
The Complete Chloroplast Genomes of Gynostemma Reveal the Phylogenetic Relationships of Species within the Genus
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material, DNA Extraction, and Sequencing
2.2. Chloroplast Genome Assembly and Annotation
2.3. Codon Usage Bias and RNA Editing Sites
2.4. Repeat Analysis
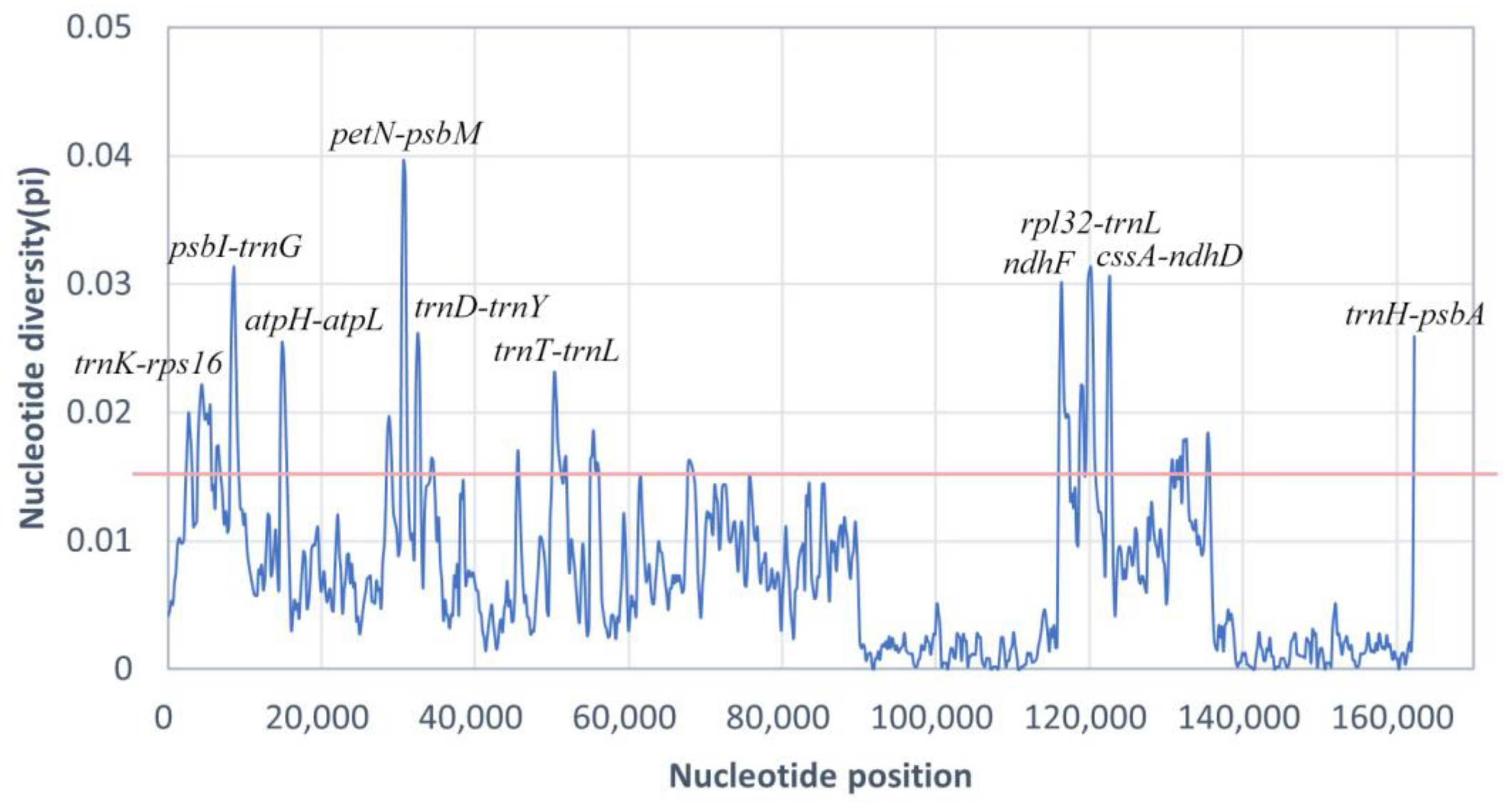
2.5. Comparative Analysis, and Identification of Polymorphic Loci
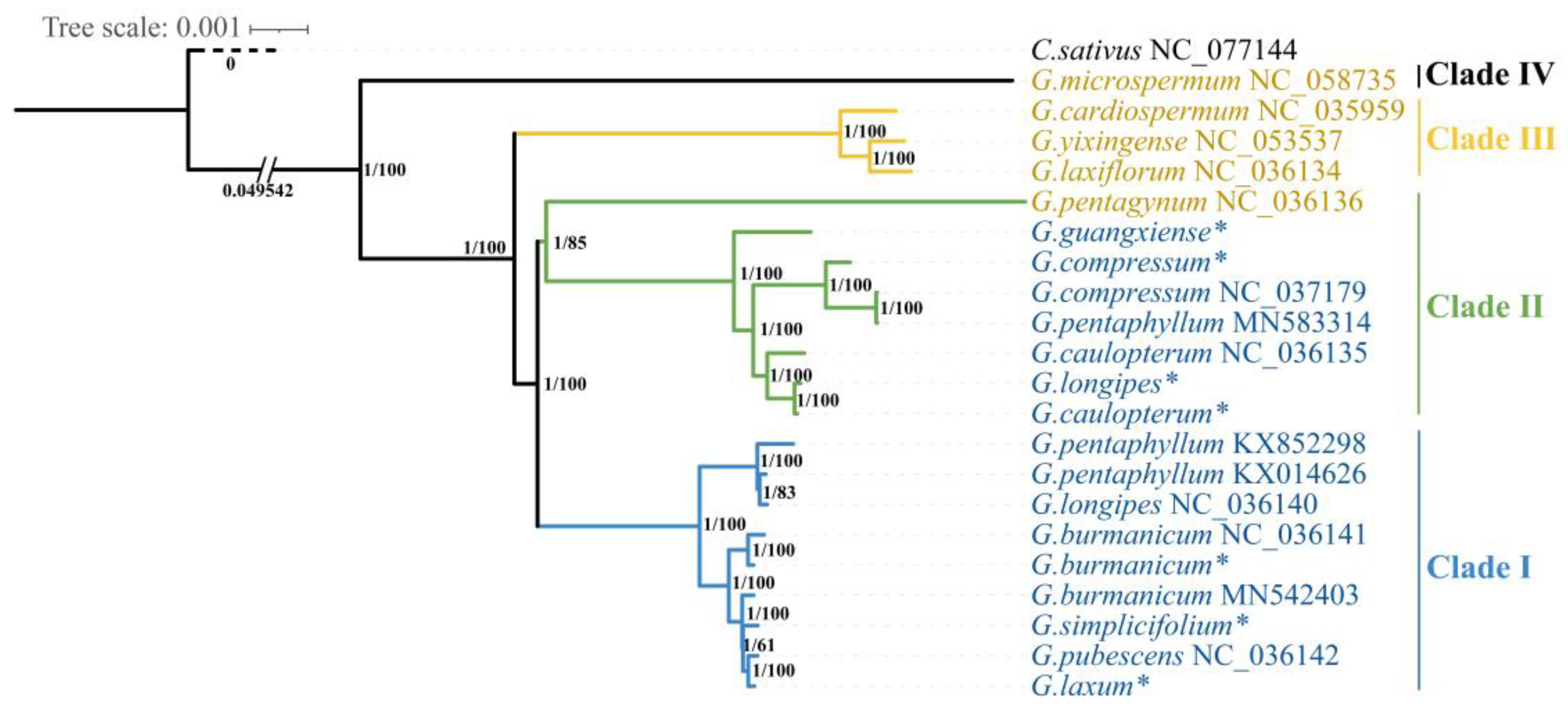
2.6. Phylogenetic Analysis
3. Results
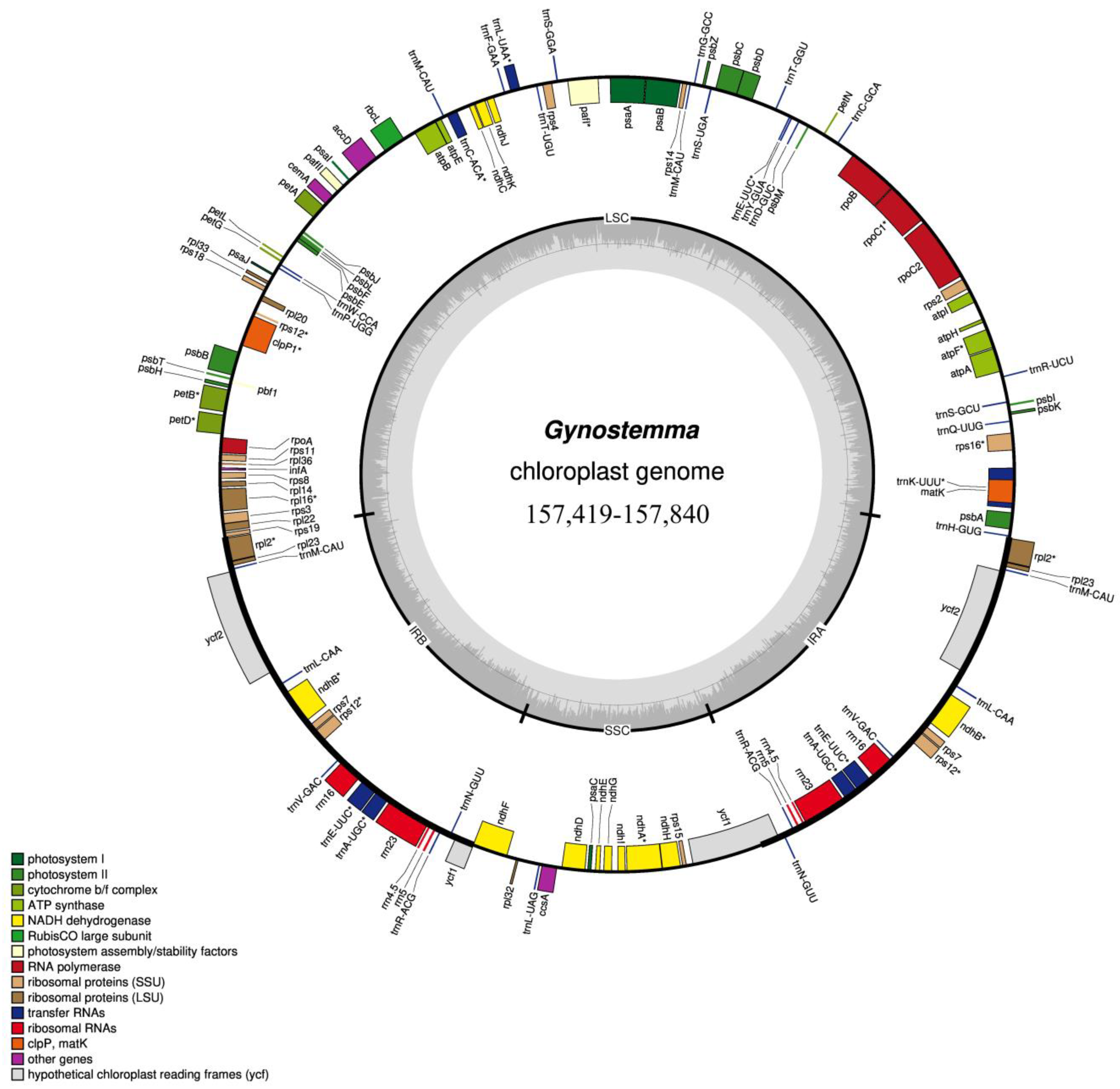
3.1. Characterization of the Chloroplast Genomes of the Gynostemma Genus
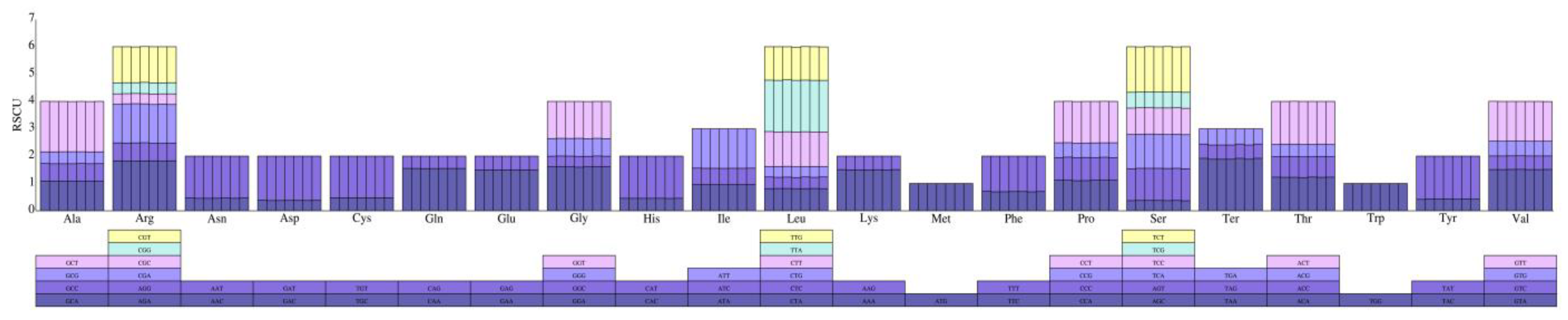
3.2. Analysis of Codon Usage Bias and Prediction of RNA Editing Sites
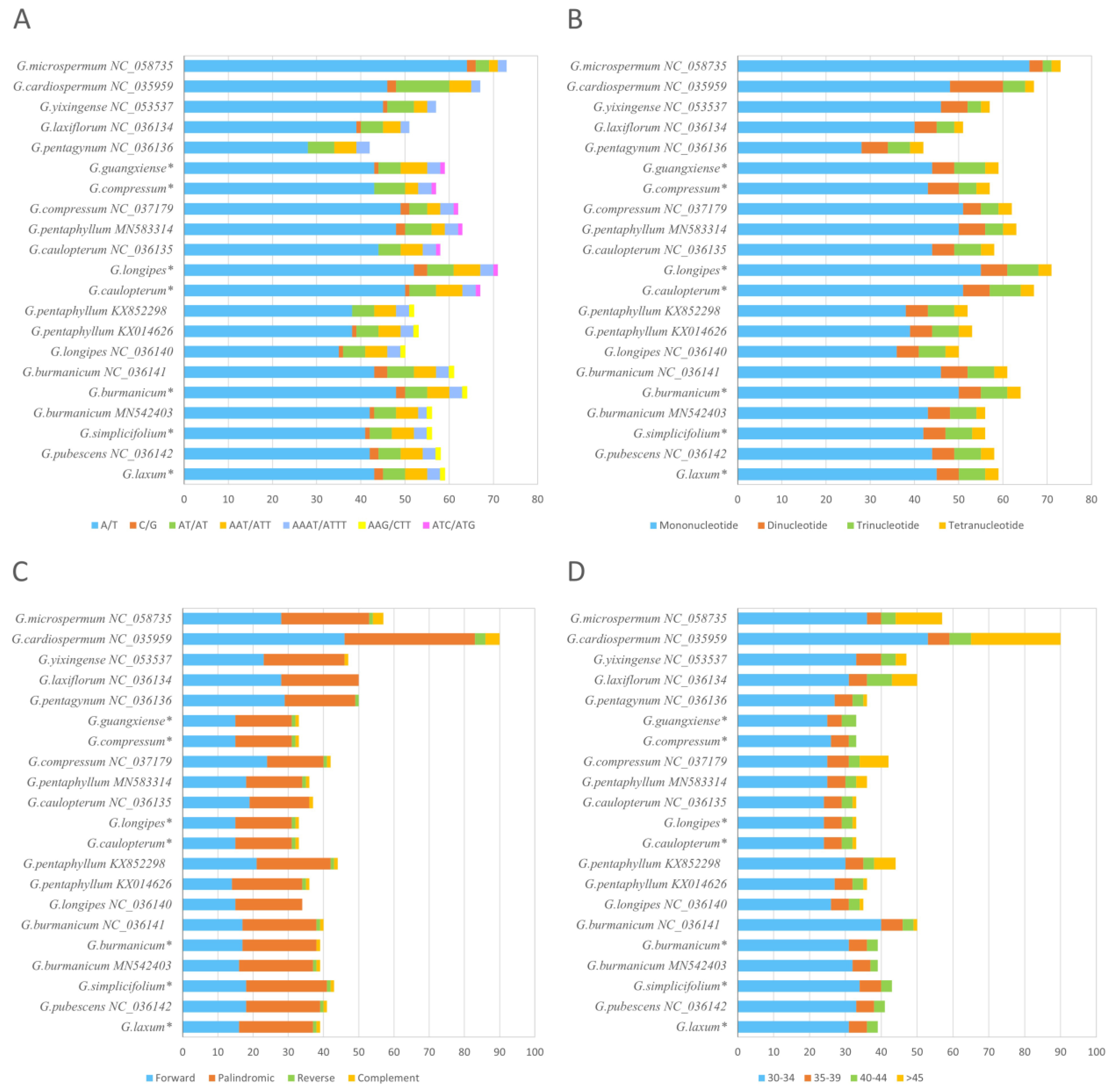
3.3. Repeat Analysis
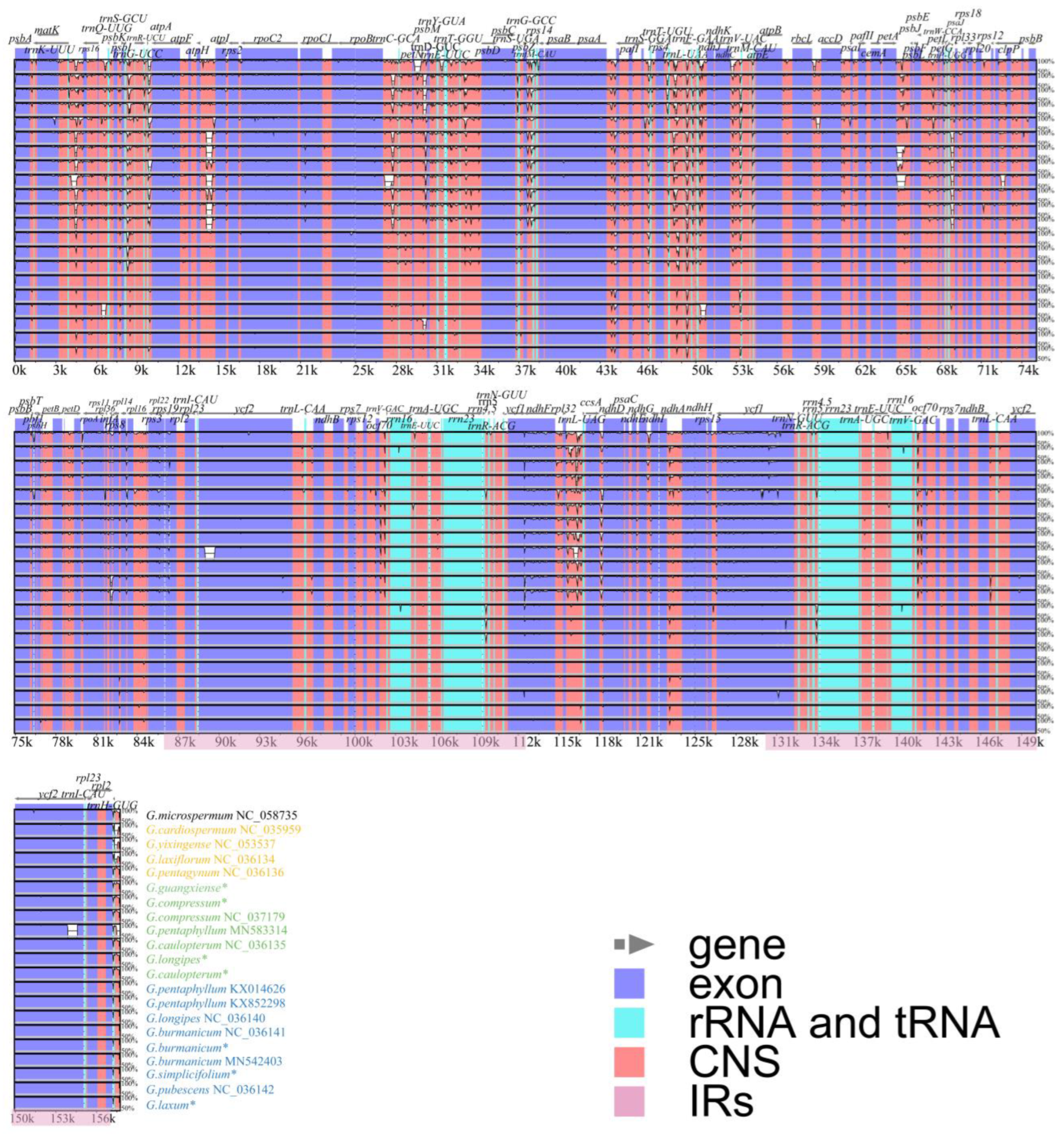
3.4. Comparative Analysis and Selection Pressure Analysis
3.5. IR Region Contraction and Expansion
3.6. Phylogenetic Relationships among the Gynostemma Species
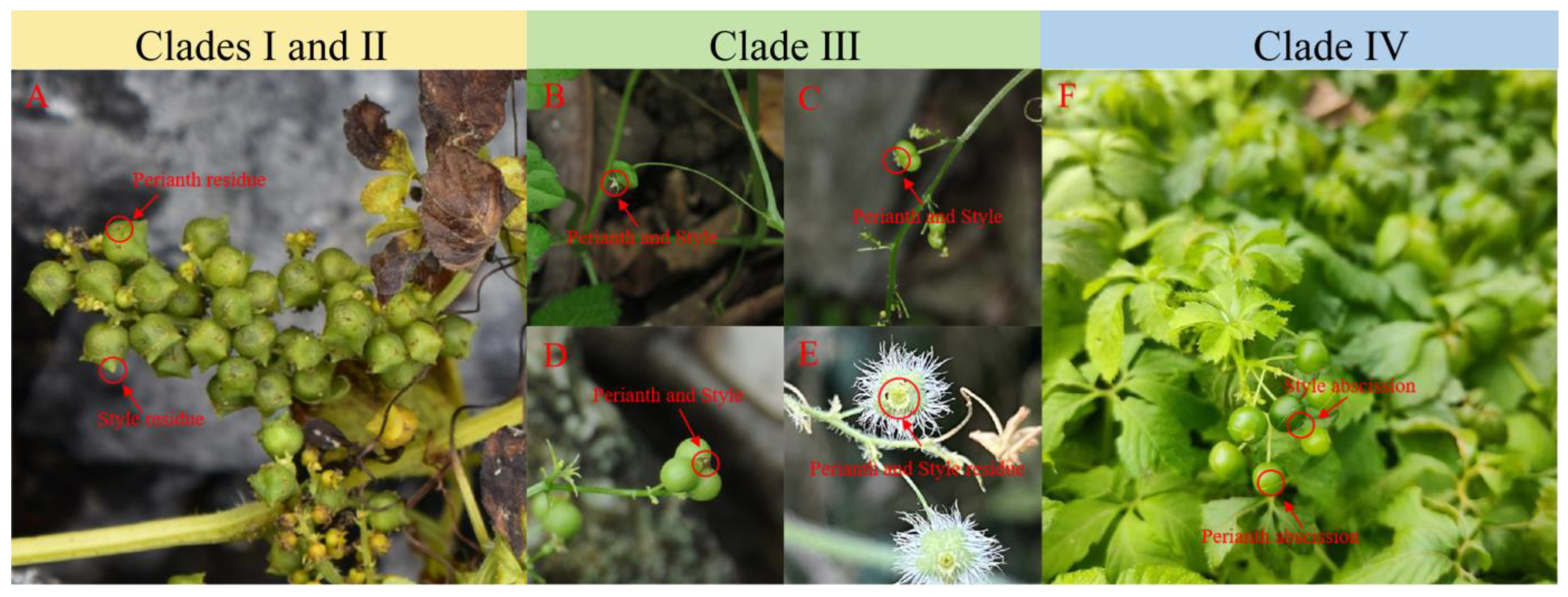
3.7. Morphological Analysis
4. Discussion
4.1. General Characteristics of the Chloroplast Genomes of the Genus Gynostemma
4.2. Analysis of Codon Usage Bias and Prediction of RNA Editing Sites
4.3. Phylogenetic Relationships
4.4. Identification of Suitable Polymorphic Loci at the Subgenus and Species Levels
4.5. Association of Fruit Shapes with the Phylogenetic Tree

5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kao, T.; Huang, S.; Inbaraj, B.S.; Chen, B. Determination of flavonoids and saponins in Gynostemma pentaphyllum (Thunb.) Makino by liquid chromatography–mass spectrometry. Anal. Chim. Acta 2008, 626, 200–211. [Google Scholar] [CrossRef] [PubMed]
- Gou, S.H.; Liu, B.J.; Han, X.F.; Wang, L.; Zhong, C.; Liang, S.; Liu, H.; Qiang, Y.; Zhang, Y.; Ni, J.M. Anti-atherosclerotic effect of Fermentum Rubrum and Gynostemma pentaphyllum mixture in high-fat emulsion-and vitamin D3-induced atherosclerotic rats. J. Chin. Med. Assoc. 2018, 81, 398–408. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhao, X.; Liu, X.; Lu, W.; Jia, S.; Hong, T.; Li, R.; Zhang, H.; Peng, L.; Zhan, X. Anti-diabetic activity evaluation of a polysaccharide extracted from Gynostemma pentaphyllum. Int. J. Biol. Macromol. 2019, 126, 209–214. [Google Scholar] [CrossRef]
- Babich, O.; Sukhikh, S.; Prosekov, A.; Asyakina, L.; Ivanova, S. Medicinal plants to strengthen immunity during a pandemic. Pharmaceuticals 2020, 13, 313. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.H.; Ha, T.K.Q.; Yang, J.L.; Pham, H.T.T.; Oh, W.K. Triterpenoids from the genus Gynostemma: Chemistry and pharmacological activities. J. Ethnopharmacol. 2021, 268, 113574. [Google Scholar] [CrossRef]
- Zhang, H.X.; Wang, Z.Z.; Du, Z.Z. Sensory-guided isolation and identification of new sweet-tasting dammarane-type saponins from Jiaogulan (Gynostemma pentaphyllum) herbal tea. Food Chem. 2022, 388, 132981. [Google Scholar] [CrossRef]
- Jiang, L.Y.; Guo, Z.G.; Wang, C.; Zhao, G.F. ITS Sequence analysis of Gynostemma pentaphyllum from different habitats in China. Chin. Tradit. Herb. Drugs 2009, 40, 1123–1127. [Google Scholar]
- Chen, S.; Lu, A.; Charles, J. Flora of China; Missouri Botanical Garden Press: Beijing, China, 2011; Volume 19, pp. 11–15. [Google Scholar]
- Piao, X.-L.; Qian, W. Progressive Studies on Gynostemma pentaphyllum. Lishizhen Med. Mater. Med. Res. 2010, 21, 1758–1760. [Google Scholar]
- Chen, S.K. A classificatory system and geographical distribution of the genus Gynostemma, B.L. (Cucurbitaceae). Acta Phytotaxon. 1995, 33, 403–410. [Google Scholar]
- Qin, S.S.; Li, H.T.; Wang, Z.Y.; Cui, Z.H.; Yu, L.Y. Analysis phylogenetic relationship of Gynostemma (Cucurbitaceae). China J. Chin. Mater. Med. 2015, 40, 1681–1687. [Google Scholar]
- Ranade, S.; Gupta, V.S.; Aggarwal, R.K. Genetic diversity and phylogenetic relationship as revealed by inter simple sequence repeat (ISSR) polymorphism in the genus Oryza. Theor. Appl. Genet. 2000, 100, 1311–1320. [Google Scholar] [CrossRef]
- Abid, S.; Mohanan, P.; Kaliraj, L.; Park, J.K.; Ahn, J.C.; Yang, D.C. Development of species-specific chloroplast markers for the authentication of Gynostemma pentaphyllum and their distribution in the Korean peninsula. Fitoterapia 2019, 138, 104295. [Google Scholar] [CrossRef] [PubMed]
- Plunkett, G.M.; Downie, S.R. Expansion and contraction of the chloroplast inverted repeat in Apiaceae subfamily Apioideae. Syst. Bot. 2000, 25, 648–667. [Google Scholar] [CrossRef]
- Dobrogojski, J.; Adamiec, M.; Luciński, R. The chloroplast genome: A review. Acta Physiol. Plant. 2020, 42, 98. [Google Scholar] [CrossRef]
- Palmer, J.D.; Jansen, R.K.; Michaels, H.J.; Chase, M.W.; Manhart, J.R. Chloroplast DNA variation and plant phylogeny. Ann. Mo. Bot. Gard. 1988, 75, 1180–1206. [Google Scholar] [CrossRef]
- Wang, L.; Lu, G.; Liu, H.; Huang, L.; Jiang, W.; Li, P.; Lu, X. The complete chloroplast genome sequence of Gynostemma yixingense and comparative analysis with congeneric species. Genet. Mol. Biol. 2020, 43, e20200092. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, X.; Zhang, H.; Zhao, Y.; Ju, M.; Zhao, G. Characterization of the complete chloroplast genome sequence of Gynostemma microspermum (Cucurbitaceae). Mitochondrial DNA Part B 2022, 7, 32–34. [Google Scholar] [CrossRef]
- Zhang, X.; Li, H.; Zhou, T.; Yang, Y.; Zhao, G. Characterization of the complete chloroplast genome sequence of Gynostemma compressum (Cucurbitaceae), an endemic plant in China. Conserv. Genet. Resour. 2018, 10, 141–144. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, X.; Zhou, T.; Chen, X.; Ding, B. Complete chloroplast genome sequence of Gynostemma guangxiense: Genome structure, codon usage bias, and phylogenetic relationships in Gynostemma (Cucurbitaceae). Braz. J. Bot. 2023, 1–15. [Google Scholar] [CrossRef]
- Zhang, X.; Zhou, T.; Kanwal, N.; Zhao, Y.; Bai, G.; Zhao, G. Completion of eight Gynostemma BL.(Cucurbitaceae) chloroplast genomes: Characterization, comparative analysis, and phylogenetic relationships. Front. Plant Sci. 2017, 8, 1583. [Google Scholar] [CrossRef]
- Ewels, P.; Magnusson, M.; Lundin, S.; Käller, M. MultiQC: Summarize analysis results for multiple tools and samples in a single report. Bioinformatics 2016, 32, 3047–3048. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.-J.; Yu, W.-B.; Yang, J.B.; Song, Y.; DePamphilis, C.W.; Yi, T.S.; Li, D.Z. GetOrganelle: A fast and versatile toolkit for accurate de novo assembly of organelle genomes. Genome Biol. 2020, 21, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Luo, R.; Liu, B.; Xie, Y.; Li, Z.; Huang, W.; Yuan, J.; He, G.; Chen, Y.; Pan, Q.; Liu, Y. SOAPdenovo2: An empirically improved memory-efficient short-read de novo assembler. Gigascience 2012, 1, 18. [Google Scholar] [CrossRef] [PubMed]
- Kent, W.J. BLAT—The BLAST-like alignment tool. Genome Res. 2002, 12, 656–664. [Google Scholar] [CrossRef] [PubMed]
- Wick, R.R.; Schultz, M.B.; Zobel, J.; Holt, K.E. Bandage: Interactive visualization of de novo genome assemblies. Bioinformatics 2015, 31, 3350–3352. [Google Scholar] [CrossRef]
- Tillich, M.; Lehwark, P.; Pellizzer, T.; Ulbricht-Jones, E.S.; Fischer, A.; Bock, R.; Greiner, S. GeSeq–versatile and accurate annotation of organelle genomes. Nucleic Acids Res. 2017, 45, W6–W11. [Google Scholar] [CrossRef]
- Finn, R.D.; Clements, J.; Eddy, S.R. HMMER web server: Interactive sequence similarity searching. Nucleic Acids Res. 2011, 39, W29–W37. [Google Scholar] [CrossRef]
- Laslett, D.; Canback, B. ARAGORN, a program to detect tRNA genes and tmRNA genes in nucleotide sequences. Nucleic Acids Res. 2004, 32, 11–16. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef]
- Somaratne, Y.; Guan, D.-L.; Wang, W.-Q.; Zhao, L.; Xu, S.-Q. The complete chloroplast genomes of two Lespedeza species: Insights into codon usage bias, RNA editing sites, and phylogenetic relationships in Desmodieae (Fabaceae: Papilionoideae). Plants 2019, 9, 51. [Google Scholar] [CrossRef]
- Beier, S.; Thiel, T.; Münch, T.; Scholz, U.; Mascher, M. MISA-web: A web server for microsatellite prediction. Bioinformatics 2017, 33, 2583–2585. [Google Scholar] [CrossRef] [PubMed]
- Thiel, T.; Michalek, W.; Varshney, R.; Graner, A. Exploiting EST databases for the development and characterization of gene-derived SSR-markers in barley (Hordeum vulgare L.). Theor. Appl. Genet. 2003, 106, 411–422. [Google Scholar] [CrossRef] [PubMed]
- Kurtz, S.; Choudhuri, J.V.; Ohlebusch, E.; Schleiermacher, C.; Stoye, J.; Giegerich, R. REPuter: The manifold applications of repeat analysis on a genomic scale. Nucleic Acids Res. 2001, 29, 4633–4642. [Google Scholar] [CrossRef] [PubMed]
- Thode, V.A.; Lohmann, L.G. Comparative chloroplast genomics at low taxonomic levels: A case study using Amphilophium (Bignonieae, Bignoniaceae). Front. Plant Sci. 2019, 10, 796. [Google Scholar] [CrossRef]
- Frazer, K.A.; Pachter, L.; Poliakov, A.; Rubin, E.M.; Dubchak, I. VISTA: Computational tools for comparative genomics. Nucleic Acids Res. 2004, 32, W273–W279. [Google Scholar] [CrossRef]
- Brudno, M.; Malde, S.; Poliakov, A.; Do, C.B.; Couronne, O.; Dubchak, I.; Batzoglou, S. Glocal alignment: Finding rearrangements during alignment. Bioinformatics 2003, 19, i54–i62. [Google Scholar] [CrossRef]
- Amiryousefi, A.; Hyvönen, J.; Poczai, P. IRscope: An online program to visualize the junction sites of chloroplast genomes. Bioinformatics 2018, 34, 3030–3031. [Google Scholar] [CrossRef]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef]
- Katoh, K.; Misawa, K.; Kuma, K.i.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef]
- Posada, D.; Crandall, K.A. MODELTEST: Testing the model of DNA substitution. Bioinformatics 1998, 14, 817–818. [Google Scholar] [CrossRef]
- Wilgenbusch, J.C.; Swofford, D. Inferring evolutionary trees with PAUP. Curr. Protoc. Bioinform. 2003. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Lian, C.; Yang, H.; Lan, J.; Zhang, X.; Zhang, F.; Yang, J.; Chen, S. Comparative analysis of chloroplast genomes reveals phylogenetic relationships and intraspecific variation in the medicinal plant Isodon rubescens. PLoS ONE 2022, 17, e0266546. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.X.; Qin, D.H. A New Species of Gynostemma from GuangXi. Acta Bot. Yunnanica 1988, 10, 195–196. [Google Scholar]
- De, P.Y.; Ying, X.Y.; You, H.B.; Ying, Y.L.; Yan, H.X. Gynostemma caulopterum, A Newly recorded Species of Cucurbitaceae from Guangxi, China. Mod. Chin. Med. 2013, 15, 1059–1159. [Google Scholar]
- Gong, X.; Ji, M.; Xu, J.; Zhang, C.; Li, M. Hypoglycemic effects of bioactive ingredients from medicine food homology and medicinal health food species used in China. Crit. Rev. Food Sci. Nutr. 2020, 60, 2303–2326. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Nguyen, V.B.; Dong, J.; Wang, Y.; Park, J.Y.; Lee, S.C.; Yang, T.J. Evolution of the Araliaceae family inferred from complete chloroplast genomes and 45S nrDNAs of 10 Panax-related species. Sci. Rep. 2017, 7, 4917. [Google Scholar] [CrossRef] [PubMed]
- Firetti, F.; Zuntini, A.R.; Gaiarsa, J.W.; Oliveira, R.S.; Lohmann, L.G.; Van Sluys, M.A. Complete chloroplast genome sequences contribute to plant species delimitation: A case study of the Anemopaegma species complex. Am. J. Bot. 2017, 104, 1493–1509. [Google Scholar] [CrossRef]
- Li, E.; Liu, K.; Deng, R.; Gao, Y.; Liu, X.; Dong, W.; Zhang, Z. Insights into the phylogeny and chloroplast genome evolution of Eriocaulon (Eriocaulaceae). BMC Plant Biol. 2023, 23, 32. [Google Scholar] [CrossRef]
- Zhang, P.; Xu, W.; Lu, X.; Wang, L. Analysis of codon usage bias of chloroplast genomes in Gynostemma species. Physiol. Mol. Biol. Plants 2021, 27, 2727–2737. [Google Scholar] [CrossRef]
- Ren, J.; Tian, J.; Jiang, H.; Zhu, X.-X.; Mutie, F.M.; Wanga, V.O.; Ding, S.-X.; Yang, J.-X.; Dong, X.; Chen, L.-L. Comparative and phylogenetic analysis based on the chloroplast genome of coleanthus subtilis (Tratt.) Seidel, a protected rare species of monotypic genus. Front. Plant Sci. 2022, 13, 828467. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, T.; Yukawa, Y.; Miyamoto, T.; Obokata, J.; Sugiura, M. Identification of RNA editing sites in chloroplast transcripts from the maternal and paternal progenitors of tobacco (Nicotiana tabacum): Comparative analysis shows the involvement of distinct trans-factors for ndhB editing. Mol. Biol. Evol. 2003, 20, 1028–1035. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Chen, Z.; He, L.; Feng, Q.; Zhang, H.; Du, G.; Shi, C.; Wang, S. Comparative chloroplast genome analysis of wax gourd (Benincasa hispida) with three Benincaseae species, revealing evolutionary dynamic patterns and phylogenetic implications. Genes 2022, 13, 461. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Yang, C.; Zhao, X.; Chen, S.; Qu, G.-Z. Complete chloroplast genome sequence of Betula platyphylla: Gene organization, RNA editing, and comparative and phylogenetic analyses. BMC Genom. 2018, 19, 950. [Google Scholar] [CrossRef] [PubMed]
- Moore, M.J.; Bell, C.D.; Soltis, P.S.; Soltis, D.E. Using plastid genome-scale data to resolve enigmatic relationships among basal angiosperms. Proc. Natl. Acad. Sci. USA 2007, 104, 19363–19368. [Google Scholar] [CrossRef]
- Zhang, X.; Zhou, T.; Yang, J.; Sun, J.; Ju, M.; Zhao, Y.; Zhao, G. Comparative analyses of chloroplast genomes of Cucurbitaceae species: Lights into selective pressures and phylogenetic relationships. Molecules 2018, 23, 2165. [Google Scholar] [CrossRef]
- Group, A.P.; Chase, M.W.; Christenhusz, M.J.; Fay, M.F.; Byng, J.; Judd, W.; Soltis, D.; Mabberley, D.; Sennikov, A.; Soltis, P. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar] [CrossRef]
- Li, Z.; Liu, Z.; Zhao, P.; Su, H.; Zhao, G. A review on studies of systematic evolution of Gynostemma Bl. Acta Bot. Boreali-Occident. Sin. 2012, 32, 2133–2138. [Google Scholar]
- Sun, H.; Chen, S.K. The Microstructural Features of Seed Surfaces and Its Taxonomic Significance in the Genus Gynostemma. Acta Bot. Yunnanica 1998, 20, 309–311. [Google Scholar]
- Ren, T.; Yang, Y.; Zhou, T.; Liu, Z.-L. Comparative plastid genomes of Primula species: Sequence divergence and phylogenetic relationships. Int. J. Mol. Sci. 2018, 19, 1050. [Google Scholar] [CrossRef]
- Yang, L.; Abduraimov, O.; Tojibaev, K.; Shomurodov, K.; Zhang, Y.-M.; Li, W.-J. Analysis of complete chloroplast genome sequences and insight into the phylogenetic relationships of Ferula L. BMC Genom. 2022, 23, 643. [Google Scholar] [CrossRef] [PubMed]
- Vieira, M.L.C.; Santini, L.; Diniz, A.L.; Munhoz, C.d.F. Microsatellite markers: What they mean and why they are so useful. Genet. Mol. Biol. 2016, 39, 312–328. [Google Scholar] [CrossRef] [PubMed]
- Provan, J.; Powell, W.; Hollingsworth, P.M. Chloroplast microsatellites: New tools for studies in plant ecology and evolution. Trends Ecol. Evol. 2001, 16, 142–147. [Google Scholar] [CrossRef] [PubMed]
- Kalia, R.K.; Rai, M.K.; Kalia, S.; Singh, R.; Dhawan, A. Microsatellite markers: An overview of the recent progress in plants. Euphytica 2011, 177, 309–334. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Functions | Biological Function | Gene List | Number |
|---|---|---|---|
| Photosynthesis | ATP synthase related | atpA atpF atpH atpI atpE atpB | 6 |
| Photosystem I | psaB psaA psaI psaJ psaC | 5 | |
| Photosystem II | psbA psbK psbI psbM psbD psbC psbZ psbJ psbL psbF psbEpsbB psbT psbH | 14 | |
| Cytochrome b/f complex | petN petA petG petL petB * petD * | 6 | |
| NADH dehydrogenase | ndhJ ndhK ndhC ndhB * ndhF ndhD ndhE ndhG ndhI ndhA * ndhH ndhB * | 12 | |
| Photosystem biogenesis factor 1 | pbf1 | 1 | |
| Photosystem I assembly protein | pafI ** pafII | 2 | |
| Self-replication | Ribosomal Structural RNAs | rrn16 rrn23 rrn4.5 rrn5 rrn5 rrn4.5 rrn23 rrn16 | 8 |
| Translation-related gene | trnK-UUU * trnQ-UUG trnS-GCU trnR-UCU trnC-GCA trnD-GUC trnY-GUA trnE-UUC trnT-GGU trnS-UGA trnG-GCC trnfM-CAU trnG-UCC trnS-GGA trnT-UGU trnL-UAA * trnF-GAA trnV-UAC * trnM-CAU trnW-CCA trnP-UGG trnl-CAU trnL-CAA trnV-GAC trnE-UUC * trnA-UGC * trnR-ACG trnN-GUU trnL-UAG trnN-GUU trnR-ACG trnA-UGC * trnE-UUC * trnV-GAC trnL-CAA trnl-CAU trnH-GUG | 37 | |
| Ribosomal Proteins (small subunit) | rps16 * rps2 rps14 rps4 rps18 rps12 * rps12 * rps11 rps8 rps3 rps19 rps7 rps15 rps7 | 14 | |
| Ribosomal Proteins (large subunit) | rpl33 rpl20 rpl36 rpl14 rpl16 * rpl22 rpl2 * rpl23 rpl32 rpl23 rpl2 * | 11 | |
| RNA polymerase | rpoC2 rpoC1 rpoB rpoA | 4 | |
| Other genes | RuBisCO large subunit | rbcL | 1 |
| Translation related | infA | 1 | |
| Acetyl-CoA carboxylase gene | accD | 1 | |
| RNA Splicing | matK | 1 | |
| Carbon metabolism | cemA | 1 | |
| c-type Cytochrome biogenesis | ccsA | 1 | |
| ATP-dependent protease subunit | clpP1 ** | 1 | |
| Unknown | ycf2 orf70 ycf1 ycf1 orf70 ycf2 | 6 |
| Subgenus | Species | Fruit | Persistence | Information Source | |||
|---|---|---|---|---|---|---|---|
| Type | Mature Performance | Shape | Perianth | Style | |||
| Trirostellum [10] | G. microspermum | capsule | split | campanulate | - | Yes | [8] and Substance |
| G. cardiospermum | capsule | split | campanulate | - | Yes | [8] | |
| G. yixingense | capsule | split | campanulate | - | Yes | [8] | |
| G. laxiflorum | capsule | split | campanulate | - | Yes | [8] | |
| G. pentagynum | - | - | 5-angled-oblate | Yes | Yes | [8] and Substance | |
| Gynostemma [10] | G. guangxiense | - | - | 3-angled-oblate | Yes | Yes | [8,45] and Substance |
| G. compressum | - | - | compressed, obtriangular | Yes | Yes | [8] and Substance | |
| G. caulopterum | - | - | compressed globose | Yes | Yes | [8,46] and Substance | |
| G. pentaphyllum | berry | not split | globose | - | - | [8] | |
| G. longipes | berry | not split | globose | - | - | [8] | |
| G. burmanicum | berry | not split | globose | - | - | [8] | |
| G. simplicifolium | berry | not split | globose | - | - | [8] | |
| G. laxum | berry | not split | globose | - | - | [8] | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gan, J.; Li, Y.; Tang, D.; Guo, B.; Li, D.; Cao, F.; Sun, C.; Yu, L.; Yan, Z. The Complete Chloroplast Genomes of Gynostemma Reveal the Phylogenetic Relationships of Species within the Genus. Genes 2023, 14, 929. https://doi.org/10.3390/genes14040929
Gan J, Li Y, Tang D, Guo B, Li D, Cao F, Sun C, Yu L, Yan Z. The Complete Chloroplast Genomes of Gynostemma Reveal the Phylogenetic Relationships of Species within the Genus. Genes. 2023; 14(4):929. https://doi.org/10.3390/genes14040929
Chicago/Turabian StyleGan, Jiaxia, Ying Li, Deying Tang, Baolin Guo, Doudou Li, Feng Cao, Chao Sun, Liying Yu, and Zhuyun Yan. 2023. "The Complete Chloroplast Genomes of Gynostemma Reveal the Phylogenetic Relationships of Species within the Genus" Genes 14, no. 4: 929. https://doi.org/10.3390/genes14040929