
Genetic Characterization and Insular Habitat Enveloping of Endangered Leaf-Nosed Bat, Hipposideros nicobarulae (Mammalia: Chiroptera) in India: Phylogenetic Inference and Conservation Implication

 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
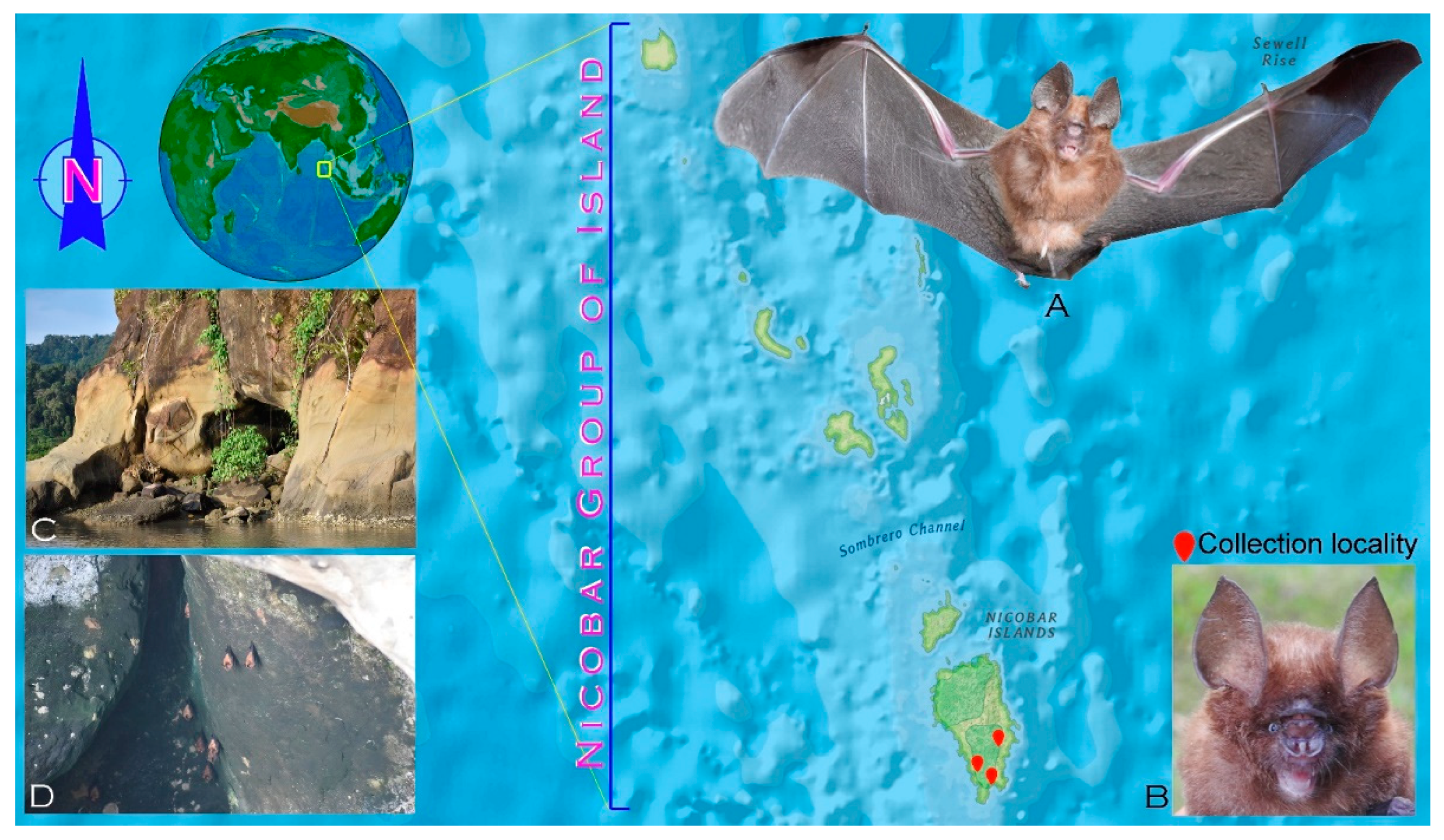
2.1. Ethics Statement and Sampling
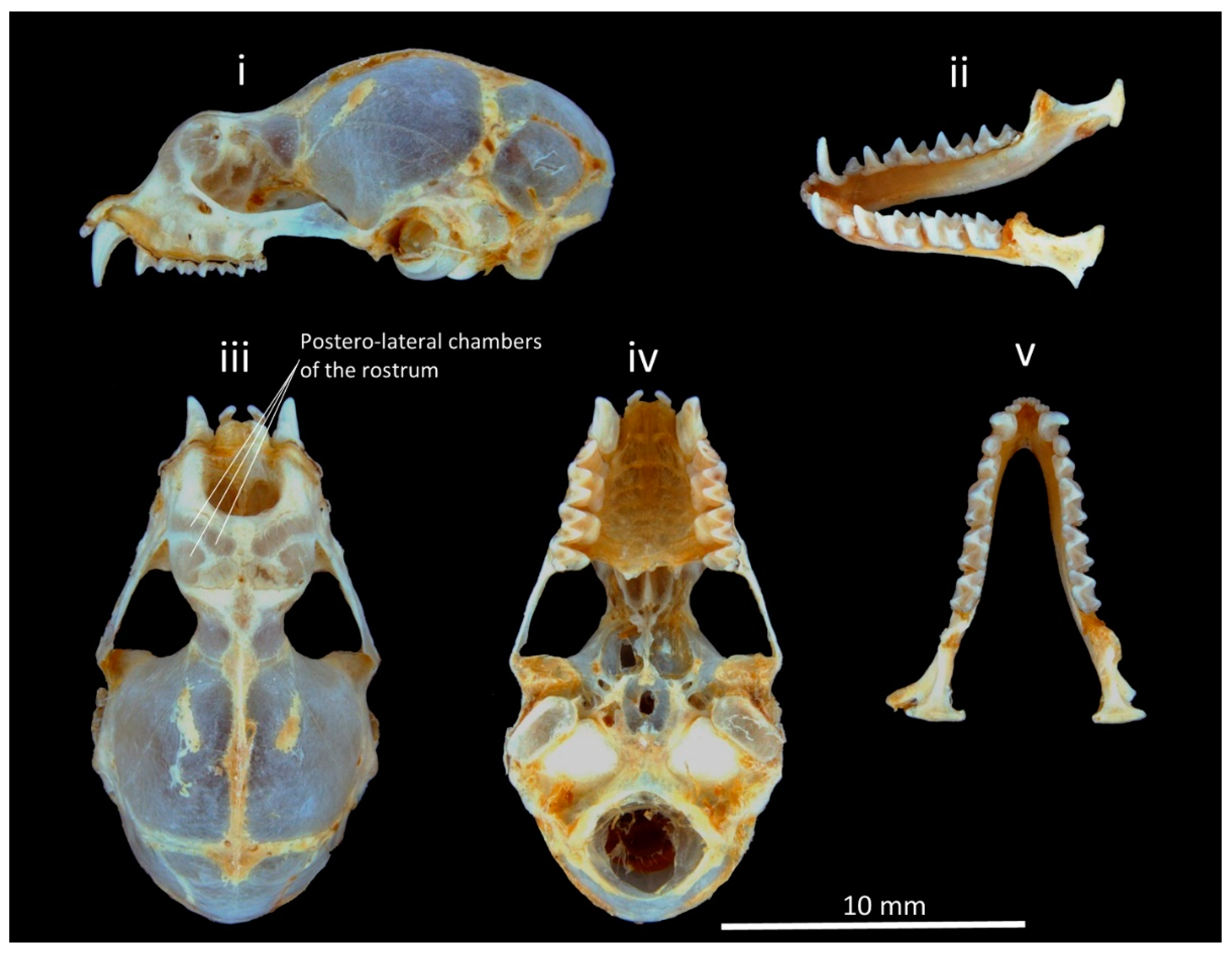
2.2. Morphological Identification
2.3. Mitochondrial DNA Extraction and Sequencing
2.4. Sequence Annotation and Dataset Construction
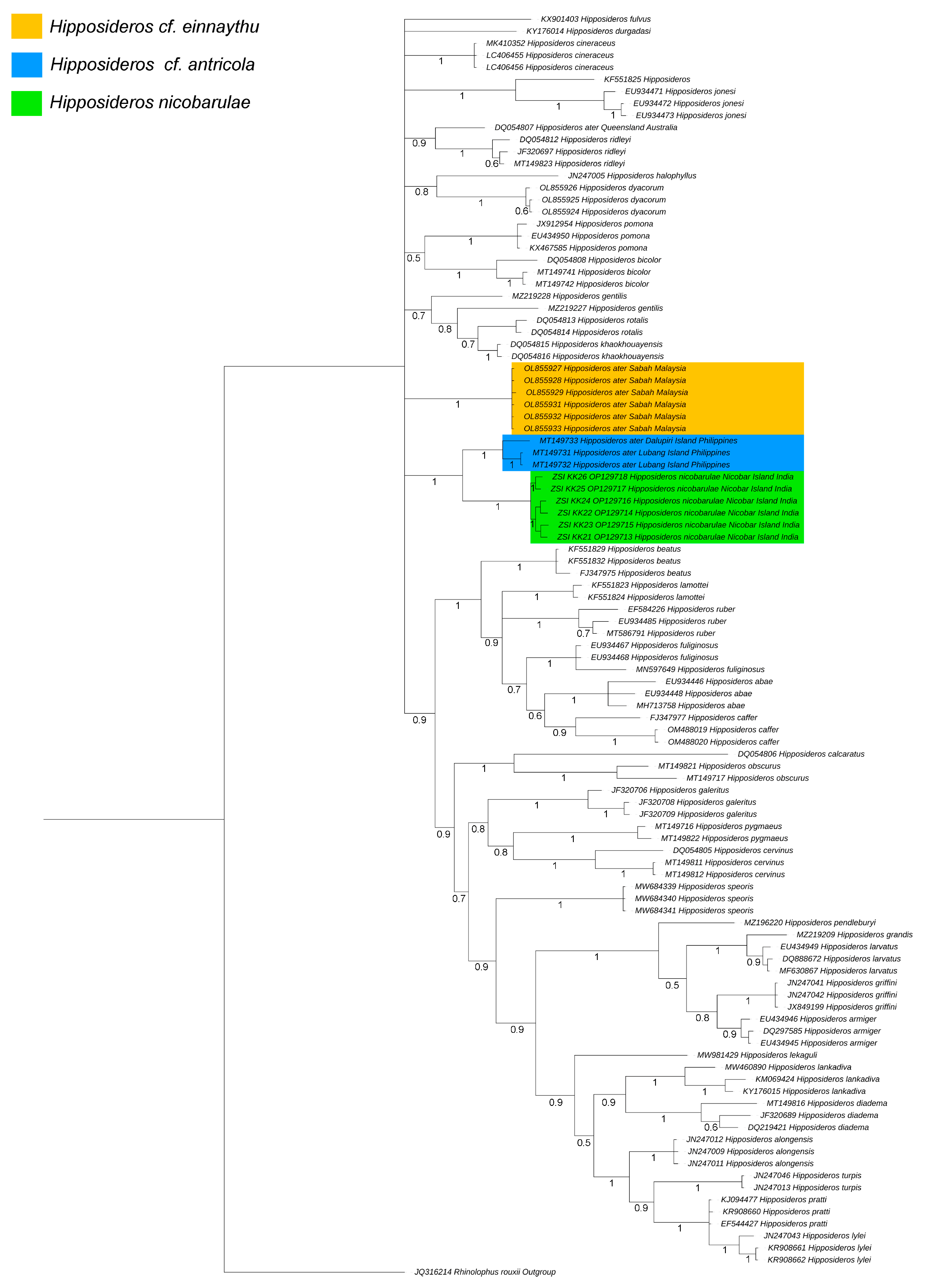
2.5. Genetic Divergence and Phylogenetic Analyses
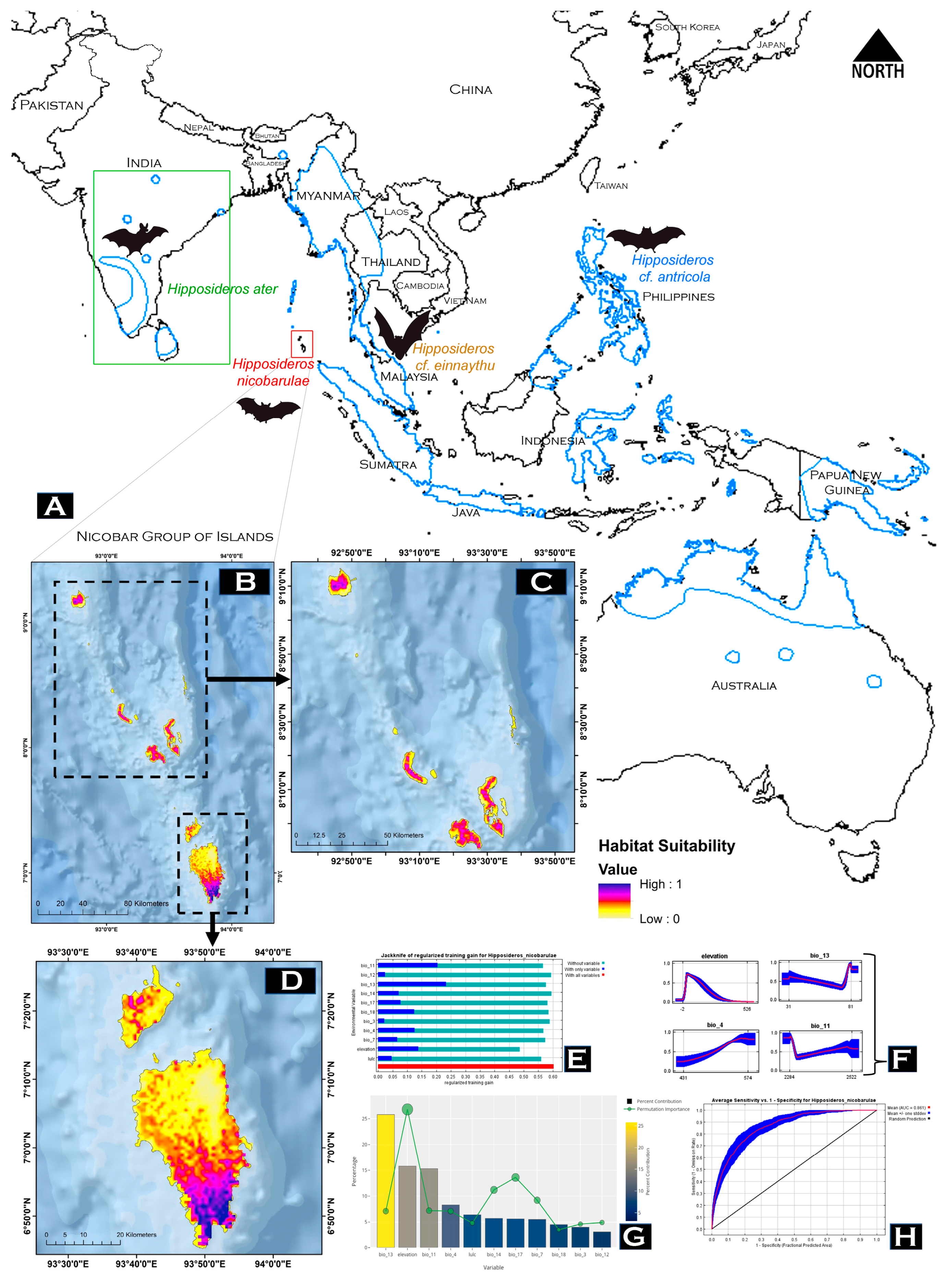
2.6. Species Occurrence Data
2.7. Model Building and Evaluation
3. Results
3.1. Taxonomic Identification
3.2. Molecular Identification
3.3. Model Performance and Habitat Suitability
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Simmons, N.B.; Cirranello, A.L. Bat Species of the World: A Taxonomic and Geographic Database. Available online: https://batnames.org (accessed on 5 February 2023).
- Burgin, C.J.; Colella, J.P.; Kahn, P.L.; Upham, N.S. How many species of mammals are there? J. Mammal. 2018, 99, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Douangboubpha, B.; Bumrungsri, S.; Satasook, C.; Soisook, P.; HlaBu, S.S.; Aul, B.; Harrison, D.L.; Pearch, M.J.; Thomas, N.M.; Bates, P.J.J. A New Species of Small Hipposideros (Chiroptera: Hipposideridae) from Myanmar and a Revaluation of the Taxon H. nicobarulae Miller, 1902 from the Nicobar Islands. Acta Chiropterologica 2011, 13, 61–78. [Google Scholar] [CrossRef]
- Aul, B.; Bates, P.J.J.; Harrison, D.L.; Marimuthu, G. Diversity, distribution and status of bats on the Andaman and Nicobar Islands, India. Oryx 2014, 48, 204–214. [Google Scholar] [CrossRef] [Green Version]
- Srinivasulu, B.; Srinivasulu, A.; Srinivasulu, C.; Dar, T.H.; Gopi, A.; Jones, G. First record of the diadem leaf-nosed bat Hipposideros diadema (E. Geoffroy, 1813) (Chiroptera, Hipposideridae) from the Andaman Islands, India with the possible occurrence of a hitherto unreported subspecies. J. Threat. Taxa 2016, 8, 9316–9321. [Google Scholar] [CrossRef]
- Srinivasulu, C.; Srinivasulu, A.; Srinivasulu, B.; Gopi, A.; Dar, T.H.; Bates, P.J.J.; Rossiter, S.J.; Jones, G. Recent surveys of bats from the Andaman Islands, India: Diversity, distribution, and echolocation characteristics. Acta Chiropterologica 2017, 19, 419–437. [Google Scholar] [CrossRef]
- Srinivasulu, B.; Srinivasulu, C. In plain sight: Bacular and noseleaf morphology supports distinct specific status of Roundleaf Bats Hipposideros pomona Andersen, 1918 and Hipposideros gentilis Andersen, 1918 (Chiroptera: Hipposideridae). J. Threat. Taxa 2018, 10, 12018–12026. [Google Scholar] [CrossRef]
- Miller, G.S. The mammals of the Andaman and Nicobar Islands. Proc. U. S. Natl. Mus. 1902, 24, 751–795. [Google Scholar] [CrossRef] [Green Version]
- Hill, J.E. A revision of the genus Hipposideros. Bull. Br. Mus. Nat. Hist. Zool. 1963, 2, 1–129. [Google Scholar] [CrossRef]
- Simmons, N.B. Order Chiroptera. In Mammal Species of the World: A Taxonomic and Geographic Reference; Wilson, D.E., Reeder, D.M., Eds.; The Johns Hopkins University Press: Baltimore, MD, USA, 2005; pp. 312–529. [Google Scholar]
- Srinivasulu, B.; Srinivasulu, C.; Waldien, D.L. Hipposideros nicobarulae. In The IUCN Red List of Threatened Species 2020: E.T80458824A95642215; Cambridge, UK, 2020; Available online: https://doi.org/10.2305/IUCN.UK.2020-3.RLTS.T80458824A95642215.en (accessed on 10 March 2023).
- Teeling, E.C.; Springer, M.S.; Madsen, O.; Bates, P.; O’brien, S.J.; Murphy, W.J. A molecular phylogeny of bats illuminates biogeography and fossil record. Science 2005, 307, 580–584. [Google Scholar] [CrossRef]
- Esselstyn, J.A.; Evans, B.J.; Sedlock, J.L.; Anwarali Khan, F.A.; Heaney, L.R. Single-locus species delimitation: A test of the mixed Yule-coalescent model, with an empirical application to Philippine round-leaf bats. Proc. Biol. Sci. 2012, 279, 3678–3686. [Google Scholar] [CrossRef]
- Hawkins, J.A.; Kaczmarek, M.E.; Müller, M.A.; Drosten, C.; Press, W.H.; Sawyer, S.L. A metaanalysis of bat phylogenetics and positive selection based on genomes and transcriptomes from 18 species. Proc. Natl. Acad. Sci. USA 2019, 116, 11351–11360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jebb, D.; Huang, Z.; Pippel, M.; Hughes, G.M.; Lavrichenko, K.; Devanna, P.; Winkler, S.; Jermiin, L.S.; Skirmuntt, E.C.; Katzourakis, A.; et al. Six reference-quality genomes reveal evolution of bat adaptations. Nature 2020, 583, 578–584. [Google Scholar] [CrossRef]
- Thong, V.D.; Puechmaille, S.J.; Denzinger, A.; Dietz, C.; Csorba, G.; Bates, P.J.J.; Teeling, E.C.; Schnitzler, H.-U. A new species of Hipposideros (Chiroptera: Hipposideridae) from Vietnam. J. Mammal. 2012, 93, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Murray, S.W.; Campbell, P.; Kingston, T.; Zubaid, A.; Francis, C.M.; Kunz, T.H. Molecular phylogeny of hipposiderid bats from Southeast Asia and evidence of cryptic diversity. Mol. Phylogenet. Evol. 2012, 62, 597–611. [Google Scholar] [CrossRef]
- Rakotoarivelo, A.R.; Willows-Munro, S.; Schoeman, M.C.; Lamb, J.M.; Goodman, S.M. Cryptic diversity in Hipposideros commersoni sensu stricto (Chiroptera: Hipposideridae) in the western portion of Madagascar. BMC Evol. Biol. 2015, 15, 235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavery, T.H.; Leung, L.K.P.; Seddon, J.M. Molecular phylogeny of hipposiderid bats (Chiroptera: Hipposideridae) from Solomon Islands and Cape York Peninsula, Australia. Zool. Scr. 2013, 43, 429–442. [Google Scholar]
- Foley, N.; Thong, V.D.; Soisook, P.; Goodman, S.; Armstrong, K.; Jacobs, D.; Puechmaille, S.J.; Teeling, E. How and why overcome the impediments to resolution: Lessons from rhinolophid and hipposiderid bats. Mol. Biol. Evol. 2014, 33, 313–333. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; He, C.; Shen, C.; Jiang, T.; Shi, L.; Sun, K.; Berquist, S.W.; Feng, J. Phylogeography and population genetic structure of the great leaf-nosed bat (Hipposideros armiger) in China. J. Hered. 2010, 101, 562–572. [Google Scholar] [CrossRef] [Green Version]
- Lin, A.Q.; Csorba, G.; Li, L.F.; Jiang, T.L.; Lu, G.J.; Thong, V.D.; Soisook, P.; Sun, K.P.; Feng, J. Phylogeography of Hipposideros armiger (Chiroptera: Hipposideridae) in the Oriental Region: The contribution of multiple Pleistocene glacial refugia and intrinsic factors to contemporary population genetic structure. J. Biogeogr. 2013, 41, 317–327. [Google Scholar] [CrossRef]
- Puechmaille, S.J.; Gouilh, M.A.; Piyapan, P.; Yokubol, M.; Mie, K.M.; Bates, P.J.; Satasook, C.; New, T.; Bu, S.S.; Mackie, I.J.; et al. The evolution of sensory divergence in the context of limited gene flow in the bumblebee bat. Nat. Commun. 2011, 2, 573. [Google Scholar] [CrossRef] [Green Version]
- Teeling, E.C.; Scally, M.; Kao, D.J.; Romagnoli, M.L.; Springer, M.S.; Stanhope, M.J. Molecular evidence regarding the origin of echolocation and flight in bats. Nature 2000, 403, 188–192. [Google Scholar] [CrossRef]
- Johns, G.C.; Avise, J.C. A comparative summary of genetic distances in the vertebrates from the mitochondrial cytochrome b gene. Mol. Biol. Evol. 1998, 15, 1481–1490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradley, R.D.; Baker, R.J. A Test of the Genetic Species Concept: Cytochrome-b Sequences and Mammals. J. Mammal. 2001, 82, 960–973. [Google Scholar] [CrossRef]
- Tobe, S.S.; Kitchener, A.C.; Linacre, A.M.T. Reconstructing Mammalian Phylogenies: A Detailed Comparison of the Cytochrome b and Cytochrome Oxidase Subunit I Mitochondrial Genes. PLoS ONE 2010, 5, e14156. [Google Scholar] [CrossRef] [Green Version]
- Walther, G.R.; Post, E.; Convey, P.; Menzel, A.; Parmesan, C.; Beebee, T.J.C.; Fromentin, J.M.; Hoegh-Guldberg, O.; Bairlein, F. Ecological responses to recent climate change. Nature 2002, 416, 389–395. [Google Scholar] [CrossRef]
- Hughes, A.C.; Satasook, C.; Bates, P.J.; Bumrungsri, S.; Jones, G. The projected effects of climatic and vegetation changes on the distribution and diversity of Southeast Asian bats. Glob. Chang. Biol. 2012, 18, 1854–1865. [Google Scholar] [CrossRef]
- Wordley, C.F.R.; Sankaran, M.; Mudappa, D.; Altringham, J.D. Landscape scale habitat suitability modelling of bats in the Western Ghats of India: Bats like something in their tea. Biol. Conserv. 2015, 191, 529–536. [Google Scholar] [CrossRef] [Green Version]
- Herkt, K.M.B.; Barnikel, G.; Skidmore, A.K.; Fahr, J. A high-resolution model of bat diversity and endemism for continental Africa. Ecol. Model. 2016, 320, 9–28. [Google Scholar] [CrossRef]
- Bandara, A.P.M.J.; Madurapperuma, B.D.; Edirisinghe, G.; Gabadage, D.; Botejue, M.; Surasinghe, T.D. Bioclimatic Envelopes for Two Bat Species from a Tropical Island: Insights on Current and Future Distribution from Ecological Niche Modeling. Diversity 2022, 14, 506. [Google Scholar] [CrossRef]
- Raman, S.; Shameer, T.T.; Pooja, U.; Hughes, A.C. Identifying priority areas for bat conservation in the Western Ghats Mountain range, peninsular India. J. Mammal. 2022, gyac060. [Google Scholar] [CrossRef]
- Bates, P.J.J.; Harrison, D.L. Bats of Indian Sub-Continent; Harrison Zoological Museum: Sevenoaks, UK, 1997; p. xv-258. [Google Scholar]
- Sinha, Y.P. Contribution to the Knowledge of Bats of North East Hills India; Occasional Paper No. 174; Zoological Survey of India: Kolakata, India, 1999; pp. 1–52. [Google Scholar]
- Thong, V.D.; Denzinger, A.; Sang, N.V.; Huyen, N.T.T.; Thanh, H.T.; Loi, D.N.; Nha, P.V.; Viet, N.V.; Tien, P.D.; Tuanmu, M.-N.; et al. Bat Diversity in Cat Ba Biosphere Reserve, Northeastern Vietnam: A Review with New Records from Mangrove Ecosystem. Diversity 2021, 13, 376. [Google Scholar] [CrossRef]
- Verma, S.K.; Singh, L. Novel universal primers establish identity of an enormous number of animal species for forensic application. Mol. Ecol. Notes 2002, 3, 28–31. [Google Scholar] [CrossRef]
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001; Volume 1. [Google Scholar]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. JModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Trifinopoulos, J.; Nguyen, L.-T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Bork, P. Interactive tree of life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [Green Version]
- GBIF. Free and Open Access to Biodiversity Data. Available online: https://doi.org/10.15468/dl.d8ph4a (accessed on 5 February 2023).
- Fick, S.E.; Hijmans, R.J. Worldclim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudık, M. Modeling of species distributions with MaxEnt: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Halvorsen, R.; Mazzoni, S.; Dirksen, J.W.; Næsset, E.; Gobakken, T.; Ohlson, M. How important are choice of model selection method and spatial autocorrelation of presence data for distribution modelling by MaxEnt? Ecol. Model. 2016, 328, 108–118. [Google Scholar] [CrossRef]
- Kamalakannan, M.; Sivaperuman, C.; Kundu, S.; Gokulakrishnan, G.; Venkatraman, C.; Chandra, K. Discovery of a new mammal species (Soricidae: Eulipotyphla) from Narcondam volcanic island, India. Sci. Rep. 2021, 11, 9416. [Google Scholar] [CrossRef] [PubMed]
- Voris, H.K. Maps of Pleistocene Sea levels in Southeast Asia: Shorelines, river systems and time durations. J. Biogeogr. 2000, 27, 1153–1167. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, S.K.; Nagendran, G.; Karthikeyan, M.; Vasu, P.; Tripathi, S.K.; Varghese, S.; Raghav, S.; Resmi, S. Morphology of submarine volcanic seamounts from inner volcanic arc of Andaman Sea. Indian J. Geosci. 2017, 71, 451–470. [Google Scholar]
- Robinson, M.F.; Jenkins, P.D.; Francis, C.M.; Fulford, A.J.C. A new species of the Hipposideros pratti group (Chiroptera, Hipposideridae) from Lao PDR and Vietnam. Acta Chiropterologica 2003, 5, 31–48. [Google Scholar] [CrossRef]
- Bates, P.J.J.; Rossiter, S.J.; Suyanto, A.; Kingston, T. A new species of Hipposideros (Chiroptera: Hipposideridae) from Sulawesi. Acta Chiropterologica 2007, 9, 13–26. [Google Scholar] [CrossRef]
- Baker, R.J.; Bradley, R.D. Speciation in Mammals and the Genetic Species Concept. J. Mammal. 2006, 87, 643–662. [Google Scholar] [CrossRef]
- Clare, E.L.; Lim, B.K.; Fenton, M.B.; Hebert, P.D.N. Neotropical Bats: Estimating Species Diversity with DNA Barcodes. PLoS ONE 2011, 6, e22648. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Li, Y.; Motokawa, M.; Wu, Y.; Harada, M.; Li, Y. The Effectiveness of Molecular, Karyotype and Morphological Methods in the Identification of Morphologically Conservative Sibling Species: An Integrative Taxonomic Case of the Crocidura attenuata Species Complex in Mainland China. Animals 2023, 13, 643. [Google Scholar] [CrossRef] [PubMed]
- Teeling, E.C.; Madsen, O.; Van den Bussche, R.A.; de Jong, W.W.; Stanhope, M.J.; Springer, M.S. Microbat paraphyly and the convergent evolution of a key innovation in Old World rhinolophoid microbats. Proc. Natl. Acad. Sci. USA 2002, 99, 1432–1436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guillén-Servent, A.; Francis, C.M. A new species of bat of the Hipposideros bicolor group (Chiroptera: Hipposideridae) from Central Laos, with evidence of convergent evolution with Sundaic taxa. Acta Chiropterologica 2006, 8, 39–61. [Google Scholar] [CrossRef]
- Malik, J.N.; Johnson, F.C.; Khan, A.; Sahoo, S.; Irshad, R.; Paul, D.; Arora, S.; Baghel, P.K.; Chopra, S. Tsunami records of the last 8000 years in the Andaman Island, India, from mega and large earthquakes: Insights on recurrence interval. Sci Rep. 2019, 9, 18463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mageswaran, T.; Sachithanandam, V.; Sridhar, R.; Mahapatra, M.; Purvaja, R.; Ramesh, R. Impact of sea level rise and shoreline changes in the tropical island ecosystem of Andaman and Nicobar region, India. Nat. Hazards 2021, 109, 1717–1741. [Google Scholar] [CrossRef]
- Brooks, T.M.; Mittermeier, R.A.; da Fonseca, G.A.; Gerlach, J.; Hoffmann, M.; Lamoreux, J.F.; Mittermeier, C.G.; Pilgrim, J.D.; Rodrigues, A.S. Global biodiversity conservation priorities. Science 2006, 313, 58–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allan, J.R.; Possingham, H.P.; Atkinson, S.C.; Waldron, A.; Di Marco, M.; Butchart, S.H.M.; Adams, V.M.; Kissling, W.D.; Worsdell, T.; Sandbrook, C.; et al. The minimum land area requiring conservation attention to safeguard biodiversity. Science 2022, 376, 1094–1101. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | HBL | FA | EL | HF | Tib | T |
|---|---|---|---|---|---|---|
| H. nicobarulae (n = 6) | 41.6–47.9 | 38.1–42.2 | 15.5–15.9 | 7.67–8.05 | 17.2–18.4 | 24.7–32.5 |
| H. ater (n = 11) | 38.0–48.0 | 34.0–38.0 | 14.8–20.0 | 5.30–7.20 | 15.2–17.8 | 20.0–30.0 |
| H. einnaythu (n = 2) | 43.3, 49.1 | 39.5, 40.3 | 16.6, 16.7 | 6.5, 7.0 | 16.2, 17.8 | 24.7, 28.7 |
| H. diadema (n = 6) | - | 80.2–90.2 | 23.3–30.8 | 13.67–18.1 | 29.7–37.8 | 44.3–57.2 |
| H. fulvus (n = 35) | 40.0–50.0 | 38.4–44.0 | 19.0–26.0 | 6.0–9.8 | 16.5–20.7 | 24.0–35.0 |
| H. grandis (n = 12) | 70.0–79.0 | 60.0–64.0 | 21.5–24.0 | 12.0–13.5 | 21.0–25.7 | 34.0–39.0 |
| H. larvatus (n = 8) | - | 48.7–59.0 | 14.4–23.5 | 3.9–6.6 | 19.0–20.1 | 24.8–35.5 |
| H. gentilis (n = 9) | 36.0–52.0 | 39.5–43.2 | 22.0–25.0 | 6.3–8.5 | 18.2–19.1 | 28.0–35.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kundu, S.; Kamalakannan, M.; Mukherjee, T.; Banerjee, D.; Kim, H.-W. Genetic Characterization and Insular Habitat Enveloping of Endangered Leaf-Nosed Bat, Hipposideros nicobarulae (Mammalia: Chiroptera) in India: Phylogenetic Inference and Conservation Implication. Genes 2023, 14, 765. https://doi.org/10.3390/genes14030765
Kundu S, Kamalakannan M, Mukherjee T, Banerjee D, Kim H-W. Genetic Characterization and Insular Habitat Enveloping of Endangered Leaf-Nosed Bat, Hipposideros nicobarulae (Mammalia: Chiroptera) in India: Phylogenetic Inference and Conservation Implication. Genes. 2023; 14(3):765. https://doi.org/10.3390/genes14030765
Chicago/Turabian StyleKundu, Shantanu, Manokaran Kamalakannan, Tanoy Mukherjee, Dhriti Banerjee, and Hyun-Woo Kim. 2023. "Genetic Characterization and Insular Habitat Enveloping of Endangered Leaf-Nosed Bat, Hipposideros nicobarulae (Mammalia: Chiroptera) in India: Phylogenetic Inference and Conservation Implication" Genes 14, no. 3: 765. https://doi.org/10.3390/genes14030765