
Genes Participating in the Ensheathment of Neurons Are Affected by Postnatal Stress and Maternal Immune Activation in the Pituitary Gland
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Pituitary Gland RNA Sequencing and Analysis
2.2. Functional Enrichment, Network Inference, and Transcriptional Factor Analysis
3. Results
3.1. Sequencing Differential Gene Expression Descriptive
3.2. Effects of Maternal Immune Activation, Stress, and Sex on the Pituitary Biological Processes and Genes
3.3. Networks of Genes in the Ensheathment of Neurons Process Affected by Maternal Immune Activation, Stress, and Sex in the Pituitary Gland
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rymut, H.E.; Bolt, C.R.; Caputo, M.P.; Houser, A.K.; Antonson, A.M.; Zimmerman, J.D.; Villamil, M.B.; Southey, B.R.; Rund, L.A.; Johnson, R.W.; et al. Long-Lasting Impact of Maternal Immune Activation and Interaction with a Second Immune Challenge on Pig Behavior. Front. Vet. Sci. 2020, 7, 561151. [Google Scholar] [CrossRef]
- French, S.S.; Chester, E.M.; Demas, G.E. Maternal immune activation affects litter success, size and neuroendocrine responses related to behavior in adult offspring. Physiol. Behav. 2013, 119, 175–184. [Google Scholar] [CrossRef]
- Knuesel, I.; Chicha, L.; Britschgi, M.; Schobel, S.A.; Bodmer, M.; Hellings, J.A.; Toovey, S.; Prinssen, E.P. Maternal immune activation and abnormal brain development across CNS disorders. Nat. Rev. Neurol. 2014, 10, 643–660. [Google Scholar] [CrossRef] [PubMed]
- Einarsson, S.; Brandt, Y.; Lundeheim, N.; Madej, A. Stress and its influence on reproduction in pigs: A review. Acta Vet. Scand. 2008, 50, 48. [Google Scholar] [CrossRef] [PubMed]
- Crawley, J.N.; Heyer, W.D.; LaSalle, J.M. Autism and Cancer Share Risk Genes, Pathways, and Drug Targets. Trends Genet. 2016, 32, 139–146. [Google Scholar] [CrossRef]
- Campbell, J.M.; Crenshaw, J.D.; Polo, J. The biological stress of early weaned piglets. J. Anim. Sci. Biotechnol. 2013, 4, 19. [Google Scholar] [CrossRef]
- Southey, B.R.; Bolt, C.R.; Rymut, H.E.; Keever, M.R.; Ulanov, A.V.; Li, Z.; Rund, L.A.; Johnson, R.W.; Rodriguez-Zas, S.L. Impact of Weaning and Maternal Immune Activation on the Metabolism of Pigs. Front. Mol. Biosci. 2021, 8, 660764. [Google Scholar] [CrossRef] [PubMed]
- Maskal, J.M.; Brito, L.F.; Duttlinger, A.W.; Kpodo, K.R.; McConn, B.R.; Byrd, C.J.; Richert, B.T.; Marchant, J.N.; Lay, D.C., Jr.; Perry, S.D.; et al. Characterizing the postnatal hypothalamic-pituitary-adrenal axis response of in utero heat stressed pigs at 10 and 15 weeks of age. Sci. Rep. 2021, 11, 22527. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.; Shen, Q.; Wu, P.; Chen, H.; Wang, S.; Chen, D.; Yu, Y.; Guo, Z.; Wang, J.; Tang, G. Pituitary-Gland-Based Genes Participates in Intrauterine Growth Restriction in Piglets. Genes 2022, 13, 2141. [Google Scholar] [CrossRef]
- Ronovsky, M.; Berger, S.; Molz, B.; Berger, A.; D Pollak, D. Animal models of maternal immune activation in depression research. Curr. Neuropharmacol. 2016, 14, 688–704. [Google Scholar] [CrossRef] [PubMed]
- Makris, G.; Agorastos, A.; Chrousos, G.P.; Pervanidou, P. Stress System Activation in Children and Adolescents with Autism Spectrum Disorder. Front. Neurosci. 2021, 15, 756628. [Google Scholar] [CrossRef]
- Southey, B.R.; Zhang, P.; Keever, M.R.; Rymut, H.E.; Johnson, R.W.; Sweedler, J.V.; Rodriguez-Zas, S.L. Effects of maternal immune activation in porcine transcript isoforms of neuropeptide and receptor genes. J. Integr. Neurosci. 2021, 20, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Zager, A.; Andersen, M.L.; Tufik, S.; Palermo-Neto, J. Maternal immune activation increases the corticosterone response to acute stress without affecting the hypothalamic monoamine content and sleep patterns in male mice offspring. Neuroimmunomodulation 2014, 21, 37–44. [Google Scholar] [CrossRef]
- Fujitani, M.; Miyajima, H.; Otani, Y.; Liu, X. Maternal and Adult Interleukin-17A Exposure and Autism Spectrum Disorder. Front. Psychiatry 2022, 13, 836181. [Google Scholar] [CrossRef]
- Rodriguez-Zas, S.L.; Southey, B.R.; Rymut, H.E.; Rund, L.A.; Johnson, R.W. Hippocampal Changes Elicited by Metabolic and Inflammatory Stressors following Prenatal Maternal Infection. Genes 2022, 14, 77. [Google Scholar] [CrossRef]
- Keever-Keigher, M.R.; Zhang, P.; Bolt, C.R.; Rymut, H.E.; Antonson, A.M.; Corbett, M.P.; Houser, A.K.; Hernandez, A.G.; Southey, B.R.; Rund, L.A.; et al. Interacting impact of maternal inflammatory response and stress on the amygdala transcriptome of pigs. G3 2021, 11, jkab113. [Google Scholar] [CrossRef]
- Rymut, H.E.; Rund, L.A.; Bolt, C.R.; Villamil, M.B.; Bender, D.E.; Southey, B.R.; Johnson, R.W.; Rodriguez-Zas, S.L. Biochemistry and Immune Biomarkers Indicate Interacting Effects of Pre- and Postnatal Stressors in Pigs across Sexes. Animals 2021, 11, 987. [Google Scholar] [CrossRef] [PubMed]
- Andrews, S. A Quality Control Tool for High Throughput Sequence Data. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 16 November 2022).
- Pruitt, K.D.; Tatusova, T.; Maglott, D.R. NCBI reference sequences (RefSeq): A curated non-redundant sequence database of genomes, transcripts and proteins. Nucleic Acids Res. 2007, 35, D61–D65. [Google Scholar] [CrossRef] [PubMed]
- Bray, N.L.; Pimentel, H.; Melsted, P.; Pachter, L. Near-optimal probabilistic RNA-seq quantification. Nat. Biotechnol. 2016, 34, 525–527. [Google Scholar] [CrossRef]
- Zhang, P.; Moye, L.S.; Southey, B.R.; Dripps, I.; Sweedler, J.V.; Pradhan, A.; Rodriguez-Zas, S.L. Opioid-Induced Hyperalgesia Is Associated with Dysregulation of Circadian Rhythm and Adaptive Immune Pathways in the Mouse Trigeminal Ganglia and Nucleus Accumbens. Mol. Neurobiol. 2019, 56, 7929–7949. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed]
- Barrett, T.; Wilhite, S.E.; Ledoux, P.; Evangelista, C.; Kim, I.F.; Tomashevsky, M.; Marshall, K.A.; Phillippy, K.H.; Sherman, P.M.; Holko, M.; et al. NCBI GEO: Archive for functional genomics data sets—Update. Nucleic Acids Res. 2013, 41, D991–D995. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Kanehisa, M. Toward understanding the origin and evolution of cellular organisms. Protein Sci. 2019, 28, 1947–1951. [Google Scholar] [CrossRef] [PubMed]
- Gene Ontology Consortium. The Gene Ontology Resource: 20 years and still GOing strong. Nucleic Acids Res. 2019, 47, D330–D338. [Google Scholar] [CrossRef]
- Liao, Y.; Wang, J.; Jaehnig, E.J.; Shi, Z.; Zhang, B. WebGestalt 2019: Gene set analysis toolkit with revamped UIs and APIs. Nucleic Acids Res. 2019, 47, W199–W205. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P.; et al. The STRING database in 2021: Customizable protein-protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2021, 49, D605–D612. [Google Scholar] [CrossRef]
- Keever, M.R.; Zhang, P.; Bolt, C.R.; Antonson, A.M.; Rymut, H.E.; Caputo, M.P.; Houser, A.K.; Hernandez, A.G.; Southey, B.R.; Rund, L.A. Lasting and sex-dependent impact of maternal immune activation on molecular pathways of the amygdala. Front. Neurosci. 2020, 14, 774. [Google Scholar] [CrossRef]
- Rymut, H.E.; Rund, L.A.; Bolt, C.R.; Villamil, M.B.; Southey, B.R.; Johnson, R.W.; Rodriguez-Zas, S.L. The Combined Effect of Weaning Stress and Immune Activation during Pig Gestation on Serum Cytokine and Analyte Concentrations. Animals 2021, 11, 2274. [Google Scholar] [CrossRef]
- Makinodan, M.; Tatsumi, K.; Manabe, T.; Yamauchi, T.; Makinodan, E.; Matsuyoshi, H.; Shimoda, S.; Noriyama, Y.; Kishimoto, T.; Wanaka, A. Maternal immune activation in mice delays myelination and axonal development in the hippocampus of the offspring. J. Neurosci. Res. 2008, 86, 2190–2200. [Google Scholar] [CrossRef] [PubMed]
- Mabuchi, Y.; Shirasawa, N.; Sakuma, E.; Wada, I.; Horiuchi, O.; Kikuchi, M.; Sakamoto, A.; Herbert, D.C.; Soji, T. Electron microscopic observations of the anterior pituitary gland: Part I. The neurons in the “transitional zone” of the anterior pituitary gland. Tissue Cell 2008, 40, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Timmons, M.; Tsokos, M.; Asab, M.A.; Seminara, S.B.; Zirzow, G.C.; Kaneski, C.R.; Heiss, J.D.; van der Knaap, M.S.; Vanier, M.T.; Schiffmann, R.; et al. Peripheral and central hypomyelination with hypogonadotropic hypogonadism and hypodontia. Neurology 2006, 67, 2066–2069. [Google Scholar] [CrossRef]
- Page, N.F.; Gandal, M.J.; Estes, M.L.; Cameron, S.; Buth, J.; Parhami, S.; Ramaswami, G.; Murray, K.; Amaral, D.G.; Van de Water, J.A.; et al. Alterations in Retrotransposition, Synaptic Connectivity, and Myelination Implicated by Transcriptomic Changes Following Maternal Immune Activation in Nonhuman Primates. Biol. Psychiatry 2021, 89, 896–910. [Google Scholar] [CrossRef]
- Kikusui, T.; Kiyokawa, Y.; Mori, Y. Deprivation of mother–pup interaction by early weaning alters myelin formation in male, but not female, ICR mice. Brain Res. 2007, 1133, 115–122. [Google Scholar] [CrossRef]
- Lee, G.A.; Lin, Y.-K.; Lai, J.-H.; Lo, Y.-C.; Yang, Y.-C.S.; Ye, S.-Y.; Lee, C.-J.; Wang, C.-C.; Chiang, Y.-H.; Tseng, S.-H. Maternal immune activation causes social behavior deficits and hypomyelination in male rat offspring with an autism-like microbiota profile. Brain Sci. 2021, 11, 1085. [Google Scholar] [CrossRef]
- Richetto, J.; Chesters, R.; Cattaneo, A.; Labouesse, M.A.; Gutierrez, A.M.C.; Wood, T.C.; Luoni, A.; Meyer, U.; Vernon, A.; Riva, M.A. Genome-Wide Transcriptional Profiling and Structural Magnetic Resonance Imaging in the Maternal Immune Activation Model of Neurodevelopmental Disorders. Cereb. Cortex 2017, 27, 3397–3413. [Google Scholar] [CrossRef]
- Zucchi, F.C.R.; Yao, Y.; Ilnytskyy, Y.; Robbins, J.C.; Soltanpour, N.; Kovalchuk, I.; Kovalchuk, O.; Metz, G.A.S. Lifetime Stress Cumulatively Programs Brain Transcriptome and Impedes Stroke Recovery: Benefit of Sensory Stimulation. PLoS ONE 2014, 9, e92130. [Google Scholar] [CrossRef] [PubMed]
- Escorcio-Bezerra, M.L.; Pinto, W.B.V.R.; Bichuetti, D.B.; Souza, P.V.S.; Nunes, R.M.; Silva, L.H.L.; Lima, K.D.F.; Manzano, G.M.; Oliveira, A.S.B.; Baeta, A.M. Immune-mediated inflammatory polyneuropathy overlapping Charcot-Marie-Tooth 1B. J. Clin. Neurosci. 2020, 75, 228–231. [Google Scholar] [CrossRef] [PubMed]
- Leyrolle, Q.; Decoeur, F.; Briere, G.; Amadieu, C.; Quadros, A.R.A.A.; Voytyuk, I.; Lacabanne, C.; Benmamar-Badel, A.; Bourel, J.; Aubert, A.; et al. Maternal dietary omega-3 deficiency worsens the deleterious effects of prenatal inflammation on the gut-brain axis in the offspring across lifetime. Neuropsychopharmacology 2021, 46, 579–602. [Google Scholar] [CrossRef]
- Sunman, B.; Yurdakök, K.; Kocer-Gumusel, B.; Özyüncü, Ö.; Akbıyık, F.; Balcı, A.; Özkemahlı, G.; Erkekoğlu, P.; Yurdakök, M. Prenatal bisphenol a and phthalate exposure are risk factors for male reproductive system development and cord blood sex hormone levels. Reprod. Toxicol. 2019, 87, 146–155. [Google Scholar] [CrossRef]
- Nandwana, V.; Nandwana, N.K.; Das, Y.; Saito, M.; Panda, T.; Das, S.; Almaguel, F.; Hosmane, N.S.; Das, B.C. The Role of Microbiome in Brain Development and Neurodegenerative Diseases. Molecules 2022, 27, 3402. [Google Scholar] [CrossRef]
- Reshetnikov, V.V.; Kisaretova, P.E.; Bondar, N.P. Transcriptome Alterations Caused by Social Defeat Stress of Various Durations in Mice and Its Relevance to Depression and Posttraumatic Stress Disorder in Humans: A Meta-Analysis. Int. J. Mol. Sci. 2022, 23, 13792. [Google Scholar] [CrossRef]
- Roshan-Milani, S.; Seyyedabadi, B.; Saboory, E.; Parsamanesh, N.; Mehranfard, N. Prenatal stress and increased susceptibility to anxiety-like behaviors: Role of neuroinflammation and balance between GABAergic and glutamatergic transmission. Stress 2021, 24, 481–495. [Google Scholar] [CrossRef]
- Rymut, H.E.; Rund, L.A.; Southey, B.R.; Johnson, R.W.; Rodriguez-Zas, S.L. Terpenoid Backbone Biosynthesis among Pig Hippocampal Pathways Impacted by Stressors. Genes 2022, 13, 814. [Google Scholar] [CrossRef]
- Gorlova, A.V.; Pavlov, D.A.; Ushakova, V.M.; Zubkov, E.A.; Zorkina, Y.A.; Morozova, A.Y.; Inozemtsev, A.N.; Chekhonin, V.P. The Induction of a Depression-Like State by Chronic Exposure to Ultrasound in Rats Is Accompanied by a Reduction in Gene Expression of GABAA-Receptor Subunits in the Brain. Neurochem. J. 2020, 14, 49–54. [Google Scholar] [CrossRef]
- Mozhui, K.; Karlsson, R.-M.; Kash, T.L.; Ihne, J.; Norcross, M.; Patel, S.; Farrell, M.R.; Hill, E.E.; Graybeal, C.; Martin, K.P.; et al. Strain Differences in Stress Responsivity Are Associated with Divergent Amygdala Gene Expression and Glutamate-Mediated Neuronal Excitability. J. Neurosci. 2010, 30, 5357–5367. [Google Scholar] [CrossRef]
- Rymut, H.E.; Rund, L.A.; Southey, B.R.; Johnson, R.W.; Sweedler, J.V.; Rodriguez-Zas, S.L. Prefrontal Cortex Response to Prenatal Insult and Postnatal Opioid Exposure. Genes 2022, 13, 1371. [Google Scholar] [CrossRef] [PubMed]
- Dorshkind, K.; Horseman, N.D. Anterior pituitary hormones, stress, and immune system homeostasis. BioEssays 2001, 23, 288–294. [Google Scholar] [CrossRef]
- Anderson, A.; Genaro-Mattos, T.C.; Allen, L.B.; Koczok, K.; Korade, Z.; Mirnics, K. Interaction of maternal immune activation and genetic interneuronal inhibition. Brain Res. 2021, 1759, 147370. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.; Fang, M.; Xu, H.; Xing, H.; Nie, Q. Transcriptome comparison in the pituitary–adrenal axis between Beagle and Chinese Field dogs after chronic stress exposure. Anim. Genet. 2015, 46, 522–534. [Google Scholar] [CrossRef] [PubMed]
- Konishi, H.; Ogawa, T.; Nakagomi, S.; Inoue, K.; Tohyama, M.; Kiyama, H. Id1, Id2 and Id3 are induced in rat melanotrophs of the pituitary gland by dopamine suppression under continuous stress. Neuroscience 2010, 169, 1527–1534. [Google Scholar] [CrossRef] [PubMed]
- Narayan, S.; Head, S.R.; Gilmartin, T.J.; Dean, B.; Thomas, E.A. Evidence for disruption of sphingolipid metabolism in schizophrenia. J. Neurosci. Res. 2009, 87, 278–288. [Google Scholar] [CrossRef] [PubMed]
- Gebicke-Haerter, P.J.; Leonardi-Essmann, F.; Haerter, J.O.; Rossner, M.J.; Falkai, P.; Schmitt, A.; Raabe, F.J. Differential gene regulation in the anterior cingulate cortex and superior temporal cortex in schizophrenia: A molecular network approach. Schizophr. Res. 2021, 232, 1–10. [Google Scholar] [CrossRef]




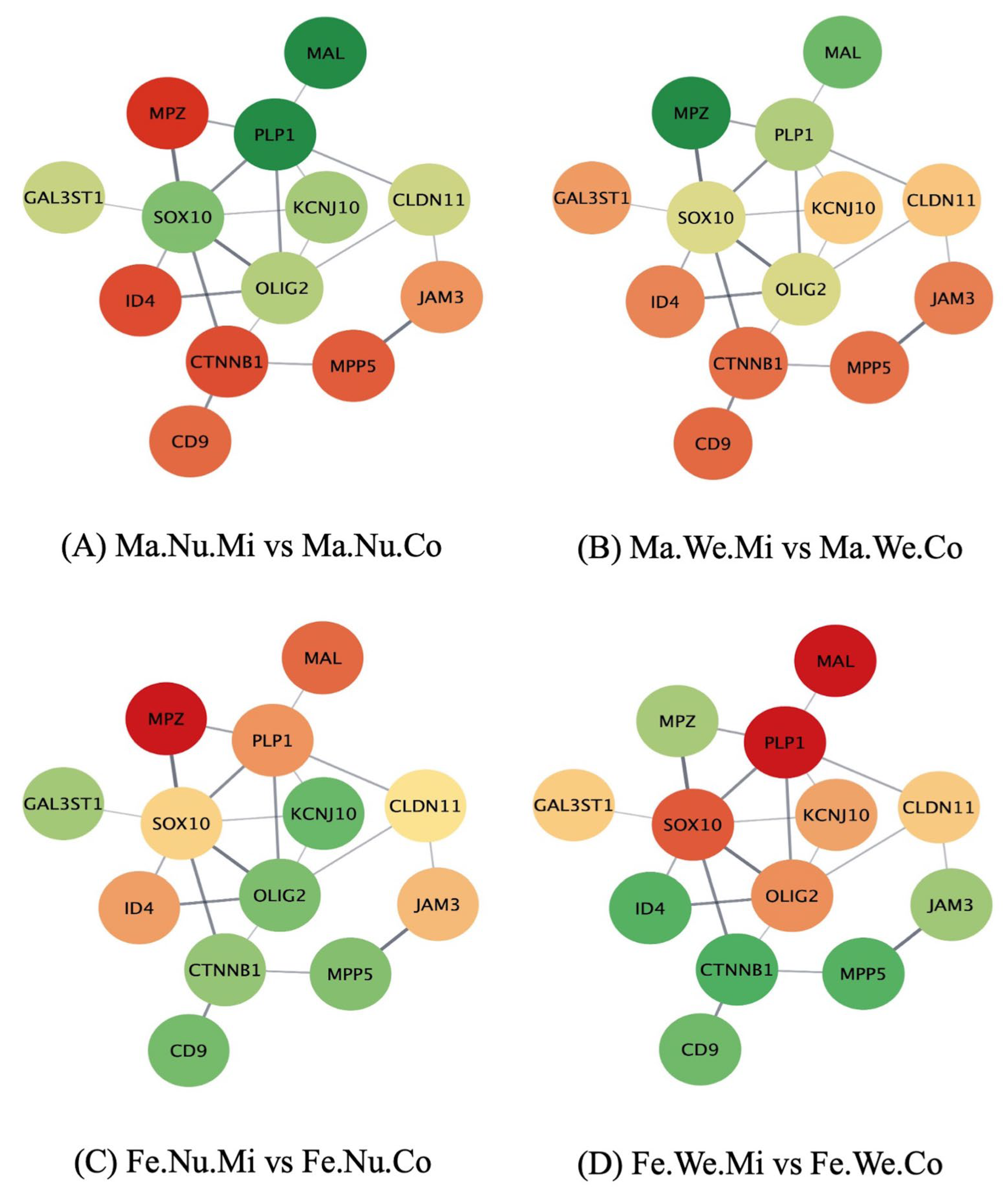{kind=link}
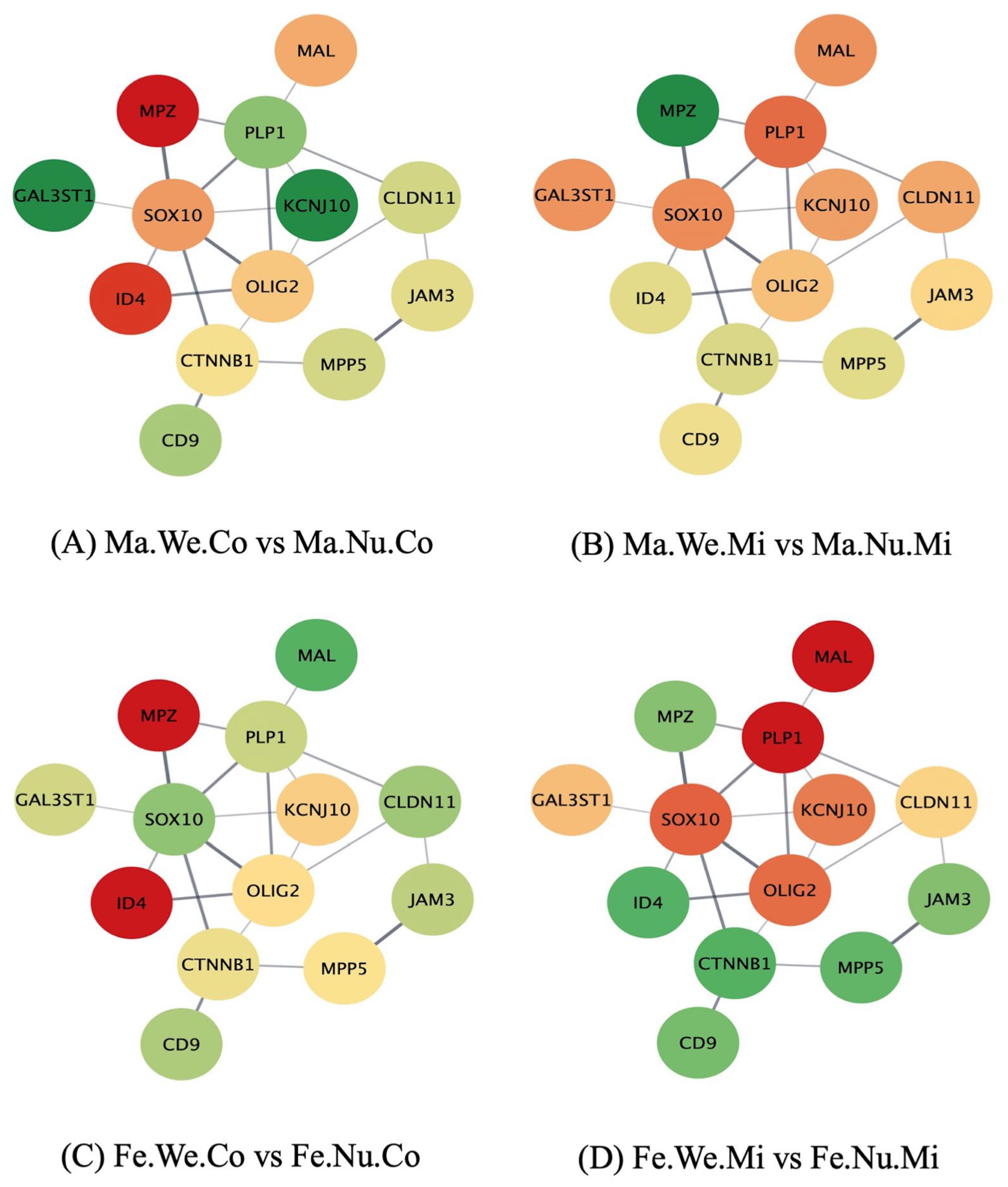{kind=link}
{kind=link}
| Category | Identifier | MIA | Weaning | Sex | MIA × Sex | Weaning × Sex | Weaning × MIA | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NES 3 | FDR | NES | FDR | NES | FDR | NES | FDR | NES | FDR | NES | FDR | ||
| GO Biological Process 1 | |||||||||||||
| Nervous system | |||||||||||||
| Ensheathment of neurons | GO:0007272 | 1.6 | 4 × 10−2 | −1.8 | 2 × 10−8 | 1.8 | 5 × 10−3 | −2.2 | 2 × 10−8 | ||||
| Regeneration | GO:0031099 | 1.6 | 4 × 10−2 | −1.7 | 1 × 10−2 | 1.7 | 3 × 10−2 | ||||||
| Neuropeptide signaling pathway | GO:0007218 | 1.8 | 2 × 10−2 | −1.9 | 3 × 10−3 | 1.9 | 5 × 10−3 | 1.8 | 2 × 10−2 | ||||
| Extracellular structure organization | GO:0043062 | 2.0 | 2 × 10−8 | 1.7 | 3 × 10−2 | ||||||||
| Reproductive system development | GO:0061458 | 1.8 | 2 × 10−2 | 1.7 | 4 × 10−2 | ||||||||
| Morphogenesis of a branching structure | GO:0001763 | 1.8 | 2 × 10−2 | −2.0 | 1 × 10−2 | ||||||||
| Pattern specification process | GO:0007389 | 1.7 | 3 × 10−2 | 1.9 | 2 × 10−2 | ||||||||
| Response to organophosphorus | GO:0046683 | 1.6 | 4 × 10−2 | 1.9 | 3 × 10−2 | ||||||||
| KEGG Pathways 2 | |||||||||||||
| Nervous system | |||||||||||||
| Nicotine addiction | ssc05033 | −2.1 | 2 × 10−8 | −1.8 | 9 × 10−3 | 1.9 | 3 × 10−3 | ||||||
| Morphine addiction | ssc05032 | −1.8 | 5 × 10−3 | 1.7 | 3 × 10−2 | ||||||||
| GABAergic synapse | ssc04727 | −1.8 | 5 × 10−3 | 1.7 | 3 × 10−2 | ||||||||
| Oxidative phosphorylation | ssc00190 | −1.8 | 1 × 10−2 | 1.9 | 2 × 10−3 | −1.9 | 2 × 10−2 | ||||||
| Protein digestion and absorption | ssc04974 | 1.9 | 2 × 10−3 | −1.7 | 2 × 10−2 | 2.0 | 2 × 10−3 | ||||||
| Ribosome | ssc03010 | −2.2 | 2 × 10−8 | 1.9 | 3 × 10−2 | 2.0 | 2 × 10−8 | −2.3 | 2 × 10−8 | ||||
| Long-term potentiation | ssc04720 | −1.7 | 4 × 10−2 | −1.7 | 4 × 10−2 | −1.8 | 3 × 10−2 | ||||||
| Immune system | |||||||||||||
| Graft-versus-host disease | ssc05332 | −2.0 | 9 × 10−4 | 2.3 | 2 × 10−8 | ||||||||
| Measles | ssc05162 | −1.8 | 2 × 10−2 | 1.7 | 2 × 10−2 | ||||||||
| Antigen processing and presentation | ssc04612 | −1.8 | 2 × 10−2 | 2.4 | 2 × 10−8 | ||||||||
| Th17 cell differentiation | ssc04659 | −1.7 | 3 × 10−2 | 1.9 | 3 × 10−3 | ||||||||
| Gene | MIA | Weaning | Sex | MIA × Sex | Weaning × Sex | Weaning × MIA | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Symbol | LogFC 1 | FDR | LogFC | FDR | LogFC | FDR | LogFC | FDR | LogFC | FDR | LogFC | FDR |
| Neuronal ensheathment | ||||||||||||
| Cldn11 | 0.7 | 4 × 10−1 | 0.1 | 1 × 100 | −3.7 | 3 × 10−5 | 4.7 | 3 × 10−5 | −0.3 | 1 × 100 | −3.0 | 3 × 10−5 |
| Olig2 | 0.3 | 1 × 100 | −0.2 | 1 × 100 | −4.7 | 3 × 10−5 | 4.7 | 3 × 10−5 | −0.3 | 1 × 100 | −4.6 | 1 × 10−5 |
| Mal | 1.2 | 1 × 10−2 | 0.2 | 1 × 100 | −7.0 | 4 × 10−5 | 8.8 | 2 × 10−5 | −0.8 | 1 × 100 | −6.8 | 1 × 10−5 |
| Plp1 | 1.1 | 5 × 10−2 | 0.0 | 1 × 100 | −7.8 | 5 × 10−5 | 8.7 | 4 × 10−5 | 0.1 | 1 × 100 | −6.3 | 2 × 10−5 |
| Immune system | ||||||||||||
| Cxcl10 | −0.2 | 1 × 100 | −0.8 | 3 × 10−1 | −1.3 | 2 × 10−3 | 4.4 | 6 × 10−8 | 1.4 | 2 × 10−1 | 2.2 | 3 × 10−4 |
| Cxcl9 | −2.2 | 3 × 10−7 | −0.5 | 1 × 100 | −0.1 | 1 × 100 | 1.7 | 3 × 10−2 | 0.0 | 1 × 100 | 2.4 | 4 × 10−4 |
| Tnfrsf8 | 1.9 | 2 × 10−5 | −1.9 | 1 × 10−5 | −2.1 | 6 × 10−8 | 2.4 | 5 × 10−5 | 2.3 | 5 × 10−4 | 1.9 | 9 × 10−3 |
| Gdf1 | 1.2 | 1 × 10−2 | −0.3 | 1 × 100 | −3.3 | 5 × 10−8 | 4.3 | 6 × 10−8 | 0.7 | 1 × 100 | −2.1 | 5 × 10−4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alsegehy, S.; Southey, B.R.; Rund, L.; Johnson, R.W.; Rodriguez-Zas, S.L. Genes Participating in the Ensheathment of Neurons Are Affected by Postnatal Stress and Maternal Immune Activation in the Pituitary Gland. Genes 2023, 14, 1007. https://doi.org/10.3390/genes14051007
Alsegehy S, Southey BR, Rund L, Johnson RW, Rodriguez-Zas SL. Genes Participating in the Ensheathment of Neurons Are Affected by Postnatal Stress and Maternal Immune Activation in the Pituitary Gland. Genes. 2023; 14(5):1007. https://doi.org/10.3390/genes14051007
Chicago/Turabian StyleAlsegehy, Samah, Bruce R. Southey, Laurie Rund, Rodney W. Johnson, and Sandra L. Rodriguez-Zas. 2023. "Genes Participating in the Ensheathment of Neurons Are Affected by Postnatal Stress and Maternal Immune Activation in the Pituitary Gland" Genes 14, no. 5: 1007. https://doi.org/10.3390/genes14051007