
The Genetics of Fitness Reorganization during the Transition to Multicellularity: The Volvocine regA-like Family as a Model


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
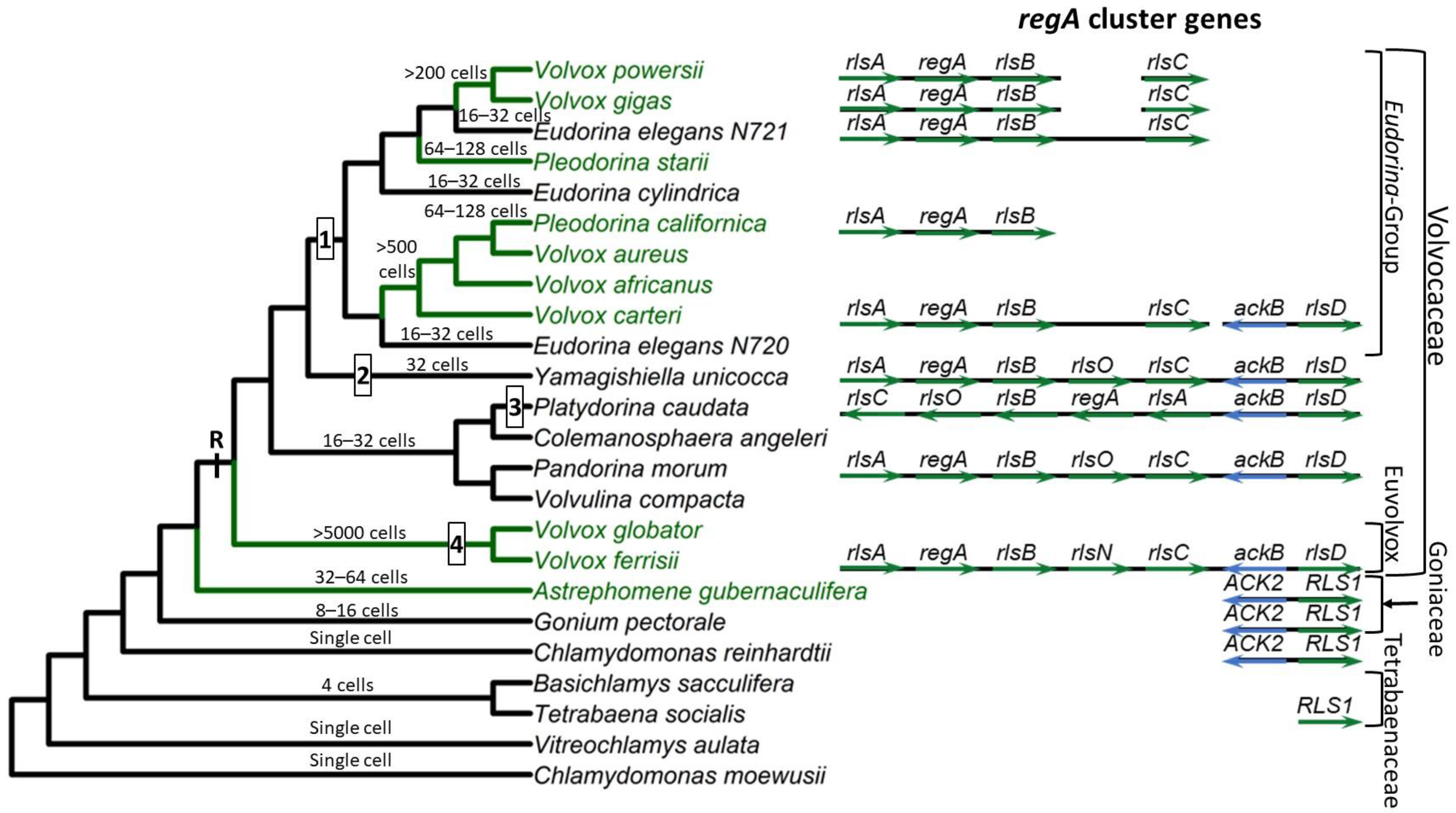
2. The Volvocine Model System
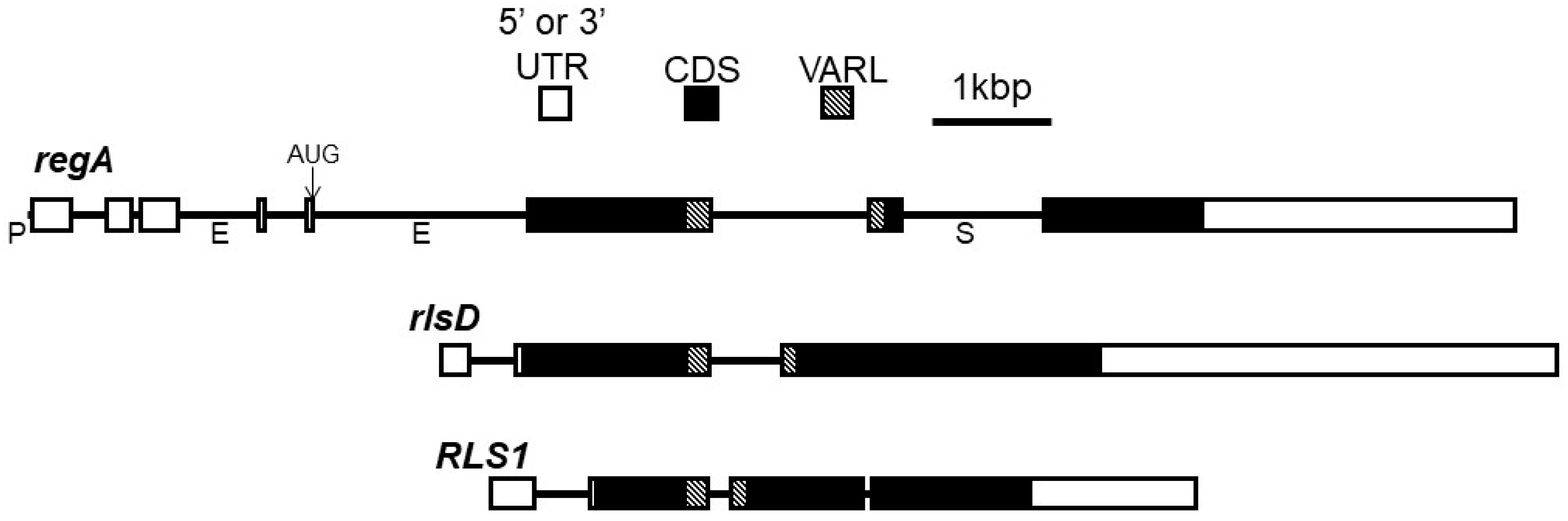
3. regA Gene Structure and Function
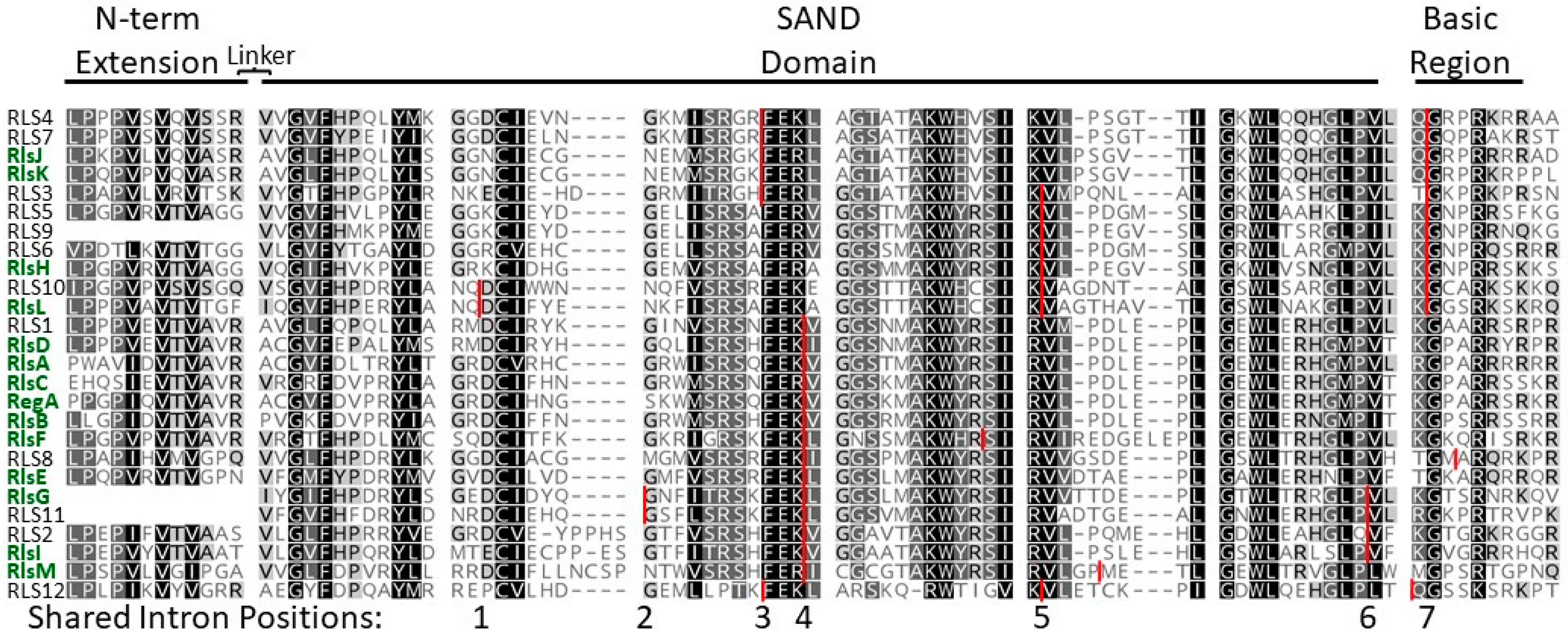
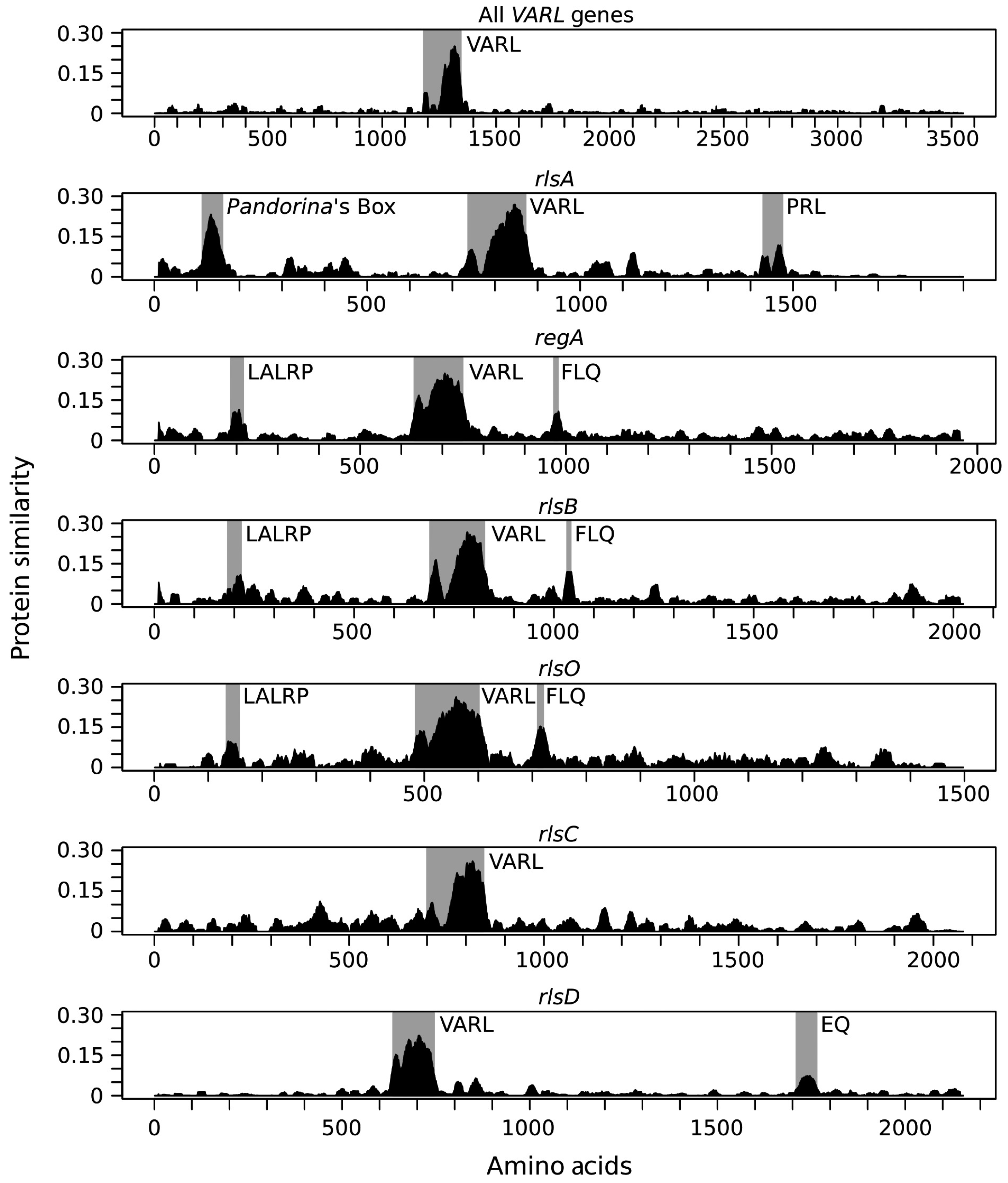
4. Evolution of the VARL Gene Family
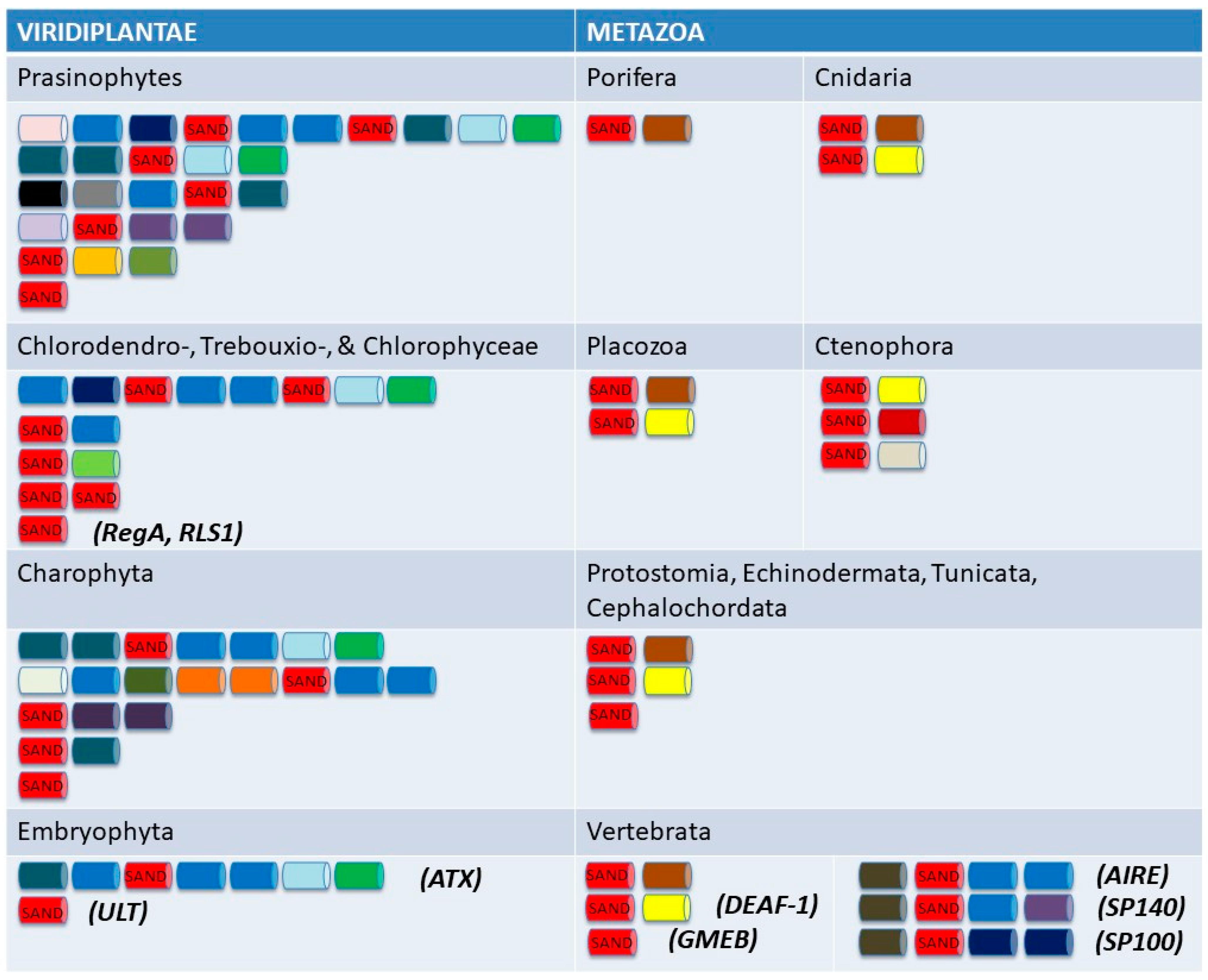
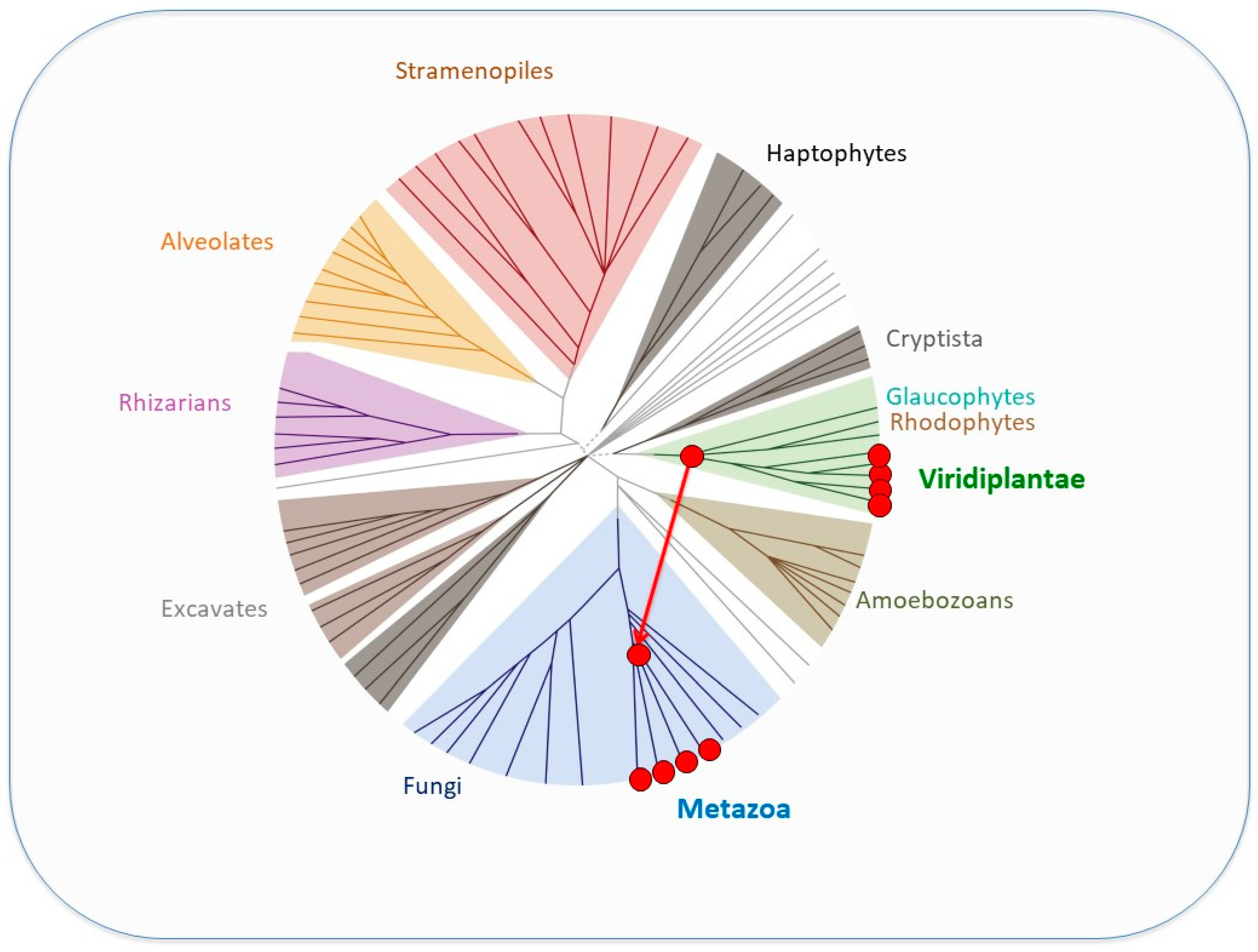
5. SAND-Domain-Containing Sequences beyond Volvocine Algae
6. Functional Evolution of regA via Co-Option of a Life History Trade-Off Gene
6.1. Life History Trade-Offs in Single-Celled Organisms
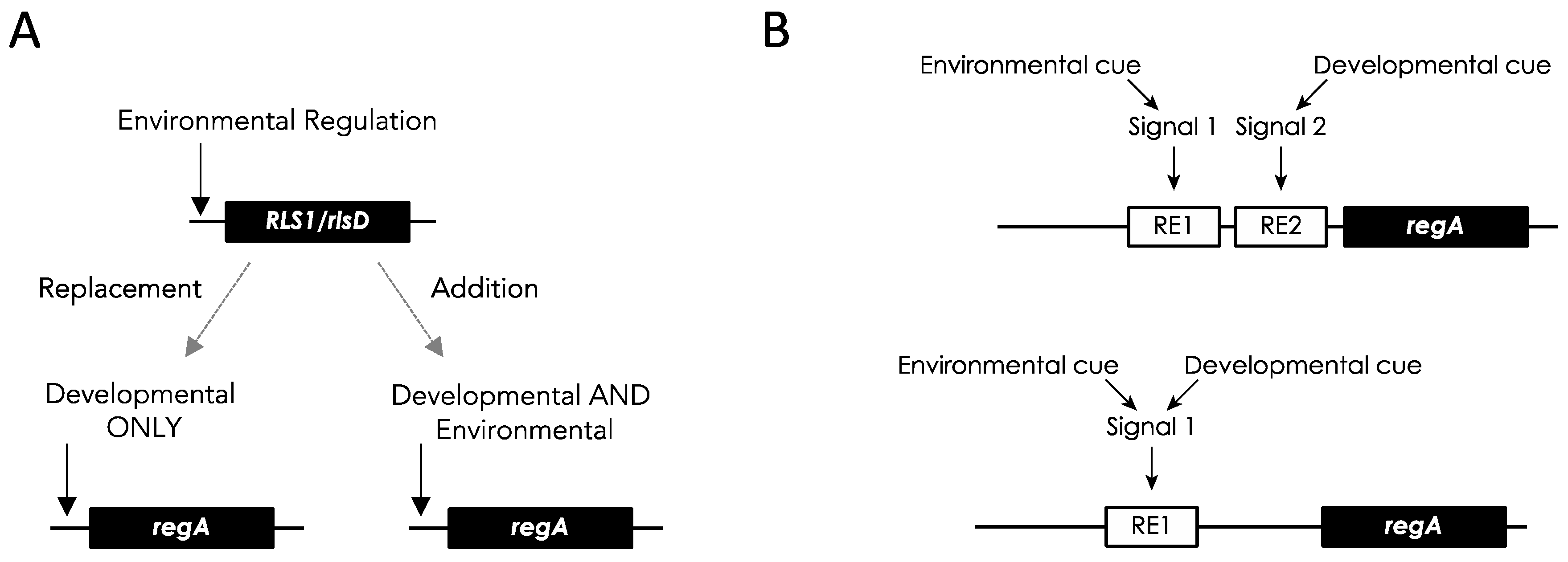
6.2. Chlamydomonas RLS1 Is a Life History Trade-Off Gene Induced via Environmental Cues
6.3. V. carteri regA Retained the Ancestral Environmental Regulation
6.4. Co-Option of an Environmentally Regulated Gene into a Developmental Master Regulator
7. The General Role of Stress and Life History Trade-Off Genes in the Re-Organization of Fitness during the Evolution of Multicellularity
8. Future Directions
9. Summary
Author Contributions
Funding
Conflicts of Interest
References
- Buss, L. The Evolution of Individuality; Princeton University Press: Princeton, NJ, USA, 1987. [Google Scholar]
- Maynard Smith, J.; Szathmáry, E. The Major Transitions in Evolution; Oxford University Press: Oxford, UK, 1995. [Google Scholar]
- Michod, R.E. Darwinian Dynamics; Princeton University Press: Princeton, NJ, USA, 1999. [Google Scholar]
- Grosberg, R.K.; Strathmann, R.R. The Evolution of Multicellularity: A Minor Major Transition? Annu. Rev. Ecol. Evol. Syst. 2007, 38, 621–654. [Google Scholar] [CrossRef]
- Herron, M.D.; Conlin, P.L.; Ratcliff, W.C. The Evolution of Multicellularity; CRC Press: Boca Raton, FL, USA, 2022. [Google Scholar] [CrossRef]
- Bonner, J.T. The origins of multicellularity. Integr. Biol. Issues News Rev. 1998, 1, 27–36. [Google Scholar] [CrossRef]
- Boraas, M.E.; Seale, D.B.; Boxhorn, J.E. Phagotrophy by flagellate selects for colonial prey: A possible origin of multicellularity. Evol. Ecol. 1998, 12, 153–164. [Google Scholar] [CrossRef]
- Herron, M.D.; Borin, J.M.; Boswell, J.C.; Walker, J.; Chen, I.K.; Knox, C.A.; Boyd, M.; Rosenzweig, F.; Ratcliff, W.C. De novo origins of multicellularity in response to predation. Sci. Rep. 2019, 9, 2328. [Google Scholar] [CrossRef]
- Ratcliff, W.C.; Denison, R.F.; Borrello, M.; Travisano, M. Experimental evolution of multicellularity. Proc. Natl. Acad. Sci. USA 2012, 109, 1595–1600. [Google Scholar] [CrossRef]
- Ratcliff, W.C.; Herron, M.D.; Howell, K.; Pentz, J.T.; Rosenzweig, F.; Travisano, M. Experimental evolution of an alternating uni- and multicellular life cycle in Chlamydomonas reinhardtii. Nat. Commun. 2013, 4, 2742. [Google Scholar] [CrossRef]
- Rose, C.J.; Hammerschmidt, K.; Rainey, P.B. Experimental evolution of nascent multicellularity: Recognizing a Darwinian transition in individuality. BioRxiv 2020. [Google Scholar] [CrossRef]
- Michod, R.E.; Nedelcu, A.M. On the reorganization of fitness during evolutionary transitions in individuality. Integr. Comp. Biol. 2003, 43, 64–73. [Google Scholar] [CrossRef]
- Nedelcu, A.M.; Michod, R.E. Evolvability, modularity, and individuality during the transition to multicellularity in volvocalean green algae. In Modularity in Development and Evolution; Schlosser, G., Wagner, G.P., Eds.; University of Chicago Press: Chicago, IL, USA, 2003. [Google Scholar]
- Michod, R.E. On the transfer of fitness from the cell to the multicellular organism. Biol. Philos. 2006, 20, 967–987. [Google Scholar] [CrossRef]
- Michod, R.E. The group covariance effect and fitness trade-offs during evolutionary transitions in individuality. Proc. Natl. Acad. Sci. USA 2006, 103, 9113–9117. [Google Scholar] [CrossRef]
- Nedelcu, A.M. Environmentally induced responses co-opted for reproductive altruism. Biol. Lett. 2009, 5, 805–808. [Google Scholar] [CrossRef]
- Nedelcu, A.M.; Michod, R.E. The evolutionary origin of an altruistic gene. Mol. Biol. Evol. 2006, 23, 1460–1464. [Google Scholar] [CrossRef] [PubMed]
- Brunet, T.; King, N. The Origin of Animal Multicellularity and Cell Differentiation. Dev. Cell 2017, 43, 124–140. [Google Scholar] [CrossRef]
- Olson, B.J.S.C.; Nedelcu, A.M. Co-option during the evolution of multicellularity and developmental complexity in the volvocine green algae. Curr. Opin. Genet. Dev. 2016, 39, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Sharpe, S.C.; Eme, L.; Brown, M.W.; Roger, A.J. Timing the Origins of Multicellular Eukaryotes Through Phylogenomics and Relaxed Molecular Clock Analyses. In Evolutionary Transitions to Multicellular Life; Nedelcu, A.M., Ruiz-Trillo, I., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 3–30. [Google Scholar]
- Hanschen, E.R.; Davison, D.R.; Grochau-Wright, Z.I.; Michod, R.E. Evolution of individuality: A case study in the volvocine green algae. Philos. Theory Pract. Biol. 2017, 9, 3. [Google Scholar] [CrossRef]
- Herron, M.D.; Hackett, J.D.; Aylward, F.O.; Michod, R.E. Triassic origin and early radiation of multicellular volvocine algae. Proc. Natl. Acad. Sci. USA 2009, 106, 3254–3258. [Google Scholar] [CrossRef]
- Umen, J.G. Volvox and volvocine green algae. EvoDevo 2020, 11, 7–15. [Google Scholar] [CrossRef]
- König, S.G.; Nedelcu, A.M. The genetic basis for the evolution of soma: Mechanistic evidence for the co-option of a stress-induced gene into a developmental master regulator. Proc. R. Soc. B Biol. Sci. 2020, 287, 20201414. [Google Scholar] [CrossRef] [PubMed]
- Grochau-Wright, Z.I.; Hanschen, E.R.; Ferris, P.J.; Hamaji, T.; Nozaki, H.; Olson, B.J.S.C.; Michod, R.E. Genetic Basis for Soma is Present in Undifferentiated Volvocine Green Algae. J. Evol. Biol. 2017, 30, 1205–1218. [Google Scholar] [CrossRef]
- Coleman, A.W. A Comparative Analysis of the Volvocaceae (Chlorophyta). J. Phycol. 2012, 48, 491–513. [Google Scholar] [CrossRef]
- Herron, M.D.; Michod, R.E. Evolution of complexity in the volvocine algae: Transitions in individuality through Darwin’s eye. Evol. Int. J. Org. Evol. 2008, 62, 436–451. [Google Scholar] [CrossRef] [PubMed]
- Kirk, D.L. A twelve-step program for evolving multicellularity and a division of labor. BioEssays News Rev. Mol. Cell. Dev. Biol. 2005, 27, 299–310. [Google Scholar] [CrossRef] [PubMed]
- Nozaki, H.; Misawa, K.; Kajita, T.; Kato, M.; Nohara, S.; Watanabe, M.M. Origin and evolution of the colonial volvocales (Chlorophyceae) as inferred from multiple, chloroplast gene sequences. Mol. Phylogenet. Evol. 2000, 17, 256–268. [Google Scholar] [CrossRef] [PubMed]
- Arakaki, Y.; Kawai-Toyooka, H.; Hamamura, Y.; Higashiyama, T.; Noga, A.; Hirono, M.; Olson, B.J.S.C.; Nozaki, H. The simplest integrated multicellular organism unveiled. PLoS ONE 2013, 8, e81641. [Google Scholar] [CrossRef]
- Nozaki, H.; Itoh, M.; Watanabe, M.M.; Kuroiwa, T. Ultrastructure of the vegetative colonies and systematic position of basichlamys (Volvocales, Chlorophyta). Eur. J. Phycol. 1996, 31, 67–72. [Google Scholar] [CrossRef]
- Hanschen, E.R.; Marriage, T.N.; Ferris, P.J.; Hamaji, T.; Toyoda, A.; Fujiyama, A.; Neme, R.; Noguchi, H.; Minakuchi, Y.; Suzuki, M.; et al. The Gonium pectorale genome demonstrates co-option of cell cycle regulation during the evolution of multicellularity. Nat. Commun. 2016, 7, 11370. [Google Scholar] [CrossRef]
- Yamashita, S.; Yamamoto, K.; Matsuzaki, R.; Suzuki, S.; Yamaguchi, H.; Hirooka, S.; Minakuchi, Y.; Miyagishima, S.-Y.; Kawachi, M.; Toyoda, A.; et al. Genome sequencing of the multicellular alga Astrephomene provides insights into convergent evolution of germ-soma differentiation. Sci. Rep. 2021, 11, 22231. [Google Scholar] [CrossRef]
- Nozaki, H.; Mahakham, W.; Athibai, S.; Yamamoto, K.; Takusagawa, M.; Misumi, O.; Herron, M.D.; Rosenzweig, F.; Kawachi, M. Rediscovery of the species of ‘ancestral Volvox’: Morphology and phylogenetic position of Pleodorina sphaerica (Volvocales, Chlorophyceae) from Thailand. ’ Phycologia 2017, 56, 469–475. [Google Scholar] [CrossRef]
- Lindsey, C.R.; Rosenzweig, F.; Herron, M.D. Phylotranscriptomics points to multiple independent origins of multicellularity and cellular differentiation in the volvocine algae. BMC Biol. 2021, 19, 182. [Google Scholar] [CrossRef]
- Kirk, D.L. Volvox: Molecular-Genetic Origins of Multicellularity and Cellular Differentiation; Cambridge University Press: Cambridge, UK, 1998. [Google Scholar]
- Kirk, D.L. Germ-soma differentiation in Volvox. Dev. Biol. 2001, 238, 213–223. [Google Scholar] [CrossRef]
- Koufopanou, V. The Evolution of Soma in the Volvocales. Am. Nat. 1994, 143, 907–931. [Google Scholar] [CrossRef]
- Kirk, M.M.; Ransick, A.; McRae, S.E.; Kirk, D.L. The Relationship between Cell Size and Cell Fate in Volvox carteri. J. Cell Biol. 1993, 123, 191–208. [Google Scholar] [CrossRef] [PubMed]
- Grochau-Wright, Z.I. The Origin and Evolution of the Reg Cluster in the Volvocine Green Algae: A Model System for the Evolution of Cellular Differentiation. Ph.D. Dissertation, University of Arizona, Tucson, AZ, USA, 2019. [Google Scholar]
- Grochau-Wright, Z.I.; Ferris, P.J.; Tumberger, J.; Jiménez-Marin, B.; Olson, B.J.S.C.; Michod, R.E. Characterization and Transformation of reg Cluster Genes in Volvox powersii Enable Investigation of Convergent Evolution of Cellular Differentiation in Volvox. Protist 2021, 172, 125834. [Google Scholar] [CrossRef]
- Ransick, A. Reproductive cell specification during Volvox obversus development. Dev. Biol. 1991, 143, 185–198. [Google Scholar] [CrossRef] [PubMed]
- Ransick, A. Specification of reproductive cells in Volvox. In Evolutionary Conservation of Developmental Mechanisms, Proceedings of the 50th Symposium of the Society for Developmental Biology, Marquette University, Milwaukee, WI, USA, 20–23 June 1991; Spradling, A., Ed.; Wiley-Liss: Hoboken, NJ, USA, 1993; pp. 55–70. [Google Scholar]
- Sessoms, A.H.; Huskey, R.J. Genetic Control of Development in Volvox: Isolation and Characterization of Morphogenetic Mutants. Proc. Natl. Acad. Sci. USA 1973, 70, 1335–1338. [Google Scholar] [CrossRef]
- Starr, R.C. Control of differentiation in Volvox. Dev. Biol. 1970, 4, 59–100. [Google Scholar]
- Huskey, R.J.; Griffin, B.E.; Cecil, P.O.; Callahan, A.M. A Preliminary Genetic Investigation of Volvox carteri. Genetics 1979, 91, 229–244. [Google Scholar] [CrossRef] [PubMed]
- Huskey, R.J.; Griffin, B.E. Genetic Control of Somatic Cell Differentiation in Volvox. Dev. Biol. 1979, 72, 226–235. [Google Scholar] [CrossRef]
- Baran, G. Analysis of Somatic Cell Differentiation in Volvox carteri f. nagariensis; University of Virginia: Charlottesville, VA, USA, 1984. [Google Scholar]
- Kirk, M.M.; Stark, K.; Miller, S.M.; Müller, W.; Taillon, B.E.; Gruber, H.; Schmitt, R.; Kirk, D.L. regA, a Volvox gene that plays a central role in germ-soma differentiation, encodes a novel regulatory protein. Development 1999, 126, 639–647. [Google Scholar] [CrossRef]
- Harryman, A. Investigating the Roles of regA and Related Genes in the Evolution of Multicellularity in the Volvocine Green Algae. Ph.D. Thesis, University of Maryland, Baltimore, MD, USA, 2012. [Google Scholar]
- Klein, B.; Wibberg, D.; Hallmann, A. Whole transcriptome RNA-Seq analysis reveals extensive cell type-specific compartmentalization in Volvox carteri. BMC Biol. 2017, 15, 1–22. [Google Scholar] [CrossRef]
- Matt, G.Y.; Umen, J.G. Cell-Type Transcriptomes of the Multicellular Green Alga Volvox carteri Yield Insights into the Evolutionary Origins of Germ and Somatic Differentiation Programs. G3 Genes|Genomes|Genet. 2018, 8, 531–550. [Google Scholar] [CrossRef] [PubMed]
- Duncan, L.; Nishii, I.; Howard, A.; Kirk, D.; Miller, S.M. Orthologs and paralogs of regA, a master cell-type regulatory gene in Volvox carteri. Curr. Genet. 2006, 50, 61–72. [Google Scholar] [CrossRef]
- Choi, G.; Przybylska, M.; Straus, D. Three abundant germ line-specific transcripts in Volvox carteri encode photosynthetic proteins. Curr. Genet. 1996, 30, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Meissner, M.; Stark, K.; Cresnar, B.; Kirk, D.L.; Schmitt, R. Volvox germline-specific genes that are putative targets of RegA repression encode chloroplast proteins. Curr. Genet. 1999, 36, 363–370. [Google Scholar] [CrossRef]
- Tam, L.; Kirk, D.L. Identification of Cell-Type-Specific Characterization of their Expression Genes of Volvox carteri and during the Asexual Life Cycle. Dev. Biol. 1991, 145, 51–66. [Google Scholar] [CrossRef]
- Stark, K.; Kirk, D.L.; Schmitt, R. Two enhancers and one silencer located in the introns of regA control somatic cell differentiation in Volvox carteri. Genes Dev. 2001, 15, 1449–1460. [Google Scholar] [CrossRef] [PubMed]
- Babinger, K.; Hallmann, A.; Schmitt, R. Translational control of regA, a key gene controlling cell differentiation in Volvox carteri. Development 2006, 133, 4045–4051. [Google Scholar] [CrossRef]
- Duncan, L.; Nishii, I.; Harryman, A.; Buckley, S.; Howard, A.; Friedman, N.R.; Miller, S.M. The VARL gene family and the evolutionary origins of the master cell-type regulatory gene, regA, in Volvox carteri. J. Mol. Evol. 2007, 65, 1–11. [Google Scholar] [CrossRef]
- Bottomley, M.J.J.; Collard, M.W.W.; Huggenvik, J.I.I.; Liu, Z.; Gibson, T.J.J.; Sattler, M. The SAND domain structure defines a novel DNA-binding fold in transcriptional regulation. Nat. Struct. Biol. 2001, 8, 626–633. [Google Scholar] [CrossRef]
- Barker, H.E.; Smyth, G.K.; Wettenhall, J.; Ward, T.A.; Bath, M.L.; Lindeman, G.J.; Visvader, J.E. Deaf-1 regulates epithelial cell proliferation and side-branching in the mammary gland. BMC Dev. Biol. 2008, 8, 94. [Google Scholar] [CrossRef]
- Veraksa, A.; Kennison, J.; McGinnis, W. DEAF-1 function is essential for the early embryonic development of Drosophila. Genesis 2002, 33, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, T.; Tsuruma, K.; Uehara, T.; Nomura, Y. GMEB1, a novel endogenous caspase inhibitor, prevents hypoxia- and oxidative stress-induced neuronal apoptosis. Neurosci. Lett. 2008, 438, 34–37. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, M.; Shakes, D.C.; Guevel, K.; Smith, H.E. SPE-44 Implements Sperm Cell Fate. PLoS Genet. 2012, 8, e1002678. [Google Scholar] [CrossRef]
- Radhakrishnan, K.; Bhagya, K.P.; Kumar, A.T.; Devi, A.N.; Sengottaiyan, J.; Kumar, P.G. Autoimmune Regulator (AIRE) Is Expressed in Spermatogenic Cells, and It Altered the Expression of Several Nucleic-Acid-Binding and Cytoskeletal Proteins in Germ Cell 1 Spermatogonial (GC1-spg) Cells. Mol. Cell. Proteom. 2016, 15, 2686–2698. [Google Scholar] [CrossRef]
- Schaller, C.E.; Wang, C.L.; Beck-engeser, G.; Goss, L.; Scott, H.S.; Anderson, M.S.; Wabl, M. Expression of Aire and the Early Wave of Apoptosis in Spermatogenesis. J. Immunol. 2008, 180, 1338–1343. [Google Scholar] [CrossRef]
- Chen, L.-Q.; Luo, J.-H.; Cui, Z.-H.; Xue, M.; Wang, L.; Zhang, X.-Y.; Pawlowski, W.P.; He, Y. ATX3, ATX4, and ATX5 Encode Putative H3K4 Methyltransferases and Are Critical for Plant Development. Plant Physiol. 2017, 174, 1795–1806. [Google Scholar] [CrossRef] [PubMed]
- Carles, C.C. ULTRAPETALA1 encodes a SAND domain putative transcriptional regulator that controls shoot and floral meristem activity in Arabidopsis. Development 2005, 132, 897–911. [Google Scholar] [CrossRef]
- Carles, C.C.; Fletcher, J.C. The SAND domain protein ULTRAPETALA1 acts as a trithorax group factor to regulate cell fate in plants. Genes Dev. 2009, 23, 2723–2728. [Google Scholar] [CrossRef]
- Fletcher, J.C. The ULTRAPETALA gene controls shoot and floral meristem size in Arabidopsis. Development 2001, 128, 1323–1333. [Google Scholar] [CrossRef]
- Nedelcu, A.M. Independent evolution of complex development in animals and plants: Deep homology and lateral gene transfer. Dev. Genes Evol. 2019, 229, 25–34. [Google Scholar] [CrossRef]
- Featherston, J.; Arakaki, Y.; Hanschen, E.R.; Ferris, P.J.; Michod, R.E.; Olson, B.J.S.C.; Nozaki, H.; Durand, P.M. The 4-celled tetrabaena socialis nuclear genome reveals the essential components for genetic control of cell number at the origin of multicellularity in the volvocine lineage. Mol. Biol. Evol. 2018, 35, 855–870. [Google Scholar] [CrossRef]
- Hanschen, E.R.; Ferris, P.J.; Michod, R.E. Early Evolution of the Genetic Basis for Soma in the Volvocaceae. Evolution 2014, 68, 2014–2025. [Google Scholar] [CrossRef] [PubMed]
- Keeling, P.J.; Palmer, J.D. Horizontal gene transfer in eukaryotic evolution. Nat. Rev. Genet. 2008, 9, 605–618. [Google Scholar] [CrossRef] [PubMed]
- Keeling, P.J.; Burki, F. Progress towards the Tree of Eukaryotes. Curr. Biol. 2019, 29, R808–R817. [Google Scholar] [CrossRef] [PubMed]
- Knoll, A.H. The Multiple Origins of Complex Multicellularity. Annu. Rev. Earth Planet. Sci. 2011, 39, 217–239. [Google Scholar] [CrossRef]
- Bochdanovits, Z.; De Jong, G. Antagonistic pleiotropy for life-history traits at the gene expression level. Proc. R. Soc. B Biol. Sci. 2004, 271 (Suppl. S3), S75–S78. [Google Scholar] [CrossRef]
- Flatt, T. Life-history evolution and the genetics of fitness components in drosophila melanogaster. Genetics 2020, 214, 3–48. [Google Scholar] [CrossRef]
- Stearns, S.C. Trade-Offs in Life-History Evolution. Funct. Ecol. 1989, 3, 259–268. [Google Scholar] [CrossRef]
- Flatt, T.; Heyland, A. Mechanisms of Life History Evolution: The Genetics and Physiology of Life History Traits and Trade-Offs; Oxford University Press: Oxford, UK, 2011. [Google Scholar]
- Hood, W.R.; Zhang, Y.; Mowry, A.V.; Hyatt, H.W.; Kavazis, A.N. Life History Trade-offs within the Context of Mitochondrial Hormesis. Integr. Comp. Biol. 2018, 58, 567–577. [Google Scholar] [CrossRef]
- Hughes, K.A.; Leips, J. Pleiotropy, constraint, and modularity in the evolution of life histories: Insights from genomic analyses. Ann. N. Y. Acad. Sci. 2017, 1389, 76–91. [Google Scholar] [CrossRef]
- Koch, R.E.; Buchanan, K.L.; Casagrande, S.; Crino, O.; Dowling, D.K.; Hill, G.E.; Hood, W.R.; McKenzie, M.; Mariette, M.M.; Noble, D.W.A.; et al. Integrating Mitochondrial Aerobic Metabolism into Ecology and Evolution. Trends Ecol. Evol. 2021, 36, 321–332. [Google Scholar] [CrossRef] [PubMed]
- Leroi, A.M. Molecular signals versus the Loi de Balancement. Trends Ecol. Evol. 2001, 16, 24–29. [Google Scholar] [CrossRef]
- Monaghan, P.; Metcalfe, N.B.; Torres, R. Oxidative stress as a mediator of life history trade-offs: Mechanisms, measurements and interpretation. Ecol. Lett. 2009, 12, 75–92. [Google Scholar] [CrossRef] [PubMed]
- Roff, D.A.; Fairbairn, D.J. The evolution of trade-offs: Where are we? J. Evol. Biol. 2007, 20, 433–447. [Google Scholar] [CrossRef] [PubMed]
- Villellas, J.; García, M.B. Life-history trade-offs vary with resource availability across the geographic range of a widespread plant. Plant Biol. 2018, 20, 483–489. [Google Scholar] [CrossRef]
- Zera, A.J.; Harshman, L.G. The Physiology of Life History Trade-Offs in Animals. Annu. Rev. Ecol. Syst. 2001, 32, 95–126. [Google Scholar] [CrossRef]
- Wykoff, D.D.; Davies, J.P.; Melis, A.; Grossman, A.R. The regulation of photosynthetic electron transport during nutrient deprivation in Chlamydomonas reinhardtii. Plant Physiol. 1998, 117, 129–139. [Google Scholar] [CrossRef]
- Grossman, A. Acclimation of Chlamydomonas reinhardtii to its nutrient environment. Protist 2000, 151, 201–224. [Google Scholar] [CrossRef]
- Saggere, R.M.S.; Lee, C.W.J.; Chan, I.C.W.; Durnford, D.G.; Nedelcu, A.M. A life-history trade-off gene with antagonistic pleiotropic effects on reproduction and survival in limiting environments. Proc. R. Soc. B 2022, 288, 20212669. [Google Scholar] [CrossRef]
- Pfannschmidt, T.; Bräutigam, K.; Wagner, R.; Dietzel, L.; Schröter, Y.; Steiner, S.; Nykytenko, A. Potential regulation of gene expression in photosynthetic cells by redox and energy state: Approaches towards better understanding. Ann. Bot. 2009, 103, 599. [Google Scholar] [CrossRef]
- Koshikawa, S.; Giorgianni, M.W.; Vaccaro, K.; Kassner, V.A.; Yoder, J.H.; Werner, T.; Carroll, S.B. Gain of cis-regulatory activities underlies novel domains of wingless gene expression in Drosophila. Proc. Natl. Acad. Sci. USA 2015, 112, 7524–7529. [Google Scholar] [CrossRef]
- Alvarez-Curto, E.; Rozen, D.E.; Ritchie, A.V.; Fouquet, C.; Baldauf, S.L.; Schaap, P. Evolutionary origin of cAMP-based chemoattraction in the social amoebae. Proc. Natl. Acad. Sci. USA 2005, 102, 6385–6390. [Google Scholar] [CrossRef]
- Zhang, J. Evolution by gene duplication: An update. Trends Ecol. Evol. 2003, 18, 292–298. [Google Scholar] [CrossRef]
- Ortega Escalante, J.A. Investigation of Volvox carteri Cell Differentiation and Its Evolution Through Functional Analysis of regA and regA Homologues. Ph.D. Dissertation, University of Maryland, Baltimore, MD, USA, 2018. [Google Scholar]
- Strand, D.D.; Livingston, A.K.; Satoh-Cruz, M.; Koepke, T.; Enlow, H.M.; Fisher, N.; Froehlich, J.E.; Cruz, J.A.; Minhas, D.; Hixson, K.K.; et al. Defects in the Expression of Chloroplast Proteins Leads to H2O2 Accumulation and Activation of Cyclic Electron Flow around Photosystem I. Front. Plant Sci. 2017, 7, 2073. [Google Scholar] [CrossRef]
- Cameron-Pack, M.E.; König, S.G.; Reyes-Guevara, A.; Reyes-Prieto, A.; Nedelcu, A.M. A personal cost of cheating can stabilize reproductive altruism during the early evolution of clonal multicellularity. Biol. Lett. 2022, 18, 20220059. [Google Scholar] [CrossRef]
- Nedelcu, A.M. Evidence for p53-like-mediated stress responses in green algae. FEBS Lett. 2006, 580, 3013–3017. [Google Scholar] [CrossRef]
- Jimenez Marin, L.B. Gene Loss, Co-Option and the Evolution of Developmental Complexity in the Volvocine Algae. Ph.D. Dissertation, Kansas State University, Manhattan, KS, USA, 2023. [Google Scholar]
- Ferenci, T. Trade-off Mechanisms Shaping the Diversity of Bacteria. Trends Microbiol. 2016, 24, 209–223. [Google Scholar] [CrossRef]
- Lang, G.I.; Murray, A.W.; Botstein, D. The cost of gene expression underlies a fitness trade-off in yeast. Proc. Natl. Acad. Sci. USA 2009, 106, 5755–5760. [Google Scholar] [CrossRef] [PubMed]
- Wenger, J.W.; Piotrowski, J.; Nagarajan, S.; Chiotti, K.; Sherlock, G.; Rosenzweig, F. Hunger Artists: Yeast Adapted to Carbon Limitation Show Trade-Offs under Carbon Sufficiency. PLoS Genet. 2011, 7, e1002202. [Google Scholar] [CrossRef] [PubMed]
- Wolf, J.B.; Howie, J.A.; Parkinson, K.; Gruenheit, N.; Melo, D.; Rozen, D.; Thompson, C.R.L. Fitness Trade-offs Result in the Illusion of Social Success. Curr. Biol. CB 2015, 25, 1086–1090. [Google Scholar] [CrossRef]
- Lee, C.; Raffaghello, L.; Brandhorst, S.; Safdie, F.M.; Bianchi, G.; Martin-Montalvo, A.; Pistoia, V.; Wei, M.; Hwang, S.; Merlino, A.; et al. Fasting Cycles Retard Growth of Tumors and Sensitize a Range of Cancer Cell Types to Chemotherapy. Sci. Transl. Med. 2012, 4, 124ra27. [Google Scholar] [CrossRef] [PubMed]
- Raffaghello, L.; Lee, C.; Safdie, F.M.; Wei, M.; Madia, F.; Bianchi, G.; Longo, V.D. Starvation-dependent differential stress resistance protects normal but not cancer cells against high-dose chemotherapy. Proc. Natl. Acad. Sci. USA 2008, 105, 8215–8220. [Google Scholar] [CrossRef] [PubMed]
- Nedelcu, A.M.; Michod, R.E. Stress Responses Co-Opted for Specialized Cell Types During the Early Evolution of Multicellularity. BioEssays 2020, 42, 2000029. [Google Scholar] [CrossRef] [PubMed]
- Schaap, P. Evolution of developmental signalling in Dictyostelid social amoebas. Curr. Opin. Genet. Dev. 2016, 39, 29–34. [Google Scholar] [CrossRef]
- Wagner, G.P.; Erkenbrack, E.M.; Love, A.C. Stress-Induced Evolutionary Innovation: A Mechanism for the Origin of Cell Types. BioEssays 2019, 41, 1800188. [Google Scholar] [CrossRef]
- Davison, D.R.; Michod, R.E. Phenotypic Plasticity and Evolutionary Transitions in Individuality. In Phenotypic Plasticity & Evolution: Causes, Consequences, Controversies; Pfennig, D.W., Ed.; CRC Press: Boca Raton, FL, USA, 2021; pp. 241–266. [Google Scholar]
- Hamaji, T.; Kawai-Toyooka, H.; Uchimura, H.; Suzuki, M.; Noguchi, H.; Minakuchi, Y.; Toyoda, A.; Fujiyama, A.; Miyagishima, S.; Umen, J.G.; et al. Anisogamy evolved with a reduced sex-determining region in volvocine green algae. Commun. Biol. 2018, 1, 17. [Google Scholar] [CrossRef]
- Yamamoto, K.; Hamaji, T.; Kawai-Toyooka, H.; Matsuzaki, R.; Takahashi, F.; Nishimura, Y.; Kawachi, M.; Noguchi, H.; Minakuchi, Y.; Umen, J.G.; et al. Three genomes in the algal genus Volvox reveal the fate of a haploid sex-determining region after a transition to homothallism. Proc. Natl. Acad. Sci. USA 2021, 118, e2100712118. [Google Scholar] [CrossRef]
- Yamashita, S.; Arakaki, Y.; Kawai-Toyooka, H.; Noga, A.; Hirono, M.; Nozaki, H. Alternative evolution of a spheroidal colony in volvocine algae: Developmental analysis of embryogenesis in Astrephomene (Volvocales, Chlorophyta). BMC Evol. Biol. 2016, 16, 243. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grochau-Wright, Z.I.; Nedelcu, A.M.; Michod, R.E. The Genetics of Fitness Reorganization during the Transition to Multicellularity: The Volvocine regA-like Family as a Model. Genes 2023, 14, 941. https://doi.org/10.3390/genes14040941
Grochau-Wright ZI, Nedelcu AM, Michod RE. The Genetics of Fitness Reorganization during the Transition to Multicellularity: The Volvocine regA-like Family as a Model. Genes. 2023; 14(4):941. https://doi.org/10.3390/genes14040941
Chicago/Turabian StyleGrochau-Wright, Zachariah I., Aurora M. Nedelcu, and Richard E. Michod. 2023. "The Genetics of Fitness Reorganization during the Transition to Multicellularity: The Volvocine regA-like Family as a Model" Genes 14, no. 4: 941. https://doi.org/10.3390/genes14040941