
Genome-Wide Identification of the KNOX Gene Family in Japanese Apricot (Prunus mume Sieb. et Zucc.) and Functional Characterization of PmKNAT2 Genes

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Identification of PmKNOX Family Members
2.3. Analysis of Structure and Conserved Motifs of PmKNOX Gene in Japanese Apricot
2.4. Chromosomal Location and Syntenic Analysis
2.5. Analysis of Cis-Acting Elements in PmKNOX Gene Upstream Promoter Region
2.6. Expression Profiling Analyses of PmKNOX Genes
2.7. PmKNAT2a and PmKNAT2b Cloning and Sequence Analysis
2.8. Vector Construction, Plant Transformation, and Identification of Transgenic Lines
2.9. Determination of Lignin Percentage Content
2.10. Measurement of Leaf Area
2.11. Statistical Analyses
3. Results
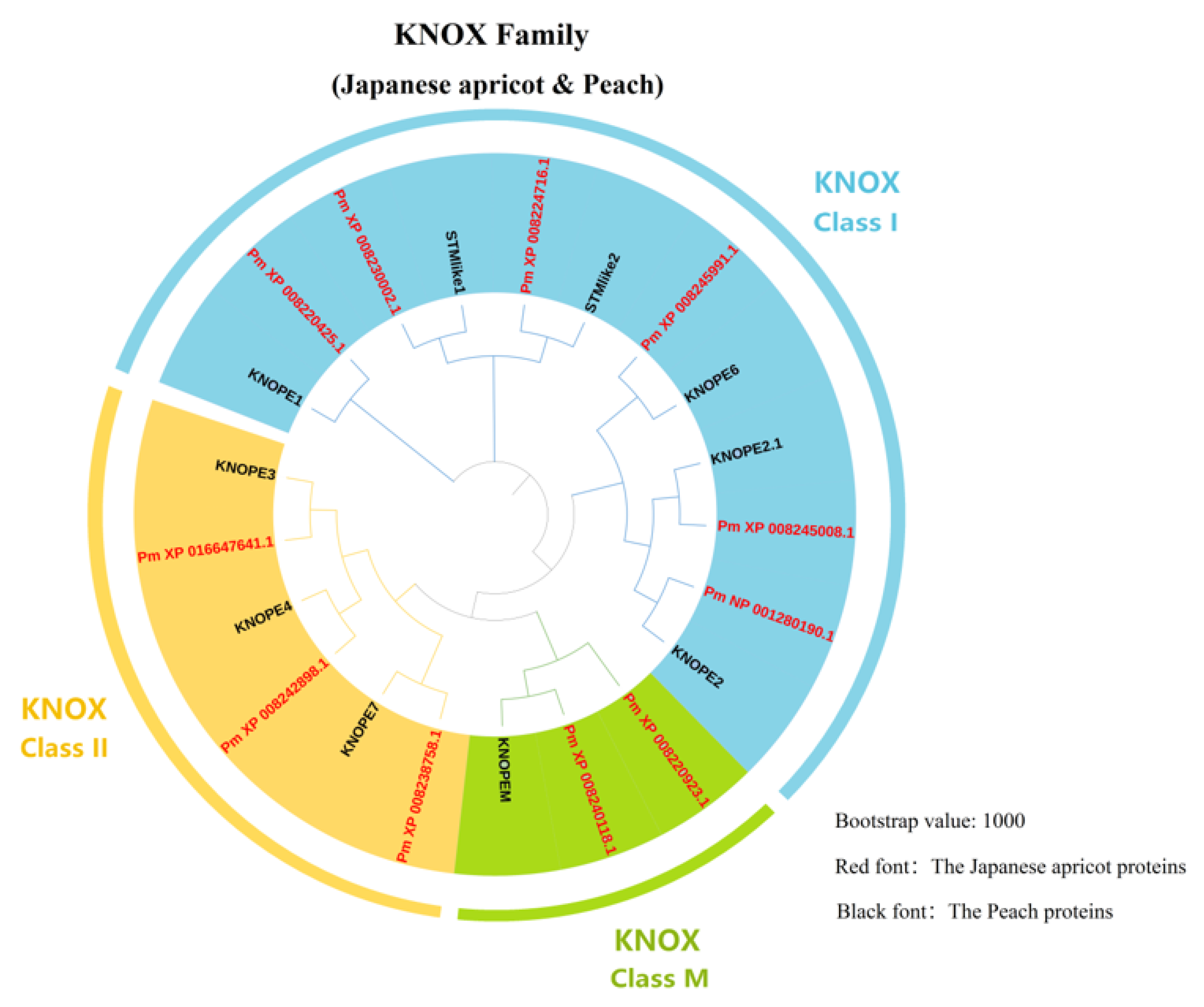
3.1. Identification of KNOX Family Members in Japanese Apricot
3.2. Secondary Structure Analysis of PmKNOX Protein
3.3. Gene Structures, Protein Conserved Domain and Motif Compositions of PmKNOX Proteins
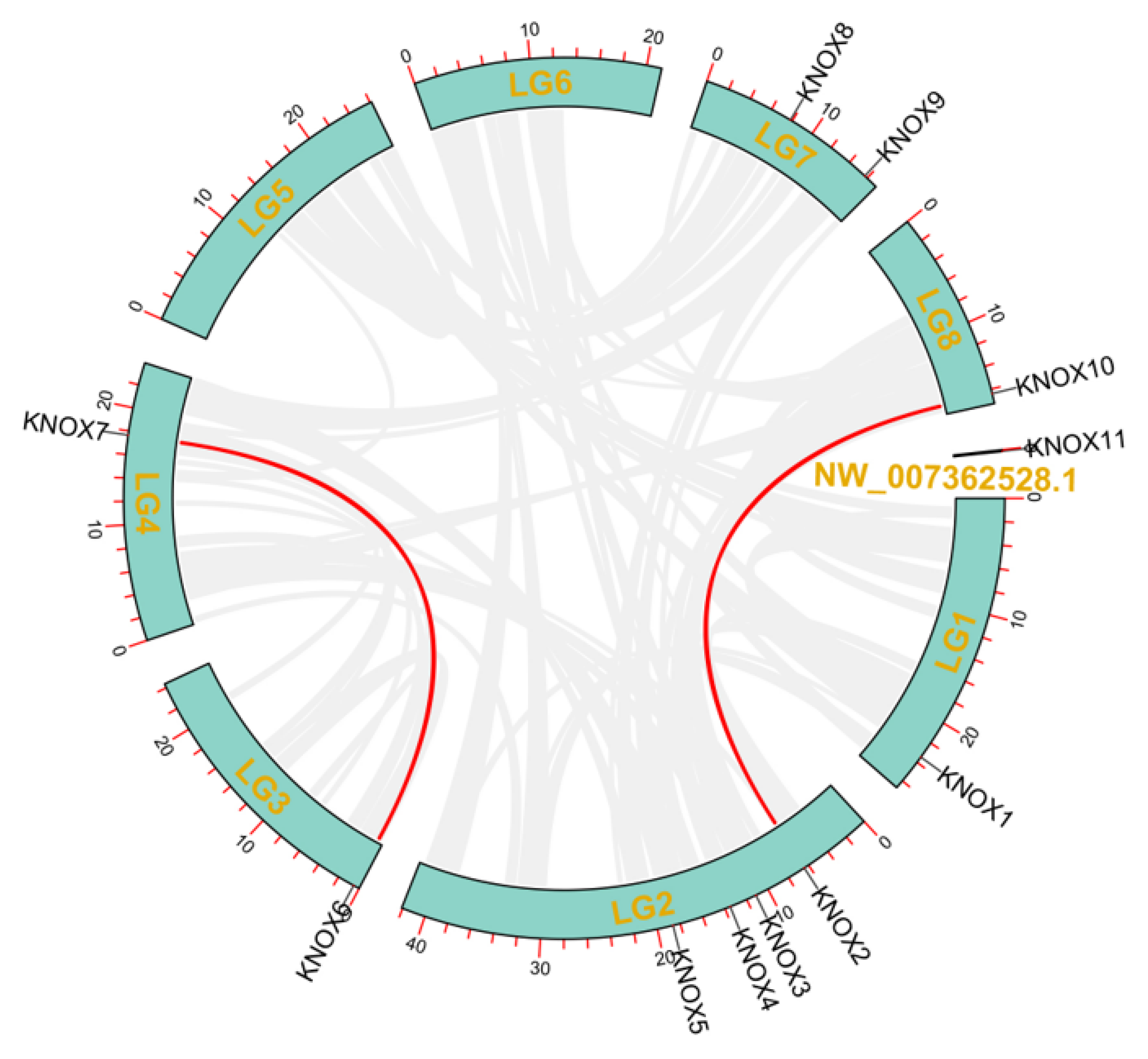
3.4. Chromosomal Distributions Analysis of PmKNOX Genes
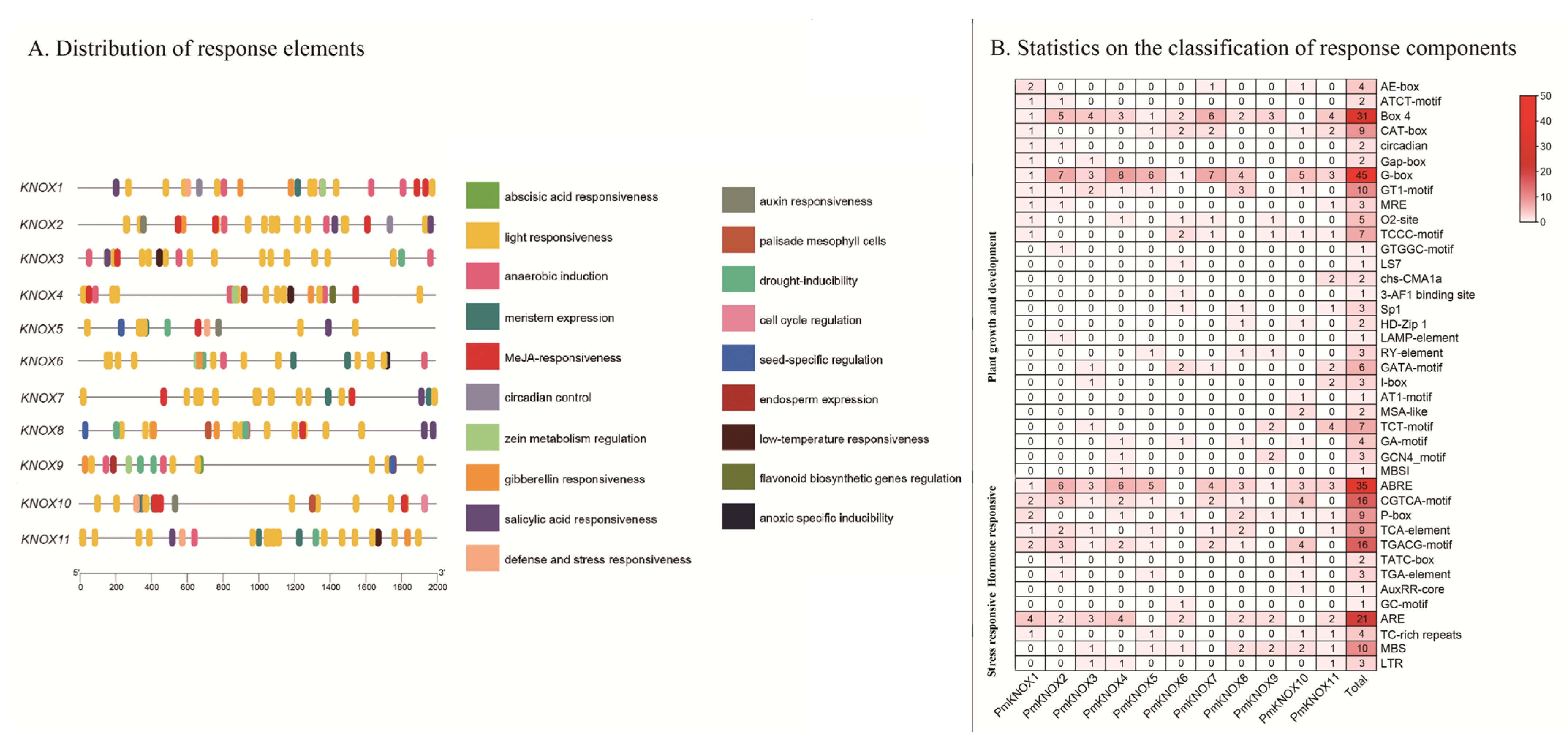
3.5. Cis-Acting Elements Analysis in PmKNOX Gene Promoter Region
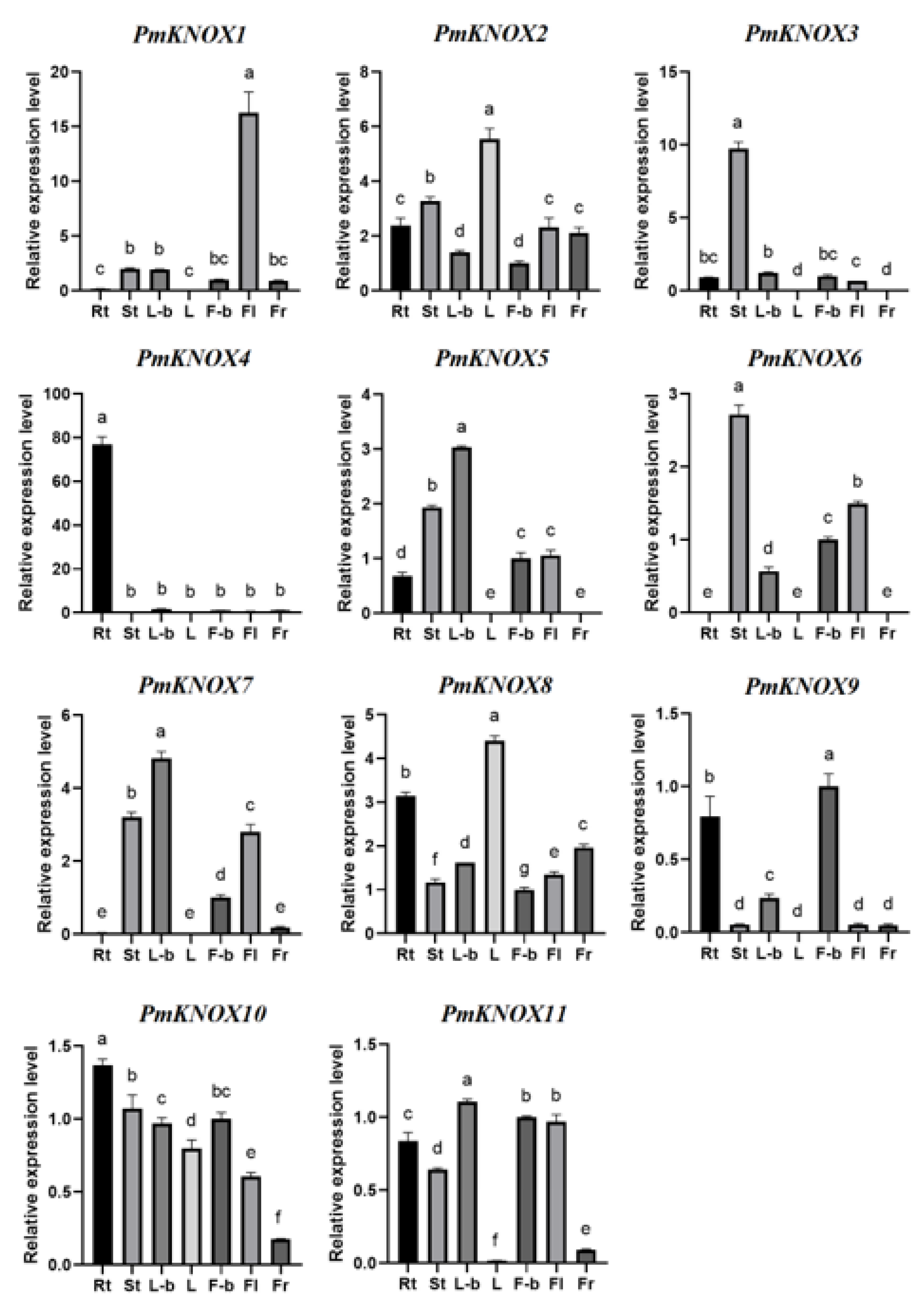
3.6. Expression Profiles Analysis of PmKNOX Genes in Different Tissues
3.7. Heterologous Expression of PmKNAT2 Genes in Arabidopsis
3.8. Effect of Heterologous Expression of PmKNAT2a and PmKNAT2b on Arabidopsis Stems
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vollbrecht, E.; Veit, B.; Sinha, N.; Hake, S. The developmental gene Knotted-1 is a member of a maize homeobox gene family. Nature 1991, 350, 241–243. [Google Scholar] [CrossRef]
- Smith, L.G.; Greene, B.; Veit, B.; Hake, S. A dominant mutation in the maize homeobox gene, Knotted-1, causes its ectopic expression in leaf cells with altered fates. Development 1992, 116, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yang, X.; Gao, Y.; Yang, S. Genome-Wide Identification and Characterization of TALE Superfamily Genes in Soybean (Glycine max L.). Int. J. Mol. Sci. 2021, 22, 4117. [Google Scholar] [CrossRef]
- Jia, P.; Zhang, C.; Xing, L.; Li, Y.; Shah, K.; Zuo, X.; Zhang, D.; An, N.; Han, M.; Ren, X. Genome-Wide Identification of the MdKNOX Gene Family and Characterization of Its Transcriptional Regulation in Malus domestica. Front. Plant Sci. 2020, 11, 128. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Lan, S.; Yin, W.L.; Liu, Z.J. Genome-Wide Identification and Expression Pattern Analysis of KNOX Gene Family in Orchidaceae. Front. Plant Sci. 2022, 13, 901089. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Pan, Y.; Zhi, C.; Zheng, Y.; Wang, X.; Li, X.; Cheng, Z. Genome-Wide Identification and Characterization of KNOTTED-Like Homeobox (KNOX) Homologs in Garlic (Allium sativum L.) and Their Expression Profilings Responding to Exogenous Cytokinin and Gibberellin. Int. J. Mol. Sci. 2021, 22, 9237. [Google Scholar] [CrossRef]
- Kerstetter, R.; Vollbrecht, E.; Lowe, B.; Veit, B.; Yamaguchi, J.; Hake, S. Sequence analysis and expression patterns divide the maize knotted1-like homeobox genes into two classes. Plant Cell 1994, 6, 1877–1887. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Cho, Y.H.; Ryu, H.; Kim, Y.; Kim, T.H.; Hwang, I. BLH1 and KNAT3 modulate ABA responses during germination and early seedling development in Arabidopsis. Plant J. 2013, 75, 755–766. [Google Scholar] [CrossRef]
- Li, E.; Bhargava, A.; Qiang, W.; Friedmann, M.C.; Forneris, N.; Savidge, R.A.; Johnson, L.A.; Mansfield, S.D.; Ellis, B.E.; Douglas, C.J. The Class II KNOX gene KNAT7 negatively regulates secondary wall formation in Arabidopsis and is functionally conserved in Populus. New Phytol. 2012, 194, 102–115. [Google Scholar] [CrossRef]
- Wang, S.; Yang, H.; Mei, J.; Liu, X.; Wen, Z.; Zhang, L.; Xu, Z.; Zhang, B.; Zhou, Y. Rice Homeobox Protein KNAT7 Integrates the Pathways Regulating Cell Expansion and Wall Stiffness. Plant Physiol. 2019, 181, 669–682. [Google Scholar] [CrossRef]
- Magnani, E.; Hake, S. KNOX lost the OX: The Arabidopsis KNATM gene defines a novel class of KNOX transcriptional regulators missing the homeodomain. Plant Cell 2008, 20, 875–887. [Google Scholar] [CrossRef]
- Bürglin, T.R. Analysis of TALE superclass homeobox genes (MEIS, PBC, KNOX, Iroquois, TGIF) reveals a novel domain conserved between plants and animals. Nucleic Acids Res. 1997, 25, 4173–4180. [Google Scholar] [CrossRef]
- Hay, A.; Tsiantis, M. The genetic basis for differences in leaf form between Arabidopsis thaliana and its wild relative Cardamine hirsuta. Nat. Genet. 2006, 38, 942–947. [Google Scholar] [CrossRef]
- Hay, A.; Jackson, D.; Ori, N.; Hake, S. Analysis of the competence to respond to KNOTTED1 activity in Arabidopsis leaves using a steroid induction system. Plant Physiol. 2003, 131, 1671–1680. [Google Scholar] [CrossRef]
- Groover, A.T.; Mansfield, S.D.; DiFazio, S.P.; Dupper, G.; Fontana, J.R.; Millar, R.; Wang, Y. The Populus homeobox gene ARBORKNOX1 reveals overlapping mechanisms regulating the shoot apical meristem and the vascular cambium. Plant Mol. Biol. 2006, 61, 917–932. [Google Scholar] [CrossRef]
- Yan, F.; Hu, G.; Ren, Z.; Deng, W.; Li, Z. Ectopic expression a tomato KNOX Gene Tkn4 affects the formation and the differentiation of meristems and vasculature. Plant Mol. Biol. 2015, 89, 589–605. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Song, X.; Zhou, H.; Wei, K.; Jiang, C.; Wang, J.; Cao, Y.; Tang, F.; Zhao, S.; Lu, M.Z. KNAT2/6b, a class I KNOX gene, impedes xylem differentiation by regulating NAC domain transcription factors in poplar. New Phytol. 2020, 225, 1531–1544. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Li, B.; Liu, J.; Guo, Z.; Liu, Y.; Li, Y.; Shen, W.H.; Huang, Y.; Huang, H.; Zhang, Y.; et al. Transcription factors AS1 and AS2 interact with LHP1 to repress KNOX genes in Arabidopsis. J. Integr. Plant Biol. 2016, 58, 959–970. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Testone, G.; Condello, E.; Verde, I.; Nicolodi, C.; Caboni, E.; Dettori, M.T.; Vendramin, E.; Bruno, L.; Bitonti, M.B.; Mele, G.; et al. The peach (Prunus persica L. Batsch) genome harbours 10 KNOX genes, which are differentially expressed in stem development, and the class 1 KNOPE1 regulates elongation and lignification during primary growth. J. Exp. Botany 2012, 63, 5417–5435. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, H.; Zhu, D.; Gao, Y.; Yan, H.; Xiang, Y. Genome-wide analysis of VQ motif-containing proteins in Moso bamboo (Phyllostachys edulis). Planta 2017, 246, 165–181. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Shi, T.; Iqbal, S.; Ayaz, A.; Bai, Y.; Pan, Z.; Ni, X.; Hayat, F.; Saqib Bilal, M.; Khuram Razzaq, M.; Gao, Z. Analyzing Differentially Expressed Genes and Pathways Associated with Pistil Abortion in Japanese Apricot via RNA-Seq. Genes 2020, 11, 1079. [Google Scholar] [CrossRef]
- Iqbal, S.; Ni, X.; Bilal, M.S.; Shi, T.; Khalil-Ur-Rehman, M.; Zhenpeng, P.; Jie, G.; Usman, M.; Gao, Z. Identification and expression profiling of sugar transporter genes during sugar accumulation at different stages of fruit development in apricot. Gene 2020, 742, 144584. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, S.; Bai, Y.; Hayat, F.; Coulibaly, D.; Khalil-Ur-Rehman, M.; Shi, T.; Gao, Z. Comprehensive transcriptomic and metabolomic analysis revealed distinct flavonoid biosynthesis regulation during abnormal pistil development in Japanese apricot. Genomics 2022, 114, 110451. [Google Scholar] [CrossRef] [PubMed]
- Hamant, O.; Pautot, V. Plant development: A TALE story. Curr. Biol. 2010, 333, 371–381. [Google Scholar] [CrossRef]
- Ezura, K.; Nakamura, A.; Mitsuda, N. Genome-wide characterization of the TALE homeodomain family and the KNOX-BLH interaction network in tomato. Plant Mol. Biol. 2022, 109, 799–821. [Google Scholar] [CrossRef]
- Cheng, X.; Li, M.; Abdullah, M.; Li, G.; Zhang, J.; Manzoor, M.A.; Wang, H.; Jin, Q.; Jiang, T.; Cai, Y.; et al. In Silico Genome-Wide Analysis of the Pear (Pyrus bretschneideri) KNOX Family and the Functional Characterization of PbKNOX1, an Arabidopsis BREVIPEDICELLUS Orthologue Gene, Involved in Cell Wall and Lignin Biosynthesis. Front. Genet. 2019, 10, 632. [Google Scholar] [CrossRef]
- Tan, F.Q.; Wang, W.; Li, J.; Lu, Y.; Zhu, B.; Hu, F.; Li, Q.; Zhao, Y.; Zhou, D.X. A coiled-coil protein associates Polycomb Repressive Complex 2 with KNOX/BELL transcription factors to maintain silencing of cell differentiation-promoting genes in the shoot apex. Plant Cell 2022, 34, 2969–2988. [Google Scholar] [CrossRef]
- Bueno, N.; Alvarez, J.M.; Ordas, R.J. Characterization of the KNOTTED1-LIKE HOMEOBOX (KNOX) gene family in Pinus pinaster Ait. Plant Sci. 2020, 301, 110691. [Google Scholar] [CrossRef]
- Hake, S.; Smith, H.M.; Holtan, H.; Magnani, E.; Mele, G.; Ramirez, J. The role of knox genes in plant development. Annu. Rev. Cell Dev. Biol. 2004, 20, 125–151. [Google Scholar] [CrossRef]
- Song, X.; Zhao, Y.; Wang, J.; Lu, M.Z. The transcription factor KNAT2/6b mediates changes in plant architecture in response to drought via down-regulating GA20ox1 in Populus alba × P. glandulosa. J. Exp. Bot. 2021, 72, 5625–5637. [Google Scholar] [CrossRef]
- Venglat, S.P.; Dumonceaux, T.; Rozwadowski, K.; Parnell, L.; Babic, V.; Keller, W.; Martienssen, R.; Selvaraj, G.; Datla, R. The homeobox gene BREVIPEDICELLUS is a key regulator of inflorescence architecture in Arabidopsis. Proc. Natl. Acad. Sci. USA 2002, 99, 4730–4735. [Google Scholar] [CrossRef]
- Mele, G.; Ori, N.; Sato, Y.; Hake, S. The knotted1-like homeobox gene BREVIPEDICELLUS regulates cell differentiation by modulating metabolic pathways. Genes Dev. 2003, 17, 2088–2093. [Google Scholar] [CrossRef]
- Shi, C.-L.; Stenvik, G.-E.; Vie, A.K.; Bones, A.M.; Pautot, V.; Proveniers, M.; Aalen, R.B.; Butenko, M.A. Arabidopsis Class I KNOTTED-like Homeobox Proteins Act Downstream in the IDA-HAE/HSL2 Floral Abscission Signaling Pathway. Plant Cell 2011, 23, 2553–2567. [Google Scholar] [CrossRef] [PubMed]
- Bueno, N.; Cuesta, C.; Centeno, M.L.; Ordás, R.J.; Alvarez, J.M. In Vitro Plant Regeneration in Conifers: The Role of WOX and KNOX Gene Families. Genes 2021, 12, 438. [Google Scholar] [CrossRef] [PubMed]
- Keren-Keiserman, A.; Shtern, A.; Levy, M.; Chalupowicz, D.; Furumizu, C.; Alvarez, J.P.; Amsalem, Z.; Arazi, T.; Alkalai-Tuvia, S.; Efroni, I.; et al. CLASS-II KNOX genes coordinate spatial and temporal ripening in tomato. Plant Physiol. 2022, 190, 657–668. [Google Scholar] [CrossRef] [PubMed]
- Ragni, L.; Belles-Boix, E.; Gunl, M.; Pautot, V. Interaction of KNAT6 and KNAT2 with BREVIPEDICELLUS and PENNYWISE in Arabidopsis inflorescences. Plant Cell 2008, 20, 888–900. [Google Scholar] [CrossRef] [PubMed]
- Butenko, M.A.; Shi, C.L.; Aalen, R.B. KNAT1, KNAT2 and KNAT6 act downstream in the IDA-HAE/HSL2 signaling pathway to regulate floral organ abscission. Plant Signal. Behav. 2012, 7, 135–138. [Google Scholar] [CrossRef]
- Long, J.A.; Moan, E.I.; Medford, J.I.; Barton, M.K. A member of the KNOTTED class of homeodomain proteins encoded by the STM gene of Arabidopsis. Nature 1996, 379, 66–69. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, K.; Hake, S. Diverse functions of KNOX transcription factors in the diploid body plan of plants. Curr. Opin. Plant Biol. 2015, 27, 91–96. [Google Scholar] [CrossRef]
- Serikawa, K.A.; Martinez-Laborda, A.; Zambryski, P. Three knotted1-like homeobox genes in Arabidopsis. Plant Mol. Biol. 1996, 32, 673–683. [Google Scholar] [CrossRef]
- Meng, L.; Liu, X.; He, C.; Xu, B.; Li, Y.; Hu, Y. Functional divergence and adaptive selection of KNOX gene family in plants. Open Life Sci. 2020, 15, 346–363. [Google Scholar] [CrossRef] [PubMed]
- Scofield, S.; Murray, J.A. KNOX gene function in plant stem cell niches. Plant Mol. Biol. 2006, 60, 929–946. [Google Scholar] [CrossRef]
- Reiser, L.; Sánchez-Baracaldo, P.; Hake, S. Knots in the family tree evolutionary relationships and functions of KNOX homeobox genes. Plant Mol. Biol. 2000, 42, 151–166. [Google Scholar] [CrossRef] [PubMed]
- Hamant, O.; Nogue, F.; Belles-Boix, E.; Jublot, D.; Grandjean, O.; Traas, J.; Pautot, V. The KNAT2 homeodomain protein interacts with ethylene and cytokinin signaling. Plant Physiol. 2002, 130, 657–665. [Google Scholar] [CrossRef]
- Vogel, S. Leaves in the lowest and highest winds: Temperature, force and shape. N. Phytol. 2009, 183, 13–26. [Google Scholar] [CrossRef]
- Hay, A.; Tsiantis, M. KNOX genes: Versatile regulators of plant development and diversity. Development 2010, 137, 3153–3165. [Google Scholar] [CrossRef] [PubMed]
- Rogers, L.A.; Dubos, C.; Cullis, I.F.; Surman, C.; Poole, M.; Willment, J.; Mansfield, S.D.; Campbell, M.M. Light, the circadian clock, and sugar perception in the control of lignin biosynthesis. J. Exp. Bot. 2005, 56, 1651–1663. [Google Scholar] [CrossRef] [PubMed]
- Townsley, B.T.; Sinha, N.R.; Kang, J. KNOX1 genes regulate lignin deposition and composition in monocots and dicots. Front. Plant Sci. 2013, 4, 121. [Google Scholar] [CrossRef]
- Khan, M.; Tabb, P.; Hepworth, S.R. BLADE-ON-PETIOLE1 and 2 regulate Arabidopsis inflorescence architecture in conjunction with homeobox genes KNAT6 and ATH1. Plant Signal Behav. 2012, 7, 788–792. [Google Scholar] [CrossRef] [PubMed]
- Mauriat, M.; Moritz, T. Analyses of GA20ox- and GID1-over-expressing aspen suggest that gibberellins play two distinct roles in wood formation. Plant J. 2009, 58, 989–1003. [Google Scholar] [CrossRef]
- Dong, Y.; Ye, X.; Xiong, A.; Zhu, N.; Jiang, L.; Qu, S. The regulatory role of gibberellin related genes DKGA2ox1 and MIR171f_3 in persimmon dwarfism. Plant Sci. 2021, 310, 110958. [Google Scholar] [CrossRef]
- Nakayama, H.; Nakayama, N.; Seiki, S.; Kojima, M.; Sakakibara, H.; Sinha, N.; Kimura, S. Regulation of the KNOX-GA gene module induces heterophyllic alteration in North American lake cress. Plant Cell 2014, 26, 4733–4748. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Banerjee, A.K.; Hannapel, D.J. The tandem complex of BEL and KNOX partners is required for transcriptional repression of ga20ox1. Plant J. 2004, 38, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Bolduc, N.; Hake, S. The maize transcription factor KNOTTED1 directly regulates the gibberellin catabolism gene ga2ox1. Plant Cell 2009, 21, 1647–1658. [Google Scholar] [CrossRef]
- Lee, H.G.; Choi, Y.R.; Seo, P.J. Increased STM expression is associated with drought tolerance in Arabidopsis. J. Plant Physiol. 2016, 201, 79–84. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Gene ID | Protein ID | Chromosomal Position | gDNA (bp) | CDS (bp) | Protein | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Length (aa) | MW (kDa) | pI | GRAVY | Subcellular Localization Prediction | ||||||
| PmKNOX1 | LOC103344141 | XP_008245991.1 | LG1: 23,453,520–23,456,592 | 3073 | 1002 | 333 | 37.683 | 5.06 | −0.67 | nucleus |
| PmKNOX2 | LOC103319886 | XP_016647641.1 | LG2: 6,437,754–6,441,461 | 3708 | 1293 | 430 | undefined | undefined | −0.844 | nucleus |
| PmKNOX3 | LOC103320515 | XP_008220425.1 | LG2: 11,142,302–11,146,201 | 3900 | 1161 | 386 | 44.011 | 6.13 | −1.011 | nucleus |
| PmKNOX4 | LOC103320960 | XP_008220923.1 | LG2: 13,532,590–13,533,609 | 1020 | 489 | 162 | 17.988 | 4.29 | −0.583 | nucleus |
| PmKNOX5 | LOC103321797 | NP_001280190.1 | LG2: 18,638,472–18,644,136 | 5665 | 1062 | 353 | 40.407 | 5.09 | −0.727 | nucleus |
| PmKNOX6 | LOC103324440 | XP_008224716.1 | LG3: 521,722–527,036 | 5315 | 1152 | 383 | 42.812 | 6.3 | −0.738 | nucleus |
| PmKNOX7 | LOC103329326 | XP_008230002.1 | LG4: 17,650,120–17,652,180 | 2061 | 990 | 329 | 36.901 | 6.53 | −0.63 | nucleus |
| PmKNOX8 | LOC103337382 | XP_008238758.1 | LG7: 7,825,616–7,830,499 | 4884 | 877 | 291 | 33.059 | 6.35 | −0.73 | nucleus |
| PmKNOX9 | LOC103338667 | XP_008240118.1 | LG7: 15,732,193–15,733,759 | 1567 | 423 | 140 | 15.732 | 4.81 | −0.759 | nucleus |
| PmKNOX10 | LOC103341196 | XP_008242898.1 | LG8: 16,386,362–16,389,098 | 2737 | 1077 | 358 | 40.833 | 5.61 | −0.795 | nucleus |
| PmKNOX11 | LOC103343109 | XP_008245008.1 | Unplaced Scaffold: 110,247–116,106 | 5860 | 1066 | 354 | 39.674 | 5.45 | −0.667 | nucleus |
| Protein Name | α Helix | Extended Strand | β Turn | Random Coil |
|---|---|---|---|---|
| PmKNOX1 | 149 (44.74%) | 19 (5.71%) | 13 (3.90%) | 152 (45.65%) |
| PmKNOX2 | 178 (41.40%) | 24 (5.58%) | 7 (1.63%) | 221 (51.40%) |
| PmKNOX3 | 155 (40.16%) | 30 (7.77%) | 12 (3.11%) | 189 (48.96%) |
| PmKNOX4 | 95 (58.64%) | 14 (8.64%) | 5 (3.09%) | 48 (29.63%) |
| PmKNOX5 | 154 (43.63%) | 11 (3.12%) | 14 (3.97%) | 174 (49.29%) |
| PmKNOX6 | 147 (38.38%) | 41 (10.70%) | 20 (5.22%) | 175 (45.69%) |
| PmKNOX7 | 176 (53.50%) | 14 (4.26%) | 13 (3.95%) | 126 (38.30%) |
| PmKNOX8 | 168 (57.73%) | 8 (2.75%) | 10 (3.44%) | 105 (36.08%) |
| PmKNOX9 | 98 (68.53%) | 2 (1.40%) | 1 (0.70%) | 42 (29.37%) |
| PmKNOX10 | 191 (53.35%) | 11 (3.07%) | 7 (1.96%) | 149 (41.62%) |
| PmKNOX11 | 180 (50.85%) | 11 (3.11%) | 24 (6.78%) | 139 (39.27%) |
| Sample Name | Leaf Area of Basal Leaves/cm2 | Leaf Area of Leaves near the Base/cm2 |
|---|---|---|
| WT | 1.22 ± 0.35 | 0.82 ± 0.08 |
| HE-PmKNAT2a | 0.65 ± 0.31 | 0.39 ± 0.06 |
| HE-PmKNAT2b | 1.14 ± 0.31 | 0.59 ± 0.20 |
| Sample Name | 0~1/cm | 1~2/cm | 2~3/cm |
|---|---|---|---|
| WT | 2.24 ± 0.08 | 2.06 ± 0.13 | 1.43 ± 0.09 |
| HE-PmKNAT2a | 1.24 ± 0.1 | 1.38 ± 0.22 | 1.24 ± 0.08 |
| HE-PmKNAT2b | 2.34 ± 0.24 | 1.46 ± 0.18 | 1.37 ± 0.14 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bai, Y.; Shi, T.; Huang, X.; Zhou, P.; Ouma, K.O.; Ni, Z.; Gao, F.; Tan, W.; Ma, C.; Ma, Y.; et al. Genome-Wide Identification of the KNOX Gene Family in Japanese Apricot (Prunus mume Sieb. et Zucc.) and Functional Characterization of PmKNAT2 Genes. Genes 2023, 14, 939. https://doi.org/10.3390/genes14040939
Bai Y, Shi T, Huang X, Zhou P, Ouma KO, Ni Z, Gao F, Tan W, Ma C, Ma Y, et al. Genome-Wide Identification of the KNOX Gene Family in Japanese Apricot (Prunus mume Sieb. et Zucc.) and Functional Characterization of PmKNAT2 Genes. Genes. 2023; 14(4):939. https://doi.org/10.3390/genes14040939
Chicago/Turabian StyleBai, Yang, Ting Shi, Xiao Huang, Pengyu Zhou, Kenneth Omondi Ouma, Zhaojun Ni, Feng Gao, Wei Tan, Chengdong Ma, Yufan Ma, and et al. 2023. "Genome-Wide Identification of the KNOX Gene Family in Japanese Apricot (Prunus mume Sieb. et Zucc.) and Functional Characterization of PmKNAT2 Genes" Genes 14, no. 4: 939. https://doi.org/10.3390/genes14040939