
Nucleotide Context Can Modulate Promoter Strength in Genes Transcribed by RNA Polymerase III

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
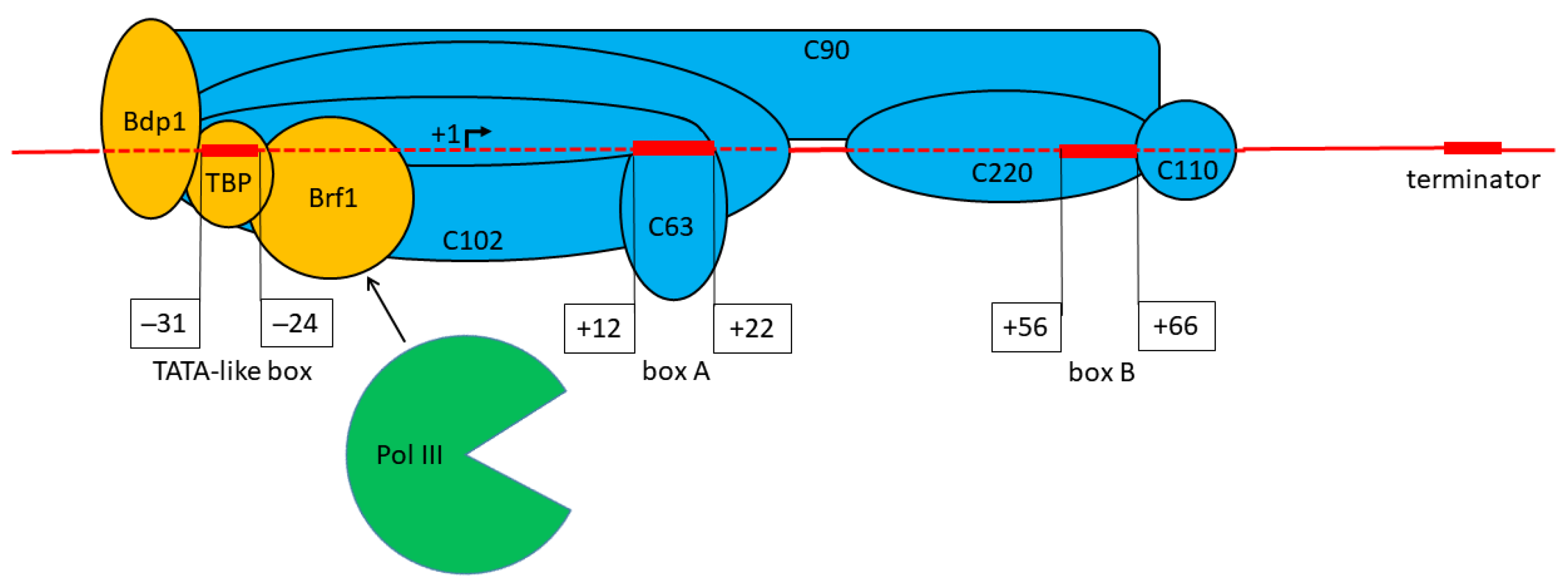
:1. Introduction
2. Materials and Methods
2.1. Plasmid Constructs
2.2. Cell Transfection and Northern Blot Analysis
3. Results and Discussion
3.1. Control Construct Transcribed by RNA Polymerase II
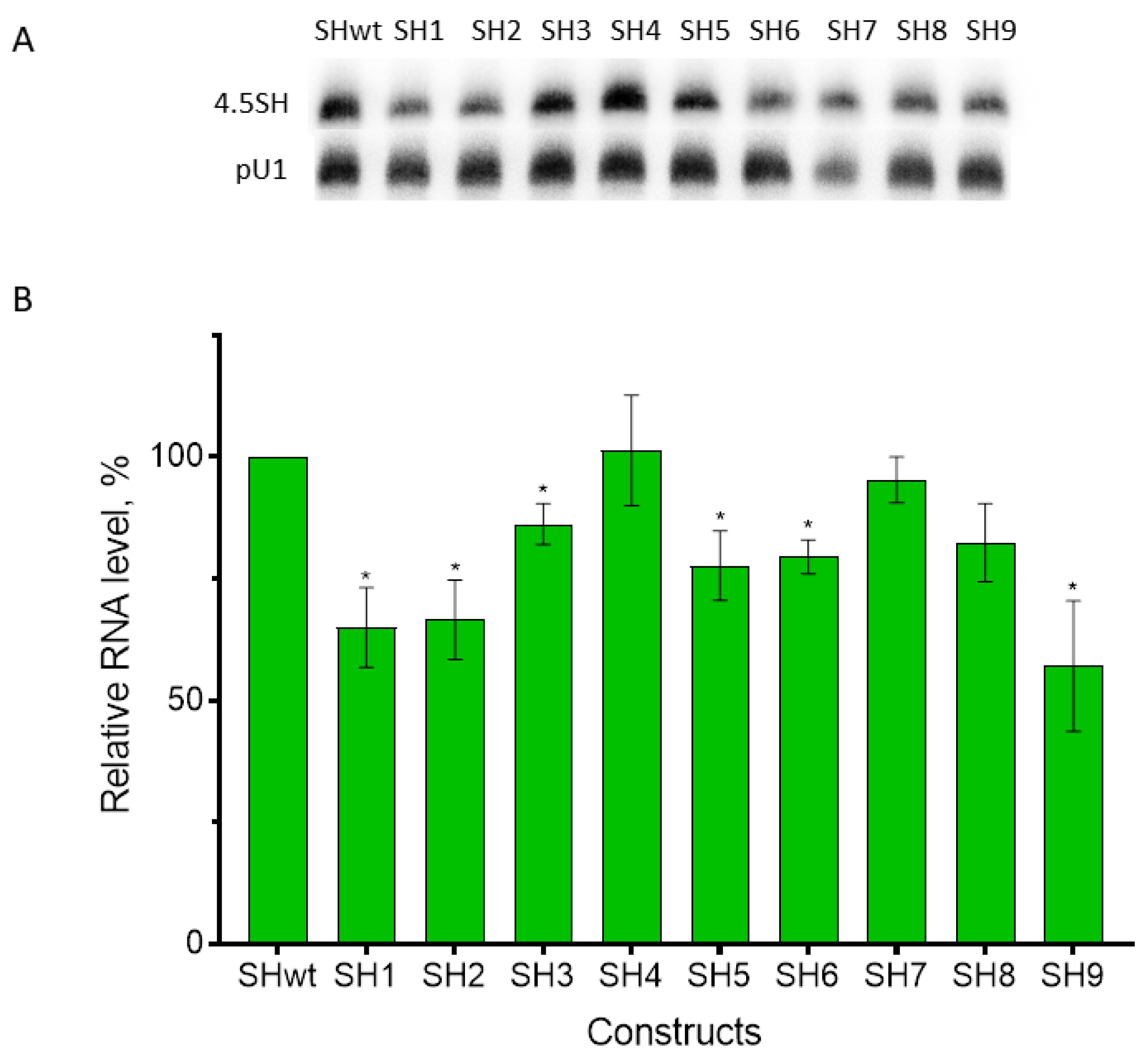
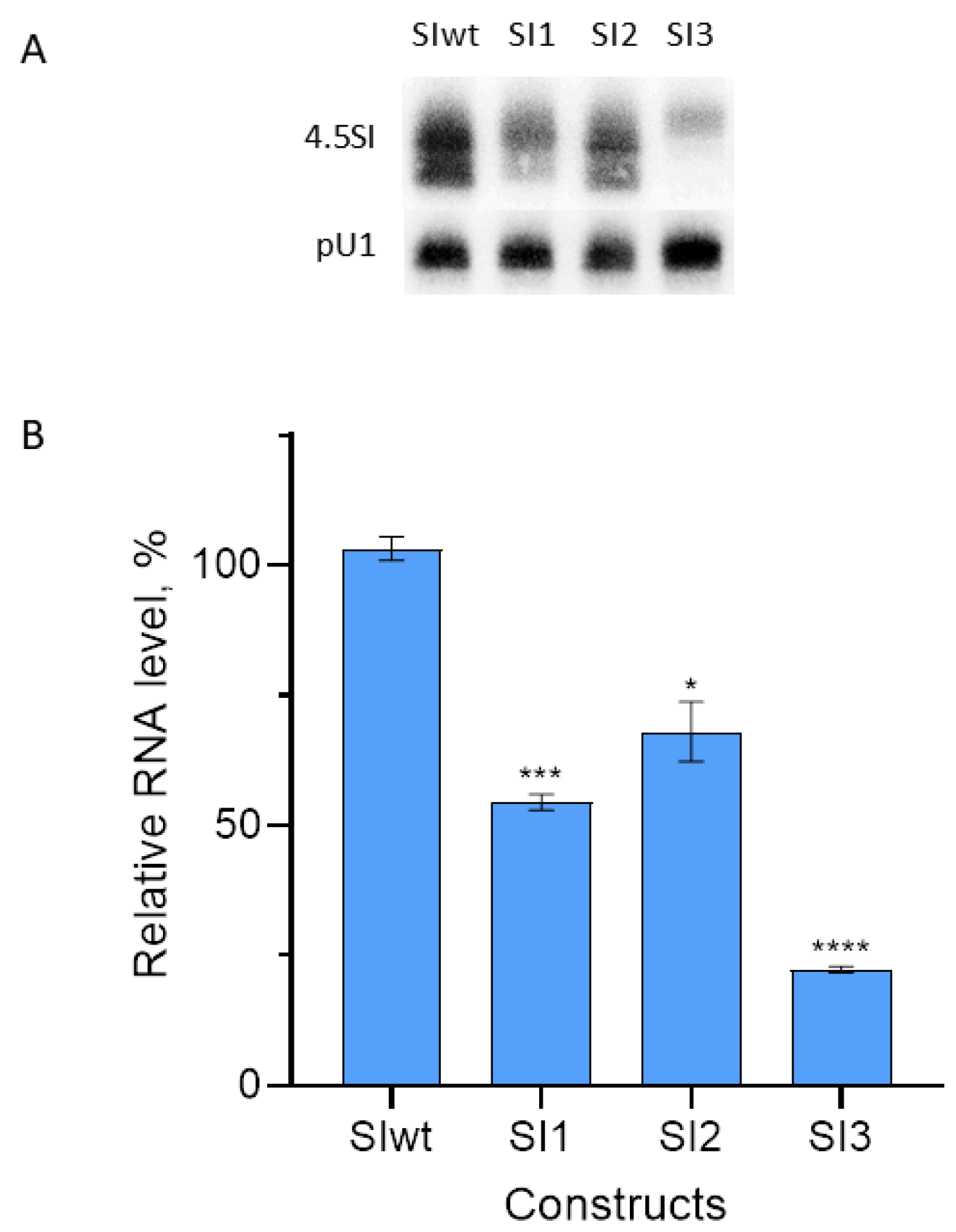
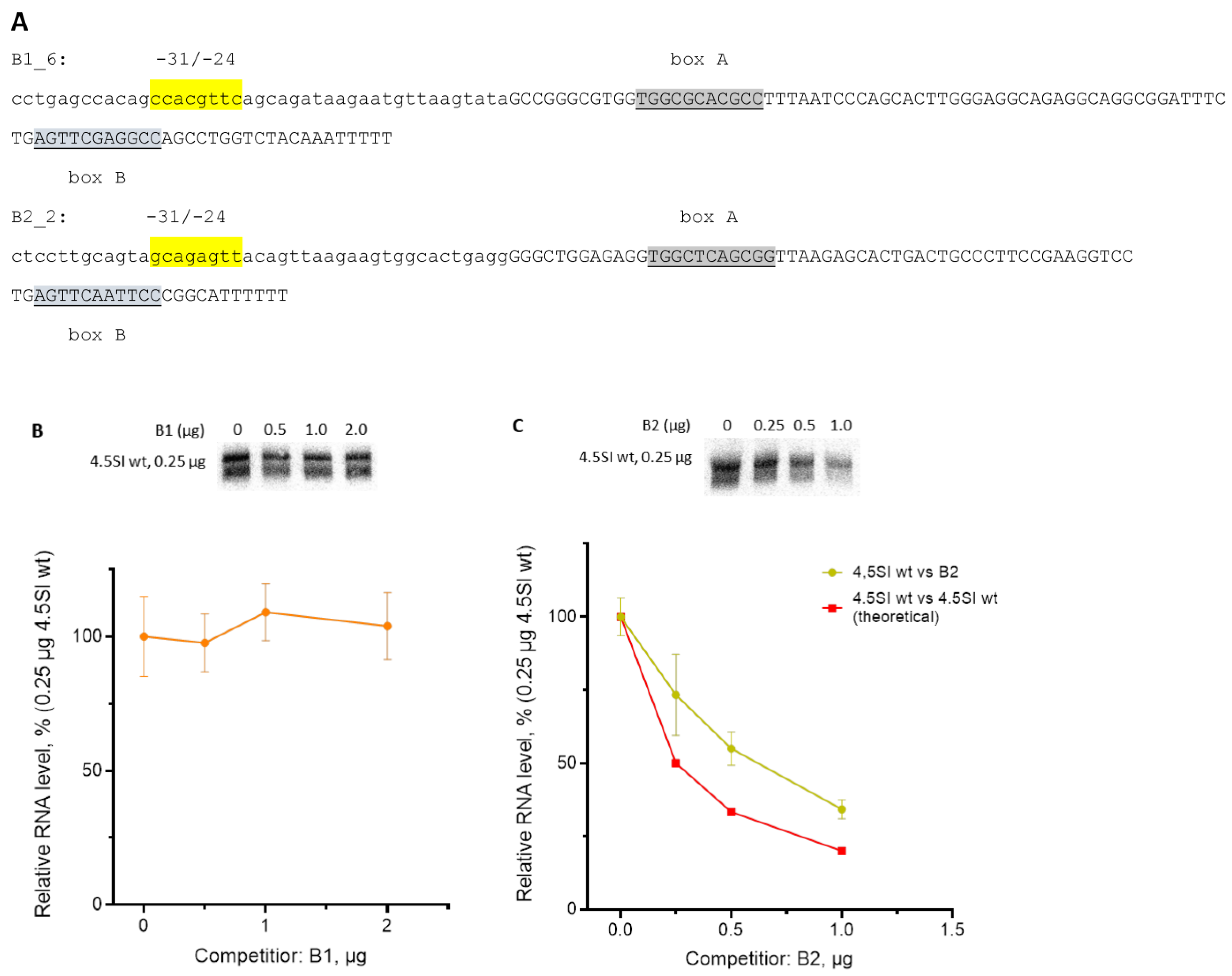
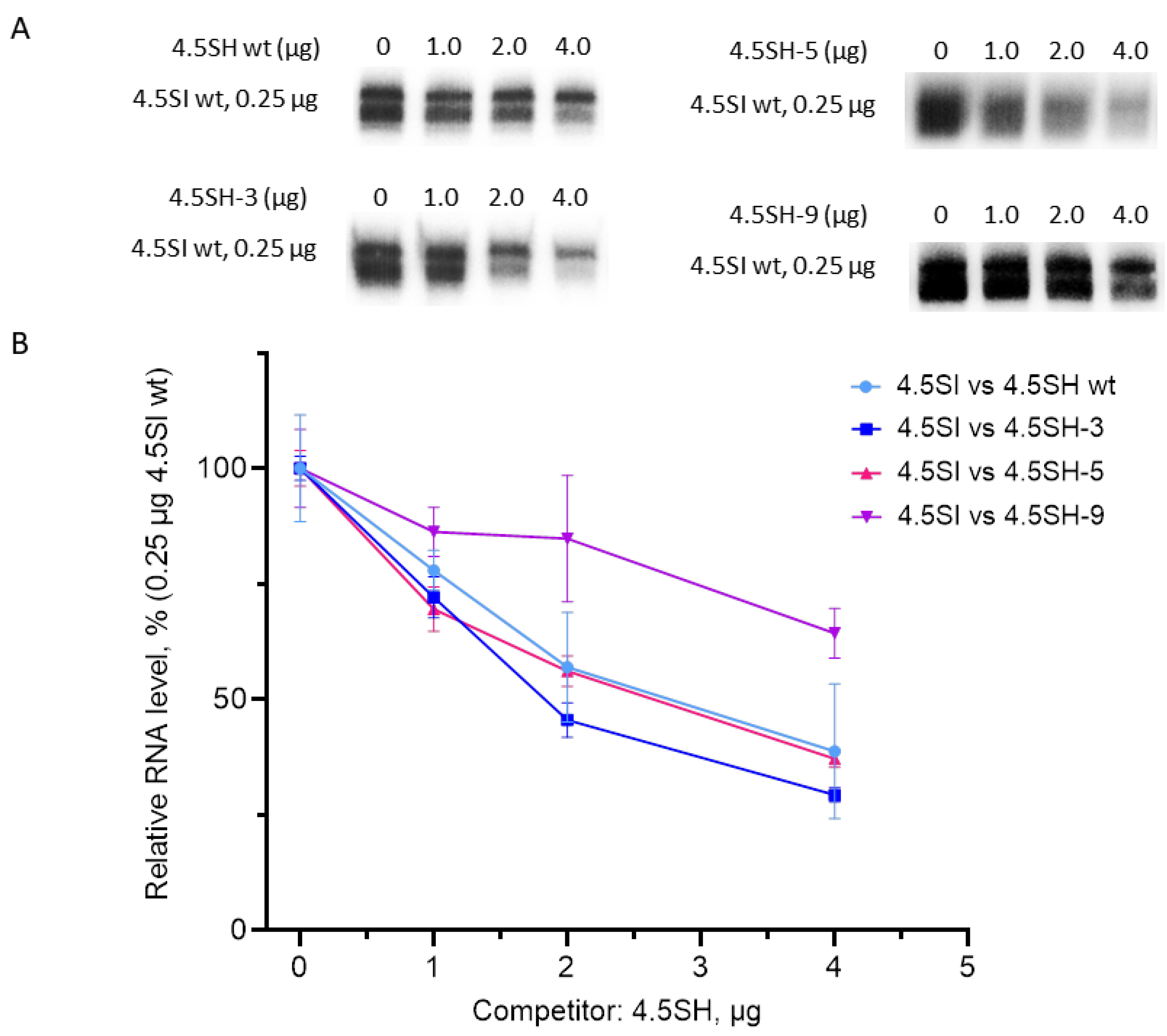
3.2. Effect of Replacement of Promoter Boxes on Pol III Transcription
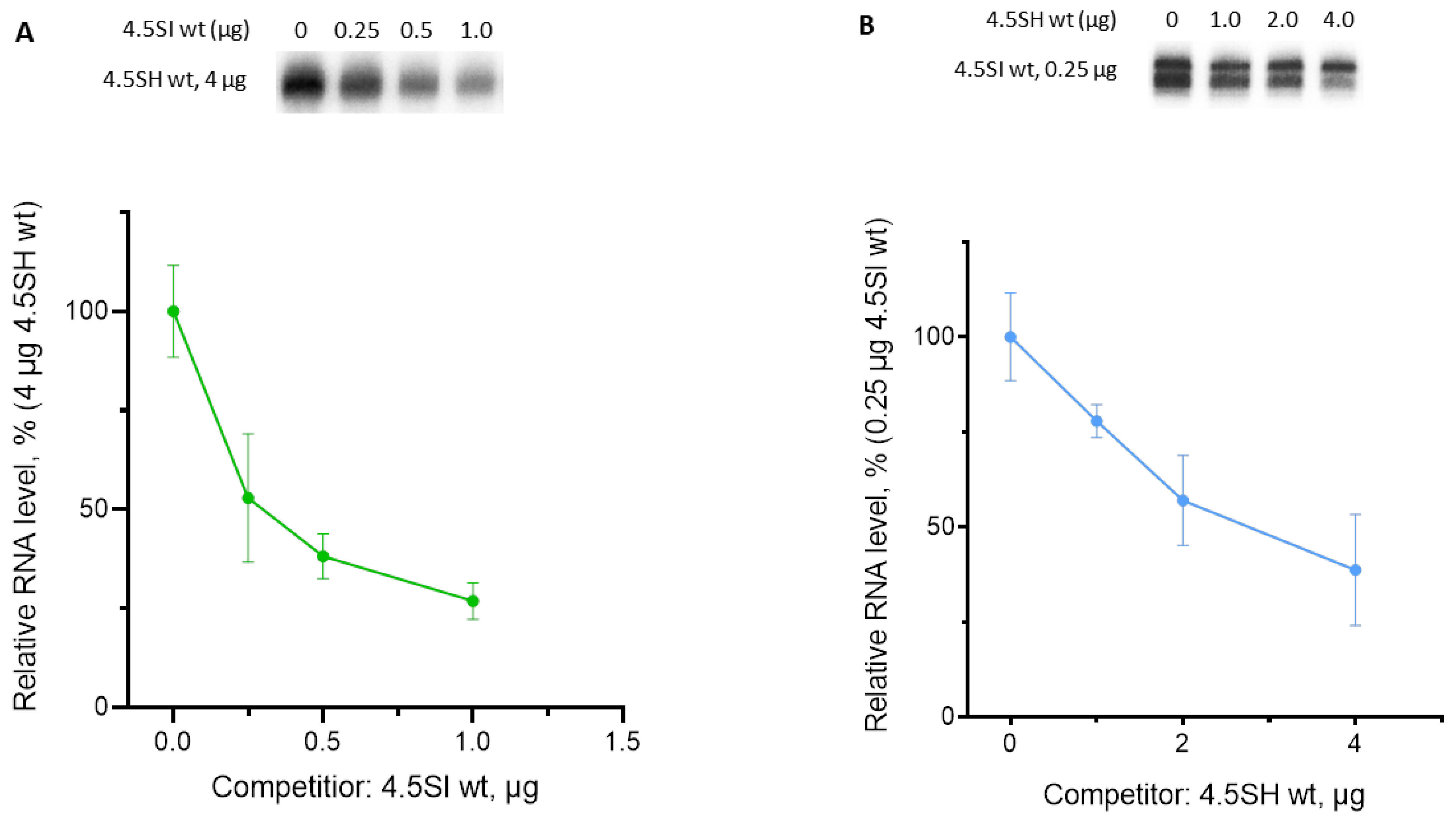
3.3. Effect of Box Replacement on Promoter Strength
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cech, T.R.; Steitz, J.A. The noncoding RNA revolution-trashing old rules to forge new ones. Cell 2014, 157, 77–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dieci, G.; Conti, A.; Pagano, A.; Carnevali, D. Identification of RNA polymerase III-transcribed genes in eukaryotic genomes. Biochim. Et Biophys. Acta (BBA)-Gene Regul. Mech. 2013, 1829, 296–305. [Google Scholar] [CrossRef] [PubMed]
- Dieci, G.; Fiorino, G.; Castelnuovo, M.; Teichmann, M.; Pagano, A. The expanding RNA polymerase III transcriptome. TRENDS Genet. 2007, 23, 614–622. [Google Scholar] [CrossRef] [PubMed]
- Horos, R.; Buscher, M.; Kleinendorst, R.; Alleaume, A.M.; Tarafder, A.K.; Schwarzl, T.; Dziuba, D.; Tischer, C.; Zielonka, E.M.; Adak, A.; et al. The Small Non-coding Vault RNA1-1 Acts as a Riboregulator of Autophagy. Cell 2019, 176, 1054–1067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kheir, E.; Krude, T. Non-coding Y RNAs associate with early replicating euchromatin in concordance with the origin recognition complex. J. Cell Sci. 2017, 130, 1239–1250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kramerov, D.A.; Vassetzky, N.S. SINEs. Wiley Interdiscip. Rev. RNA 2011, 2, 772–786. [Google Scholar] [CrossRef]
- Vassetzky, N.S.; Kramerov, D.A. SINEBase: A database and tool for SINE analysis. Nucleic Acids Res. 2013, 41, D83–D89. [Google Scholar] [CrossRef] [Green Version]
- Schramm, L.; Hernandez, N. Recruitment of RNA polymerase III to its target promoters. Genes Dev. 2002, 16, 2593–2620. [Google Scholar] [CrossRef] [Green Version]
- Oler, A.J.; Alla, R.K.; Roberts, D.N.; Wong, A.; Hollenhorst, P.C.; Chandler, K.J.; Cassiday, P.A.; Nelson, C.A.; Hagedorn, C.H.; Graves, B.J.; et al. Human RNA polymerase III transcriptomes and relationships to Pol II promoter chromatin and enhancer-binding factors. Nat. Struct. Mol. Biol. 2010, 17, 620–628. [Google Scholar] [CrossRef]
- Geiduschek, E.P.; Kassavetis, G.A. The RNA polymerase III transcription apparatus. J. Mol. Biol. 2001, 310, 1–26. [Google Scholar] [CrossRef]
- Hamada, M.; Huang, Y.; Lowe, T.M.; Maraia, R.J. Widespread use of TATA elements in the core promoters for RNA polymerases III, II, and I in fission yeast. Mol. Cell. Biol. 2001, 21, 6870–6881. [Google Scholar] [CrossRef] [Green Version]
- Giuliodori, S.; Percudani, R.; Braglia, P.; Ferrari, R.; Guffanti, E.; Ottonello, S.; Dieci, G. A composite upstream sequence motif potentiates tRNA gene transcription in yeast. J. Mol. Biol. 2003, 333, 1–20. [Google Scholar] [CrossRef]
- Zhang, G.; Lukoszek, R.; Mueller-Roeber, B.; Ignatova, Z. Different sequence signatures in the upstream regions of plant and animal tRNA genes shape distinct modes of regulation. Nucleic. Acids. Res. 2011, 39, 3331–3339. [Google Scholar] [CrossRef] [Green Version]
- Howe, J.G.; Shu, M.D. Upstream basal promoter element important for exclusive RNA polymerase III transcription of the EBER 2 gene. Mol. Cell. Biol. 1993, 13, 2655–2665. [Google Scholar] [CrossRef]
- Niller, H.H.; Salamon, D.; Ilg, K.; Koroknai, A.; Banati, F.; Bauml, G.; Rucker, O.; Schwarzmann, F.; Wolf, H.; Minarovits, J. The in vivo binding site for oncoprotein c-Myc in the promoter for Epstein-Barr virus (EBV) encoding RNA (EBER) 1 suggests a specific role for EBV in lymphomagenesis. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2003, 9, HY1–HY9. [Google Scholar]
- Fowlkes, D.M.; Shenk, T. Transcriptional control regions of the adenovirus VAI RNA gene. Cell 1980, 22, 405–413. [Google Scholar] [CrossRef]
- Piras, G.; Dittmer, J.; Radonovich, M.F.; Brady, J.N. Human T-cell leukemia virus type I Tax protein transactivates RNA polymerase III promoter in vitro and in vivo. J. Biol. Chem. 1996, 271, 20501–20506. [Google Scholar] [CrossRef] [Green Version]
- Englert, M.; Felis, M.; Junker, V.; Beier, H. Novel upstream and intragenic control elements for the RNA polymerase III-dependent transcription of human 7SL RNA genes. Biochimie 2004, 86, 867–874. [Google Scholar] [CrossRef]
- Ludwig, A.; Rozhdestvensky, T.S.; Kuryshev, V.Y.; Schmitz, J.; Brosius, J. An unusual primate locus that attracted two independent Alu insertions and facilitates their transcription. J. Mol. Biol. 2005, 350, 200–214. [Google Scholar] [CrossRef]
- Tatosyan, K.A.; Stasenko, D.V.; Koval, A.P.; Gogolevskaya, I.K.; Kramerov, D.A. TATA-Like Boxes in RNA Polymerase III Promoters: Requirements for Nucleotide Sequences. Int. J. Mol. Sci. 2020, 21, 3706. [Google Scholar] [CrossRef]
- White, R.J.; Jackson, S.P. Mechanism of TATA-binding protein recruitment to a TATA-less class III promoter. Cell 1992, 71, 1041–1053. [Google Scholar] [CrossRef]
- Joazeiro, C.A.; Kassavetis, G.A.; Geiduschek, E.P. Identical components of yeast transcription factor IIIB are required and sufficient for transcription of TATA box-containing and TATA-less genes. Mol. Cell. Biol. 1994, 14, 2798–2808. [Google Scholar] [CrossRef]
- Gogolevskaya, I.K.; Stasenko, D.V.; Tatosyan, K.A.; Kramerov, D.A. Influence of 5′-flanking sequence on 4.5SI RNA gene transcription by RNA polymerase III. Genome 2018, 61, 367–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gogolevskaya, I.K.; Koval, A.P.; Kramerov, D.A. Evolutionary history of 4.5SH RNA. Mol. Biol. Evol. 2005, 22, 1546–1554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gogolevskaya, I.K.; Kramerov, D.A. Evolutionary history of 4.5SI RNA and indication that it is functional. J. Mol. Evol. 2002, 54, 354–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harada, F.; Kato, N. Nucleotide sequences of 4.5S RNAs associated with poly(A)-containing RNAs of mouse and hamster cells. Nucleic Acids Res. 1980, 8, 1273–1285. [Google Scholar] [CrossRef] [Green Version]
- Ro-Choi, T.S.; Redy, R.; Henning, D.; Takano, T.; Taylor, C.W.; Busch, H. Nucleotide sequence of 4.5 S ribonucleic acid of Novikoff hepatoma cell nuclei. J. Biol. Chem. 1972, 247, 3205–3222. [Google Scholar] [CrossRef]
- Gogolevskaya, I.K.; Kramerov, D.A. 4.5SI RNA genes and the role of their 5′-flanking sequences in the gene transcription. Gene 2010, 451, 32–37. [Google Scholar] [CrossRef]
- Schoeniger, L.O.; Jelinek, W.R. 4.5S RNA is encoded by hundreds of tandemly linked genes, has a short half-life, and is hydrogen bonded in vivo to poly(A)-terminated RNAs in the cytoplasm of cultured mouse cells. Mol. Cell. Biol. 1986, 6, 1508–1519. [Google Scholar] [CrossRef]
- Krayev, A.S.; Markusheva, T.V.; Kramerov, D.A.; Ryskov, A.P.; Skryabin, K.G.; Bayev, A.A.; Georgiev, G.P. Ubiquitous transposon-like repeats B1 and B2 of the mouse genome: B2 sequencing. Nucleic Acids Res. 1982, 10, 7461–7475. [Google Scholar] [CrossRef] [Green Version]
- Serdobova, I.M.; Kramerov, D.A. Short retroposons of the B2 superfamily: Evolution and application for the study of rodent phylogeny. J. Mol. Evol. 1998, 46, 202–214. [Google Scholar] [CrossRef]
- Quentin, Y. Emergence of master sequences in families of retroposons derived from 7sl RNA. Genetica 1994, 93, 203–215. [Google Scholar] [CrossRef]
- Veniaminova, N.A.; Vassetzky, N.S.; Kramerov, D.A. B1 SINEs in different rodent families. Genomics 2007, 89, 678–686. [Google Scholar] [CrossRef] [Green Version]
- Ullu, E.; Tschudi, C. Alu sequences are processed 7SL RNA genes. Nature 1984, 312, 171–172. [Google Scholar] [CrossRef]
- Ishida, K.; Miyauchi, K.; Kimura, Y.; Mito, M.; Okada, S.; Suzuki, T.; Nakagawa, S. Regulation of gene expression via retrotransposon insertions and the noncoding RNA 4.5S RNAH. Genes Cells 2015, 20, 887–901. [Google Scholar] [CrossRef]
- Tatosyan, K.A.; Koval, A.P.; Gogolevskaya, I.K.; Kramerov, D.A. 4.5SI and 4.5SH RNAs: Expression in various rodent organs and abundance and distribution in the cell. Mol. Biol. 2017, 51, 142–149. [Google Scholar] [CrossRef]
- Koval, A.P.; Gogolevskaya, I.K.; Tatosyan, K.A.; Kramerov, D.A. Complementarity of end regions increases the lifetime of small RNAs in mammalian cells. PLoS ONE 2012, 7, e44157. [Google Scholar] [CrossRef] [Green Version]
- Tatosyan, K.A.; Kramerov, D.A. Heat shock increases lifetime of a small RNA and induces its accumulation in cells. Gene 2016, 587, 33–41. [Google Scholar] [CrossRef]
- Yoshimoto, R.; Nakayama, Y.; Yamamoto, I.; Tanaka, S.; Kurihara, M.; Suzuki, Y.; Kobayashi, T.; Kozuka-Hata, H.; Oyama, M.; Mito, M.; et al. 4.5SH RNA counteracts deleterious exonization of SINE B1 in mice. Res. Sq. 2022. [Google Scholar] [CrossRef]
- Koval, A.P.; Kramerov, D.A. 5′-flanking sequences can dramatically influence 4.5SH RNA gene transcription by RNA-polymerase III. Gene 2009, 446, 75–80. [Google Scholar] [CrossRef]
- Chomczynski, P.; Sacchi, N. The single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction: Twenty-something years on. Nat. Protoc. 2006, 1, 581–585. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, R.; Bernhart, S.H.; Honer Zu Siederdissen, C.; Tafer, H.; Flamm, C.; Stadler, P.F.; Hofacker, I.L. ViennaRNA Package 2.0. Algorithms Mol. Biol. 2011, 6, 26. [Google Scholar] [CrossRef] [PubMed]
- Vlahovicek, K.; Kajan, L.; Pongor, S. DNA analysis servers: Plot.it, bend.it, model.it and IS. Nucleic Acids Res. 2003, 31, 3686–3687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koval, A.P.; Gogolevskaya, I.K.; Tatosyan, K.A.; Kramerov, D.A. A 5′-3′ terminal stem in small non-coding RNAs extends their lifetime. Gene 2015, 555, 464–468. [Google Scholar] [CrossRef]
- Kriegs, J.O.; Churakov, G.; Jurka, J.; Brosius, J.; Schmitz, J. Evolutionary history of 7SL RNA-derived SINEs in Supraprimates. Trends Genet. 2007, 23, 158–161. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stasenko, D.V.; Tatosyan, K.A.; Borodulina, O.R.; Kramerov, D.A. Nucleotide Context Can Modulate Promoter Strength in Genes Transcribed by RNA Polymerase III. Genes 2023, 14, 802. https://doi.org/10.3390/genes14040802
Stasenko DV, Tatosyan KA, Borodulina OR, Kramerov DA. Nucleotide Context Can Modulate Promoter Strength in Genes Transcribed by RNA Polymerase III. Genes. 2023; 14(4):802. https://doi.org/10.3390/genes14040802
Chicago/Turabian StyleStasenko, Danil V., Karina A. Tatosyan, Olga R. Borodulina, and Dmitri A. Kramerov. 2023. "Nucleotide Context Can Modulate Promoter Strength in Genes Transcribed by RNA Polymerase III" Genes 14, no. 4: 802. https://doi.org/10.3390/genes14040802