
TPD1-like Gene as a Suitable Marker for Early Sex Determination in Date Palm (Phoenix dactylifera L.)

1
Institute of Plant Breeding and Biotechnology, MNS-University of Agriculture, Multan 60000, Pakistan
2
Institute of Horticultural Sciences, University of Agriculture, Faisalabad 38040, Pakistan
3
Department of Agronomy, MNS-University of Agriculture, Multan 60000, Pakistan
*
Author to whom correspondence should be addressed.
Genes 2023, 14(4), 907; https://doi.org/10.3390/genes14040907
Submission received: 13 February 2023
/
Revised: 22 March 2023
/
Accepted: 30 March 2023
/
Published: 13 April 2023
(This article belongs to the Special Issue Plant Breeding: Molecular Genetics, Challenges, Opportunities and Future Perspectives)
Abstract
:Date palm (Phoenix dactylifera L.) is a considerably beneficial and economically profitable fruit crop. Female date palm plants produce fruit that is rich in fiber and sugar. Date palm is propagated by two means: suckers and seed. The propagation of date palm through seeds is very necessary for germplasm conservation and breeding. The late reproductive age (4–5 years) and dioecious nature of date palm make genetic improvement and breeding difficult. Early sex determination is the only way to improve breeding by selecting experimental male and female plants at the seedling stage. The primers for Tapetum Determinant 1 (TPD1-like) were designed using Amplify software. The DNA amplification of selected date palm suckers of three genotypes (Ajwa, Amber, and Medjool) was observed through PCR. Expression profiling of selected genotypes was carried out through semi-q PCR and RT-PCR by using the cDNA of suckers and unknown seedlings. Different in silico analyses were performed for the gene and protein characterization and identification of cis-acting elements in the promoter region. The promoter was identified along with the protein’s properties and functionality. The expression of TPD1-like gene was found in the leaves of three selected genotypes of male sucker and in some plants of selected unknown seedlings that are considered male plants, and no expression was observed in female suckers and unknown seedlings that are considered female plants. The findings suggested that the TPD1-like gene has the potential for sex differentiation at the seedling stage, as the TPD1-like gene is essential to the specialization of tapetal cells and plays a critical role in plant reproduction.
Keywords:
Ajwa; Amber; Medjool; suckers; seedling; protein; gene sequence; polymerase chain reaction1. Introduction
Date palm (P. dactylifera) is a dioecious, monocotyledonous fruit crop belonging to the family Arecaceae [1]. There are 14 dioecious species of Phoenix, but only date palm produces fruit [2]. It is cultivated in many regions of the world, particularly in North Africa, the Middle East and South Asia. It is also called the “tree of life” due to its ability to grow in harsh weather conditions that are not fit for growing other crops. Date palm is extensively grown in dry areas because it can tolerate drought and salinity, and can grow well in alkaline soils [3,4]. The fruit of date palm is a main source of macro- and micronutrients, providing nutritional fiber, soluble sugars, and an ample supply of lipids, proteins and carbohydrates [5,6]. Certain varieties such as Ajwa, Medjool, and Amber are considered to have a high value due to unique characteristics such as their taste, size and texture. These varieties are often in high demand and command a premium price in the market. This can result in a significantly higher income for farmers who grow these varieties compared to those who grow ordinary varieties [7,8].
Seed propagation is required to preserve the genetic variation that promotes date palm breeding and the development of new, improved cultivars. Its breeding is limited due to the need to determine sex at an early stage [9]. Sex identification is an important step in date palm cultivation, as it helps to identify the gender of an individual plant before planting the seedlings. Early-sex identification is used to select a balanced ratio of male and female plants for date palm orchards. By planting productive female plants, growers can reduce the need for the costly maintenance and pruning of unproductive date palm [10,11]. Several morphological, biochemical, and molecular characteristics can be used to identify the sex of dioecious plants at an early stage of development. It is often assumed that the percentage of male and female seedlings in the progeny is equal, which supports the hypothesis that sex in date palm is hereditarily determined [12].
Male and female date palm plants of the same variety can exhibit morphological differences. In general, male date palm plants tend to have a larger leaf length than female plants. However, these morphological differences can be influenced by environmental factor such as soil conditions, water availability, and temperature [13]. The biochemical analysis of date palm leaves does not provide much information about the sex identification of immature plants. The sex of date palm is determined genetically Cytological studies such as chromomycine A3 (CMA3) staining have also been used for sex determination, which depict sex as a genetic factor. The staining technique can reveal differences in the structure and composition of chromosomes, which can help to identify the sex [14]. Over the past decade, various biomarkers, including AFLP, RAPD and microsatellite, have been used to differentiate the sex of date palm plants [15].
Gene expression profiling is the most significant and effective technique used to determine an organism’s gene function. In plant biology, a variety of expression profiling techniques have been utilized, and technological development is still ongoing. Additionally, hybridization, sequencing, and polymerase chain reaction (PCR) are three key methods that support these widespread techniques and practices [16]. Real-time polymerase chain reaction (RT-PCR) is one of the best techniques used to evaluate different levels of gene expression. When a certain gene is amplified using gene-specific primers, the fluorescent signals allow for real-time measurements of the target gene level [17]. It has been reported that the TPD1 gene is required for the development of tapetal cells in Arabidopsis anther. Analysis of the male-sterile mutant, tpd1 showed that the functional interruption of TPD1 caused the precursors of tapetal cells to differentiate and develop into microsporocytes instead of tapetum and led to the formation of extra microsporocytes, and tapetum was found to be absent in developing anthers. TPD1 plays an important role in the differentiation of tapetal cells, possibly in coordination with the EMS1/EXS gene product, a Leu-rich repeat receptor protein kinase. It was observed that TPD1 is initially expressed in leaves and young seedlings [18].
The TPD1-like expression in leaves makes it possible to differentiate male and female plants from unknown seedlings, which would be advantageous for early sex identification in date palm [19]. In the current study, we used PCR-based techniques to perform expression profiling of the TPD1-like gene for sex determination in specific date palm cultivars. The objective of the study was to find a reliable, stable and durable technique for sex determination in date palm to assist farmers and breeders.
2. Materials and Methods
2.1. Plant Material
The leaf samples from suckers of both female and male date palm plants and unknown seedlings of three selected genotypes (Ajwa, Amber, and Medjool) were collected from Horticulture Research Institute, Bahawalpur, Pakistan to evaluate the differential expression of certain sex-determining genes. Date palm leaf tissues from the studied genotypes were thoroughly cleaned in RNase-free water, then dried and frozen for further extraction of RNA and DNA.
2.2. DNA Sequence Retrieval and Primer Designing
The TPD1-like mRNA sequence of Phoenix dactylifera was downloaded from the online portal of the NCBI database with accession number XM-026801684.2 (https://www.ncbi.nlm.nih.gov/ accessed on 11 March 2022). The nucleotide sequence of TPD1-like was translated into amino acid sequences through the EXPASY tool (https://web.expasy.org/translate/ accessed on 11 March 2022). PCR primers for TPD1-like gene were designed using the software Amplify version 1.7.0. Primer sequences are shown in (Table 1).
2.3. Extraction of DNA
The leaf tissue of 300 mg frozen date palm cultivars was ground in 700 μL CTAB extraction buffer until a homogenous mixture. The sample was shifted to a 1.5 mL Eppendorf tube and 50 μL of 2-β-mercaptoethanol was added. This mixture was incubated in water bath for 15 min at 65 °C. Then, the incubated sample was centrifuged at 14,000 rpm for ten minutes. Then, the supernatant was separated and shifted to another clean microfuge tube, and 500 μL of chloroform: Isoamyl Alcohol (24:1) was added and mixed by gentle inversion for 20 min. After mixing, the tube was centrifuged at 14,000 rpm for 10 min. The supernatant was transferred into a new tube, then 500 μL of cold absolute isopropanol was added and mixed carefully by gently inversing the Eppendorf tube. The mixture was incubated for 2 h at −20 °C. The tube was centrifuged at 12,000 rpm for 10 min, and the supernatant was discarded. The DNA pellet was washed twice with 100 μL ice cold 75% ethanol with subsequent centrifugation at 5000 rpm for 5 min. The DNA pellet was air dried and resuspended in 30 μL of distilled water and was stored at −20 °C [20].
2.4. RNA Extraction and cDNA Synthesis
Leaf samples kept in liquid nitrogen were crushed to powder for total RNA extraction. RNA extraction of selected samples was performed using TRIzol method [21]. The sample was ground in liquid nitrogen; then, 500 μL of RNA reagent was added and kept on ice for 5 min and spun for 5 min. Then, 500 μL NaCl (5M) + 300 μL chloroform was added to supernatant. Then, 200 μL of chilled isopropanol was added to the solution and held on ice for 10 min. The supernatant was discarded after a 10 min spin and the pellet was resuspended in 200 μL of 70% ethanol and spun for 1–2 min. The liquid was removed by rotating the pellet for 5–6 s while retaining it inside the tube. The pellet was held to dry for 10–15 min. Finally, 30 μL of DEPC water was added to dissolve the pellet. The isolated RNA was used to synthesise cDNA using the cDNA synthesis kit (Thermo Scientific Inc., Waltham, MA, USA) following the manufacturer’s protocols.
2.5. Polymerase Chain Reaction
The PCR was conducted using specific primers designed for TPD1-like gene. DreamTaq Green PCR Master Mix (2X) (Thermo Scientific Inc., Waltham, MA, USA) was used to prepare PCR reaction following the manufacturer’s protocol. The PCR was performed under the following conditions: initial denaturation at 95 °C for 3 min, followed by 30 cycles of 95 °C for 1 min, 55 °C for 1 min, and 72 °C for 50 s, with final extension at 72 °C for 10 min [22]. Amplicons were analyzed on 1.5% agarose gel using 1× TAE buffer. The integrity of gel bands was checked under UV light using the gel documentation system.
2.6. Semi qPCR Analysis of TPD1-like Gene
A reaction mixture (20 µL) of semi-quantitative PCR contained 1 µL template (cDNA reaction product), 1.6 µL MgCl2, 2 µL PCR buffer solution, 1 µL dNTP’s mixture, 0.2 µL DNA Taq Polymerase, 1 µL of each primer and 13.2 µL PCR-grade water. PCR program and further procedures were similar to those in Section 2.5.
2.7. Real-Time PCR Analysis of TPD1-like Gene
Real-time PCR was performed using real-time system (Bio-Rad Laboratories, Inc., Hercules, CA, USA) with SYBR Green as the reaction mixture, following the manufacturer’s instructions. The reaction mixture (20 µL) contains 5 µL SYBR Green, 0.5 µL of each primer (forward and reverse), 1 µL template of cDNA and 13 µL PCR-grade water. Conditions for amplification were as follows: initial denaturation for 8 min at 95 °C; 36 cycles of quantification consisting of denaturation step for 45 s at 95 °C, annealing for 30 s at 60 °C and extension for 30 s at 72 °C. Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was used as an internal control gene for the relative expression analysis. Expression was calculated using the 2−ΔΔCt method [23].
2.8. In Silico Analysis of TPD1-like Gene
2.8.1. Functional Annotation of Protein
An amino acid sequence of TPD1-like protein was analyzed using various bioinformatics tools, including TMHMM (https://services.healthtech.dtu.dk/service.php?DeepTMHMM accessed on 31 March 2022), PROTTER (http://wlab.ethz.ch/protter/ accessed on 31 March 2022), SignalP (https://services.healthtech.dtu.dk/service.php?SignalP-5.0 accessed on 31 March 2022) and NetPhos (http://cbs.dtu.dk/services/NetPhos/ accessed on 31 March 2022), to evaluate biochemical and physical characteristics.
2.8.2. Subcellular Localization and Signal Peptide Determination
The SignalP (https://services.healthtech.dtu.dk/service.php?SignalP-5.0 accessed on 1 April 2022) database, which predicts the existence of signal peptides and the locations of their cleavage sites in proteins, was used to determine the position of signal sequence in TPD1-like gene [24].
2.8.3. Protein Structure Analysis
The STRING database was used (https://string-db.org accessed on 1 April 2022) to integrate all known and predicted associations between proteins. These connections consist of direct (physical) and indirect (functional) interactions discovered by quantitative modelling, the information conveyed between species, and interactions discovered in other (primary) databases.
2.8.4. Glycosylation Sites Prediction
The web servers GlycoEP (http://www.imtech.res.in/raghava/glycoep accessed on 1 April 2022) and NetNGly (http://www.cbs.dtu.dk/services/NetNGlyc/ accessed on 1 April 2022) were used to more accurately evaluate N-linked, O-linked, and C-linked glycosides in eukaryotic glycoproteins.
2.8.5. Acetylation and Phosphorylation Sites
NetPhos 3.1 (http://www.cbs.dtu.dk/services/NetPhos/ accessed on 5 April 2022) webtoolwas used to examine acetylation and phosphorylation sites. NetAcet 1.0 server (http://www.cbs.dtu.dk/services/NetAcet/ accessed on 5 April 2022) was also used to examine protein acetylation and phosphorylation sites. This included both generic and kinase-specific protein prediction.
2.8.6. Sequence Analysis of Promoter
The promoter date palm TPD1-like gene (XM_026801684.1) was taken from NCBI (http://www.ncbi.nlm.nih.gov/ accessed on 15 April 2022). In BLAST searches, the BLASTx was carried out on the upstream regulatory regions of selected genes. The start codon of the TPD1 gene was identified and, using several bioinformatics tools, an upstream gene region of about 2.5 kb was evaluated. The chosen sequence was shown to be a non-coding region. About 2 kb of the of TPD1-promoter region was identified for isolation and investigated to confirm the noncoding sequences.
Each promoter’s transcription initiation site was located via the BDGP (https://www.fruitfly.org/seq_tools/promoter.html accessed on 15 April 2022) webtool. PlantCARE and plantPAN can be used to identify cis-regulatory elements (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/ accessed on 15 April 2022). PlantPAN 3.0 (http://plantpan.itps.ncku.edu.tw/promoter.php accessed on 15 April 2022) databases were used. Other important regulatory features in the promoter sequence, such as CpG islands and tandem repeats, were also determined by PlantPAN.
3. Results
3.1. TPD1-like Gene Amplification on DNA of Selected Date Palm Suckers
PCR amplification was carried out using a primer designed from the TPD1-like gene (Table 1) on the DNA of selected suckers whose sex was known (male and female plants) to confirm its presence in date palm. The TPD1-like gene was amplified in the DNA obtained from the leaves of male and female date palm plants of three selected genotypes, confirming its presence in date palm (Figure 1).
3.2. Semi qPCR Analysis of TPDI-like Gene on the Leaves of Date Palm Suckers
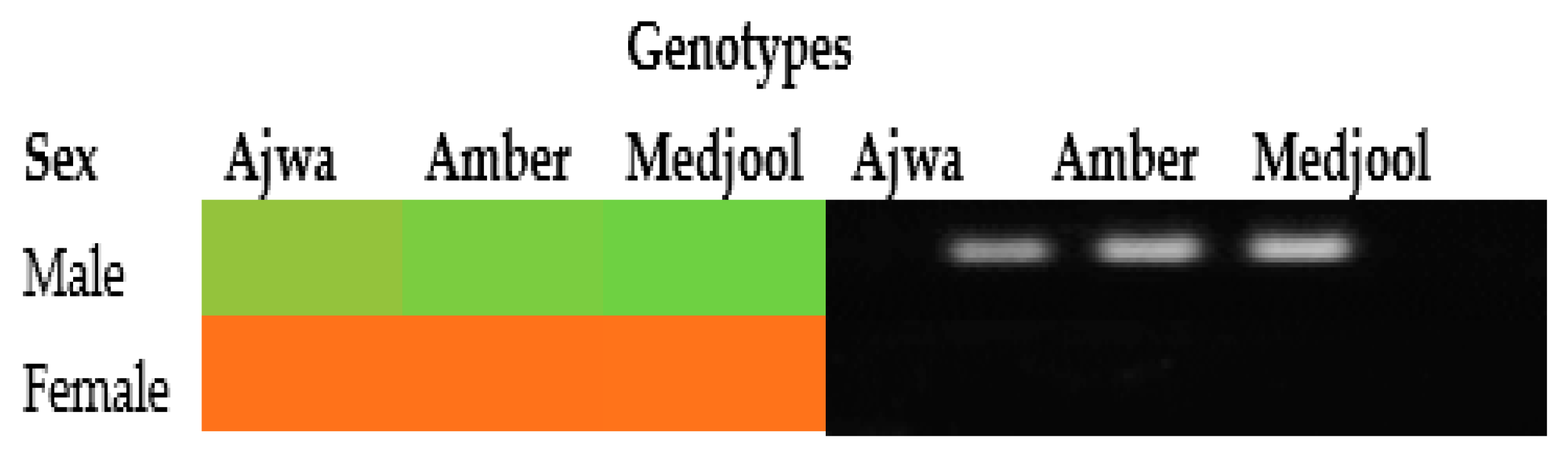
To validate the TPD1-like gene for early sex determination at seedling stage in date palm, its expression pattern was determined in the leaves of date palm suckers whose sex was known. It was shown to have varied expression in the leaves of three male genotypes (Ajwa, Amber and Medjool) and showed no expression in the leaves of these female genotypes. The semi-qPCR results were analyzed using ImageJ software [25] to generate a heat map. The heat map generated from this analysis can provide valuable information on the expression of the TPD1-like gene in the leaves of selected male and female genotypes through varied color intensity. The dark green color denoted higher expression and orange color denoted no expression (Figure 2).
3.3. Real-Time PCR Analysis of TPD1-like Gene on the Leaves of Date Palm Suckers
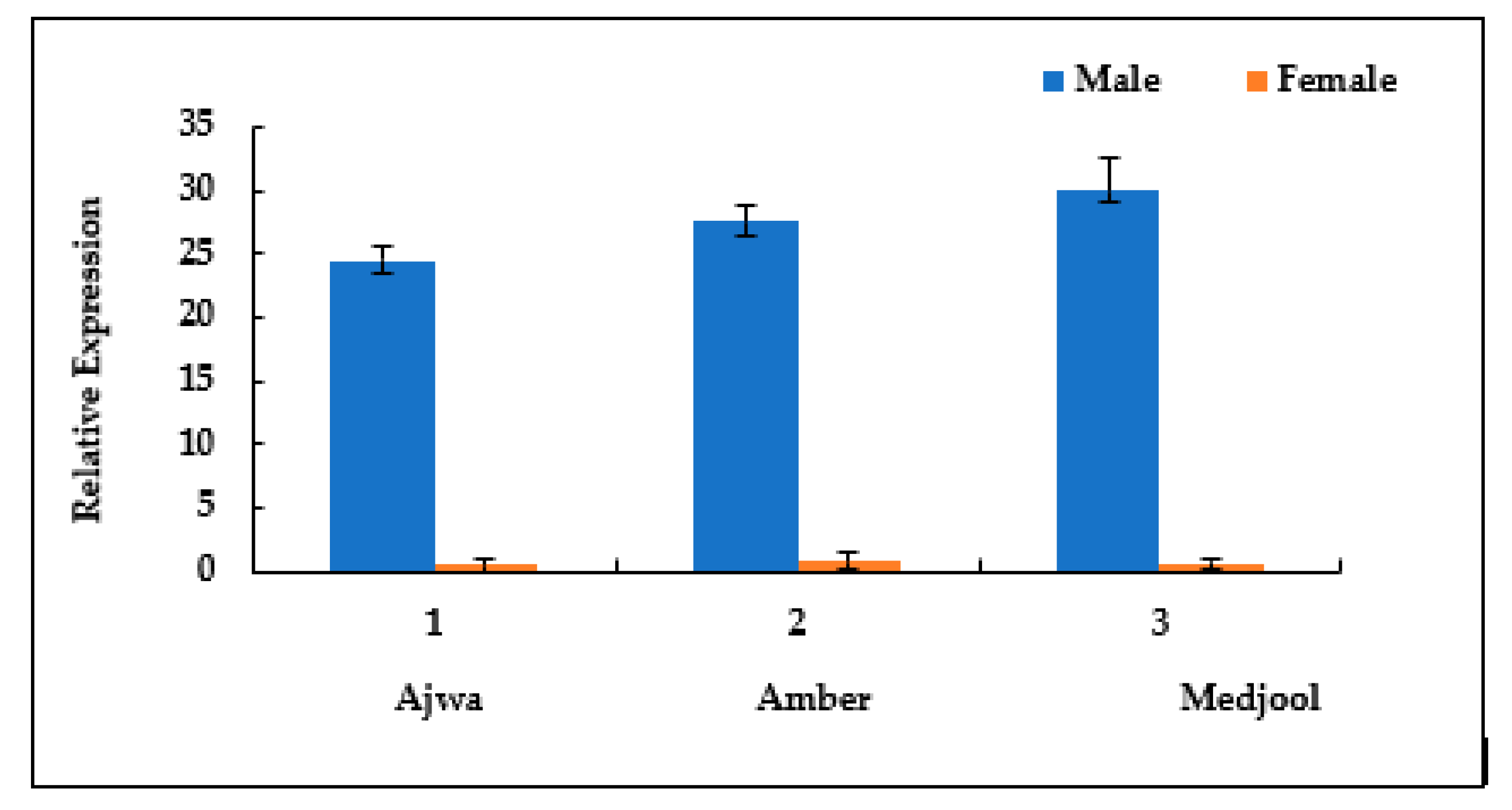
From semi-qPCR, the selected leaf tissue of date palm was further analyzed through qPCR, and an expression graph was made that showed that the TPD1-like gene was involved in tapetum development and only expressed in male date palm leaves. A higher relative expression was noted in male Medjool plants as compared to male Ajwa and Amber male and no relative expression was observed in female plants of the selected genotypes (Figure 3).
3.4. Semi-qPCR Analysis of TPD1-like Gene on the Leaves of Unknown Seedling of Date Palm
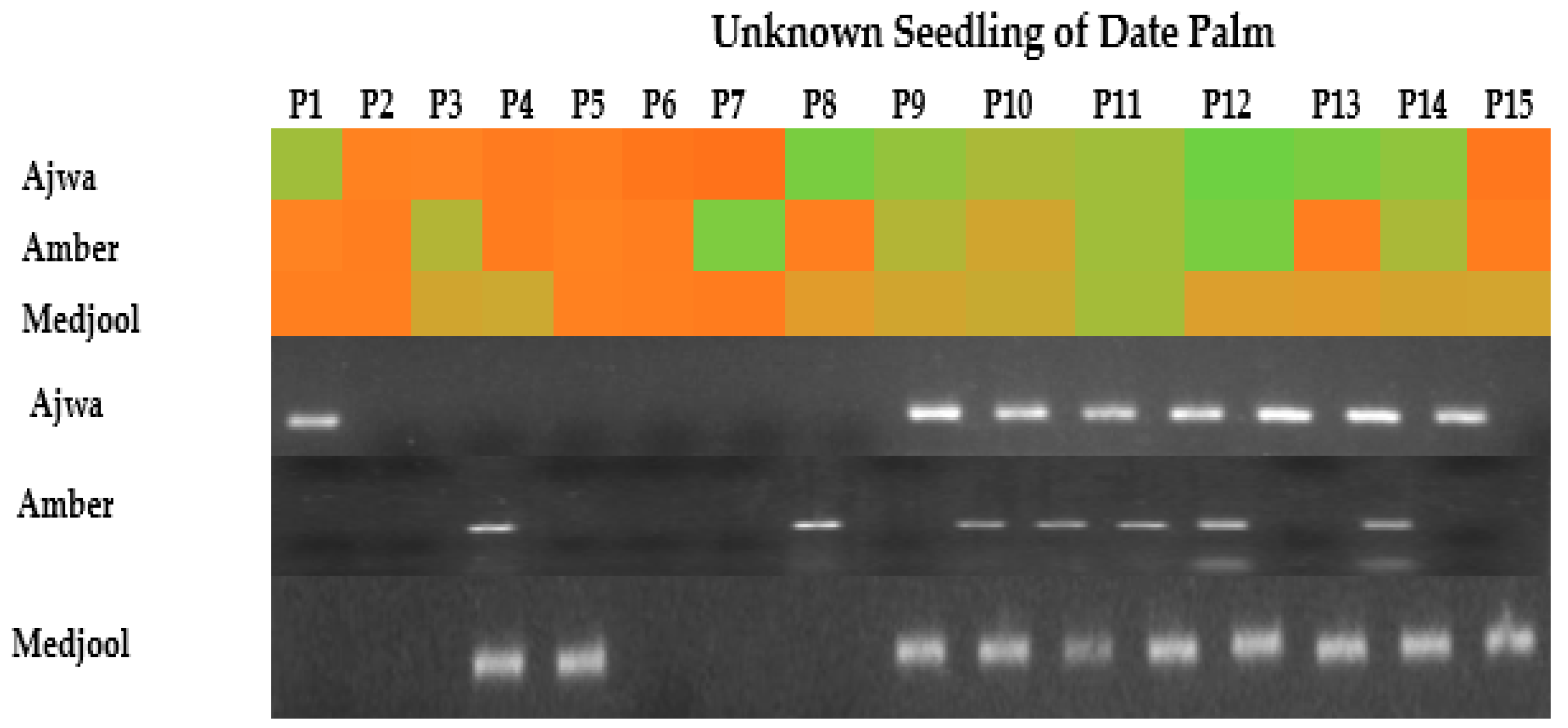
The unknown seedlings of Amber, Medjool, and Ajwa were identified on the basis of TPD1-like expression analysis. Male plants showed an enhanced expression of TPD1-like while female plants of all selected genotypes showed no expression. The heat map generated for unknown date palm seedlings showed varied color expression in selected unknown seedlings of Ajwa, Amber and Medjool plants. In the heat map, a light green to green color showed low–high expression, and orange showed no expression. In the case of Ajwa, out of fifteen seedlings 1, 8, 9, 10, 11, 12, 13 and 14 showed a varied color expression from light green to green for TPD1-like and were considered male plants, while seedlings 2, 3, 4, 5, 6, 7 and 15, showed an orange color and were recorded as female plants. Similarly, unknown Amber seedlings showed a varied color intensity for TPD1-like; plant numbers 3, 7, 9, 10, 11, 12 and 14 were screened as male, showing a light green and green color, while plants with numbers 1, 2, 4, 5, 6, 8, 13 and 15 had no expression, showing an orange color, and were assumed to be female plants. Plants 1, 2, 5, 6, and 7 for Medjool seedlings showed an orange color and were female while the rest of the plants showed a light green to green color and were considered male plants (Figure 4).
3.5. Real-Time PCR Analysis of TPD1-like Gene on the Leaves of Unknown Seedling of Date Palm
The relative expression of selected unknown seedlings was recorded. Ajwa seedlings with the number 1, 8, 9, 10, 11, 12, 13, 14 showed some expression and were considered male plants, while the rest of the unknown seedlings among the fifteen plants were female plants showing no relative expression (Figure 5A). Unknown Amber seedlings 3, 7, 9, 10, 11, 12 and 14 showed relative expression and were considered male plants (Figure 5B). The Medjool unknown seedlings 3, 4, 8, 9, 10, 11, 12, 13, 14, 15 showed relative expression and were considered male plants, while the rest of the unknown seedlings were considered as female plants showing no expression (Figure 5C).
3.6. In Silico Analysis of TPD1-like Gene
3.6.1. Coding Sequence of TPD1-like Gene
BLASTp was carried out for the sequence identity of TPD1-like genes. EXPASY (https://web.expasy.org/translate/ accessed on 11 March 2022) was used to translate the TPD1-like gene sequence using six reading frames (Figure 6 and Figure 7).
3.6.2. Subcellular Localization
SignalP was used to predict signal peptides from amino acid sequences. Proteins with signal peptides are targeted to the secretory pathway but are not necessarily secreted. The neural networks in SignalP generated three output scores (C-score, Y-score and S-score) for each position in the input sequence. The findings demonstrate that a similar protein lacks a signal peptide. The output scores and their positions were graphically represented. There is no peptide signal when the C, S, and Y score values are less than 0.45. The sequence from the upstream region of the coding sequence contains a signal peptide with a value of 0.0009 (Sec/SPI), indicating that the signal peptide is located outside the membrane (Figure 8).
3.6.3. Signal Peptide Analysis
The TMHMM membrane protein topology prediction method describes transmembrane helices and various proteins embedded in the cellular membrane. TMHMM analysis showed that TPD1 protein had no transmembrane proteins, as shown in (Figure 9). The prediction provides the most likely position and direction of TM helices in the sequence. The plot represents the possibilities of the protein both inside and outside of the trans membrane helix.
3.6.4. Acetylation and Phosphorylation Sites
The probable N- phosphorylation sites in protein sequence are predicted using the NetPhos 3.1 server. Phosphorylation is an effective mechanism through which protein activity can be altered once established. Netphos result shows the phosphorylation sites in the TPD1-like protein sequence with threshold value 0.5 (Figure 10). Netphos analysis showed that protein has many phosphorylation sites.
3.6.5. Promoter Analysis of TPD1-like Gene
A promoter analysis of the TPD1-like gene showed predicted transcription initiation site (TIS) with probabilities of 88, 96, 98, 80 and percentages at positions of 33, 150, 209, 294 and 746, respectively. TIS is shown in bold letters in all sequences (Figure 11).
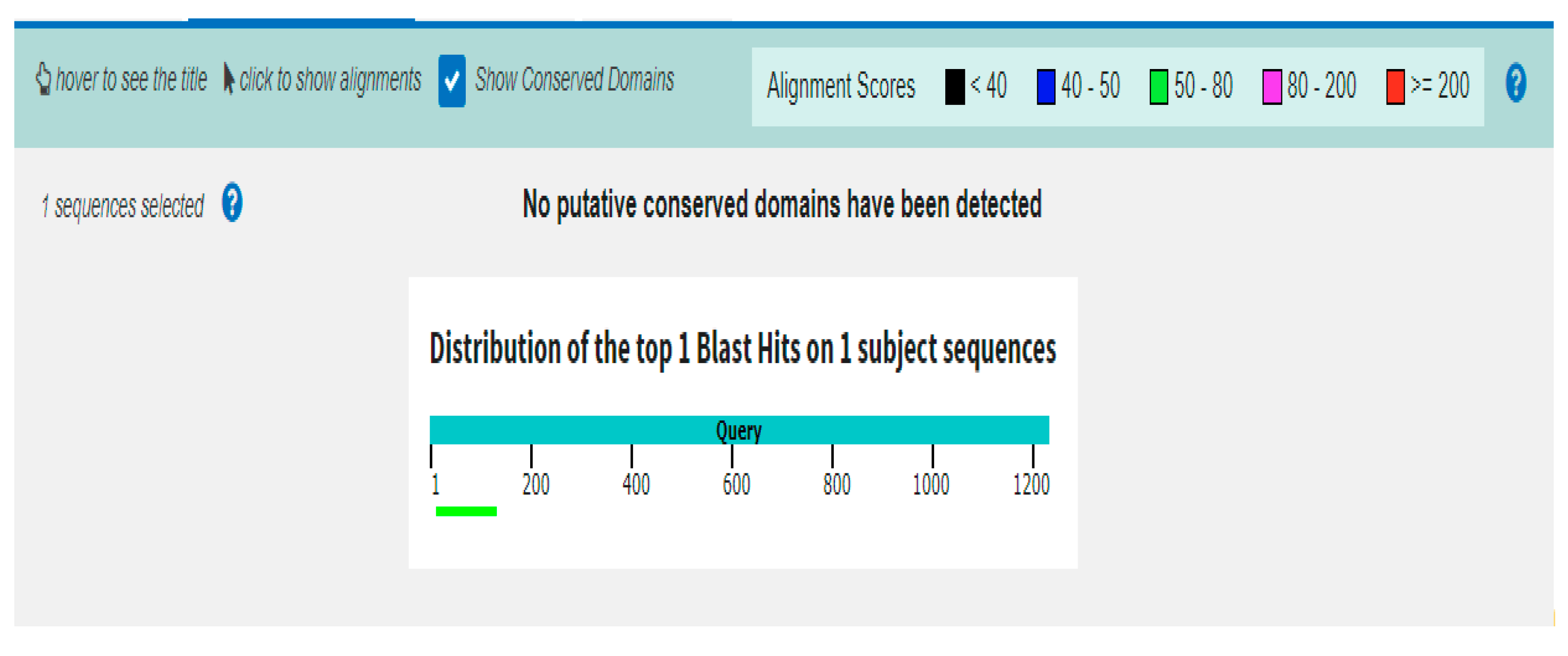
3.6.6. BLASTx Analysis of TPD1-like Promoter Sequence
BLASTx results show that the promoter sequence has no similarity with the coding sequence or protein, as shown in Figure 12.
3.6.7. PLANTPAN Analysis of Promoter
PLANTPAN analysis shows various transcription factor binding sites in the promoter region of TPD1-like promoter (Figure 13).
3.6.8. Promoter Cis-Acting Regulatory Elements’ Analysis
Using the PLANTCARE database, other cis-regulatory elements were also discovered to be present in the TPD1 gene’s promoter. PlantCARE analysis shows the expression of different regulatory elements of TPD1-like. These regulatory elements include AFI, ARE, TCA element, p-box, CCAATT-box, TATA-box, CAAT-box, and TATC-box, as shown in (Figure 14).
The full-length TPD1-like promoter showed binding sites for various transcription factors (Table 2). The full TPD1 promoter fragment containing core promoter elements, including a CAAT box with consensus sequence CAAAT and CAAT, were also reported in Pisum sativum and Nicotiana glutinosa. Some light-inducible motifs (G box, AE box, sp1 box) and enhancer elements (CGTCA-motif) were found through bioinformatics analysis.
4. Discussion
The sex identification of date palm seedlings is difficult to distinguish before the first flowering stage. Different morphological parameters plant height, leaf length, leaf width and number of leaves have been used to identify the sex of dioecious plants, but these parameters are environmentally dependent. Biochemical studies provide little information about the identification of sex in date palm plants [26]. Molecular techniques offer potential for sex determination as they are stable and independent of age and environment [27]. In the current study, it was found that the TPD1-like gene has the potential for early sex determination in different date palm genotypes. Our research suggests that this gene could be a successful approach to determine the sex of date palms at the early seedling stage. Ajwa, Amber and Medjool male plants showed enhanced expression of TPD1-like. It was found that female date palm suckers and seedlings of these genotypes did not express the TPD1-like gene, which was only expressed in the leaves of male plants. Based on the relative expression of TPD1-like, unknown date palm seedlings were differentiated as male and female plants. The TPD1-like was markedly expressed in Arabidopsis thaliana’s leaves, seedlings, flowering buds, and anthers. TPD1-like serves as a ligand for EMS1 receptor kinase to signal cell fate determination during plant sexual reproduction. Its ectopic expression causes the abnormal differentiation of somatic and reproductive cells in anthers. Torres et al. [28] proposed two single copy genes, CYP-703 and GPAT3, the were present in males, involved in male flower development, and absent in females. Our approach to the gene-specific identification of date palm plants at the seedling stage represents a significant success. Transcription factors are proteins that bind to certain DNA sequences to regulate the transcriptional level of gene expression. Therefore, an important aspect of the functional analysis of transcription factors is predicting the DNA-binding motifs. The precise control of gene expression depends on the presence of cis-regulatory elements, and genomic sequences in gene-promoter regions to which transcription factors bind. Due to the central role that these transcription factors play during development, it is necessary to identify anther-development-specific genes that produce transcription factors [29]. Several studies have been proposed regarding the utilization of molecular markers for distinguishing between male and female date palm cultivars, including that of Bekheet and Hanafy [30], Al-Dous et al. [31], Al-Mahmoud et al. [32], Elmeer and Mattat [33], Zhao et al. [34], and Cherif et al. [35]. However, the accuracy of discrimination between sex significantly varies; they are more sensitive and require careful handling and a greater number of repeated experiments. Wang et al. [36] used an SSR marker to differentiate sex in date palm using the qPCR technique. Intha and Chaiprasart [37] reported DNA markers for sex identification in date palm that could differentiate male and female seedlings. Khan et al. [38] proposed reliable and authentic spectroscopic methods that used chemo-informatics regression analysis to discriminate among male and female plants. Our gene-based sex identification of date palm seedlings is a more efficient and time-saving technique. It was found that plant MYB proteins have an impact on a number of developmental processes, as well as signaling and secondary metabolisms [39]. Anthers and pollen contain several genes that could encode transcription factors containing MYB-related genes specified to tapetum from Arabidopsis [40]. A similar MYB motif is identified in the TPD1-like promoter analysis, demonstrating its significance in the development of male characters. The expression profiling of genes is a major tool to identify sex at the seedling stage. A few studies provide evidence for the expression of the TPD1-like gene in male plants, which supports the idea that this gene is involved in the development of the male character. It was observed that modifying the TPD1-like gene in the T-DNA causes metabolic and structural changes, and even male sterility [41].
5. Conclusions
The current research is an inclusive study of the identification of sex in date palm using TPD1-like gene expression analysis. One-year-old unknown date palm seedlings were identified as male and female plants on the basis of the expression of the TPD1-like gene. These findings would support effective genetic resource management, selection, and breeding strategies for date palm. It was concluded that the TPD1-like gene is a promising gene to identify the sex of date palm.
Author Contributions
Conceptualization and design of research work: A.A.K., A.G., I.A.K., P.K. and Z.K.; execution of the experiments: P.K., A.G. and Z.K.; data curation and statistical analysis: P.K. and Z.K.; write-up of manuscript: P.K., Z.K., A.G. and I.A.K. The final draft of the work has been read and approved for publication by all authors. All authors have read and agreed to the published version of the manuscript.
Funding
The current research was supported by the departmental funds provided by Institute of Plant Breeding & Biotechnology, MNS University of Agriculture Multan Pakistan.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The corresponding author can provide access to all the data that was used and examined in the current field investigation.
Acknowledgments
The current work was carried out as a part of doctoral thesis of Plosha Khanum, Institute of Plant Breeding & Biotechnology, MNS University of Agriculture Multan Pakistan. The authors are thankful to the SINO-PAK Joint Research Lab, Institute of Plant Breeding & Biotechnology and the Horticulture Research Institute Bahawalpur for the provision of lab facility, plant material and technical support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dransfield, J.; Baker, W.J.; Harley, M.M.; Asmussen, C.; Lewis, C.E. Genera Palmarum, the Evolution and Classification of Palms; Royal Botanic Gardens: London, UK, 2008. [Google Scholar]
- Al-Khalifha, N.; Askari, S.E.; Khan, A.S. Molecular and Morphological identification of some elite varieties of date palms in Saudi Arabia. Emir. J. Food Agric. 2012, 24, 456–461. [Google Scholar]
- Ghnimi, S.; Syed, U.; Azharul, K.; Kamal-Eldin, A. Date fruit as food security crop in the arid lands seeking industrial valorization. Nutr. Food Sci. J. 2017, 5, 10–15. [Google Scholar]
- Yaish, M.W.; Kumar, P.P. Salt tolerance research in date palm tree (Phoenix dactylifera L.), past, present and future perspectives. Front. Plant Sci. 2015, 6, 348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Farsi, M.A.; Lee, C.Y. Nutritional and functional properties of dates: A review. Crit. Rev. Food Sci. Nutr. 2008, 48, 877–887. [Google Scholar] [CrossRef] [PubMed]
- Abdullahi, M.; Garko, M. Medicinal value of date palm (Phoenix dactylifera L. In ). In Proceedings of the Agricultural Society of Nigeria Conference, Nsukka, Nigeria, 11–14 March 2012. [Google Scholar]
- Khalid, S.; Khalid, N.; Khan, R.S.; Ahmed, H.; Ahmad, A. A review on chemistry and pharmacology of Ajwa date fruit and pit. Trends Food Sci. Tech. 2017, 63, 60–69. [Google Scholar] [CrossRef]
- Salomón-Torres, R.; Ortiz-Uribe, N.; Valdez-Salas, B.; Rosas-González, N.; García-González, C.; Chávez, D.; Krueger, R. Nutritional assessment, phytochemical composition and antioxidant analysis of the pulp and seed of medjool date grown in Mexico. Peer J. 2019, 7, e6821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carpenter, J.B. Breeding of date palm in California. Date Grow. Inst. Rep. 1979, 54, 13–16. [Google Scholar]
- Aberlenc-Bertossi, F.; Daher, A.; Chabrillange, N.; Tregear, J.; Mohamed, N. Sex determination in date palm: New perspectives on an old theme. In Proceedings of the Plant and Animal Genomes XIX Conference, San Diego, CA, USA, 15–19 January 2011. [Google Scholar]
- Sarkar, S.; Banerjee, J.; Gantait, S. Sex-oriented research on dioecious crops of Indian subcontinent: An updated review. Biotechnol. J. 2017, 7, 93–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heikrujam, M.; Sharma, K.; Prasad, M.; Agrawal, V. Review on different mechanisms of sex determination and sex-linked molecular markers in dioecious crops: A current update. Euphytica 2015, 201, 161–194. [Google Scholar] [CrossRef]
- Ageez, A.; Madboly, E.A. Identification of male specific molecular markers in date palm sewi cultivar. Egypt. J. Genet. Cytol. 2011, 40, 201–214. [Google Scholar] [CrossRef] [Green Version]
- Siljak-Yakovlev, S.; Cerbah, M.; Sarr, A.; Benmalek, S.; Bounaga, N.; Coba de la Pena, T.; Brown, S.C. Chromosomal sex determination and heterochromatin structure in date palm. Sex. Plant Reprod. 1996, 9, 127–132. [Google Scholar] [CrossRef]
- El-Kharbotly, A.; El-Mardi, M.; Al-Saadi, N.; Al-Maharuki, Y. Towards the construction of a genetic map of date palm using the amplified fragment length polymorphism technique (AFLP). In Proceedings of the First International Conference on Date Palm, Al-Ain, United Arab Emirates, 8–10 March 1998. [Google Scholar]
- Lohman, B.K.; Stutz, W.E.; Bolnick, D.I. Gene expression stasis and plasticity following migration into a foreign environment. Mol. Ecol. 2017, 26, 4657–4670. [Google Scholar] [CrossRef]
- Garson, J.; Grant, P.R.; Ayliffe, U.; Ferns, R.B.; Tedder, R.S. Real-time PCR quantitation of hepatitis B virus DNA using automated sample preparation and murine cytomegalovirus internal control. J. Virol. Methods 2005, 126, 207–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.L.; Xie, L.F.; Mao, H.Z.; Puah, C.S.; Yang, W.C.; Jiang, L.; Ye, D. Tapetum determinant1 is required for cell specialization in the Arabidopsis anther. Plant Cell 2003, 15, 2792–2804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, G.; Liu, X.; Owen, H.A.; Zhao, D. Signaling of cell fate determination by the TPD1 small protein and EMS1 receptor kinase. Proc. Natl. Acad. Sci. USA 2008, 105, 2220–2225. [Google Scholar] [CrossRef] [Green Version]
- Aboul-Maaty, N.A.F.; Oraby, H.A.S. Extraction of high-quality genomic DNA from different plant orders applying a modified CTAB-based method. Bull. Natl. Res. Cent. 2019, 43, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Han, R. A method for extracting high-quality total RNA from plant rich in polysaccharides and polyphenols using Dendrobium huoshanense. PLoS ONE 2018, 13, e0196592. [Google Scholar] [CrossRef]
- Lorenz, T.C. Polymerase chain reaction: Basic protocol plus troubleshooting and optimization strategies. J. Vis. Exp. 2012, 63, e3998. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Armenteros, J.J.; Salvatore, A.; Emanuelsson, M.; Winther, O.; Von Heijne, G.; Elofsson, A.; Nielsen, H. Detecting sequence signals in targeting peptides using deep learning. Life Sci. Alliance 2019, 2, e201900429. [Google Scholar] [CrossRef] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Qacif, N.; Baaziz, M.; Bendiab, K. Biochemical investigations on peroxidase contents of male and female inflorescences of date palm (Phoenix dactylifera). Sci. Hortic. 2007, 114, 298–301. [Google Scholar] [CrossRef]
- Dhawan, C.; Kharb, P.; Sharma, R.; Uppal, S.; Aggarwal, R.K. Development of male-specific SCAR marker in date palm (Phoenix dactylifera L.). Tree Genet. Genomes 2013, 9, 1143–1150. [Google Scholar] [CrossRef]
- Torres, M.F.; Mathew, L.S.; Ahmed, I.; Al-Azwani, I.K.; Krueger, R.; Rivera-Nuñez, D.; Malek, J.A. Genus-wide sequencing supports a two-locus model for sex-determination in Phoenix. Nat. Commun. 2018, 9, 3969. [Google Scholar] [CrossRef] [Green Version]
- Mitsis, T.; Efthimiadou, A.; Bacopoulou, F.; Vlachakis, D.; Chrousos, G.P.; Eliopoulos, E. Transcription factors and evolution: An integral part of gene expression. World Acad. Sci. J. 2020, 2, 3–8. [Google Scholar] [CrossRef] [Green Version]
- Bekheet, S.A.; Hanafy, M.S. Towards sex determination of date palm. Arab J. Biotechnol. 2011, 26, 551–566. [Google Scholar]
- Al-Dous, E.K.; George, B.E.; Al-Mahmoud, M.; Al-Jaber, M.Y.; Wang, H.; Salameh, Y.M.; Al-Azwani, E.K.; Chaluvadi, S.R.; Pontaroli, A.C.; DeBarry, J. De novo genome sequencing and comparative genomics of date palm (Phoenix dactylifera). Nat. Biotechnol. 2011, 29, 521–527. [Google Scholar] [CrossRef] [PubMed]
- Al-Mahmoud, M.E.; Al-Dous, E.K.; Al-Azwani, E.K.; Malek, J.A. DNA-based assays to distinguish date palm (Arecaceae) gender. Am. J. Bot. 2012, 99, 7–10. [Google Scholar] [CrossRef]
- Elmeer, K.; Mattat, I. Marker-assisted sex differentiation in date palm using simple sequence repeats. Biotechnol. J. 2012, 2, 241–247. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Williams, R.; Prakash, C.S.; He, G. Identification and characterization of gene-based SSR markers in date palm (Phoenix dactylifera L.). BMC Plant Biol. 2012, 12, 237. [Google Scholar] [CrossRef] [Green Version]
- Cherif, E.; Zehdi, S.; Castillo, K.; Chabrillange, N.; Abdoulkader, S.; Pintaud, J.C.; Santoni, S.; Salhi-Hannachi, A.; Glémin, S.; Aberlenc-Bertossi, F. Male-specific DNA markers provide genetic evidence of an XY chromosome system, a recombination arrest and allow the tracing of paternal lineages in date palm. New Phytol. 2013, 197, 409–415. [Google Scholar] [CrossRef]
- Wang, Y.; Ihase, L.O.; Htwe, Y.M.; Shi, P.; Zhang, D.; Li, D.; Iserhienrhien, A. Development of sex-linked SSR marker in the genus Phoenix and validation in P. dactylifera. Crop Sci. 2020, 60, 2452–2466. [Google Scholar] [CrossRef]
- Intha, N.; Chaiprasart, P. Sex determination in date palm (Phoenix dactylifera L.) by PCR based marker analysis. Sci. Hortic. 2018, 236, 251–255. [Google Scholar] [CrossRef]
- Khan, A.L.; Al-Harrasi, A.; Numan, M.; AbdulKareem, N.M.; Mabood, F.; Al-Rawahi, A. Spectroscopic and molecular methods to differentiate gender in immature date palm (Phoenix dactylifera L.). Plants 2021, 10, 536. [Google Scholar] [CrossRef]
- Jin, H.; Martin, C. Multifunctionality and diversity within the plant MYB-gene family. Plant Mol. Biol. 1999, 41, 577–585. [Google Scholar] [CrossRef] [PubMed]
- Li, S.F.; Higginson, T.; Parish, R.W. A novel MYB-related gene from Arabidopsis thaliana expressed in developing anthers. Plant Cell Physiol. 1999, 40, 343–347. [Google Scholar] [CrossRef] [Green Version]
- Albrecht, C.; Russinova, E.; Hecht, V.; Baaijens, E.; de Vries, S. The Arabidopsis thaliana somatic embryogenesis receptor-like kinases1 and 2 control male sporogenesis. Plant Cell 2005, 17, 3337–3349. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
TPD1-like gene amplification in leaves using date palm suckers’ DNA as template. PCR product was run on 1.5% agarose gel using a 100 bp ladder (Thermo Scientific Inc., Waltham, MA, USA). A band of 152 bps was observed on the gel. M representing male plants while F representing female plants.
Figure 1.
TPD1-like gene amplification in leaves using date palm suckers’ DNA as template. PCR product was run on 1.5% agarose gel using a 100 bp ladder (Thermo Scientific Inc., Waltham, MA, USA). A band of 152 bps was observed on the gel. M representing male plants while F representing female plants.

Figure 2.
TPD1-like amplification in the leaves using date palm sucker cDNA as a template. PCR product was run on 1.5% agarose gel using a 100 bp ladder (Thermo Scientific Inc., Waltham, MA, USA). The heat map indicated the expression intensity: green for high expression, light green for low expression and orange for no expression.
Figure 2.
TPD1-like amplification in the leaves using date palm sucker cDNA as a template. PCR product was run on 1.5% agarose gel using a 100 bp ladder (Thermo Scientific Inc., Waltham, MA, USA). The heat map indicated the expression intensity: green for high expression, light green for low expression and orange for no expression.

Figure 3.
Comparative TPD1-like gene expression in the leaves of selected date palm suckers. Error bar indicates the ±SE of three replicates.
Figure 3.
Comparative TPD1-like gene expression in the leaves of selected date palm suckers. Error bar indicates the ±SE of three replicates.

Figure 4.
TPD1-like gene amplification in leaves using date palm seedling cDNA as template. PCR product was run on 1.5% agarose gel, using a 100 bp ladder (Thermo Scientific Inc., Waltham, MA, USA). The heat map indicated the expression intensity: green for higher expression, light green for low expression and orange for no expression.
Figure 4.
TPD1-like gene amplification in leaves using date palm seedling cDNA as template. PCR product was run on 1.5% agarose gel, using a 100 bp ladder (Thermo Scientific Inc., Waltham, MA, USA). The heat map indicated the expression intensity: green for higher expression, light green for low expression and orange for no expression.

Figure 5.
Expression profiling of TPD1-like in unknown seedlings of date palm. Error bar indicates ±SE of three experimental replicates. (A) is showing expression of TPD1-like gene in Ajwa, (B) is showing expression of TPD1-like gene in Amber and (C) is showing expression of TPD1-like gene in Medjool.
Figure 5.
Expression profiling of TPD1-like in unknown seedlings of date palm. Error bar indicates ±SE of three experimental replicates. (A) is showing expression of TPD1-like gene in Ajwa, (B) is showing expression of TPD1-like gene in Amber and (C) is showing expression of TPD1-like gene in Medjool.

Figure 6.
The coding sequence of TPD1-like gene (XM_026801684.1) is highlighted.

Figure 7.
Translated protein sequence of TPD 1-like gene containing 132 amino acids (highlighted).

Figure 8.
Graphically signal peptide representation of TPD1-like protein.

Figure 9.
TransMembrane prediction using Hidden Markov Models (TMHMM) of TPD1-like protein.

Figure 10.
TPD1-like protein’s predicted phosphorylation locations.

Figure 11.
Predicted transcription start site in TPD1 promoter with % probability.

Figure 12.
BLASTx analysis of TPD1 promoter showing no coding region and conserved domains.

Figure 13.
Transcriptional factor binding sites in TPD1-promoter region.

Figure 14.
Promoter sequence of TPD1 gene; important motifs are highlighted in various colors.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Primer used for selected date palm suckers and unknown seedlings for PCR amplification.
| Gene Abbreviation | Name of Gene and Accession No. | Primer Length (bp) | Forward (5′-3′) | Reverse (5′-3′) |
|---|---|---|---|---|
| Tapetum Determinant 1-Like | PdTPD1-like (>XM_026801684.2) | 152 | ATCGGACGCATCCACCTCAG | GAGTTGGCGTACTGGAAGGACA |
Table 2.
Site name, position, sequence and function of primers of organisms.
| Site Name | Organism | Position Standard | Sequence | Function |
|---|---|---|---|---|
| ABRE | A. thaliana | +1030 | ACGTG | cis-acting component involved in abscisic acid responsiveness |
| ABRE | Hordeum vulgare | +205 | CGCACGTGTC | cis-acting component involved in abscisic acid responsiveness |
| ABRE | A. thaliana | +207 | CACGTG | cis-acting component involved in abscisic acid responsiveness |
| ABRE | A. thaliana | +208 | ACGTG | cis-acting element involved in the abscisic acid responsiveness |
| ABRE | A. thaliana | −547 | ACGTG | cis-acting component involved in abscisic acid responsiveness |
| AE-box | A. thaliana | +1195 | AGAAACAA | part of a module for light response |
| CAAT-box | P. sativum | +60 | CAAAT | common cis-acting element in promoter and enhancer regions |
| CAAT-box | P. sativum | +139 | CAAAT | cis-acting component involved in abscisic acid responsiveness |
| CAAT-box | N. glutinosa | +725 | CAAT | common cis-acting element in promoter and enhancer regions |
| CGTCA-motif | H. vulgare | −285 | CGTCA | cis-acting regulatory element involved in the MeJA-responsiveness |
| G-box | A. thaliana | +207 | CACGTG | cis-acting regulatory element involved in light responsiveness |
| G-box | Zea mays | −479 | CACGAC | cis-acting regulatory element involved in light responsiveness |
| GA-motif | A. thaliana | +763 | ATAGATAA | part of a light-responsive element |
| GC-motif | Z. mays | −231 | CCCCCG | enhancer-like element involved in anoxic specific inducibility |
| GC-motif | Z. mays | −371 | CCCCCG | enhancer-like element involved in anoxic specific inducibility |
| LTR | H. vulgare | −76 | CCGAAA | cis-acting element involved in low-temperature responsiveness |
| MBS | A. thaliana | +355 | CAACTG | MYB binding site involved in drought-inducibility |
| Myb | A. thaliana | +355 | CAACTG | Stress inducubility |
| Myc | A. thaliana | −1095 | TCTCTTA | Stress inducubility |
| P-box | Oryza sativa | +611 | CCTTTTG | gibberellin-responsive element |
| Sp1 | O. sativa | +416 | GGGCGG | Light-responsive element |
| TGACG-motif | H. vulgare | +285 | TGACG | cis-acting regulatory element involved in the MeJA-responsiveness |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Khanum, P.; Khan, A.A.; Khan, I.A.; Ghaffar, A.; Khan, Z. TPD1-like Gene as a Suitable Marker for Early Sex Determination in Date Palm (Phoenix dactylifera L.). Genes 2023, 14, 907. https://doi.org/10.3390/genes14040907
AMA Style
Khanum P, Khan AA, Khan IA, Ghaffar A, Khan Z. TPD1-like Gene as a Suitable Marker for Early Sex Determination in Date Palm (Phoenix dactylifera L.). Genes. 2023; 14(4):907. https://doi.org/10.3390/genes14040907
Chicago/Turabian StyleKhanum, Plosha, Asif Ali Khan, Iqrar Ahmad Khan, Abdul Ghaffar, and Zulqurnain Khan. 2023. "TPD1-like Gene as a Suitable Marker for Early Sex Determination in Date Palm (Phoenix dactylifera L.)" Genes 14, no. 4: 907. https://doi.org/10.3390/genes14040907
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.