
Genome-Wide Identification, Characterization, and Expression of TCP Genes Family in Orchardgrass

 , ,
, ,
Abstract
:1. Introduction
2. Material and Methods
2.1. Identification of Dactylis glomerata TCP Genes
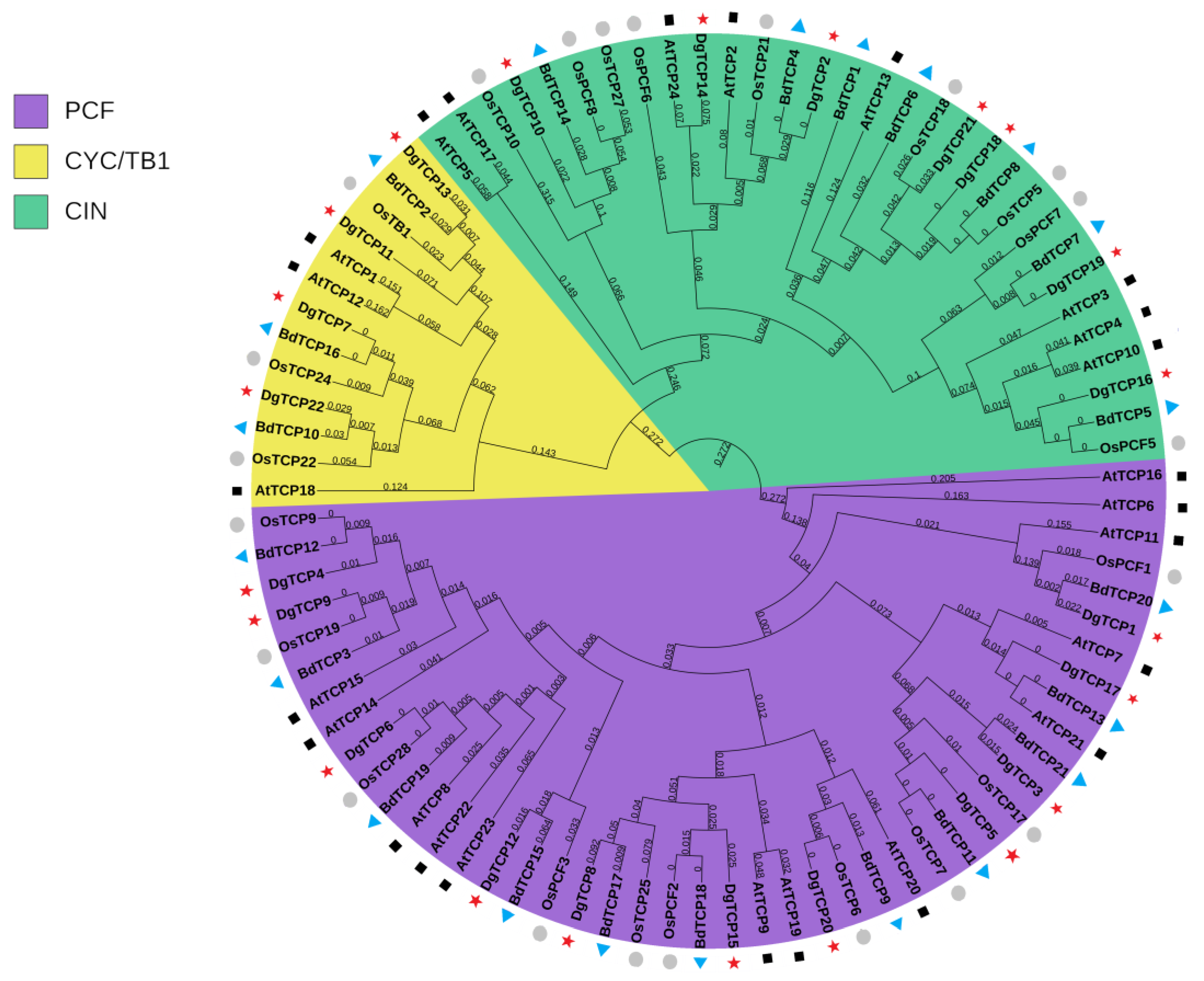
2.2. Phylogenetic Analysis and Classification of DgTCP Genes
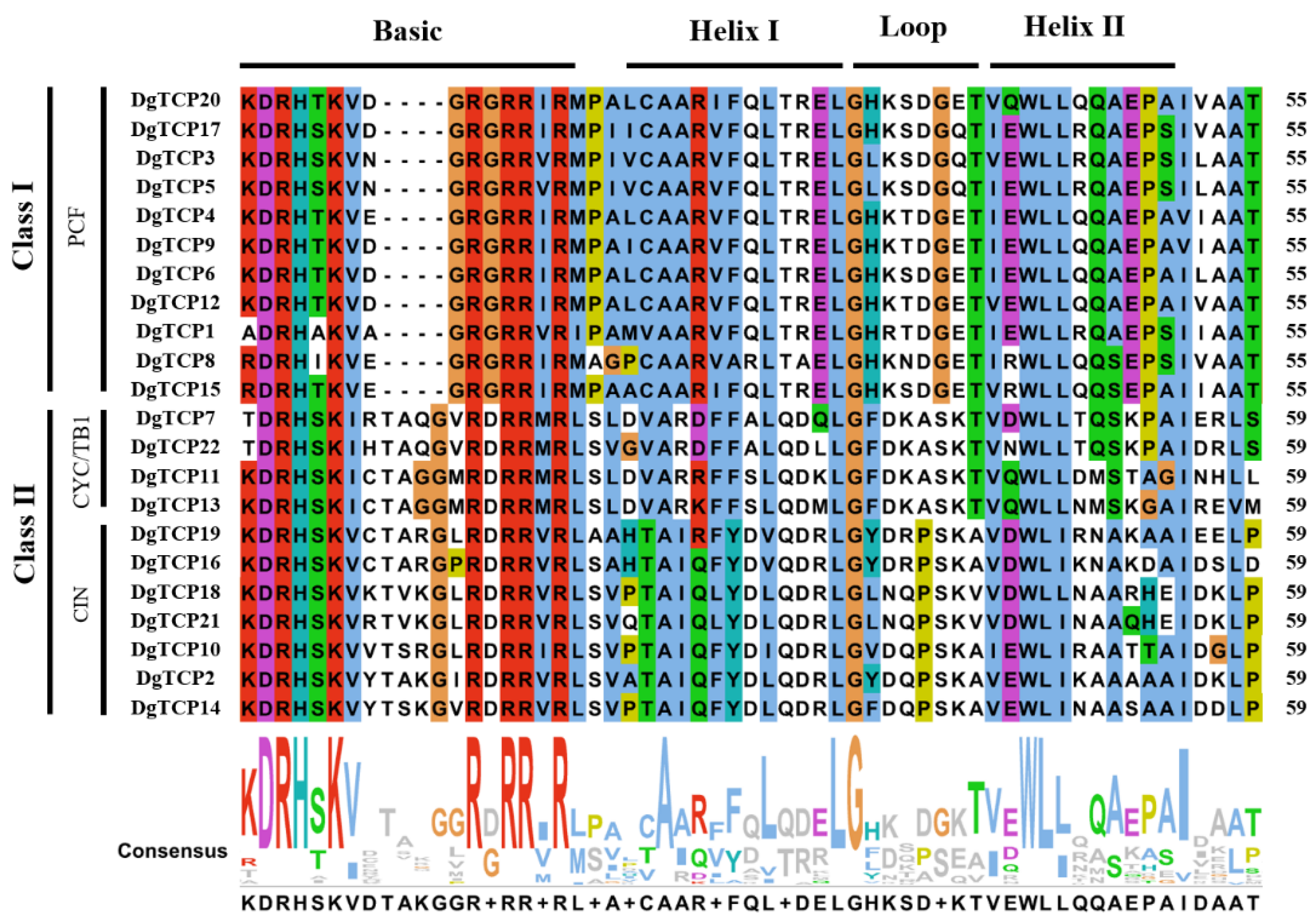
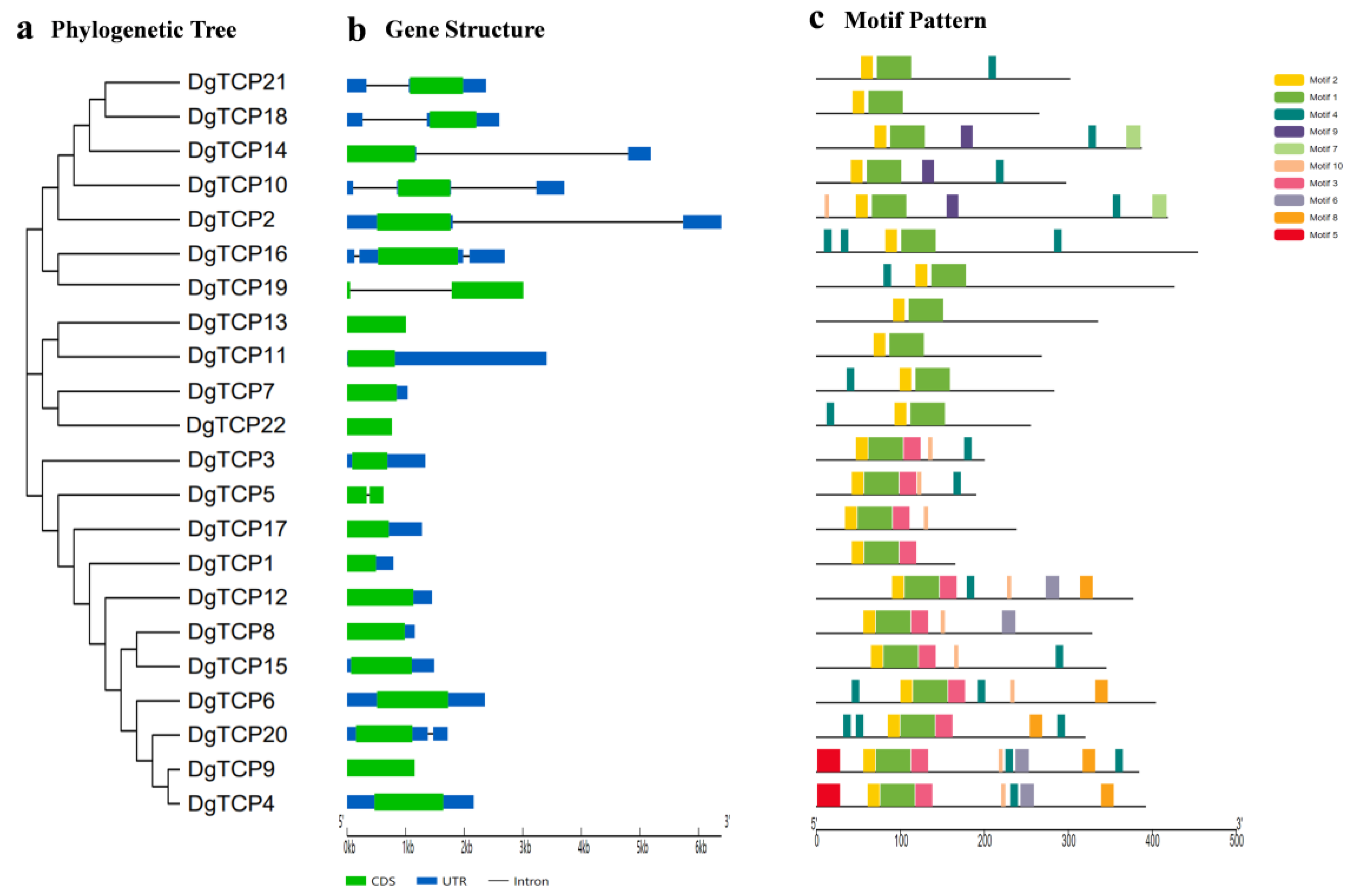
2.3. Gene Structure and Motif Analysis
2.4. Putative Promoter Cis-Acting Element Analysis
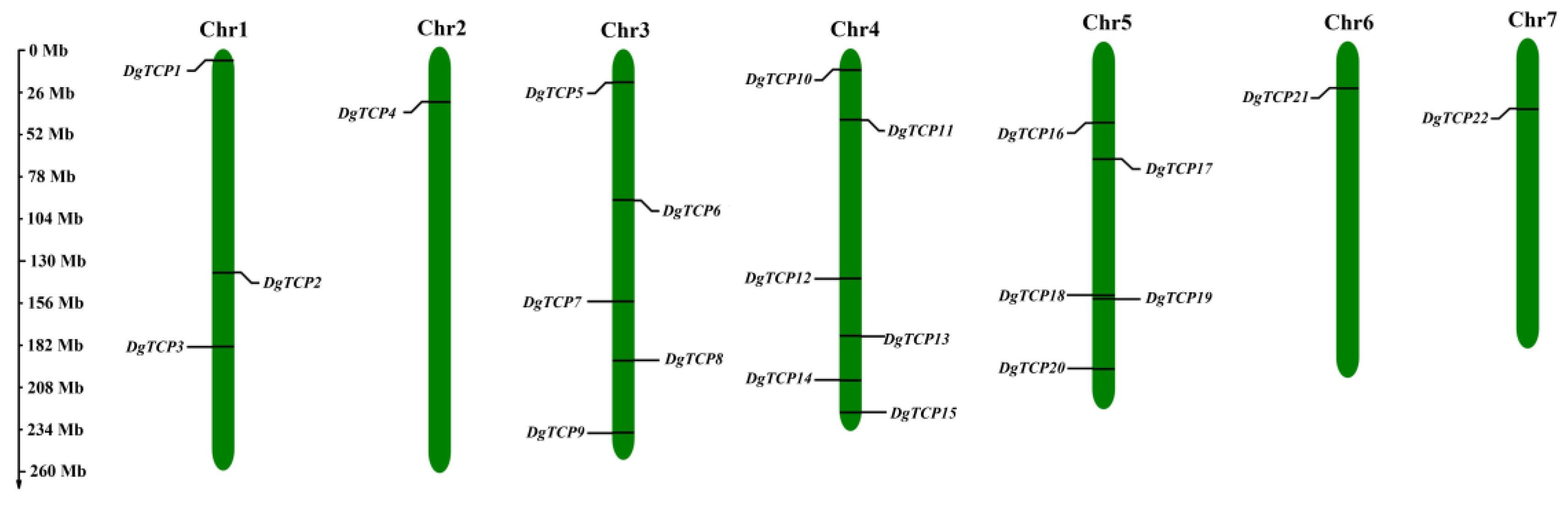
2.5. Chromosomal Mapping and Synteny Analysis
2.6. Plant Material and Treatments
2.7. Expression Profiles of DgTCP Family Members
2.8. Expression of 14 Selected DgTCP Genes in qRT-PCR
3. Results
3.1. Identifying TCP Genes in Orchardgrass
3.2. Phylogeny and Classification of the DgTCP Proteins
3.3. The DgTCP Gene Structure and Protein Motif
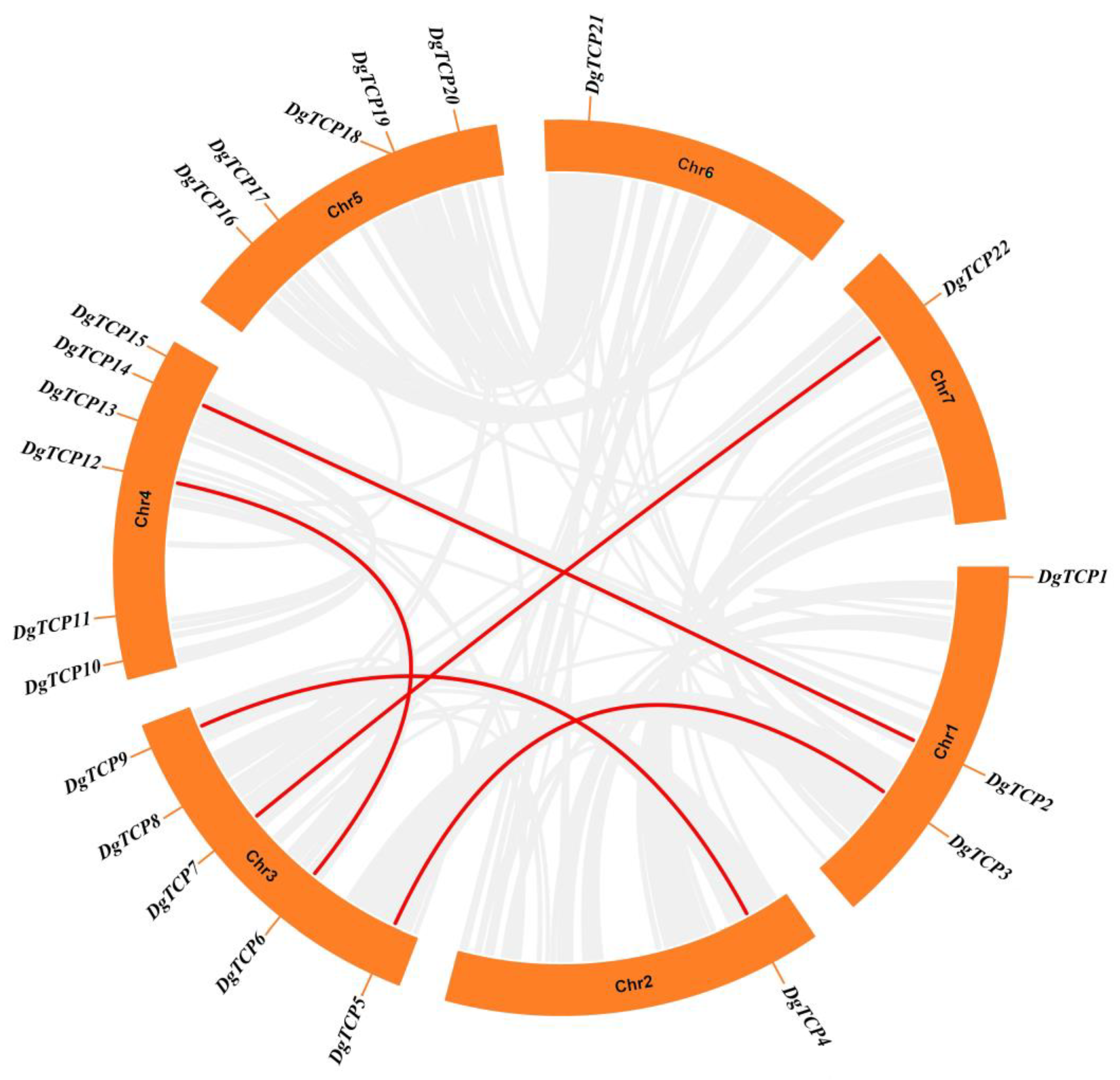
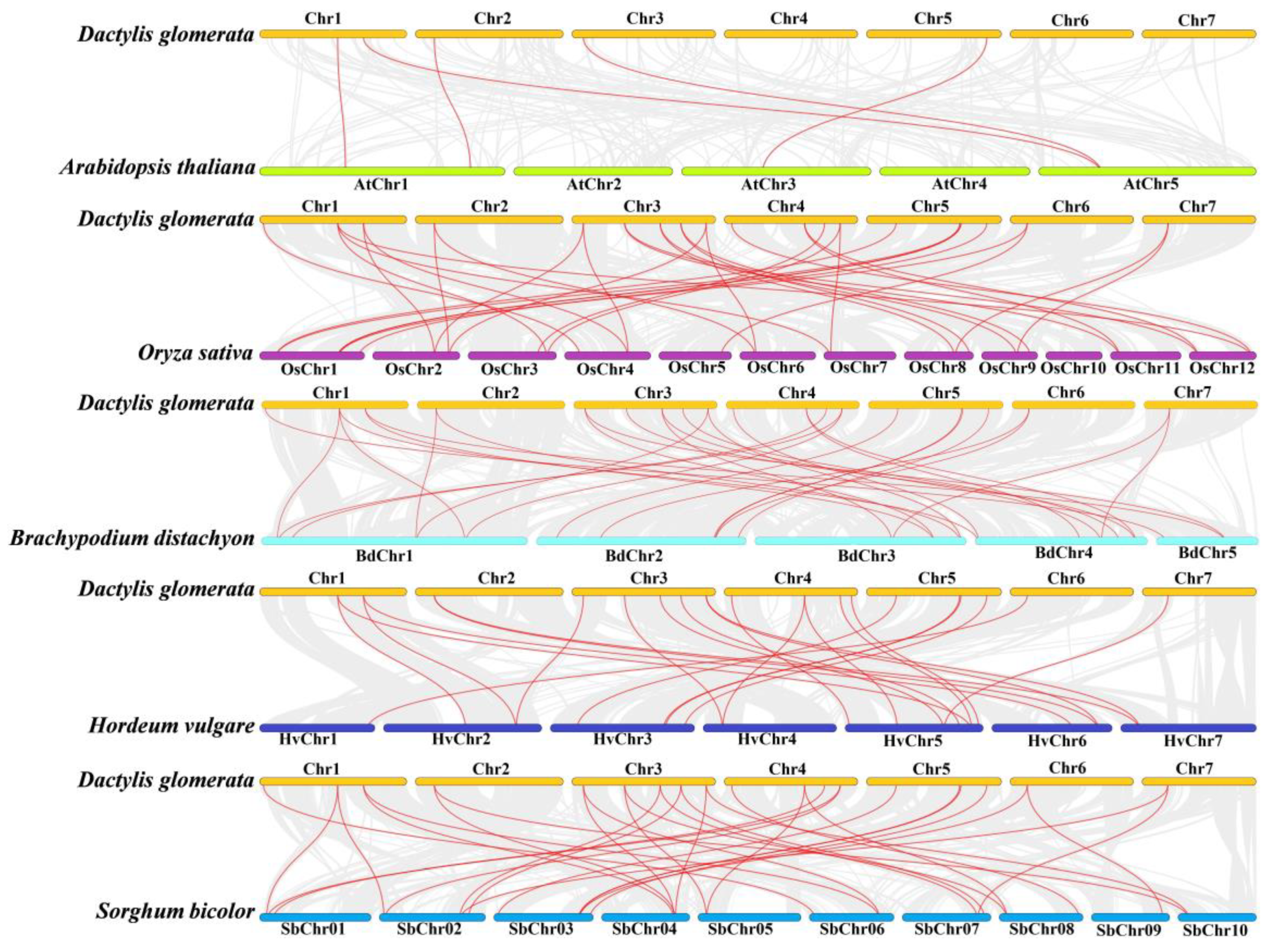
3.4. Chromosomal Localization, Gene Duplication, and Synteny Analysis
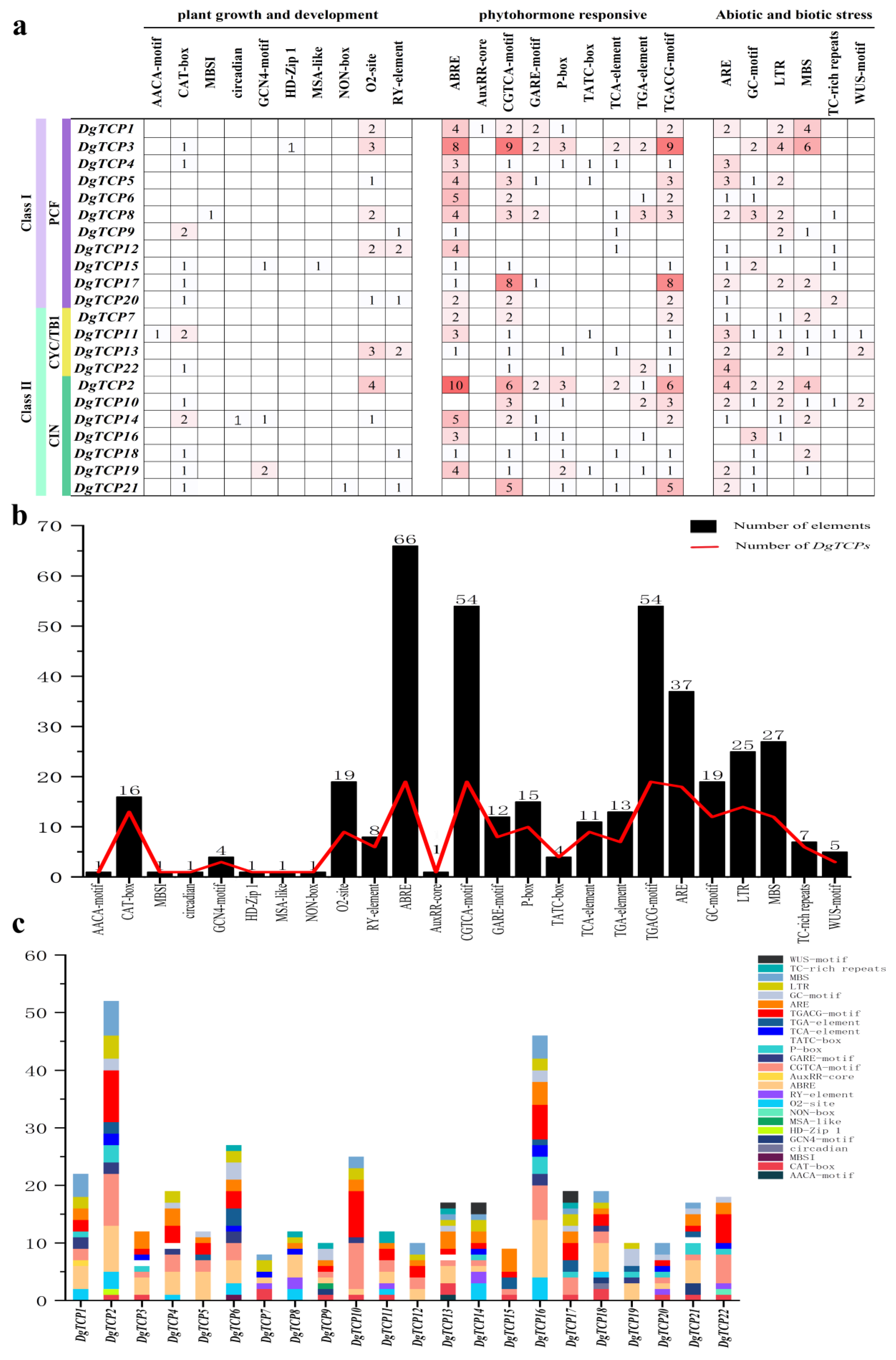
3.5. Putative Cis-Acting Elements of Orchardgrass DgTCPs
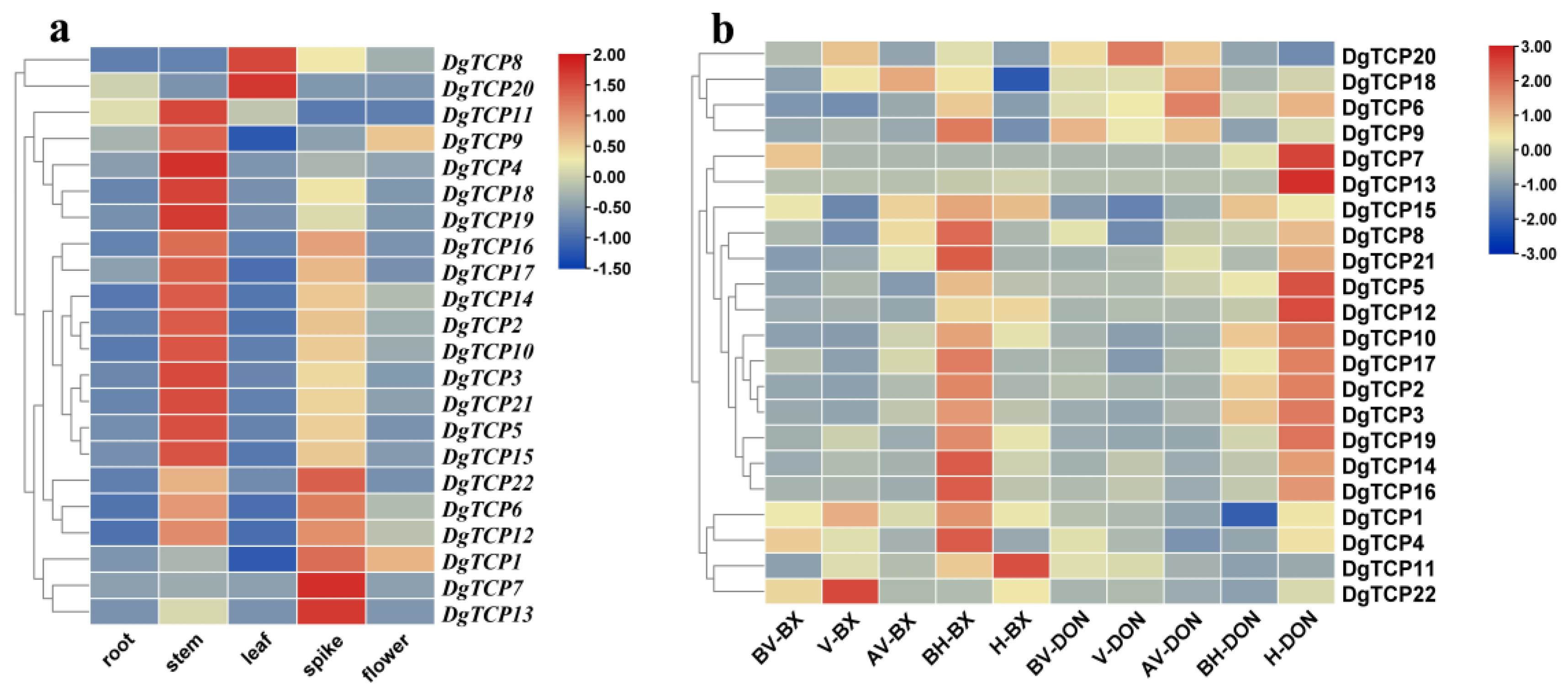
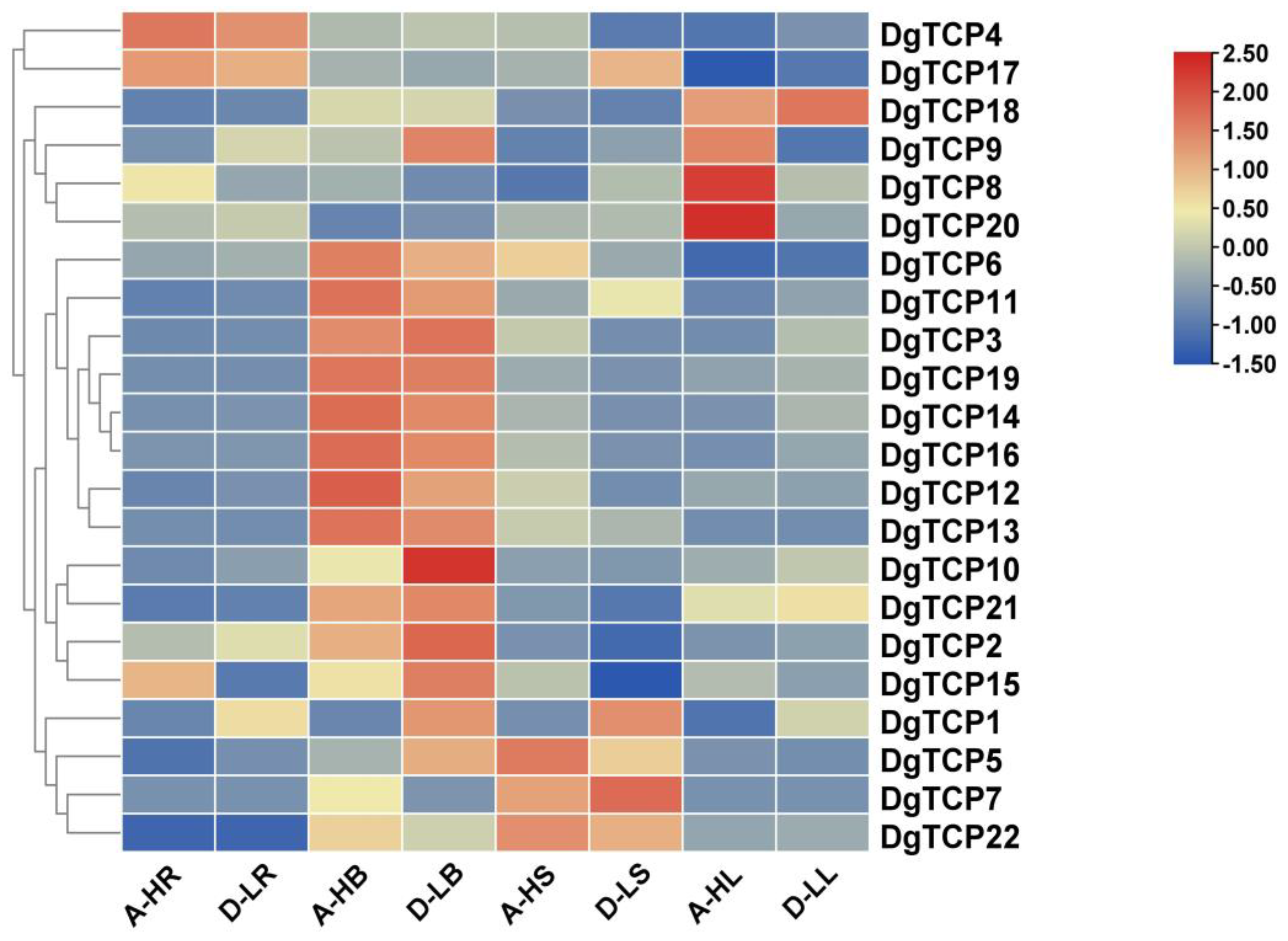
3.6. Expression Profiles of DgTCPs in Different Tissues and Developmental Stages
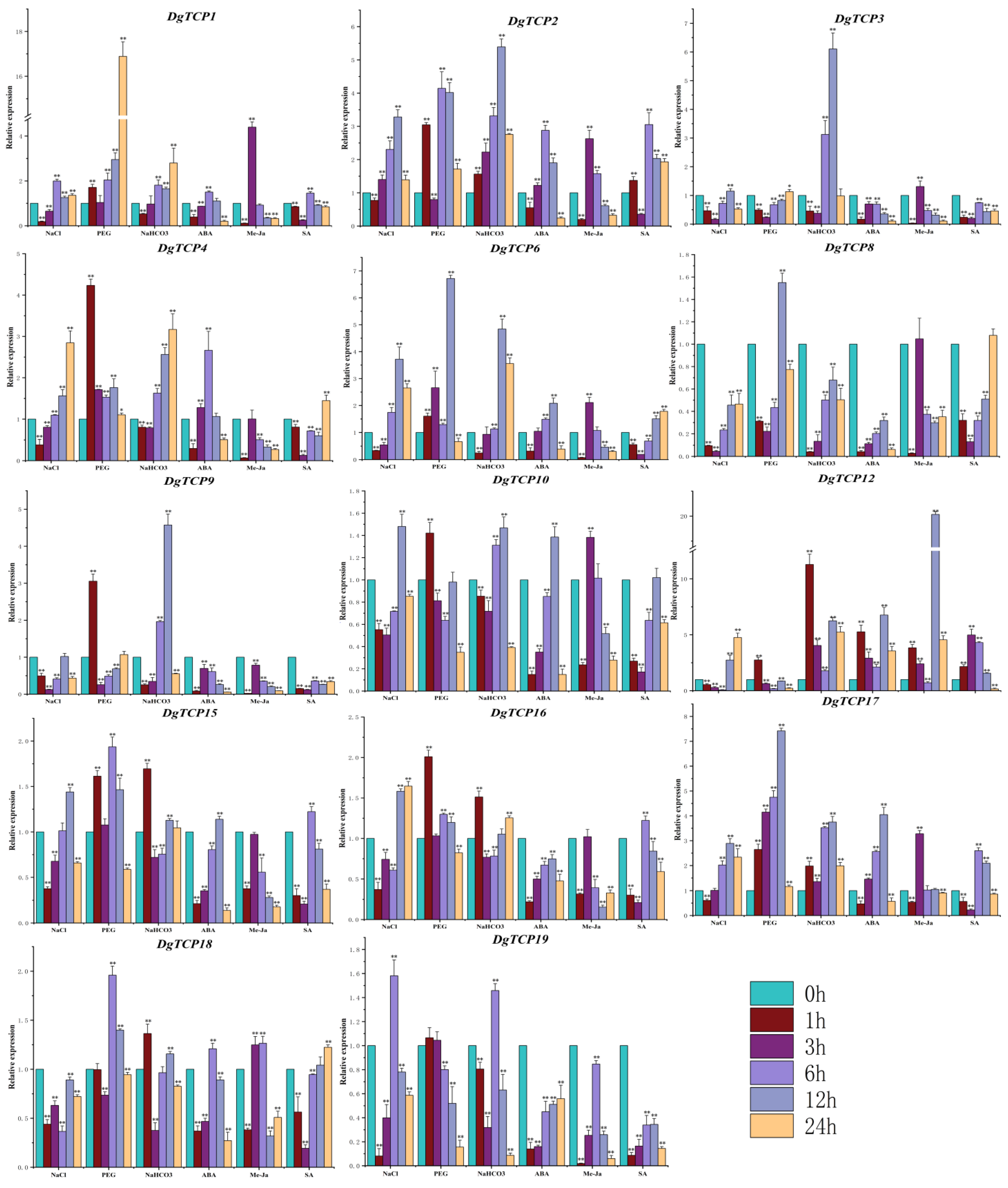
3.7. The Expression of DgTCP Genes under Six Abiotic Stresses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cubas, P.; Lauter, N.; Doebley, J.; Coen, E. The TCP Domain: A Motif Found in Proteins Regulating Plant Growth and Development. Plant J. 1999, 18, 215–222. [Google Scholar] [CrossRef]
- Doebley, J.; Stec, A.; Hubbard, L. The Evolution of Apical Dominance in Maize. Nature 1997, 386, 485–488. [Google Scholar] [CrossRef]
- Luo, D.; Carpenter, R.; Vincent, C.; Copsey, L.; Coen, E. Origin of Floral Asymmetry in Antirrhinum. Nature 1996, 383, 794–799. [Google Scholar] [CrossRef]
- Kosugi, S.; Ohashi, Y. PCF1 and PCF2 Specifically Bind to Cis Elements in the Rice Proliferating Cell Nuclear Antigen Gene. Plant Cell 1997, 9, 1607–1619. [Google Scholar] [CrossRef] [PubMed]
- Martín-Trillo, M.; Cubas, P. TCP Genes: A Family Snapshot Ten Years Later. Trends Plant Sci. 2010, 15, 31–39. [Google Scholar] [CrossRef]
- Horn, S.; Pabón-Mora, N.; Theuß, V.S.; Busch, A.; Zachgo, S. Analysis of the CYC/TB1 Class of TCP Transcription Factors in Basal Angiosperms and Magnoliids. Plant J. 2015, 81, 559–571. [Google Scholar] [CrossRef]
- Nicolas, M.; Cubas, P. TCP Factors: New Kids on the Signaling Block. Curr. Opin. Plant Biol. 2016, 33, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Navaud, O.; Dabos, P.; Carnus, E.; Tremousaygue, D.; Hervé, C. TCP Transcription Factors Predate the Emergence of Land Plants. J. Mol. Evol. 2007, 65, 23–33. [Google Scholar] [CrossRef]
- Schommer, C.; Palatnik, J.F.; Aggarwal, P.; Chételat, A.; Cubas, P.; Farmer, E.E.; Nath, U.; Weigel, D. Control of Jasmonate Biosynthesis and Senescence by MiR319 Targets. PLoS Biol. 2008, 6, e230. [Google Scholar] [CrossRef] [PubMed]
- Giraud, E.; Ng, S.; Carrie, C.; Duncan, O.; Low, J.; Lee, C.P.; Van Aken, O.; Millar, A.H.; Murcha, M.; Whelan, J. TCP Transcription Factors Link the Regulation of Genes Encoding Mitochondrial Proteins with the Circadian Clock in Arabidopsis thaliana. Plant Cell 2011, 22, 3921–3934. [Google Scholar] [CrossRef]
- Aguilar-Martínez, J.A.; Poza-Carrión, C.; Cubas, P. Arabidopsis BRANCHED1 Acts as an Integrator of Branching Signals within Axillary Buds. Plant Cell 2007, 19, 458–472. [Google Scholar] [CrossRef] [PubMed]
- Palatnik, J.F.; Allen, E.; Wu, X.; Schommer, C.; Schwab, R.; Carrington, J.C.; Weigel, D. Control of Leaf Morphogenesis by MicroRNAs. Nature 2003, 425, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Madrigal, Y.; Alzate, J.F.; Pabón-Mora, N. Evolution and Expression Patterns of TCP Genes in Asparagales. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef]
- Li, S. The Arabidopsis thaliana TCP Transcription Factors: A Broadening Horizon beyond Development. Plant Signal. Behav. 2015, 10, e1044192. [Google Scholar] [CrossRef]
- Schommer, C.; Debernardi, J.M.; Bresso, E.G.; Rodriguez, R.E.; Palatnik, J.F. Repression of Cell Proliferation by MiR319-Regulated TCP4. Mol. Plant 2014, 7, 1533–1544. [Google Scholar] [CrossRef]
- Liu, Y.; Guan, X.; Liu, S.; Yang, M.; Ren, J.; Guo, M.; Huang, Z.; Zhang, Y. Genome-Wide Identification and Analysis of TCP Transcription Factors Involved in the Formation of Leafy Head in Chinese Cabbage. Int. J. Mol. Sci. 2018, 19, 847. [Google Scholar] [CrossRef] [PubMed]
- Yin, Z.; Li, Y.; Zhu, W.; Fu, X.; Han, X.; Wang, J.; Lin, H.; Ye, W. Identification, Characterization, and Expression Patterns of TCP Genes and MicroRNA319 in Cotton. Int. J. Mol. Sci. 2018, 19, 3655. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Ma, J.; Fan, D.; Li, C.; Jiang, Y.; Luo, K. Genome-Wide Identification of TCP Family Transcription Factors from Populus Euphratica and Their Involvement in Leaf Shape Regulation. Sci. Rep. 2016, 6, 32795. [Google Scholar] [CrossRef] [PubMed]
- Kieffer, M.; Master, V.; Waites, R.; Davies, B. TCP14 and TCP15 Affect Internode Length and Leaf Shape in Arabidopsis: TCP14/15 Affect Internode Length and Leaf Shape. Plant J. 2011, 68, 147–158. [Google Scholar] [CrossRef]
- Resentini, F.; Felipo-Benavent, A.; Colombo, L.; Blázquez, M.A.; Alabadi, D.; Masiero, S. TCP14 and TCP15 Mediate the Promotion of Seed Germination by Gibberellins in Arabidopsis thaliana. Mol. Plant 2021, 14, 1771. [Google Scholar] [CrossRef]
- Danisman, S.; van der Wal, F.; Dhondt, S.; Waites, R.; de Folter, S.; Bimbo, A.; van Dijk, A.D.; Muino, J.M.; Cutri, L.; Dornelas, M.C.; et al. Arabidopsis Class I and Class II TCP Transcription Factors Regulate Jasmonic Acid Metabolism and Leaf Development Antagonistically. Plant Physiol. 2012, 159, 1511–1523. [Google Scholar] [CrossRef] [PubMed]
- Takeda, T.; Amano, K.; Ohto, M.; Nakamura, K.; Sato, S.; Kato, T.; Tabata, S.; Ueguchi, C. RNA Interference of the Arabidopsis Putative Transcription Factor TCP16 Gene Results in Abortion of Early Pollen Development. Plant. Mol. Biol. 2006, 61, 165–177. [Google Scholar] [CrossRef] [PubMed]
- Nag, A.; King, S.; Jack, T. MiR319a Targeting of TCP4 Is Critical for Petal Growth and Development in Arabidopsis. Proc. Natl. Acad. Sci. USA 2009, 106, 22534–22539. [Google Scholar] [CrossRef]
- Challa, K.R.; Aggarwal, P.; Nath, U. Activation of YUCCA5 by the Transcription Factor TCP4 Integrates Developmental and Environmental Signals to Promote Hypocotyl Elongation in Arabidopsis. Plant Cell 2016, 28, 2117–2130. [Google Scholar] [CrossRef] [PubMed]
- Nagpal, P.; Ellis, C.M.; Weber, H.; Ploense, S.E.; Barkawi, L.S.; Guilfoyle, T.J.; Hagen, G.; Alonso, J.M.; Cohen, J.D.; Farmer, E.E.; et al. Auxin Response Factors ARF6 and ARF8 Promote Jasmonic Acid Production and Flower Maturation. Development 2005, 132, 4107–4118. [Google Scholar] [CrossRef]
- Rubio-Somoza, I.; Zhou, C.-M.; Confraria, A.; Martinho, C.; von Born, P.; Baena-Gonzalez, E.; Wang, J.-W.; Weigel, D. Temporal Control of Leaf Complexity by MiRNA-Regulated Licensing of Protein Complexes. Curr. Biol. 2014, 24, 2714–2719. [Google Scholar] [CrossRef]
- Wang, J.; Wang, Y.; Luo, D. LjCYC Genes Constitute Floral Dorsoventral Asymmetry in Lotus Japonicus: LjCYC Genes Control Dorsoventral Asymmetry. J. Integr. Plant Biol. 2010, 52, 959–970. [Google Scholar] [CrossRef]
- Koyama, T.; Sato, F.; Ohme-Takagi, M. A Role of TCP1 in the Longitudinal Elongation of Leaves in Arabidopsis. Biosci. Biotechnol. Biochem. 2010, 74, 2145–2147. [Google Scholar] [CrossRef]
- Broholm, S.K.; Tähtiharju, S.; Laitinen, R.A.E.; Albert, V.A.; Teeri, T.H.; Elomaa, P. A TCP Domain Transcription Factor Controls Flower Type Specification along the Radial Axis of the Gerbera (Asteraceae) Inflorescence. Proc. Natl. Acad. Sci. USA 2008, 105, 9117–9122. [Google Scholar] [CrossRef]
- Doebley, J.; Stec, A.; Gustus, C. Teosinte Branched1 and the Origin of Maize: Evidence for Epistasis and the Evolution of Dominance. Genetics 1995, 141, 333–346. [Google Scholar] [CrossRef]
- Takeda, T.; Suwa, Y.; Suzuki, M.; Kitano, H.; Ueguchi-Tanaka, M.; Ashikari, M.; Matsuoka, M.; Ueguchi, C. The OsTB1 Gene Negatively Regulates Lateral Branching in Rice. Plant J. 2003, 33, 513–520. [Google Scholar] [CrossRef]
- Wu, J.-F.; Tsai, H.-L.; Joanito, I.; Wu, Y.-C.; Chang, C.-W.; Li, Y.-H.; Wang, Y.; Hong, J.C.; Chu, J.-W.; Hsu, C.-P.; et al. LWD–TCP Complex Activates the Morning Gene CCA1 in Arabidopsis. Nat. Commun. 2016, 7, 13181. [Google Scholar] [CrossRef]
- Arab, M.M.; Marrano, A.; Abdollahi-Arpanahi, R.; Leslie, C.A.; Askari, H.; Neale, D.B.; Vahdati, K. Genome-Wide Patterns of Population Structure and Association Mapping of Nut-Related Traits in Persian Walnut Populations from Iran Using the Axiom J. Regia 700K SNP Array. Sci. Rep. 2019, 9, 6376. [Google Scholar] [CrossRef]
- Mukhopadhyay, P.; Tyagi, A.K. OsTCP19 Influences Developmental and Abiotic Stress Signaling by ModulatingABI4-Mediated Pathways. Sci. Rep. 2015, 5, 9998. [Google Scholar] [CrossRef]
- Wang, S.; Sun, X.; Hoshino, Y.; Yu, Y.; Jia, B.; Sun, Z.; Sun, M.; Duan, X.; Zhu, Y. MicroRNA319 Positively Regulates Cold Tolerance by Targeting OsPCF6 and OsTCP21 in Rice (Oryza Sativa L.). PLoS ONE 2014, 9, e91357. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Li, D.; Mao, D.; Liu, X.; Ji, C.; Li, X.; Zhao, X.; Cheng, Z.; Chen, C.; Zhu, L. Overexpression of MicroRNA319 Impacts Leaf Morphogenesis and Leads to Enhanced Cold Tolerance in Rice (O Ryza Sativa L.): Rice MiR319 and Cold Response. Plant Cell Environ. 2013, 36, 2207–2218. [Google Scholar] [CrossRef]
- Almeida, D.M.; Gregorio, G.B.; Oliveira, M.M.; Saibo, N.J.M. Five Novel Transcription Factors as Potential Regulators of OsNHX1 Gene Expression in a Salt Tolerant Rice Genotype. Plant Mol. Biol. 2017, 93, 61–77. [Google Scholar] [CrossRef] [PubMed]
- Guan, P.; Ripoll, J.-J.; Wang, R.; Vuong, L.; Bailey-Steinitz, L.J.; Ye, D.; Crawford, N.M. Interacting TCP and NLP Transcription Factors Control Plant Responses to Nitrate Availability. Proc. Natl. Acad. Sci. USA 2017, 114, 2419–2424. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Li, D.; Li, Z.; Hu, Q.; Yang, C.; Zhu, L.; Luo, H. Constitutive Expression of a MiR319 Gene Alters Plant Development and Enhances Salt and Drought Tolerance in Transgenic Creeping Bentgrass. Plant Physiol. 2013, 161, 1375–1391. [Google Scholar] [CrossRef]
- Zhou, M.; Luo, H. Role of MicroRNA319 in Creeping Bentgrass Salinity and Drought Stress Response. Plant Signal. Behav. 2014, 9, e28700. [Google Scholar] [CrossRef] [PubMed]
- İlhan, E.; Büyük, İ.; İnal, B. Transcriptome—Scale Characterization of Salt Responsive Bean TCP Transcription Factors. Gene 2018, 642, 64–73. [Google Scholar] [CrossRef]
- Xu, Y.; Liu, H.; Gao, Y.; Xiong, R.; Wu, M.; Zhang, K.; Xiang, Y. The TCP Transcription Factor PeTCP10 Modulates Salt Tolerance in Transgenic Arabidopsis. Plant Cell Rep. 2021, 40, 1971–1987. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Yan, H.; Jiang, X.; Zhang, Y.; Zhang, X.; Ji, Y.; Zeng, B.; Xu, B.; Yin, G.; Lee, S.; et al. Reference Gene Selection for Quantitative Real-Time Reverse-Transcriptase PCR in Orchardgrass Subjected to Various Abiotic Stresses. Gene 2014, 553, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Casler, M.D.; Fales, S.L.; Undersander, D.J.; McElroy, A.R. Genetic Progress from 40 Years of Orchardgrass Breeding in North America Measured under Management-Intensive Rotational Grazing. Can. J. Plant Sci. 2001, 81, 713–721. [Google Scholar] [CrossRef]
- Finn, R.D.; Clements, J.; Eddy, S.R. HMMER Web Server: Interactive Sequence Similarity Searching. Nucleic Acids Res. 2011, 39, W29–W37. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Feng, G.; Yan, H.; Zhang, Z.; Bushman, B.S.; Wang, J.; Bombarely, A.; Li, M.; Yang, Z.; Nie, G.; et al. Genome Assembly Provides Insights into the Genome Evolution and Flowering Regulation of Orchardgrass. Plant Biotechnol. J. 2020, 18, 373–388. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X Version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef]
- Gasteiger, E. ExPASy: The Proteomics Server for in-Depth Protein Knowledge and Analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef]
- Rhee, S.Y. The Arabidopsis Information Resource (TAIR): A Model Organism Database Providing a Centralized, Curated Gateway to Arabidopsis Biology, Research Materials and Community. Nucleic Acids Res. 2003, 31, 224–228. [Google Scholar] [CrossRef]
- Jin, J.; Tian, F.; Yang, D.-C.; Meng, Y.-Q.; Kong, L.; Luo, J.; Gao, G. PlantTFDB 4.0: Toward a Central Hub for Transcription Factors and Regulatory Interactions in Plants. Nucleic Acids Res. 2017, 45, D1040–D1045. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular Evolutionary Genetics Analysis Using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Jin, J.; Guo, A.-Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An Upgraded Gene Feature Visualization Server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME Suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef] [PubMed]
- Lescot, M. PlantCARE, a Database of Plant Cis-Acting Regulatory Elements and a Portal to Tools for in Silico Analysis of Promoter Sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; DeBarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.-H.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A Toolkit for Detection and Evolutionary Analysis of Gene Synteny and Collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Feng, G.; Huang, L.; Li, J.; Wang, J.; Xu, L.; Pan, L.; Zhao, X.; Wang, X.; Huang, T.; Zhang, X. Comprehensive Transcriptome Analysis Reveals Distinct Regulatory Programs during Vernalization and Floral Bud Development of Orchardgrass (Dactylis Glomerata L.). BMC Plant Biol 2017, 17, 216. [Google Scholar] [CrossRef]
- Xu, X.; Feng, G.; Liang, Y.; Shuai, Y.; Liu, Q.; Nie, G.; Yang, Z.; Hang, L.; Zhang, X. Comparative Transcriptome Analyses Reveal Different Mechanism of High- and Low-Tillering Genotypes Controlling Tiller Growth in Orchardgrass (Dactylis Glomerata L.). BMC Plant Biol. 2020, 20, 369. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Huo, Y.; Xiong, W.; Su, K.; Li, Y.; Yang, Y.; Fu, C.; Wu, Z.; Sun, Z. Genome-Wide Analysis of the TCP Gene Family in Switchgrass (Panicum Virgatum L.). Int. J. Genom. 2019, 2019, 8514928. [Google Scholar] [CrossRef]
- Ding, S.; Cai, Z.; Du, H.; Wang, H. Genome-Wide Analysis of TCP Family Genes in Zea Mays L. Identified a Role for ZmTCP42 in Drought Tolerance. Int. J. Mol. Sci. 2019, 20, 2762. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Ma, H.; Wang, J.; Zhang, D. Genome-Wide Comparative Analysis and Expression Pattern of TCP Gene Families in Arabidopsis thaliana and Oryza sativa. J. Integr. Plant Biol. 2007, 49, 885–897. [Google Scholar] [CrossRef]
- Liu, H.-L.; Wu, M.; Li, F.; Gao, Y.-M.; Chen, F.; Xiang, Y. TCP Transcription Factors in Moso Bamboo (Phyllostachys Edulis): Genome-Wide Identification and Expression Analysis. Front. Plant Sci. 2018, 9, 1263. [Google Scholar] [CrossRef]
- Jiu, S.; Xu, Y.; Wang, J.; Wang, L.; Wang, S.; Ma, C.; Guan, L.; Abdullah, M.; Zhao, M.; Xu, W.; et al. Genome-Wide Identification, Characterization, and Transcript Analysis of the TCP Transcription Factors in Vitis Vinifera. Front. Genet. 2019, 10, 1276. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Hu, Y.; Cui, M.-Y.; Han, Y.-T.; Gao, K.; Feng, J.-Y. Identification and Transcript Analysis of the TCP Transcription Factors in the Diploid Woodland Strawberry Fragaria Vesca. Front. Plant Sci. 2016, 07. [Google Scholar] [CrossRef]
- Francis, A.; Dhaka, N.; Bakshi, M.; Jung, K.-H.; Sharma, M.K.; Sharma, R. Comparative Phylogenomic Analysis Provides Insights into TCP Gene Functions in Sorghum. Sci. Rep. 2016, 6, 38488. [Google Scholar] [CrossRef]
- Feng, Z.-J.; Xu, S.-C.; Liu, N.; Zhang, G.-W.; Hu, Q.-Z.; Gong, Y.-M. Soybean TCP Transcription Factors: Evolution, Classification, Protein Interaction and Stress and Hormone Responsiveness. Plant Physiol. Biochem. 2018, 127, 129–142. [Google Scholar] [CrossRef]
- Chen, L.; Chen, Y.Q.; Ding, A.M.; Chen, H.; Xia, F.; Wang, W.F.; Sun, Y.H. Genome-Wide Analysis of TCP Family in Tobacco. Genet. Mol. Res. 2016, 15, gmr.15027728. [Google Scholar] [CrossRef]
- Vatansever, R.; Koc, I.; Ozyigit, I.I.; Sen, U.; Uras, M.E.; Anjum, N.A.; Pereira, E.; Filiz, E. Genome-Wide Identification and Expression Analysis of Sulfate Transporter (SULTR) Genes in Potato (Solanum Tuberosum L.). Planta 2016, 244, 1167–1183. [Google Scholar] [CrossRef]
- Chai, W.; Jiang, P.; Huang, G.; Jiang, H.; Li, X. Identification and Expression Profiling Analysis of TCP Family Genes Involved in Growth and Development in Maize. Physiol. Mol. Biol. Plants 2017, 23, 779–791. [Google Scholar] [CrossRef]
- Danisman, S. TCP Transcription Factors at the Interface between Environmental Challenges and the Plant’s Growth Responses. Front. Plant Sci. 2016, 7, 1930. [Google Scholar] [CrossRef]
- Wen, Y.; Raza, A.; Chu, W.; Zou, X.; Cheng, H.; Hu, Q.; Liu, J.; Wei, W. Comprehensive In Silico Characterization and Expression Profiling of TCP Gene Family in Rapeseed. Front. Genet. 2021, 12, 794297. [Google Scholar] [CrossRef]
- Nakashima, K.; Yamaguchi-Shinozaki, K. ABA Signaling in Stress-Response and Seed Development. Plant Cell Rep. 2013, 32, 959–970. [Google Scholar] [CrossRef]
- Wang, X.; Gao, J.; Zhu, Z.; Dong, X.; Wang, X.; Ren, G.; Zhou, X.; Kuai, B. TCP Transcription Factors Are Critical for the Coordinated Regulation of ISOCHORISMATE SYNTHASE 1 Expression in Arabidopsis thaliana. Plant J. 2015, 82, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Zhang, S.; Chen, F.; Liu, B.; Wu, L.; Li, F.; Zhang, J.; Bao, M.; Liu, G. Genome-Wide Identification and Characterization of the SBP-Box Gene Family in Petunia. BMC Genom. 2018, 19, 193. [Google Scholar] [CrossRef]
- Balsemão-Pires, E.; Andrade, L.R.; Sachetto-Martins, G. Functional Study of TCP23 in Arabidopsis thaliana during Plant Development. Plant Physiol. Biochem. 2013, 67, 120–125. [Google Scholar] [CrossRef]
- Li, D.; Zhang, H.; Mou, M.; Chen, Y.; Xiang, S.; Chen, L.; Yu, D. Arabidopsis Class II TCP Transcription Factors Integrate with the FT–FD Module to Control Flowering. Plant Physiol. 2019, 181, 97–111. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, G.; Liang, Y.; Hu, L.; Zhu, B.; Qi, D.; Cui, S.; Zhao, H. TCP7 Interacts with Nuclear Factor-Ys to Promote Flowering by Directly Regulating SOC1 in Arabidopsis. Plant J. 2021, 108, 1493–1506. [Google Scholar] [CrossRef] [PubMed]
- Jewiss, O.R. Tillering in grasses-its significance and control. Grass Forage Sci. 1972, 27, 65–82. [Google Scholar] [CrossRef]
- Langer, R. Growth of the Grass Plant in Relation to Seed Production; Lincoln College: Lincoln, UK, 1980. [Google Scholar]
- Liu, Y.; Wang, H.; Jiang, Z.; Wang, W.; Xu, R.; Wang, Q.; Zhang, Z.; Li, A.; Liang, Y.; Ou, S.; et al. Genomic Basis of Geographical Adaptation to Soil Nitrogen in Rice. Nature 2021, 590, 600–605. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Chr | Protein Length (aa) | Length (bp) | Molecular Weight (kDa) | Isoelectric Point (pI) | Start | End |
|---|---|---|---|---|---|---|---|---|
| DgTCP1 | DG1C00255.1 | Chr1 | 165 | 495 | 17,433.51 | 5.18 | 6,629,906 | 6,630,695 |
| DgTCP2 | DG1C03379.1 | Chr1 | 418 | 1254 | 44,766.77 | 9.34 | 146,031,371 | 146,037,761 |
| DgTCP3 | DG1C04745.1 | Chr1 | 200 | 600 | 21,319.20 | 9.92 | 194,567,533 | 194,568,868 |
| DgTCP4 | DG2C01041.1 | Chr2 | 392 | 1176 | 39,526.73 | 9.42 | 35,226,595 | 35,228,755 |
| DgTCP5 | DG3C00669.1 | Chr3 | 190 | 570 | 20,377.03 | 9.41 | 20,872,684 | 20,873,306 |
| DgTCP6 | DG3C02184.1 | Chr3 | 404 | 1212 | 41,896.68 | 5.71 | 98,406,794 | 98,409,146 |
| DgTCP7 | DG3C03958.1 | Chr3 | 283 | 849 | 30,759.21 | 6.21 | 165,195,655 | 165,196,687 |
| DgTCP8 | DG3C05144.1 | Chr3 | 328 | 984 | 33,731.69 | 5.09 | 204,201,074 | 204,202,229 |
| DgTCP9 | DG3C06789.1 | Chr3 | 384 | 1152 | 40,053.43 | 8.49 | 251,633,645 | 251,634,796 |
| DgTCP10 | DG4C00330.1 | Chr4 | 297 | 891 | 31,333.77 | 6.55 | 13,226,786 | 13,230,494 |
| DgTCP11 | DG4C01109.1 | Chr4 | 268 | 804 | 29,570.36 | 9.15 | 45,855,850 | 45,859,255 |
| DgTCP12 | DG4C03206.1 | Chr4 | 377 | 1131 | 39,248.24 | 7.99 | 150,217,992 | 150,219,441 |
| DgTCP13 | DG4C04368.1 | Chr4 | 335 | 1005 | 36,236.47 | 9.01 | 187,896,786 | 187,897,790 |
| DgTCP14 | DG4C05327.1 | Chr4 | 387 | 1161 | 39,895.82 | 9.07 | 217,216,581 | 217,221,767 |
| DgTCP15 | DG4C06102.1 | Chr4 | 345 | 1035 | 35,534.55 | 5.80 | 238,269,030 | 238,270,515 |
| DgTCP16 | DG5C01593.1 | Chr5 | 454 | 1362 | 47,382.56 | 6.46 | 55,126,146 | 55,128,837 |
| DgTCP17 | DG5C02296.1 | Chr5 | 238 | 714 | 24,165.04 | 7.15 | 80,396,971 | 80,398,250 |
| DgTCP18 | DG5C04172.1 | Chr5 | 265 | 795 | 28,827.16 | 6.90 | 174,582,071 | 174,584,669 |
| DgTCP19 | DG5C04259.1 | Chr5 | 426 | 1278 | 45,754.54 | 6.09 | 176,990,965 | 176,993,974 |
| DgTCP20 | DG5C05852.1 | Chr5 | 320 | 960 | 33,538.98 | 6.05 | 225,545,972 | 225,547,688 |
| DgTCP21 | DG6C01132.1 | Chr6 | 302 | 906 | 32,382.92 | 6.50 | 31,588,652 | 31,591,023 |
| DgTCP22 | DG7C01355.1 | Chr7 | 255 | 765 | 27,479.72 | 6.14 | 48,396,599 | 48,397,363 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, C.; Feng, G.; Xu, X.; Huang, L.; Nie, G.; Li, D.; Zhang, X. Genome-Wide Identification, Characterization, and Expression of TCP Genes Family in Orchardgrass. Genes 2023, 14, 925. https://doi.org/10.3390/genes14040925
Wang C, Feng G, Xu X, Huang L, Nie G, Li D, Zhang X. Genome-Wide Identification, Characterization, and Expression of TCP Genes Family in Orchardgrass. Genes. 2023; 14(4):925. https://doi.org/10.3390/genes14040925
Chicago/Turabian StyleWang, Cheng, Guangyan Feng, Xiaoheng Xu, Linkai Huang, Gang Nie, Dandan Li, and Xinquan Zhang. 2023. "Genome-Wide Identification, Characterization, and Expression of TCP Genes Family in Orchardgrass" Genes 14, no. 4: 925. https://doi.org/10.3390/genes14040925