; visualization, A.B., S.G., M.F.G. and D.V.; writing—original draft preparation, A.B. and M.F.G.; writing—review and editing, A.B., S.G., M.F.G., D.V.; project administration, M.F.G. All authors have read and agreed to the published version of the manuscript.
Funding
We are in debt to the Carabinieri Biodiversity Group—Biodiversity Department of Rome for financing the monitoring activities of the Riserva Naturale Statale “Saline di Tarquinia” and supporting this study.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
The authors wish to thank in particular the Carabinieri Biodiversity Nucleus “Saline di Tarquinia”, in the persons of the Brigadiere Capo Valentina Sabbatini and Appuntati Scelti Valerio Finori, Alfredo Massi, Decimo Rosi and Alfredo Cea, for their availability, spirit of collaboration and consistently demonstrated professionalism, which made the field and study activities envisaged for monitoring easier.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Javor, B. Hypersaline Environments: Microbiology and Biogeochemistry; Brock/Springer Series in Contemporary Bioscience; Springer: Berlin/Heidelberg, Germany, 1989; ISBN 978-3-642-74372-6. [Google Scholar]
- Perthuisot, J.-P.; Guelorget, O.; Groupe d’étude du Domaine Paralique. Le Domaine Paralique: Expressions Géologiques, Biologiques et Économiques du Confinement; Presses de l’École Normale Supérieure: Paris, France, 1983; ISBN 978-2-7288-0094-0. [Google Scholar]
- Basset, A.; Sabetta, L.; Fonnesu, A.; Mouillot, D.; Do Chi, T.; Viaroli, P.; Giordani, G.; Reizopoulou, S.; Abbiati, M.; Carrada, G.C. Typology in Mediterranean Transitional Waters: New Challenges and Perspectives. Aquat. Conserv. Mar. Freshw. Ecosyst. 2006, 16, 441–455. [Google Scholar] [CrossRef]
- Basset, A.; Pinna, M.; Sabetta, L.; Barbone, E.; Galuppo, N. Hierarchical Scaling of Biodiversity in Lagoon Ecosystems. Transit. Waters Bull. 2008, 2, 75–86. [Google Scholar] [CrossRef]
- Gravina, M.F.; Cabiddu, S.; Como, S.; Floris, A.; Padedda, B.M.; Pusceddu, A.; Magni, P. Disentangling Heterogeneity and Commonalities in Nanotidal Mediterranean Lagoons through Environmental Features and Macrozoobenthic Assemblages. Estuar. Coast. Shelf Sci. 2020, 237, 106688. [Google Scholar] [CrossRef]
- Barghini, P.; Silvi, S.; Aquilanti, A.; Marcelli, M.; Fenice, M. Bacteria from Marine Salterns as a Model of Microorganisms Adapted to High Environmental Variations. J. Environ. Prot. Ecol. 2014, 15, 897–906. [Google Scholar]
- Oren, A. The Ecology of Dunaliella in High-Salt Environments. J. Biol. Res. 2014, 21, 23. [Google Scholar] [CrossRef] [Green Version]
- Elloumi, J.; Carrias, J.-F.; Ayadi, H.; Sime-Ngando, T.; Bouaïn, A. Communities Structure of the Planktonic Halophiles in the Solar Saltern of Sfax, Tunisia. Estuar. Coast. Shelf Sci. 2009, 81, 19–26. [Google Scholar] [CrossRef]
- Franciscolo, M.E. Coleoptera. Haliplidae, Hygrobiidae, Gyrinidae, Dytiscidae. In Fauna d’Italia; Edizioni Calderini: Bologna, Italy, 1979; 804p. [Google Scholar]
- Toledo, M. Revision in Part of the Genus Nebrioporus Régimbart, 1906, with Emphasis on the N. laeviventris-Group (Coleoptera: Dytiscidae). Zootaxa 2009, 2040, 1–111. [Google Scholar] [CrossRef]
- Cimmaruta, R.; Blasi, S.; Angeletti, D.; Nascetti, G. The Recent History of the Tarquinia Salterns Offers the Opportunity to Investigate Parallel Changes at the Habitat and Biodiversity Levels. Transit. Waters Bull. 2011, 4, 53–59. [Google Scholar] [CrossRef]
- Sangiorgio, F.; Pinna, M.; Gravili, C. Macroinvertebrati bentonici. In Nuovi Approcci Metodologici per la Classificazione dello Stato di Qualità Degli Ecosistemi Acquatici di Transizione; Basset, A., Sangiorgio, F., Sabetta, L., Eds.; ISPRA, Roma—Università del Salento: Lecce, Italy, 2009; pp. 64–97. [Google Scholar]
- Anderson, M.J. A New Method for Non-Parametric Multivariate Analysis of Variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]
- Clarke, K.; Gorley, R.; Somerfield, P.; Warwick, R. Change in Marine Communities: An Approach to Statistical Analysis; Primer-E Ltd: Plymouth, UK, 2014; 14p. [Google Scholar]
- Semprucci, F.; Gravina, M.F.; Magni, P. Meiofaunal Dynamics and Heterogeneity along Salinity and Trphic Gradients in a Mediterranean Transitional System. Water 2019, 11, 1488. [Google Scholar] [CrossRef] [Green Version]
- Foti, A.; Fenzi, G.A.; Di Pippo, F.; Gravina, M.F.; Magni, P. Testing the Saprobity Hypothesis in a Mediterranean Lagoon: Effects of Confinement and Organic Enrichment on Benthic Communities. Mar. Environ. Res. 2014, 99, 85–94. [Google Scholar] [CrossRef]
- Galuppo, N.; Maci, S.; Pinna, M.; Basset, A. Habitat Types and Distribution of Benthic Macroinvertebrates in a Transitional Water Ecosystem: Alimini Grande (Puglia, Italy). Transit. Waters Bull. 2007, 1, 9–19. [Google Scholar] [CrossRef]
- Whitfield, A.K.; Elliott, M.; Basset, A.; Blaber, S.J.M.; West, R.J. Paradigms in Estuarine Ecology—A Review of the Remane Diagram with a Suggested Revised Model for Estuaries. Estuar. Coast. Shelf Sci. 2012, 97, 78–90. [Google Scholar] [CrossRef]
- Zapparoli, M.; De Mattheis, E.; Vigna Taglianti, A. Invertebrati terrestri e dulcacquicoli della Riserva Naturale Saline di Tarquinia e delle aree adiacenti. In La Riserva Naturale Statale “Saline di Tarquinia”; Colletti, L., Ed.; Corpo Forestale dello Stato, Ufficio Territoriale della Biodiversità di Roma: Rome, Italy, 2014; pp. 159–171. [Google Scholar]
- Angelini, F. Catalogo topografico Hydroadephaga dei Coleoptera. Haliplidae, Hygrobiidae, Gyrinidae, Dytiscidae d’Italia. Mem. Soc. Entomol. Ital. 1984, 41, 45–126. [Google Scholar]
- Nardi, G.; Maltzeff, P. Gli Idroadefagi della Tenuta Presidenziale di Castelporziano (Coleoptera, Gyrinidae, Haliplidae, Noteridae, Hygrobiidae, Dytiscidae). Boll. Ass. Romana Entomol. 2001, 56, 175–232. [Google Scholar]
- Di Giulio, A.; Nardi, G. Description of the pupa of Nebrioporus ceresyi (Aubé) (Coleoptera: Dytiscidae). Aquat. Insects 2006, 28, 269–275. [Google Scholar] [CrossRef]
- Latella, L.; Ruffo, S.; Stoch, F. The project CKmap (Checklist and distribution of the Italian fauna) methods and informatical techniques. Mem. Mus. Civ. St. Nat. Verona 2007, 17, 15–19. [Google Scholar]
- Bologna, M.A.; Bonato, L.; Cianferoni, F.; Minelli, A.; Oliverio, M.; Stoch, F.; Zapparoli, M. Towards the new Checklist of the Italian fauna. Biogeogr.-J. Integr. Biogeogr. 2022, 37, 1–6. [Google Scholar] [CrossRef]
- Luigioni, P. I coleotteri d’Italia. Catalogo Sinonimico—Topografico -Bibliografico. Mem. Pontif. Accad. Sci. I Nuovi Lincei 1929, 13, 1160. [Google Scholar]
- Porta, A. Fauna Coleopterorum Italica. Supplementum II; Stabilimento Tipografico Piacentino: Piacenza, Italy, 1949; 386p. [Google Scholar]
- Angelini, F. Hydroadephaga inediti per Puglie e Lucania (Coleoptera. Haliplidae, Hygrobiidae, Gyrinidae, Dytiscidae). Boll. Soc. Entomol. Ital. 1972, 104, 179–194. [Google Scholar]
- Angelini, F. Hydroadephaga nuovi per Calabria e Sila. Boll. Soc. Entomol. Ital. 1973, 105, 7–12. [Google Scholar]
- Fery, H.; Fresneda, J.; Millán, A. Bemerkungen zur Nebrioporus ceresyi-Gruppe sowie Beschreibung von Nebrioporus schoedli n. sp. (Coleoptera: Dytiscidae). Entomol. Z. 1996, 106, 306–328. [Google Scholar]
- Ragusa, E. Coleotteri nuovi o poco conosciuti della Sicilia. Nat. Sicil. 1882, 1, 248–251. [Google Scholar]
- Ragusa, E. Catalogo ragionato dei Coleotteri di Sicilia (1° parte). Nat. Sicil. 1887, 6, 221–228. [Google Scholar]
- Ragusa, E. Catalogo dei Coleotteri di Sicilia. Nat. Sicil. 1891, 10, 1–32. [Google Scholar]
- Focarile, A. Ricerche entomologiche nell’arcipelago delle Eolie e nell’isola di Ustica (Sicilia). La coleotterofauna dello stagno salmastro a Punta Lingua nell’Isola di Salina. Mem. Soc. Entomol. Ital. 1972, 51, 19–37. [Google Scholar]
- Detter, K. Adephagan water beetles of Elba Island (Tuscany) (Coleoptera Haliplidae, Dytiscidae, Noteridae, Gyrinidae). Mem. Soc. Entomol. Ital. 2006, 85, 85–122. [Google Scholar] [CrossRef] [Green Version]
- Baccetti, N.; Dall’Antonia, P.; Magagnoli, P.; Melega, L.; Serra, L.; Soldatini, C.; Zenatello, M. Risultati dei Censimenti Degli Uccelli Acquatici Svernanti in Italia: Distribuzione, Stima e Trend delle Popolazioni nel 1991–2000; ISPRA: Rome, Italy, 2002; 240p. [Google Scholar]
- Gravina, M.F.; Colozza, N.; D’Ambrosio, L.; Giorgi, M.; Martinoli, M.; Talarico, L.; Tancioni, L. Monitoraggio Dell’habitat “Lagune Costiere” Di Interesse Comunitario Prioritario Presente Nella Riserva Naturale Statale “Saline di Tarquinia”. Relazione Finale. Giugno 2016; Corpo Forestale dello Stato, Ufficio Territoriale per la Biodiversità di Roma: Rome, Italy, 2016. [Google Scholar]
- Gravina, M.F.; Bontempi, D.; D’Ambrosio, L.; Giorgi, M.; Talarico, L.; Ventura, D. Monitoraggio Dell’habitat “Lagune Costiere” Di Interesse Comunitario Prioritario Presente Nella Riserva Naturale Statale “Saline di Tarquinia”. Relazione Finale. Dicembre 2017; Raggruppamento Carabinieri Biodiversità—Reparto Biodiversità di Roma—Nucleo Carabinieri Biodiversità di Saline di Tarquinia: Roma, Italy, 2017. [Google Scholar]
- Gravina, M.F.; Giorgi, M.; Puthod, P. Monitoraggio Dell’habitat “Lagune Costiere” di Interesse Comunitario Prioritario Presente Nella Riserva Naturale Statale “Saline di Tarquinia”. Relazione Finale. Giugno 2021; Raggruppamento Carabinieri Biodiversità—Reparto Biodiversità di Roma—Nucleo Carabinieri Biodiversità di Saline di Tarquinia: Roma, Italy, 2021. [Google Scholar]
- Zerunian, S. Pesci delle acque interne d’Italia. In Quaderni di Conservazione della Natura; Ministero dell’Ambiente—Istituto Nazionale Fauna Selvatica: Bologna, Italy, 2004; Volume 20. [Google Scholar]
Figure 1.
Location of sampling sites (A, B, and C) in the Saline di Tarquinia salterns. The symbol in the top-right map indicates the location of the study site. Image source: Google Earth.
Figure 1.
Location of sampling sites (A, B, and C) in the Saline di Tarquinia salterns. The symbol in the top-right map indicates the location of the study site. Image source: Google Earth.

Figure 2.
Mean values (±SE) for salinity, dissolved oxygen (DO), temperature and pH at sites A, B and C in each month of the year.
Figure 2.
Mean values (±SE) for salinity, dissolved oxygen (DO), temperature and pH at sites A, B and C in each month of the year.

Figure 3.
Mean values (±SE) for macrozoobenthic community synthetic measures at sites A (white), B (grey) and C (black) in spring (Sp), summer (Su), autumn (Au) and winter (Wi).
Figure 3.
Mean values (±SE) for macrozoobenthic community synthetic measures at sites A (white), B (grey) and C (black) in spring (Sp), summer (Su), autumn (Au) and winter (Wi).

Figure 4.
nMDS of macrozoobenthos abundance (stress = 0.12) with superimposed environmental variables.
Figure 4.
nMDS of macrozoobenthos abundance (stress = 0.12) with superimposed environmental variables.

Figure 5.
Nebrioporus ceresyi: (a) late larva; (b) imago, dorsal view; (c) detail of the posterior angles of the pronotum; (d) detail of the punctuation of the elytra; (e) tarsomeres of left limb, dorsal view; (f) apex of the elytra.
Figure 5.
Nebrioporus ceresyi: (a) late larva; (b) imago, dorsal view; (c) detail of the posterior angles of the pronotum; (d) detail of the punctuation of the elytra; (e) tarsomeres of left limb, dorsal view; (f) apex of the elytra.

Figure 6.
Number of specimens of Nebrioporus ceresyi, both larval and adult, during the year in sites B (a) and C (b).
Figure 6.
Number of specimens of Nebrioporus ceresyi, both larval and adult, during the year in sites B (a) and C (b).

Figure 7.
Distribution of the number of specimens by the size of the larvae of Nebrioporus ceresyi in April (a) and May (b).
Figure 7.
Distribution of the number of specimens by the size of the larvae of Nebrioporus ceresyi in April (a) and May (b).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Results of two-way PERMANOVA (Bray-Curtis-(dis)similarity-based) testing differences between sites and seasons and their interactions for each of the considered environmental parameters.
Table 1.
Results of two-way PERMANOVA (Bray-Curtis-(dis)similarity-based) testing differences between sites and seasons and their interactions for each of the considered environmental parameters.
| Variable | Factors | Total Sum of Squares | Df | Mean Square | Pseudo-F | p |
|---|---|---|---|---|---|---|
| Sites | 2.26 | 2 | 1.13 | 142.31 | 0.0001 | |
| Seasons | 0.11 | 3 | 0.036 | 4.56 | 0.0041 | |
| Salinity | Interaction | 0.13 | 6 | 0.022 | 2.73 | 0.014 |
| Residual | 0.62 | 78 | 0.0079 | |||
| Total | 3.11 | 89 | ||||
| Sites | 57.37 | 2 | 28.69 | 7.85 | 0.0005 | |
| Seasons | 27.41 | 3 | 9.14 | 2.50 | 0.061 | |
| Dissolved oxygen | Interaction | 20.65 | 6 | 3.44 | 0.94 | 0.47 |
| Residual | 321.64 | 88 | 3.65 | |||
| Total | 427.07 | 99 | ||||
| Sites | 0.094 | 2 | 0.047 | 3.40 | 0.036 | |
| Seasons | 1.39 | 3 | 0.46 | 33.4 | 0.0001 | |
| Temperature | Interaction | 0.019 | 6 | 0.0032 | 0.23 | 0.98 |
| Residual | 1.22 | 88 | 0.0139 | |||
| Total | 2.72 | 99 | ||||
| Sites | 0.014 | 2 | 0.0071 | 21.9 | 0.0001 | |
| Seasons | 0.00045 | 3 | 0.00015 | 0.46 | 0.71 | |
| pH | Interaction | 0.0046 | 6 | 0.00076 | 2.35 | 0.035 |
| Residual | 0.029 | 88 | 0.00032 | |||
| Total | 0.048 | 99 |
Table 2.
Results of Similarity Percentage (SIMPER) analysis showing the average dissimilarity (Av. Dissim.), the contribution of each taxon (Contrib. %), the cumulative value for the considered site couple (Cum. %) and the average abundance of each site (Mean ab.).
Table 2.
Results of Similarity Percentage (SIMPER) analysis showing the average dissimilarity (Av. Dissim.), the contribution of each taxon (Contrib. %), the cumulative value for the considered site couple (Cum. %) and the average abundance of each site (Mean ab.).
| A vs. B | |||||
|---|---|---|---|---|---|
| Taxa | Av. Dissim. | Contrib. % | Cum. % | Mean ab. A | Mean ab. B |
| Gammarus aequicauda | 9.65 | 20.04 | 20.04 | 1.40 | 0.21 |
| Chironomidae | 7.63 | 15.85 | 35.89 | 1.50 | 2.06 |
| Hydrobia acuta | 6.05 | 12.56 | 48.45 | 1.88 | 2.20 |
| Cerastoderma glaucum | 5.37 | 11.14 | 59.60 | 0.40 | 0.60 |
| Idotea chelipes | 3.87 | 8.04 | 67.64 | 0.62 | 0.016 |
| Gammaridae juv. | 2.73 | 5.68 | 73.32 | 0.38 | 0.10 |
| Monocorophium insidiosum | 2.63 | 5.47 | 78.79 | 0.44 | 0 |
| Nemertea | 2.33 | 4.84 | 83.64 | 0.34 | 0.025 |
| Cerithium vulgatum | 2.08 | 4.31 | 87.95 | 0.24 | 0 |
| Nebrioporus ceresyi | 0.99 | 2.06 | 90.01 | 0 | 0.13 |
| Perinereis cultrifera | 0.96 | 2.00 | 92.01 | 0.14 | 0.032 |
| Corophiidae juv. | 0.88 | 1.83 | 93.84 | 0.16 | 0 |
| Nereididae juv. | 0.74 | 1.53 | 95.37 | 0.11 | 0.016 |
| Monocorophium sextonae | 0.72 | 1.49 | 96.87 | 0.13 | 0 |
| Melita palmata | 0.68 | 1.41 | 98.28 | 0.068 | 0.016 |
| Neodexiospira pseudocorrugata | 0.42 | 0.86 | 99.14 | 0.053 | 0 |
| Victorella pavida | 0.41 | 0.86 | 100 | 0 | 0.061 |
| Ephydra bivittata | 0 | 0 | 100 | 0 | 0 |
| A vs. C | |||||
| Taxa | Av. Dissim. | Contrib. % | Cum. % | Mean ab. A | Mean ab. C |
| Hydrobia acuta | 15.43 | 22.30 | 22.30 | 1.88 | 0.37 |
| Gammarus aequicauda | 11.88 | 17.17 | 39.47 | 1.40 | 0 |
| Chironomidae | 10.66 | 15.40 | 54.87 | 1.50 | 2.17 |
| Nebrioporus ceresyi | 6.46 | 9.33 | 64.21 | 0 | 0.64 |
| Idotea chelipes | 4.45 | 6.42 | 70.63 | 0.62 | 0 |
| Cerastoderma glaucum | 3.73 | 5.39 | 76.02 | 0.40 | 0.028 |
| Monocorophium insidiosum | 3.02 | 4.36 | 80.39 | 0.44 | 0 |
| Nemertea | 2.65 | 3.83 | 84.21 | 0.34 | 0 |
| Gammaridae juv. | 2.53 | 3.66 | 87.87 | 0.38 | 0 |
| Cerithium vulgatum | 2.52 | 3.64 | 91.51 | 0.24 | 0 |
| Ephydra bivittata | 1.16 | 1.68 | 93.19 | 0 | 0.13 |
| Corophiidae | 1.00 | 1.44 | 94.63 | 0.16 | 0 |
| Perinereis cultrifera | 0.94 | 1.36 | 95.99 | 0.14 | 0 |
| Monocorophium sextonae | 0.82 | 1.18 | 97.17 | 0.13 | 0 |
| Nereididae juv. | 0.76 | 1.10 | 98.26 | 0.11 | 0 |
| Melita palmata | 0.71 | 1.02 | 99.28 | 0.068 | 0 |
| Neodexiospira pseudocorrugata | 0.72 | 0.72 | 100 | 0.053 | 0 |
| Victorella pavida | 0 | 0 | 100 | 0 | 0 |
| B vs. C | |||||
| Taxa | Av. Dissim. | Contrib. % | Cum. % | Mean ab. B | Mean ab. C |
| Hydrobia acuta | 21.42 | 42.73 | 42.73 | 2.20 | 0.37 |
| Chironomidae | 9.23 | 18.42 | 61.15 | 2.06 | 2.17 |
| Nebrioporus ceresyi | 7.35 | 14.67 | 75.82 | 0.13 | 0.64 |
| Cerastoderma glaucum | 6.07 | 12.11 | 87.94 | 0.60 | 0.028 |
| Gammarus aequicauda | 1.99 | 3.97 | 91.91 | 0.21 | 0 |
| Ephydra bivittata | 1.37 | 2.73 | 94.64 | 0 | 0.13 |
| Gammaridae juv. | 0.99 | 1.98 | 96.62 | 0.10 | 0 |
| Victorella pavida | 0.56 | 1.12 | 97.75 | 0.061 | 0 |
| Perinereis cultrifera | 0.37 | 0.73 | 98.48 | 0.032 | 0 |
| Nemertea | 0.26 | 0.51 | 98.99 | 0.025 | 0 |
| Melita palmata | 0.21 | 0.42 | 99.41 | 0.016 | 0 |
| Idotea chelipes | 0.15 | 0.31 | 99.71 | 0.016 | 0 |
| Nereididae juv. | 0.14 | 0.29 | 100 | 0.016 | 0 |
| Neodexiospira pseudocorrugata | 0 | 0 | 100 | 0 | 0 |
| Monocorophium sextonae | 0 | 0 | 100 | 0 | 0 |
| Monocorophium insidiosum | 0 | 0 | 100 | 0 | 0 |
| Corophiidae juv. | 0 | 0 | 100 | 0 | 0 |
| Cerithium vulgatum | 0 | 0 | 100 | 0 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Bonifazi, A.; Galli, S.; Gravina, M.F.; Ventura, D. Macrozoobenthos Structure and Dynamics in a Mediterranean Hypersaline Ecosystem with Implications for Wetland Conservation. Water 2023, 15, 1411. https://doi.org/10.3390/w15071411
AMA Style
Bonifazi A, Galli S, Gravina MF, Ventura D. Macrozoobenthos Structure and Dynamics in a Mediterranean Hypersaline Ecosystem with Implications for Wetland Conservation. Water. 2023; 15(7):1411. https://doi.org/10.3390/w15071411
Chicago/Turabian StyleBonifazi, Andrea, Simone Galli, Maria Flavia Gravina, and Daniele Ventura. 2023. "Macrozoobenthos Structure and Dynamics in a Mediterranean Hypersaline Ecosystem with Implications for Wetland Conservation" Water 15, no. 7: 1411. https://doi.org/10.3390/w15071411
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.