1. Introduction
Rice is an important staple crop whose production is crucial to global food security [
1]. In current agricultural practice, diseases and insect pests are significant challenges to safe and sustainable rice production, and reducing their impact is critical for ensuring continued food security [
2].
Chilo suppressalis is an economically important pest, which can cause serious damage to rice. It is estimated that the loss caused by
C. suppressalis accounts for 10–30% of total rice production [
3]. In many rice-growing countries in Asia and Africa, rice blast has re-emerged as the main factor affecting the stable production of rice and food security. For example, in the highlands of eastern India alone, 50% of rice production is lost on average every year [
4]. From 2005 to 2006, serious pests occurred in southern Vietnam. More than 485,000 hectares of rice fields were affected by viral diseases that seemed to be transmitted by
N. lugens and other insect vectors, resulting in a loss of 120 million US dollars [
5]. At present, chemical prevention is still the main method of ensuring stable yields; however, long-term pesticide use, especially the excessive use of chemical pesticides, has enabled many pests to develop resistance to commonly used chemical insecticides and fungicides [
6]. Cloning disease and pest resistance genes and incorporating these genes into new rice varieties through molecular breeding is therefore crucial for protecting the environment and promoting sustainable agricultural development [
7].
Most cultivated rice varieties were bred for strong single resistance traits. However, in actual production, a variety of diseases and pests typically occur simultaneously or successively [
8]. To prevent loss of resistance after large-scale planting of resistant varieties, it is necessary to further expand the sources of resistance genes, express resistance genes with different resistance spectra, and develop multi-gene-resistance varieties with strong field adaptability. Many breeders have therefore begun to combine multiple resistance genes in the same elite variety in hopes of extending its useful lifespan [
9]. Aggregating different resistance genes into one variety can not only further improve its resistance level but also enhance its broad-spectrum and durable resistance. Therefore, aggregation of two or more resistance genes into the same variety is an important breeding goal [
10].
Gene stacking breeding involves the collection and stacking of target genes, followed by the fixation of polymeric genes, that is, their homozygous stacking [
11]. There are two basic approaches to plant molecular breeding by gene stacking [
12]. The first is molecular breeding by gene stacking and genetic transformation. This method combines conventional breeding techniques with plant transformation methods such as plant DNA virus–mediated transformation,
Agrobacterium-mediated transformation, microinjection, electric shock, and pollen tube pathway–mediated transformation. Such strategies are used to transfer two or more artificially modified and isolated genes into the recipient plant. Expression of the imported genes causes heritable modifications of plant characteristics, enabling the creation of new plant varieties with specific target traits. The second approach involves gene aggregation by molecular marker–assisted selection. In this method, favorable genes are aggregated into the same genome through techniques such as pair hybridization, synthetic hybridization, additive hybridization, multi-paternity mixed pollination hybridization, and backcrossing. Using molecular markers to select a single plant with multiple target genes from offspring, and then select highly resistant strains to realize the stacking of favorable genes.
A number of studies in rice and maize demonstrate the possibilities of gene stacking breeding. Sequence-tagged site (STS) marker-assisted selection was used to pyramid two bacterial blight resistance genes,
Xa21 and
Xa13, and one aroma gene,
(fgr), into multiple transgenic rice lines. In the BC
2F
3 and BC
3F
3 generations, the transgenic plants exhibited a wider resistance spectrum and stronger resistance than plants with a single resistance gene [
13]. Similarly, the rice blast resistance genes
Pi-ta,
Pi-b, and
Pi-d(t) from Pi-4, BL-1, and Digu were polymerized into G46B, significantly improving its blast resistance [
14]. Molecular marker-assisted selection and backcross breeding techniques can therefore effectively improve rice resistance.
The use of diverse resistance genes is important for coping with variations among biotypes or species of pests and pathogens and for enhancing the breadth and durability of plant resistance [
15]. In this study, resistance genes carried in rice lines MH63, CK30, and PSL were transferred into three rice restorer lines, CH121, CH891, and R205X, by molecular marker-assisted selection and backcrossing techniques. A group of polygenic improved lines carrying resistance genes to
C. suppressalis (striped rice stemborer),
Magnaporthe oryzae (rice blast), and
N. lugens (brown planthopper) were identified, and their resistance levels and agronomic characteristics were comprehensively evaluated.
2. Materials and Methods
2.1. Plant Materials
The donor parents were MH63(
CRY1C), MH63(
CRY2A), CK30, and PSL. MH63(
CRY1C) and MH63(
CRY2A), which carry the
C. suppressalis resistance genes
CRY1C and
CRY2A, were provided by Huazhong Agricultural University.
Figures S1 and S2 shows the information of transgenes-
CRY 1C and
CRY 2A. CK30 carries the
Pibgom and
Pikm rice blast resistance genes and was provided by Sichuan Agricultural University. PSL carries the
N. lugens resistance gene
Bph29 and was provided by Chengdu Institute of Biology, Chinese Academy of Sciences. The acceptor parents were CH121, CH891, and R205X, high-quality restorer lines bred by Jiangxi Agricultural University. They have good adaptability in the middle and lower reaches of the Yangtze River, the main rice producing area in China. Descriptions of plant materials description are shown in
Table 1.
Hybridization and breeding were performed at the transgenic experimental base of Jiangxi Agricultural University, Nanchang City, Jiangxi Province beginning in 2010. With two seasons per year, we obtained stable genetic strains by 2020. At the beginning of the experiment, the upper 15 cm of soil at the test site had the following properties: pH 5.01, 1.26 g kg−1 total N, 105.6 mg kg−1 available P, 125.2 mg kg−1 K, and 20.56 g kg−1 organic matter. The study was conducted at the Transgenic Test Base of Jiangxi Agricultural University (28°48′10″ N, 115°49′55″ E) in the economic and technological development zone, Nanchang City, Jiangxi Province during the 2020 rice growing season.
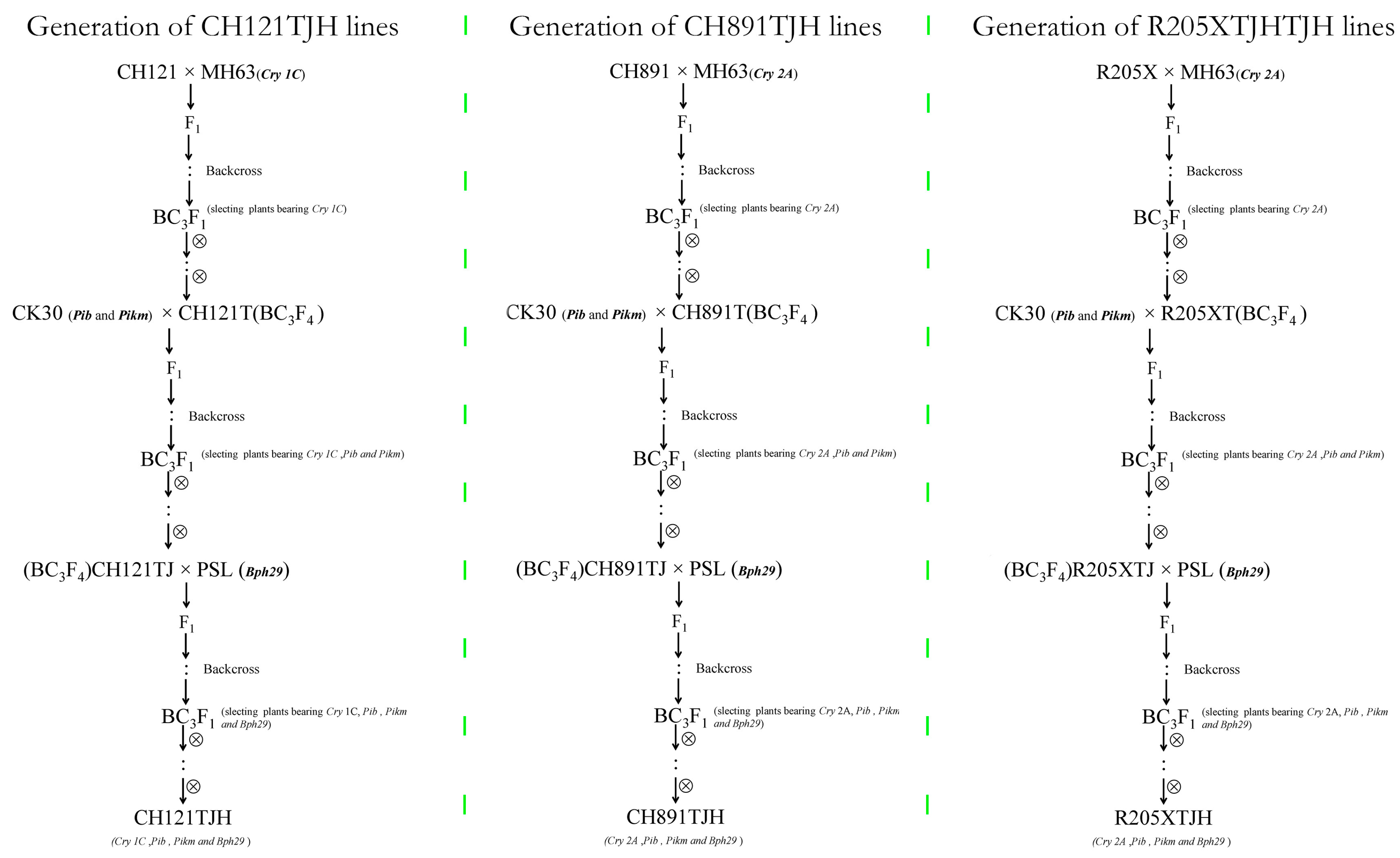
For CRY1C, CH121 was the recipient parent, and MH63(CRY1C) carrying the CRY1C borer resistance gene was the donor parent. For CRY2A, CH891 and R205X were the recipient parents, and MH63(CRY2A) carrying the CRY2A borer resistance gene was the donor parent. Through backcrossing, heterozygous single plants carrying the target genes were screened out from the BC1F1 generation. Among these heterozygous single plants, individuals with a leaf morphology similar to that of their respective recurrent parent were further screened out until the BC3F1 generation. The BC3F2 population obtained after self-crossing was screened using molecular markers, and excellent single strains carrying homozygous target genes were selected. The BC3F4 population was obtained by self-crossing, and a stable line with a leaf morphology similar to the recurrent parent and the insect resistance gene CRY1C was selected and named CH121T. The CH891T and R205XT lines containing the CRY2A gene were produced using the same process. CH121T, CH891T, and R205XT were then used as the receptor parents and CK30 as the donor parent to obtain lines with the rice blast resistance genes Pib and Pikm and agronomic traits similar to those of their parents; these lines were named CH121TJ, CH89TJ1, and R205XTJ. Similarly, Bph29 was added to CH121TJ, CH89TJ, and R205XTJ by backcrossing with PSL. The polymeric resistance gene varieties CH121TJH (containing CRY1C, Pib, Pikm, and Bph29) and CH89TJH and R205XTJH (containing CRY2A, Pib, Pikm, and Bph29) were obtained by self-crossing and provided new genetic materials for subsequent resistance experiments.
2.2. Molecular Marker-Assisted Detection
Genomic DNA was extracted from the parent varieties and the new varieties with polymerized resistance genes by the CTAB method [
16]. The parent varieties were MH63(
Cry1C), MH63(
Cry2A), CK30, PSL, CH121, CH891, and R205X, and the varieties with polymerized resistance genes were CH121TJH, CH89TJH, and R205XTJH. DNA concentration and purity were assessed using a NanoDrop 2000 spectrophotometer (Thermo Scientific) based on the absorbance ratio of 260/280 nm [
17]. The diluted DNA was amplified by PCR using the primer sequences shown in
Table S1. The PCR products were separated and visualized by agarose gel electrophoresis. The band sizes of
CRY1C and
CRY2A were 799 bp and 600 bp. The
Pikm blast resistance gene is composed of two closely related, adjacent genes,
Pikm1 and
Pikm2. They were amplified using the specific primers DKM1 and DKM2 and had band sizes of 191 bp and 223 bp. Previous work has shown that blast resistance requires the presence of both bands in a single plant [
18]. Primers for
Pib included
Pib, which detects the presence of the resistant allele (band size 360 bp) and
Lys145, which detects the presence of the susceptible allele (800 bp). The appearance of both bands indicated that the tested material was heterozygous [
19]. The band produced by amplification of the
N. lugens resistance gene
Bph29 was 570 bp.
2.3. Genetic Background Detection Based on SSR Markers
Whole genomes of the recurrent parents CH121, CH891, and R205X and the corresponding new strains CH121TJH, CH891TJH, and R205XTJH were analyzed using 560 pairs of SSR primers evenly distributed on the 12 rice chromosomes. Polymorphic primer markers were identified, and the genetic background recovery rate was analyzed. The SSR primer sequence reference (
http://www.gramene.org/microsat/ accessed on 1 October 2021.) was synthesized by Beijing Tsingske Biotechnology Company. Diluted genomic DNA was used for PCR amplification, and the PCR products were separated by SDS–PAGE on an 8% polyacrylamide gel with silver staining. The physical locations of molecular markers are shown in
Table S2. A single plant with a homozygous band pattern was marked as 1, a single plant with a heterozygous band pattern was marked as 2, and a single plant with no bands was marked as 0.
The numb of that polymorphic molecular markers screen out between the donor parent and each corresponding parent is L, and further screening is carried out between the correspond parent and the polymeric resistance gene strain to obtain the number of the molecular markers of the same band type between the corresponding polymeric resistance gene strain and the recurrent new strain is X. According to the actual recovery rate formula [
20]:
R[G(g)] is actual recovery rate after backcrossing g generations. The actual recovery rate of that rice plant line with the stacking resistance gene can be calculated (
Table S8) through one-time hybridization, three-time backcross, and three-time selfing, according to a theoretical recovery rate formula:
G(g) is theoretical recovery rate after backcrossing g generations. The theoretical recovery rate of each transgenic line after the introduction of the CRY1C or CRY 2A gene was calculated to be 93.75%; Through one-time hybridization, three times of backcross and three times of selfing, the theoretical recovery rate of each transgenic plant line after the rice blast resistance gene is introduced is calculated to be 88.25%; and finally, after the Bph 29 gene is introduced through one-time hybridization, three times of backcross and three times of selfing, the theoretical recovery rate of each transgenic plant line after the anti-rice blast gene is introduced is calculated to be 83.34%, and the recovery rate is the final theoretical recovery rate of the polymerized gene rice.
2.4. Insect Resistance in the Laboratory
Insect feeding assays were performed using 10 replicates of each transgenic line and non-transgenic control. In August 2021, female
C. suppressalis were captured using light in the experimental field of Jiangxi Agricultural University, then maintained at 28 °C under humid conditions for spawning. When black spots gradually appeared in the eggs, they were transferred to fresh stalks of rice without
C. suppressalis resistance, and the larvae were allowed to feed for about 10 days [
21]. Second instar larvae were fed fresh transgenic rice leaves and stems that had been collected from rice plants at the tillering and heading stages; non-transgenic rice tissues at the same growth stages were used as negative controls. Second instar larvae were placed in individual Petri dishes that contained a piece of leaf (4 g) or stem (5 g), and ddH
2O was added to the filter paper to maintain humid conditions. The initial number of larvae per Petri dish was 30. Petri dishes were sealed with parafilm membranes to prevent larvae from escaping. All Petri dishes were stored in a hermetic box in the dark at 27 ± 1 °C and 75 ± 5% relative humidity [
22]. Larval mortality was determined after 48 h, and the mortality rate was calculated. The calculations [
23] and statistical methods are shown in
Table S3.
Statistical equation of larval mortality:
Statistical equation of corrected larval mortality:
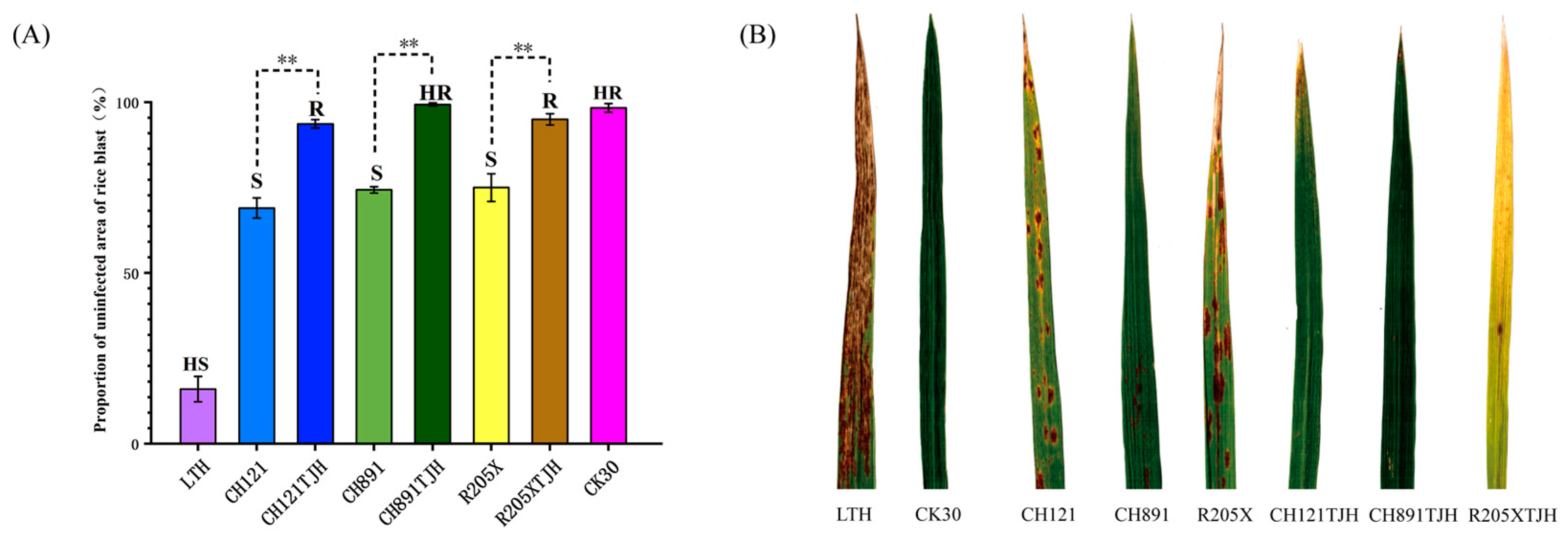
2.5. Assessment of Rice Blast Resistance at the Seedling Stage
The control materials were CK30 (with high rice blast resistance), LTH (with high rice blast sensitivity), CH121, CH891, and R205X. The experimental materials were the new lines CH121TJH, CH891TJH, and R205XTJH. Two strains of
M. oryzae were isolated and purified from a field in Jiangxi Province by researchers from Jiangxi Academy of Agricultural Sciences and named F1-6 and 253. PDA medium was used as the activation medium and rice bran medium as the sporulation medium (formulas shown in
Table S4). Spores of
M. oryzae were activated on PDA medium for 7 days and then transferred to rice bran medium. The two strains were cultured separately. After inoculation, both media were cultured in an incubator at 28 °C. After mycelium overgrew on the rice bran medium plate (about 7 days), it was placed on a tissue culture rack at 25–28 °C for 24 h of light culture. After full sporulation (about 5 days), conidia of
M. oryzae were eluted with 0.02% TWEEN-20; this procedure was repeated 2–3 times, and the eluant was filtered through sterilized gauze. The filtered spore suspensions were counted at 100× under a microscope and inoculated when there was an average of 30–50 spores per field of view [
24]. The concentration of spores was about 30 × 10
4 conidia/mL.
Seeds of the experimental and control groups were immersed in clear water at 25 °C for 48 h, germinated at 35 °C, and then sown in seedling trays. When rice seedlings had grown to the 3–4 leaf stage, spray inoculation was performed in an inoculation box using an airbrush with pressurized air. Spore suspension (40 mL) was sprayed onto each seedling tray, and there were three replicate trays per genotype and treatment. Control trays were sprayed with water only. After inoculation, trays were maintained in a dark, humid incubator at 28 °C. The incidence of rice blast was investigated 10 days after inoculation; incidence, lesion size, and lesion number were recorded on the basis of visual observation. The disease condition was recorded, and the resistance level was determined according to the six-point scale of Mackill et al. [
25]. The identification criteria are shown in
Table S5.
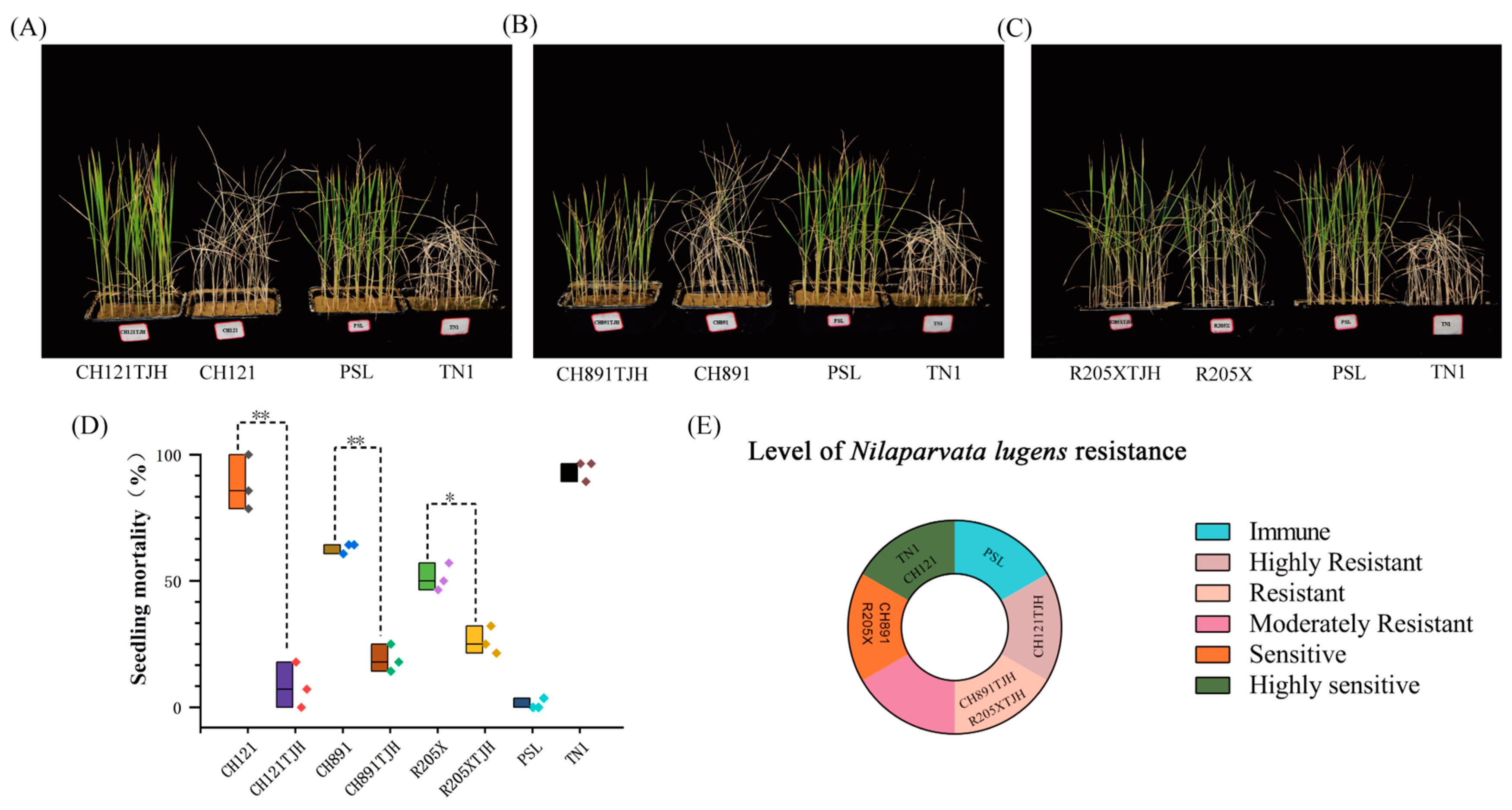
2.6. Identification of Resistance to N. lugens at the Seedling Stage
The Standard Seedbox Screening Technique (SSST) [
26] was used to determine the resistance of rice seedlings to
N. lugens. The mixed biotype insect source was collected from the experimental field of Jiangxi Agricultural University and was fed with conventional rice that lacked the
N. lugens resistance gene. Seeds of new materials (CH121TJH, CH891TJH, and R205XTJH), recurrent parents (CH121, CH891, and R205X), and control materials (susceptible TN1 and resistant PSL) were immersed in clear water at 25 °C for 48 h, germinated at 35 °C, and then sown in seedling trays. Twenty-eight seeds were sown in each tray, and there were three replicate trays per genotype.
N. lugens was added when the plants reached the three-leaf stage. Six to seven nymphs of 2–3 instars of
N. lugens were added to each plate and allowed to feed and breed. When almost all TN1 plants had withered, resistance was scored on the basis of damage symptoms using the criteria shown in
Table S6.
2.7. Measurement of Agronomic Traits
Transgenic rice lines were planted in paddy fields at the Transgenic Experimental Plots of Jiangxi Agricultural University (Nanchang, Jiangxi, China) to evaluate their agronomic performance. Non-transgenic lines CH121, CH891 and R205X, were planted in paddy fields adjacent to the transgenic lines as controls. We used pesticides and fungicides in the cultivation process in order to create an environment to control pests and diseases. The purpose of this is to examine whether there is a huge yield difference between the transgenic rice lines and their corresponding parents under normal conditions without pests and diseases. Eight yield traits were measured: plant height, total number of grains, effective panicle number, panicle length, seed-setting rate, yield per plant, filled grain number per panicle, and 1000-grain weight. Filled and unfilled grains of the main panicle were separated manually for measurement of seed-setting rate (filled grains/(filled grains + unfilled grains) × 100). Yield per plant was calculated as panicles per plant × grains per panicle × 1000-grain weight × seed-setting rate × 10
−6. Details are shown in
Table S7. Agronomic traits of transgenic plants and the recurrent parents were compared using one-way analysis of variance (ANOVA), and values are presented as means ± SD. Figures were constructed using Origin 2017 (OriginLab Corp., Northampton, MA, USA).
4. Discussion
Diseases and pests often occur simultaneously or in succession during rice production, limiting rice yield and threatening food security [
27]. Here, we used marker-assisted backcross breeding (MABB) to aggregate the stemborer resistance gene
CRY1C/
CRY2A, the rice blast resistance genes
Pikm and
Pib, and the brown planthopper resistance gene
Bph29 into three high-quality indica rice restorer lines, obtaining the new lines CH121TJH, CH891TJH and R205XTJH. In previous research, researchers have typically aimed to optimize a plant’s resistance to a specific resistance gene by introducing multiple resistance genes. Selected three broad spectrum potato R genes (
Rpi),
Rpi-sto1 (
Solanum stoloniferum),
Rpi-vnt1.1 (
S. venturii), and
Rpi-blb3 (
S. bulbocastanum), combined into a single binary vector pBINPLUS and transformed into the susceptible cultivar Desiree. Through genetic transformation, potato had three R genes that are naturally resistant to single disease. [
28]. Two antifungal genes were stacked into transgenic pea (
Pisum sativum L.) to enhance resistance against fungal diseases, which proved the stable inheritance of the antifungal genes in the transgenic plants [
29]. Compared with the technology aiming at a single disease gene, introducing more than one gene into crop simultaneously or sequentially (called transgenic stacking) is a more effective strategy to endow transgenic plants with higher and more lasting insects and disease resistance [
30]. Two
Bacillus thuringiensis (Bt) insecticidal genes,
Cry 1Ac and
Cry 1Ig, and a modified glyphosate-tolerant 5-enolpyruvylshikimate-3-phosphate synthase (
EPSPS) gene (
G 10) were combined into Elite rice (
Oryza sativa spp.
japonica) cultivar Xiushui 134, was found to be highly resistant to striped stem borer and rice leaf roller, and tolerant to glyphosate [
31]. Three insecticidal genes (the Bt gene
Cry 1Ac and
Cry 2A, and the snowdrop lectin gene
gna) were introduced into important commercial indica rice varieties M7 and Basmati370 at the same time. The bioassay using triple transgenic plants showed that rice leaf roller and yellow rice borer were completely eradicated, and the survival rate of brown planthopper was reduced by 25%. The greatest decrease in insect survival rate and plant damage occurred in plants expressing all three transgenes lines [
32]. It is effective and feasible to stack R. gene into plants to enhance disease resistance and insect resistance. However, too much introduction of transgenes will also lead to unintended effects. Such as statistically significant differences in the phenotype, response, or composition of the GM plant compared with the parent from which it is derived [
33]. Therefore, if more cis-genes can be used instead of transgenes, the risk of these unintended effects and unknown mutations will be reduced. Some scholars believe that cis-genes does not add an extra trait [
34].
Molecular marker-assisted detection is a commonly used method in MABB. By comparing the size of a target gene amplified by specific primers between the parent and the progeny line, it is possible to determine whether the target gene has been successfully introduced [
35,
36]. MABB has previously been used to introduce
crtRB1 and
o2 into maize (
Zea mays L.) to increase β-carotene, lysine, and tryptophan levels. Likewise, genes for tolerance/resistance to submergence (
Sub1), salinity (
Saltol), rice blast (
Pi2,
Pi9), and gall midge (
Gm1,
Gm4) were identified and introduced into the rice cultivar Tapaswini, which already contained four bacterial blight resistance genes [
37]. In the present study, insect and blast resistance genes were detected using molecular markers, thereby enabling their aggregation into new rice varieties via backcross breeding, consistent with the approaches used in previous studies.
In crop backcross breeding, genetic background recovery rate is an important standard for measuring the similarity of background traits between parents and offspring [
38], and genic microsatellite markers are commonly used for this purpose [
39]. The
Sub1 site of FR13A, a submergence-tolerant variety, was introduced by MABB into CO 43, a rice variety prevalent in southern India. Genotyping and phenotyping of the BC
3F
3 generation showed that high-quality, near-isogenic lines of CO 43 contained the
Sub1 locus and had 94.37–95.78% of the recurrent parent CO 43 genome [
40]. In another study, the rice variety MR219 was used as the recurrent parent, and Pongsu Seribu 1 carrying the rice blast resistance genes
Piz,
Pi2, and
Pi9 was used as the resistance gene donor. Using seventy microsatellite markers, the recovery rate of the recurrent parent genome in the improved lines was determined to be 95.98–97.70% [
41]. After continuous backcrossing and self-crossing for multiple generations, the genetic background recovery rate of offspring of different recurrent parents will be different, even with the same donor parent. As expected, the actual recovery rates differed among the three rice lines created here; some were higher than the theoretical recovery rate, and some were slightly lower.
The presence of R gene(s) is the key to the resistance of resistant plants. After introduction of the stemborer resistance gene CRY1C, the rice blast resistance genes Pib and Pikm, and the brown planthopper resistance gene Bph29, CH121TJH showed significantly improved resistance to borers, rice blast, and brown planthopper compared with its parent CH121. The resistance levels of the parent CH121 to borers, rice blast, and brown planthopper were S, S, and HS, whereas those of CH121TJH were R, R, and HR. Introduction of CRY2A, Pib, Pikm, and Bph29 into CH891TJH and R205XTJH had a similar effect on resistance. The resistance levels to borers, rice blast, and brown planthopper were S, S, and S in CH891 and R205X. By contrast, these levels were HR, HR, and R in CH891TJH and R, R, and R in R205XTJH. Differences in resistance to the same pest or pathogen in plants with different genetic backgrounds may be related to the different expression levels of resistance genes or to the different recovery rates of their genetic backgrounds.
Agronomic traits of the resistant strains were similar to those of their recurrent parents, although there were some differences. Thousand-grain weight was slightly lower in the resistant strains, although this difference was only significant for R205XTJH. Resistant strains also tended to be taller, although again this difference was not significant. In general, introduction of resistance genes had relatively minor effects on the agronomic characters of the improved strains; only R205XTJH showed significant reductions in more than one trait, and CH891TJH showed no significant reductions in any trait.
The two varieties CH121TJH and CH891TJH with actual recovery rates higher than the theoretical recovery rate showed good performance in inheriting the resistance of the donor parents, and in only one case did an agronomic trait differ from that of the original parents (total grain number in CH121TJH). R205XTJH, whose actual recovery rate was slightly lower than the theoretical rate, also inherited the resistance of the donors parent but differed from the recurrent parent in several agronomic traits.


 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}