
Intrafollicular Retinoic Acid Signaling Is Important for Luteinizing Hormone-Induced Oocyte Meiotic Resumption
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Animals
2.2. Culture of Preovulatory Follicles
2.3. Culture of COCs
2.4. Culture of Mouse MGCs
2.5. RNA Extraction and Quantitative RT-PCR Analysis (qRT-PCR)
2.6. Western Blotting
2.7. GSEA Analysis
2.8. Cleavage under Targets & Release Using Nuclease (CUT&RUN)
2.9. Reanalysis of Published RNA-Sequencing Data
2.10. Statistical Analysis
3. Results
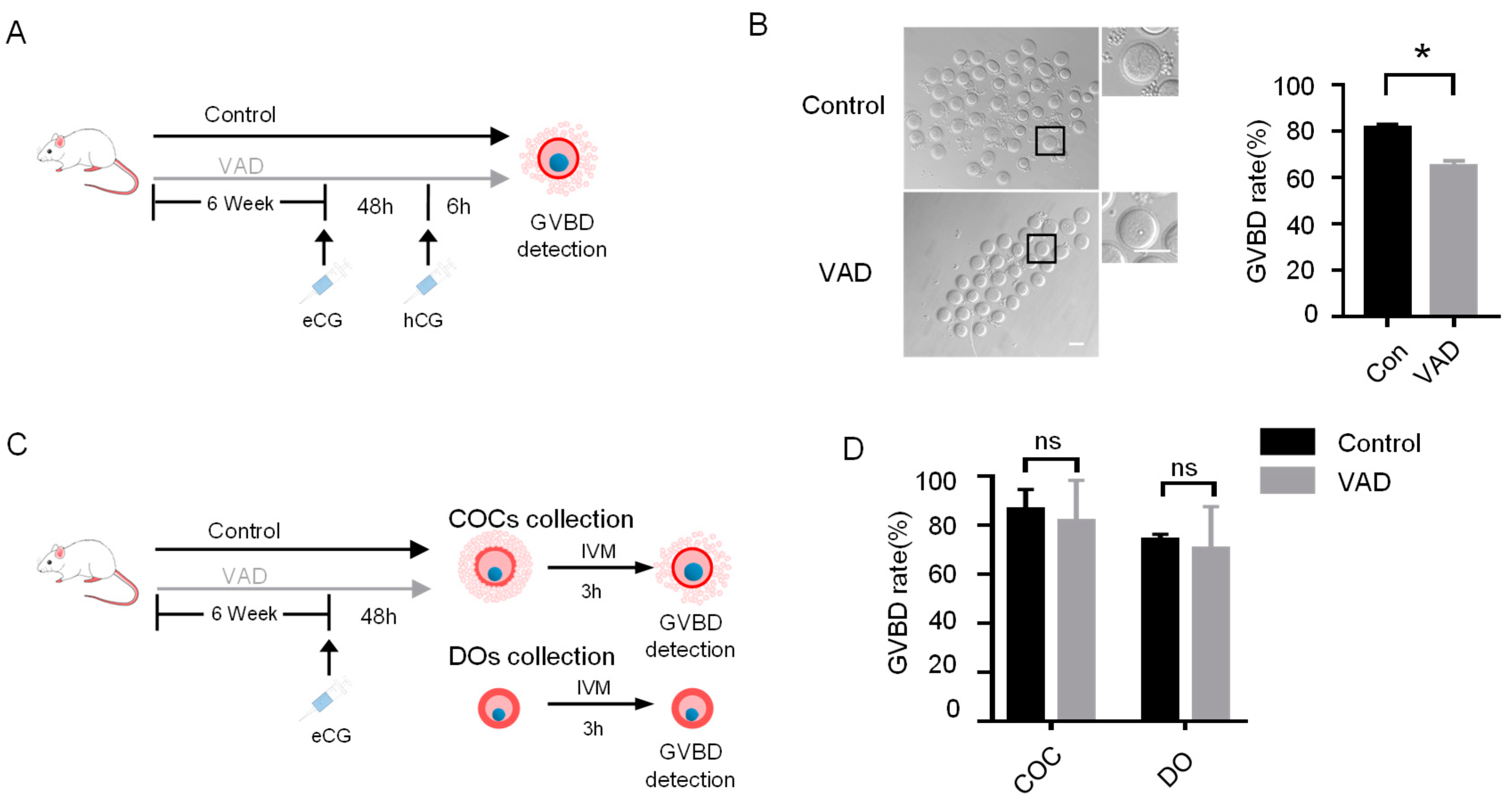
3.1. VA Deficiency Impairs Oocyte Meiotic Resumption
3.2. LH Activates RA Synthetic and Metabolic Pathway in Preovulatory MGCs
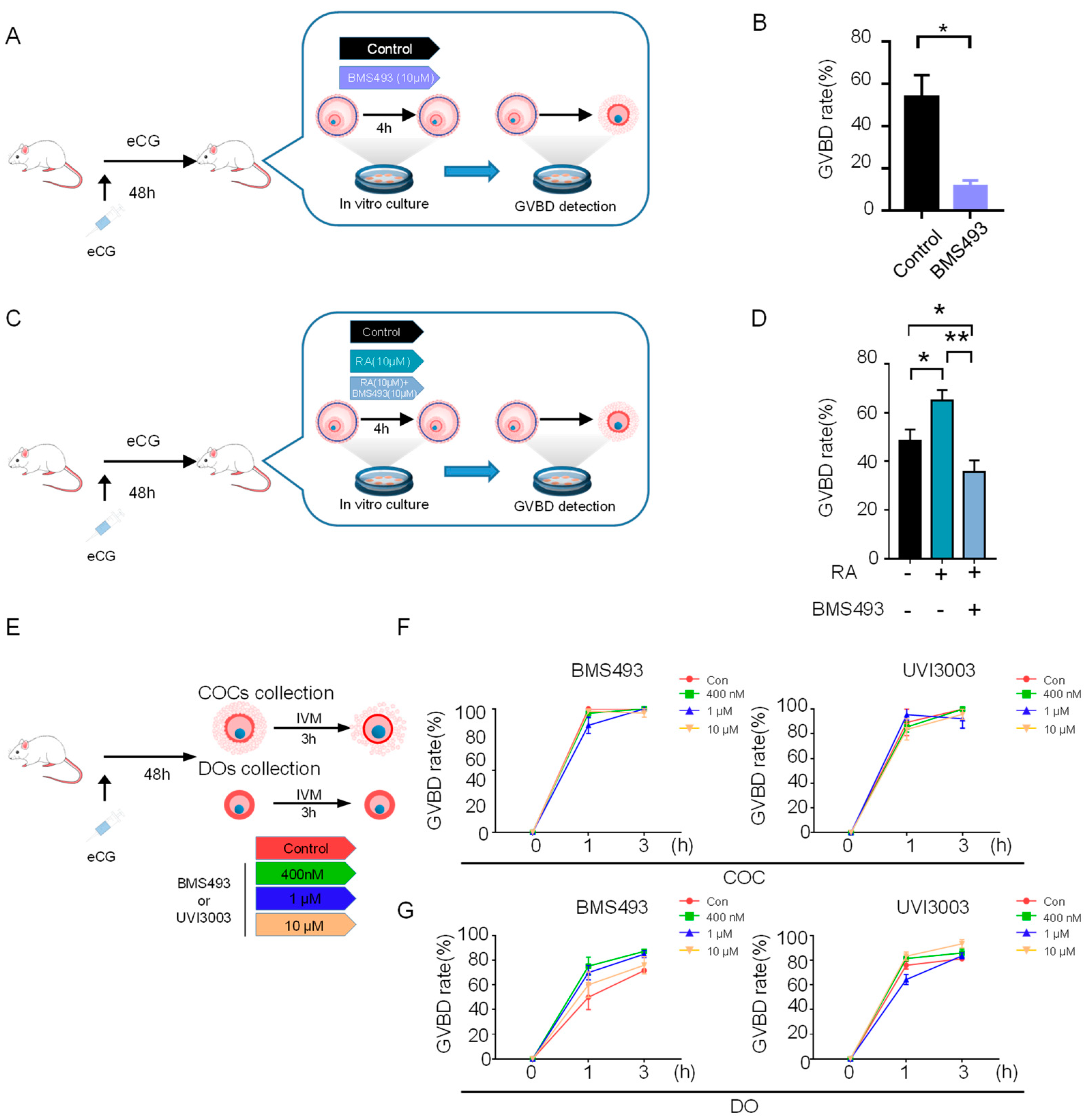
3.3. RA Signaling Disruption Impairs Oocyte Meiotic Resumption
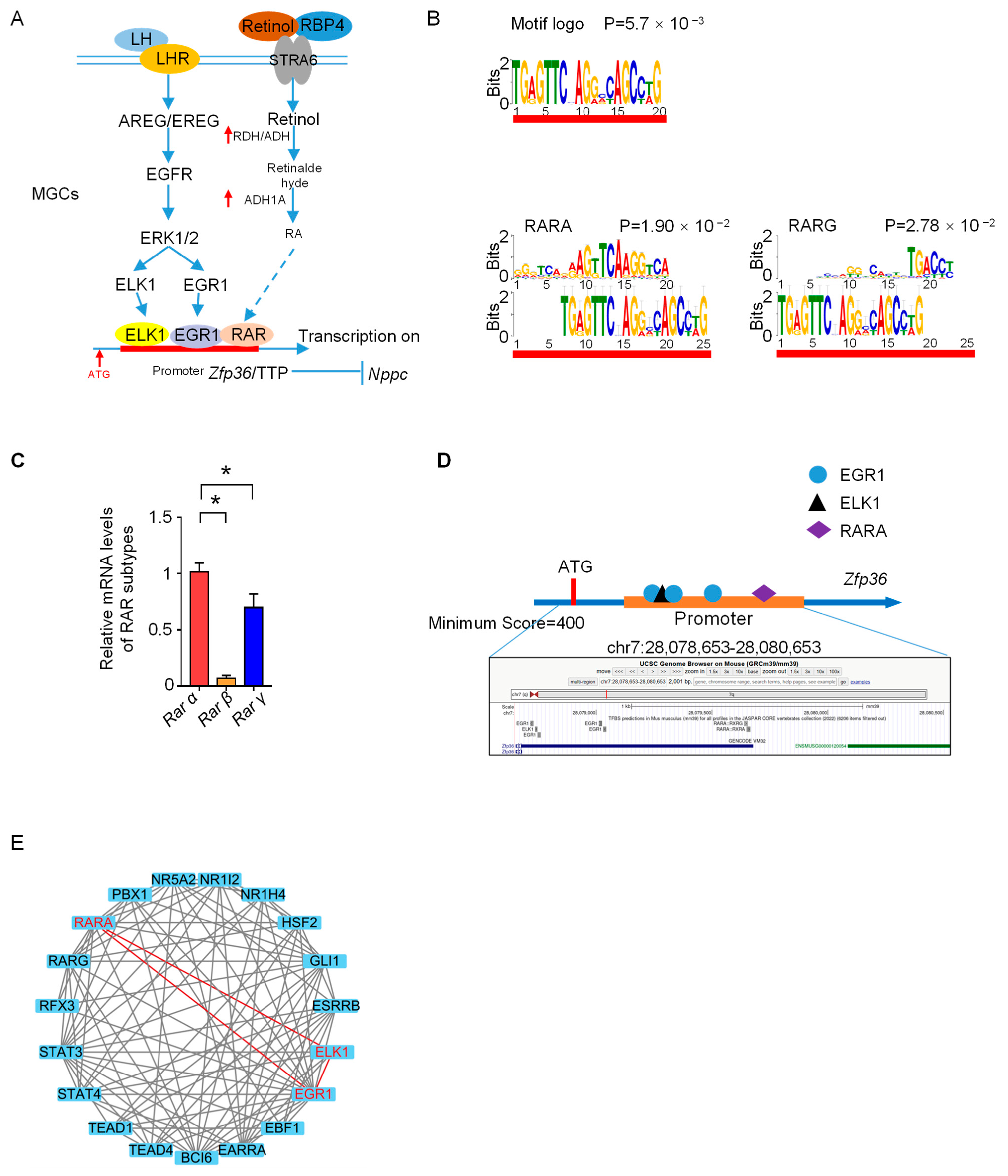
3.4. Zfp36 Is the Potential Transcriptional Target of RAR
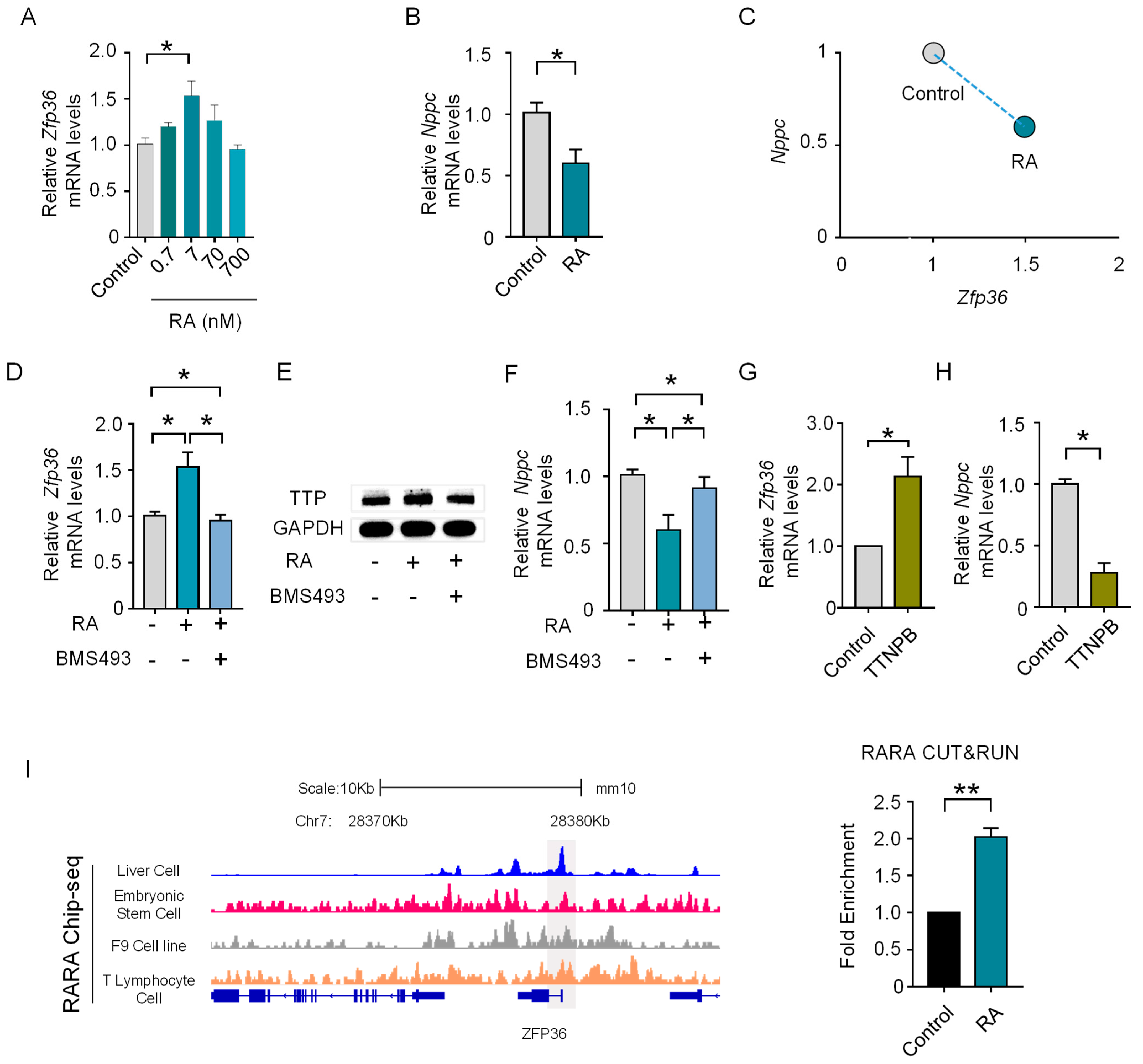
3.5. RA Prompts the Expression of Zfp36 in MGC via RAR
3.6. RA Signaling Cooperates with Parallel EGF Signaling to Upregulate Zfp36 through an ERK-Dependent Pathway
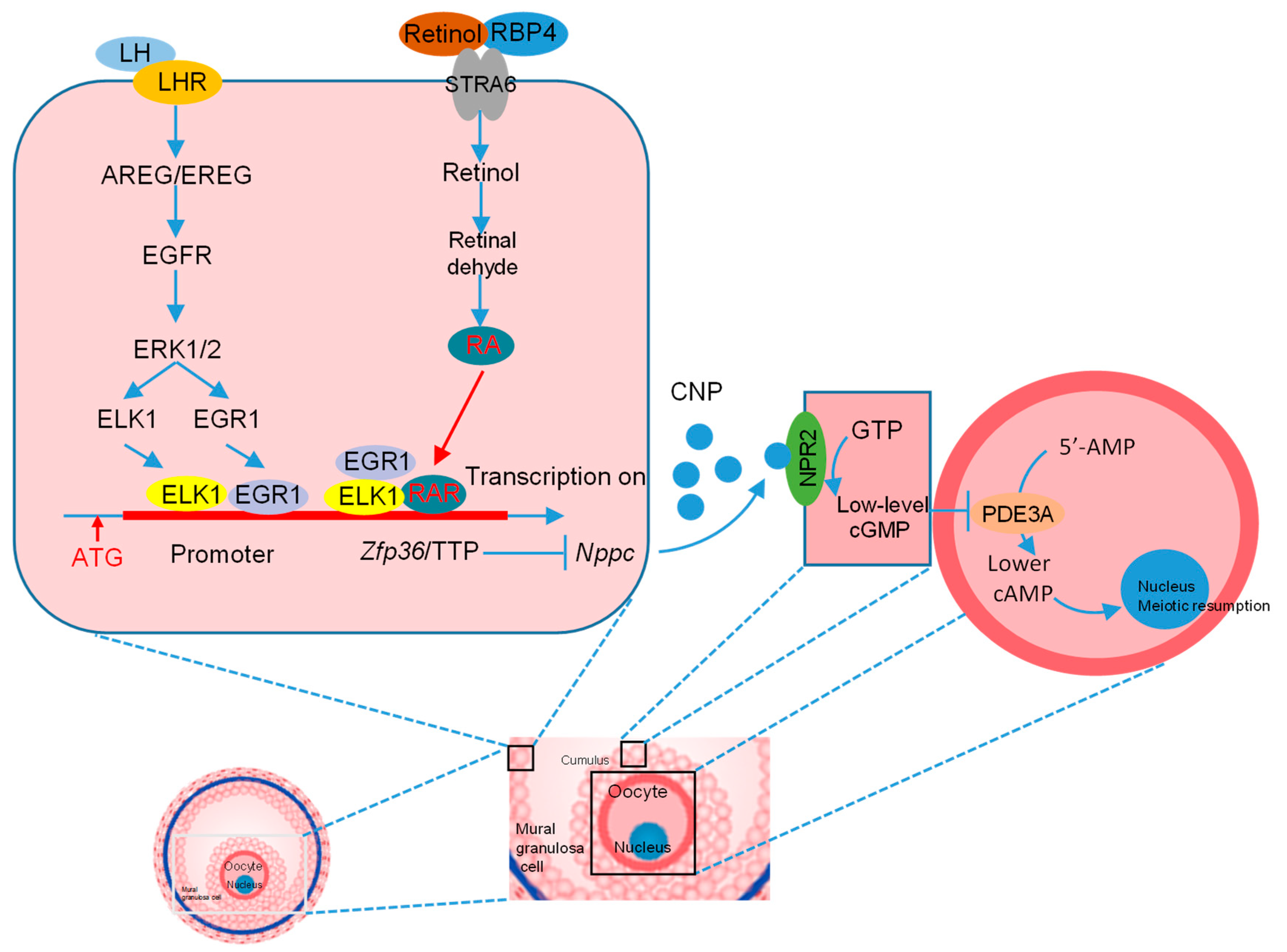
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Das, B.C.; Thapa, P.; Karki, R.; Das, S.; Mahapatra, S.; Liu, T.; Torregroza, I.; Wallace, D.P.; Kambhampati, S.; Van Veldhuizen, P.; et al. Retinoic acid signaling pathways in development and diseases. Bioorg. Med. Chem. 2014, 22, 673–683. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Yanaka, N.; Richards, J.S.; Shimada, M. De Novo-Synthesized Retinoic Acid in Ovarian Antral Follicles Enhances FSH-Mediated Ovarian Follicular Cell Differentiation and Female Fertility. Endocrinology 2016, 157, 2160–2172. [Google Scholar] [CrossRef] [PubMed]
- Kin Ting Kam, R.; Deng, Y.; Chen, Y.; Zhao, H. Retinoic acid synthesis and functions in early embryonic development. Cell Biosci. 2012, 2, 11. [Google Scholar] [CrossRef]
- Bowles, J.; Koopman, P. Retinoic acid, meiosis and germ cell fate in mammals. Development 2007, 134, 3401–3411. [Google Scholar] [CrossRef]
- Koubova, J.; Menke, D.B.; Zhou, Q.; Capel, B.; Griswold, M.D.; Page, D.C. Retinoic acid regulates sex-specific timing of meiotic initiation in mice. Proc. Natl. Acad. Sci. USA 2006, 103, 2474–2479. [Google Scholar] [CrossRef]
- Menke, D.B.; Koubova, J.; Page, D.C. Sexual differentiation of germ cells in XX mouse gonads occurs in an anterior-to-posterior wave. Dev. Biol. 2003, 262, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Abouzaripour, M.; Fathi, F.; Daneshi, E.; Mortezaee, K.; Rezaie, M.J.; Abdi, M. Combined Effect of Retinoic Acid and Basic Fibroblast Growth Factor on Maturation of Mouse Oocyte and Subsequent Fertilization and Development. Int. J. Fertil. Steril. 2018, 12, 68–71. [Google Scholar] [CrossRef]
- Ara Ahmed, J.; Dutta, D.; Nashiruddullah, N. Comparative efficacy of antioxidant retinol, melatonin, and zinc during in vitro maturation of bovine oocytes under induced heat stress. Turk. J. Vet. Anim. Sci. 2016, 40, 365–373. [Google Scholar] [CrossRef]
- Lilibeth, A.; Cajuday, A.A.H.D. Effect of Retinoic Acid on the Development of Water Buffalo Embryos In Vitro. Philipp. J. Vet. Anim. Sci. 2012, 2, 107–116. [Google Scholar]
- Tahaei, L.S.; Eimani, H.; Yazdi, P.E.; Ebrahimi, B.; Fathi, R. Effects of retinoic acid on maturation of immature mouse oocytes in the presence and absence of a granulosa cell co-culture system. J. Assist. Reprod. Genet. 2011, 28, 553–558. [Google Scholar] [CrossRef]
- Liang, S.; Kang, J.; Jin, H.; Liu, X.; Li, J.; Li, S.; Lu, Y.; Wang, W.; Yin, X. The influence of 9-cis-retinoic acid on nuclear and cytoplasmic maturation and gene expression in canine oocytes during in vitro maturation. Theriogenology 2012, 77, 1198–1205. [Google Scholar] [CrossRef] [PubMed]
- Vahedi, V.; Zeinoaldini, S.; Kohram, H.; Farahavar, A. Retinoic acid effects on nuclear maturation of bovine oocytes in vitro. Afr. J. Biotechnol. 2009, 8, 3974–3978. [Google Scholar]
- Richards, J.S.; Ascoli, M. Endocrine, Paracrine, and Autocrine Signaling Pathways That Regulate Ovulation. Trends Endocrinol. Metab. 2018, 29, 313–325. [Google Scholar] [CrossRef]
- Gómez, E.; Royo, L.; Duque, P.; Carneiro, G.; Hidalgo, C.; Goyache, F.; Lorenzo, P.L.; Alvarez, I.; Facal, N.; Díez, C. 9-cis-retinoic acid during in vitro maturation improves development of the bovine oocyte and increases midkine but not IGF-I expression in cumulus-granulosa cells. Mol. Reprod. Dev. 2003, 66, 247–255. [Google Scholar] [CrossRef]
- Almiñana, C.; Gil, M.A.; Cuello, C.; Caballero, I.; Roca, J.; Vazquez, J.M.; Gomez, E.; Martinez, E.A. In vitro maturation of porcine oocytes with retinoids improves embryonic development. Reprod. Fertil. Dev. 2008, 20, 483. [Google Scholar] [CrossRef]
- Zhang, M.; Su, Y.; Sugiura, K.; Xia, G.; Eppig, J.J. Granulosa Cell Ligand NPPC and Its Receptor NPR2 Maintain Meiotic Arrest in Mouse Oocytes. Science 2010, 330, 366–369. [Google Scholar] [CrossRef]
- Norris, R.P.; Ratzan, W.J.; Freudzon, M.; Mehlmann, L.M.; Krall, J.; Movsesian, M.A.; Wang, H.; Ke, H.; Nikolaev, V.O.; Jaffe, L.A. Cyclic GMP from the surrounding somatic cells regulates cyclic AMP and meiosis in the mouse oocyte. Development 2009, 136, 1869–1878. [Google Scholar] [CrossRef] [PubMed]
- Robinson, J.W.; Zhang, M.; Shuhaibar, L.C.; Norris, R.P.; Geerts, A.; Wunder, F.; Eppig, J.J.; Potter, L.R.; Jaffe, L.A. Luteinizing hormone reduces the activity of the NPR2 guanylyl cyclase in mouse ovarian follicles, contributing to the cyclic GMP decrease that promotes resumption of meiosis in oocytes. Dev. Biol. 2012, 366, 308–316. [Google Scholar] [CrossRef]
- Xi, G.; An, L.; Wang, W.; Hao, J.; Yang, Q.; Ma, L.; Lu, J.; Wang, Y.; Wang, W.; Zhao, W.; et al. The mRNA-destabilizing protein Tristetraprolin targets “meiosis arrester” Nppc mRNA in mammalian preovulatory follicles. Proc. Natl. Acad. Sci. USA 2021, 118, e2018345118. [Google Scholar] [CrossRef]
- Shirafuta, Y.; Tamura, I.; Ohkawa, Y.; Maekawa, R.; Doi-Tanaka, Y.; Takagi, H.; Mihara, Y.; Shinagawa, M.; Taketani, T.; Sato, S.; et al. Integrated Analysis of Transcriptome and Histone Modifications in Granulosa Cells During Ovulation in Female Mice. Endocrinology 2021, 162, bqab128. [Google Scholar] [CrossRef]
- Liu, X.; Xie, F.; Zamah, A.M.; Cao, B.; Conti, M. Multiple Pathways Mediate Luteinizing Hormone Regulation of cGMP Signaling in the Mouse Ovarian Follicle1. Biol. Reprod. 2014, 91, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Campos-Junior, P.H.; Silva, C.A.; Grazia, J.G.; Soares, M.B.; Santos, R.R.; Viana, J.H. Use of ultrasound biomicroscopy to evaluate induced ovarian follicular growth and ovulation in mice. Lab. Anim. 2011, 45, 254–258. [Google Scholar] [CrossRef]
- Crispo, M.; Meikle, M.N.; Schlapp, G.; Menchaca, A. Ovarian superstimulatory response and embryo development using a new recombinant glycoprotein with eCG-like activity in mice. Theriogenology 2021, 164, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Lima, P.F.; Oliveira, M.A.L.; Santos, M.H.B.; Reichenbach, H.D.; Weppert, M.; Paula-Lopes, F.F.; Neto, C.C.C.; Gonçalves, P.B.D. Effect of retinoids and growth factor on in vitro bovine embryos produced under chemically defined conditions. Anim. Reprod. Sci. 2006, 95, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Cheng, Y.; Kawamura, K.; Takae, S.; Hsueh, A.J.W. C-Type Natriuretic Peptide Stimulates Ovarian Follicle Development. Mol. Endocrinol. 2012, 26, 1158–1166. [Google Scholar] [CrossRef]
- Simona Marc Zarcula, G.G.C.M.; Tulcan, H.C.G.O.; Andreea Ciobota, I.H.C.M. Morphological aspects of cumulus –oocyte complexes in different species. In Proceedings of the 53th Annual Meeting of Veterinary Medicine—“Towards a Global Health”, Iasi, Romania, October 2014; pp. 91–97. [Google Scholar]
- Choi, H.; Lee, J.; Yoon, J.D.; Hwang, S.U.; Cai, L.; Kim, M.; Kim, G.; Oh, D.; Kim, E.; Hyun, S.H. The effect of copper supplementation on in vitro maturation of porcine cumulus-oocyte complexes and subsequent developmental competence after parthenogenetic activation. Theriogenology 2021, 164, 84–92. [Google Scholar] [CrossRef]
- Ming, T.; Nielsen, H.; Chen, Z. Maturation arrest of human oocytes at germinal vesicle stage. J. Hum. Reprod. Sci. 2010, 3, 153. [Google Scholar] [CrossRef]
- Tombes, R.M.; Simerly, C.; Borisy, G.G.; Schatten, G. Meiosis, egg activation, and nuclear envelope breakdown are differentially reliant on Ca2+, whereas germinal vesicle breakdown is Ca2+ independent in the mouse oocyte. J. Cell Biol. 1992, 117, 799–811. [Google Scholar] [CrossRef]
- Hainer, S.J.; Bošković, A.; Mccannell, K.N.; Rando, O.J.; Fazzio, T.G. Profiling of Pluripotency Factors in Single Cells and Early Embryos. Cell 2019, 177, 1319–1329. [Google Scholar] [CrossRef]
- Skene, P.J.; Henikoff, J.G.; Henikoff, S. Targeted in situ genome-wide profiling with high efficiency for low cell numbers. Nat. Protoc. 2018, 13, 1006–1019. [Google Scholar] [CrossRef]
- Hainer, S.J.; Fazzio, T.G. High-Resolution Chromatin Profiling Using CUT&RUN. Curr. Protoc. Mol. Biol. 2018, 126, e85. [Google Scholar] [CrossRef]
- White, J.C.; Highland, M.; Clagett-Dame, M. Abnormal development of the sinuatrial venous valve and posterior hindbrain may contribute to late fetal resorption of vitamin A-deficient rat embryos. Teratology 2000, 62, 374–384. [Google Scholar] [CrossRef]
- See, A.W.; Kaiser, M.E.; White, J.C.; Clagett-Dame, M. A nutritional model of late embryonic vitamin A deficiency produces defects in organogenesis at a high penetrance and reveals new roles for the vitamin in skeletal development. Dev. Biol. 2008, 316, 171–190. [Google Scholar] [CrossRef] [PubMed]
- Florkowska, M.; Tymoszuk, P.; Balwierz, A.; Skucha, A.; Kochan, J.; Wawro, M.; Stalinska, K.; Kasza, A. EGF activates TTP expression by activation of ELK-1 and EGR-1 transcription factors. BMC Mol. Biol. 2012, 13, 8. [Google Scholar] [CrossRef] [PubMed]
- Koubova, J.; Hu, Y.C.; Bhattacharyya, T.; Soh, Y.Q.; Gill, M.E.; Goodheart, M.L.; Hogarth, C.A.; Griswold, M.D.; Page, D.C. Retinoic acid activates two pathways required for meiosis in mice. PLoS Genet. 2014, 10, e1004541. [Google Scholar] [CrossRef] [PubMed]
- Nasiri, E.; Mahmoudi, R.; Bahadori, M.H.; Amiri, I. The Effect of Retinoic Acid on in vitro Maturation and Fertilization Rate of Mouse Germinal Vesicle Stage Oocytes. Cell J. 2011, 13, 19–24. [Google Scholar]
- Whaley, S.L.; Hedgpeth, V.S.; Farin, C.E.; Martus, N.S.; Jayes, F.C.; Britt, J.H. Influence of vitamin A injection before mating on oocyte development, follicular hormones, and ovulation in gilts fed high-energy diets. J. Anim. Sci. 2000, 78, 1598–1607. [Google Scholar] [CrossRef]
- Jaffe, L.A.; Egbert, J.R. Regulation of Mammalian Oocyte Meiosis by Intercellular Communication Within the Ovarian Follicle. Annu. Rev. Physiol. 2017, 79, 237–260. [Google Scholar] [CrossRef]
- He, M.; Zhang, T.; Yang, Y.; Wang, C. Mechanisms of Oocyte Maturation and Related Epigenetic Regulation. Front. Cell. Dev. Biol. 2021, 9, 654028. [Google Scholar] [CrossRef]
- Egbert, J.R.; Yee, S.; Jaffe, L.A. Luteinizing hormone signaling phosphorylates and activates the cyclic GMP phosphodiesterase PDE5 in mouse ovarian follicles, contributing an adsditional component to the hormonally induced decrease in cyclic GMP that reinitiates meiosis. Dev. Biol. 2018, 435, 6–14. [Google Scholar] [CrossRef]
- Hsieh, M.; Conti, M. G-protein-coupled receptor signaling and the EGF network in endocrine systems. Trends Endocrinol. Metab. 2005, 16, 320–326. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, F.; Tang, Y.; Cai, Y.; Yang, R.; Wang, Z.; Wang, X.; Yang, Q.; Wang, W.; Tian, J.; An, L. Intrafollicular Retinoic Acid Signaling Is Important for Luteinizing Hormone-Induced Oocyte Meiotic Resumption. Genes 2023, 14, 946. https://doi.org/10.3390/genes14040946
Wang F, Tang Y, Cai Y, Yang R, Wang Z, Wang X, Yang Q, Wang W, Tian J, An L. Intrafollicular Retinoic Acid Signaling Is Important for Luteinizing Hormone-Induced Oocyte Meiotic Resumption. Genes. 2023; 14(4):946. https://doi.org/10.3390/genes14040946
Chicago/Turabian StyleWang, Fupeng, Yawen Tang, Yijie Cai, Ran Yang, Zongyu Wang, Xiaodong Wang, Qianying Yang, Wenjing Wang, Jianhui Tian, and Lei An. 2023. "Intrafollicular Retinoic Acid Signaling Is Important for Luteinizing Hormone-Induced Oocyte Meiotic Resumption" Genes 14, no. 4: 946. https://doi.org/10.3390/genes14040946