
A Population Genetic Perspective on Subsistence Systems in the Sahel/Savannah Belt of Africa and the Historical Role of Pastoralism


{kind=link}
{kind=link}
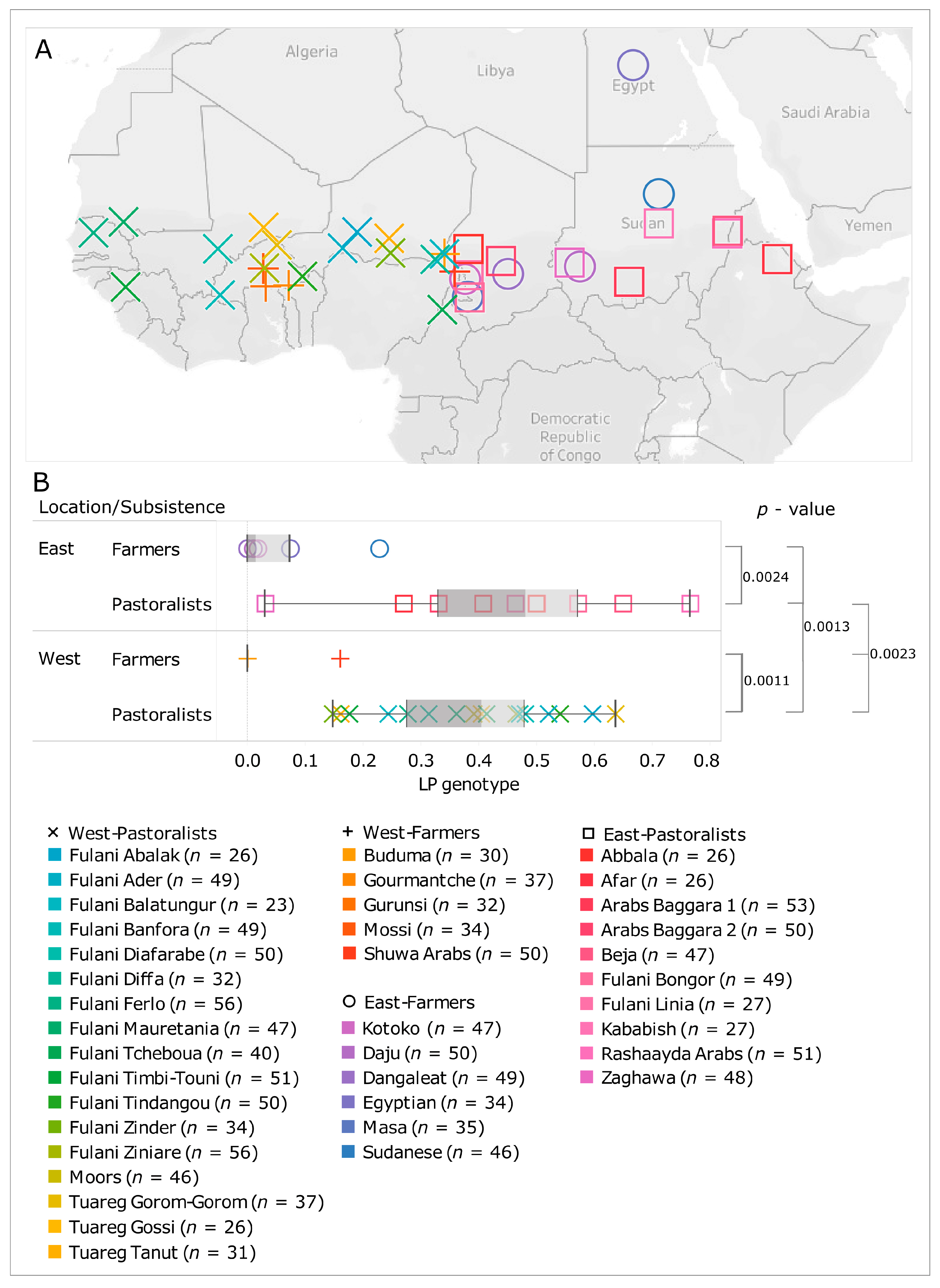{kind=link}
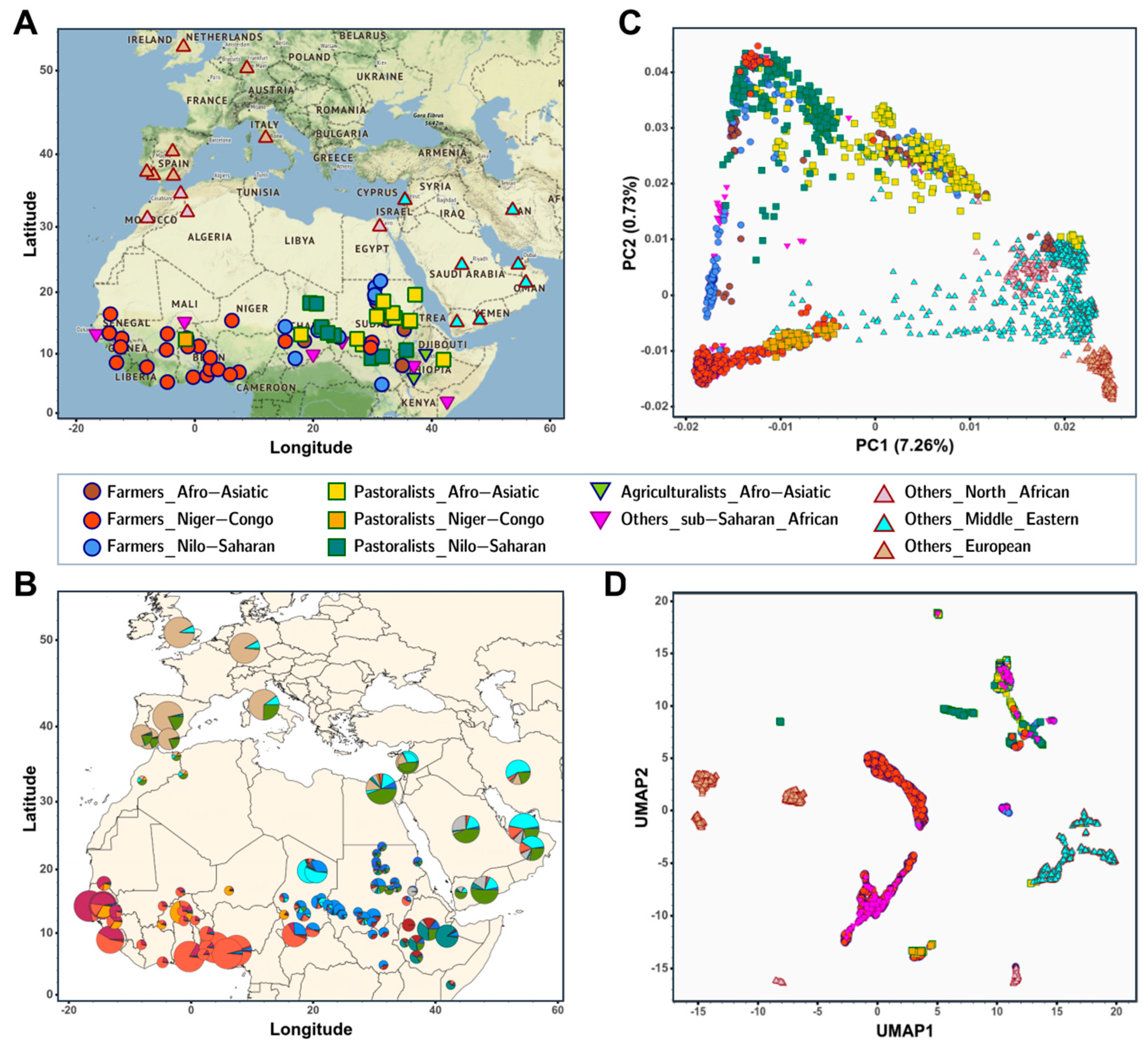{kind=link}
Abstract
:1. Introduction
2. Archaeological Imprints of Pastoralism and Pastoralist’s Continuity
3. Contributions of Population Genetics to the Understanding of Human Health
4. The Contribution of Population Genetics to Disentangling Human History
5. The Sociocultural Dynamics of Sahelian Populations
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Raynaut, C.; Delville, P.L. Un espace partagé: Complémentarité et concurrence des usages. In Sahels Diversité et Dynamiques des Relations Sociétés-Nature; Raynaut, C., Ed.; Karthala: Paris, France, 1997; pp. 143–174. [Google Scholar]
- Pedersen, J.; Benjaminsen, T.A. One Leg or Two? Food Security and Pastoralism in the Northern Sahel. Hum. Ecol. 2008, 36, 43–57. [Google Scholar] [CrossRef]
- Hardin, G. The tragedy of the commons. The population problem has no technical solution; it requires a fundamental extension in morality. Science 1968, 162, 1243–1248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behnke, R. Earth system sciences. In The End of Desertification? Disputing Environmental Change in the Drylands; Mortimore, M., Ed.; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Dupire, M. Peuls Nomads. In Étude Descriptive des Wodaabe du Sahel Nigérien; Karthala: Paris, France, 1962. [Google Scholar]
- Stenning, D.J. Savannah Nomads; A Study of the Wodaabe Pastoral Fulani of Western Bornu Province, Northern Region, Nigeria; International African Institute by Oxford University Press: London, UK, 1959. [Google Scholar]
- Asad, T. The Kababish Arabs. In Power, Authority and Consent in a Nomadic Tribe; C Hurst and Company: New York, NY, USA, 1970. [Google Scholar]
- MacMichael, H.A. The Tribes of Northern and Central Kordofán; Frank Cass & Co., Ltd.: London, UK, 1967. [Google Scholar]
- Baroin, C. Anarchie et Cohésion Sociale chez les Toubou: Les Daza Késerda (Niger); Cambridge University Press: Cambridge, UK; Editions de la Maison des Sciences de L’homme: Paris, France, 1985. [Google Scholar]
- Chapelle, J. Les Toubous. In Nomades Noires du Sahara; Harmattan: Paris, France, 1967. [Google Scholar]
- Tubiana, M.-J. De Trupeaux et de Femmes. Mariage et Transferts de Biens Chez les Beri (Zaghawa et Bideyat) du Tchad et du Soudan; Harmattan: Paris, France, 1985. [Google Scholar]
- Lhote, H. Les Touaregs du Hoggar (Ahaggar); Payot: Paris, France, 1955. [Google Scholar]
- Rodd, R. People of the Veil; Being an Account of the Habits, Organisation and History of the Wandering Tuareg Tribes Which Inhabit the Mountains of Air, or Asben in the Central Sahara; Macmillan: London, UK, 1926. [Google Scholar]
- Casajus, D. La tente dans la Solitude. In La Société et les Morts Chez les Touaregs Kel Ferwan; Cambridge University Press: Cambridge, UK, 1987. [Google Scholar]
- Xiao, N.; Cai, S.; Moritz, M.; Garabed, R.; Pomeroy, L.W. Spatial and Temporal Characteristics of Pastoral Mobility in the Far North Region, Cameroon: Data Analysis and Modeling. PLoS ONE 2015, 10, e0131697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prussin, L. Afrucan Nomadic Architecture: Space, Place and Gender; Smithsonian: Washington, WA, USA, 1995. [Google Scholar]
- Breman, H.; de Wit, C.T. Rangeland productivity and exploitation in the sahel. Science 1983, 221, 1341–1347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brierley, C.; Manning, K.; Maslin, M. Pastoralism may have delayed the end of the green Sahara. Nat. Commun 2018, 9, 4018. [Google Scholar] [CrossRef] [Green Version]
- Homewood, K. Ecology of African Pastoralist Societies; James Currey: Oxford, UK, 2008. [Google Scholar]
- Wendorf, F.; Schild, R. Holocene Settlement of the Egyptian Sahara: Volume 1: The Archaeology of Nabta Playa; Kluwer Academic/Plenum Publishers: New York, NY, USA, 2001. [Google Scholar]
- Wendorf, F.; Schild, R.; Close, A.E. Cattle-Keepers of the Eastern Sahara. The Neolithic of Bir Kiseiba; Department of Anthropology and Institute for the Study of Earth and Man, Southern Methodist University: Dallas, TX, USA, 1984. [Google Scholar]
- Dunne, J.; di Lernia, S.; Chłodnicki, M.; Kherbouche, F.; Evershed, R. Timing and pace of dairying inception and animal husbandry practices across Holocene North Africa. Quat. Int. 2018, 471, 147–159. [Google Scholar] [CrossRef]
- Neumann, K. The romance of farming: Plant cultivation and domestication in Africa. In African Archaeology: A Critical Introduction; Stahl, A.B., Ed.; Blackwell Publishing Ltd.: Malden, MA, USA, 2005; pp. 249–275. [Google Scholar]
- Phelps, L.N.; Broennimann, O.; Manning, K.; Timpson, A.; Jousse, H.; Mariethoz, G.; Fordham, D.A.; Shanahan, T.M.; Davis, B.A.S.; Guisan, A. Reconstructing the climatic niche breadth of land use for animal production during the African Holocene. Glob. Ecol. Biogeogr. 2020, 29, 127–147. [Google Scholar] [CrossRef]
- Lhote, H. The search for the Tassili frescoes. In The Story of the Prehistoric Rock-Paintings of the Sahara; Lhote, H., Ed.; Dutton: New York, NY, USA, 1959. [Google Scholar]
- Le Quellec, J.-L. Recent Work on Saharan rock art (2010–2014). In Rock Art Studies: News of the World V; Bahn, P., Franklin, N., Strecker, M., Devlet, E., Eds.; Archaeopress Publishing: Oxford, UK, 2016; pp. 55–74. [Google Scholar]
- Wright, D.K. Humans as Agents in the Termination of the African Humid Period. Front. Earth Sci. 2017, 5, 4. [Google Scholar] [CrossRef] [Green Version]
- Linseele, V. From the first stock keepers to specialised pastoralists in the West African Savannah. In Pastoralism in Africa: Past, Present and Future; Bollig, M., Schnegg, M., Wotzka, H.-P., Eds.; Berghan Books: New York, NY, USA, 2013; pp. 145–170. [Google Scholar]
- Sereno, P.C.; Garcea, E.A.; Jousse, H.; Stojanowski, C.M.; Saliege, J.F.; Maga, A.; Ide, O.A.; Knudson, K.J.; Mercuri, A.M.; Stafford, T.W., Jr.; et al. Lakeside cemeteries in the Sahara: 5000 years of holocene population and environmental change. PLoS ONE 2008, 3, e2995. [Google Scholar] [CrossRef]
- Bâ, A.H.; Daget, J.; Monod, T. L’empire Peul du Macina: (1818–1853); Editions de l’Ecole des Hautes Études en Sciences Sociales: Paris, France, 1984. [Google Scholar]
- Shahack-Gross, R. Taphonomy of abandoned Maasai settlememnts in southern Kenya with special reference to livestock enclosures. In Implications for the Spread of Pastoralism in East Africa; Saint Louis University: Saint Louis, MS, USA, 2002. [Google Scholar]
- Černý, V.; Pereira, L.; Musilová, E.; Kujanová, M.; Vašíková, A.; Blasi, P.; Garofalo, L.; Soares, P.; Diallo, I.; Brdička, R.; et al. Genetic structure of pastoral and farmer populations in the African Sahel. Mol. Biol. Evol. 2011, 28, 2491–2500. [Google Scholar] [CrossRef] [Green Version]
- Kulichová, I.; Fernandes, V.; Deme, A.; Nováčková, J.; Stenzl, V.; Novelletto, A.; Pereira, L.; Černý, V. Internal diversification of non-Sub-Saharan haplogroups in Sahelian populations and the spread of pastoralism beyond the Sahara. Am. J. Phys. Anthropol. 2017, 164, 424–434. [Google Scholar] [CrossRef]
- Nováčková, J.; Čížková, M.; Mokhtar, M.G.; Duda, P.; Stenzl, V.; Tříska, P.; Hofmanová, Z.; Černý, V. Subsistence strategy was the main factor driving population differentiation in the bidirectional corridor of the African Sahel. Am. J. Phys. Anthropol. 2020, 171, 496–508. [Google Scholar] [CrossRef]
- Ranciaro, A.; Campbell, M.C.; Hirbo, J.B.; Ko, W.Y.; Froment, A.; Anagnostou, P.; Kotze, M.J.; Ibrahim, M.; Nyambo, T.; Omar, S.A.; et al. Genetic origins of lactase persistence and the spread of pastoralism in Africa. Am. J. Hum. Genet. 2014, 94, 496–510. [Google Scholar] [CrossRef] [Green Version]
- Ségurel, L.; Bon, C. On the Evolution of Lactase Persistence in Humans. Annu. Rev. Genomics. Hum. Genet. 2017, 18, 297–319. [Google Scholar] [CrossRef]
- Priehodová, E.; Austerlitz, F.; Čížková, M.; Nováčková, J.; Ricaut, F.-X.; Hofmanová, Z.; Schlebusch, C.M.; Černý, V. Sahelian pastoralism from the perspective of variants associated with lactase persistence. Am. J. Phys. Anthropol. 2020, 173, 423–436. [Google Scholar] [CrossRef]
- Priehodová, E.; Austerlitz, F.; Čížková, M.; Mokhtar, M.G.; Poloni, E.S.; Černý, V. The historical spread of Arabian Pastoralists to the eastern African Sahel evidenced by the lactase persistence −13,915*G allele and mitochondrial DNA. Am. J. Hum. Biol. 2017, 29, e22950. [Google Scholar] [CrossRef]
- Priehodová, E.; Abdelsawy, A.; Heyer, E.; Černý, V. Lactase Persistence Variants in Arabia and in the African Arabs. Hum. Biol. 2014, 86, 7–18. [Google Scholar] [CrossRef] [Green Version]
- Černý, V.; Fortes-Lima, C.; Tříska, P. Demographic history and admixture dynamics in African Sahelian populations. Hum. Mol. Genet. 2021, 30, R29–R36. [Google Scholar] [CrossRef]
- Campbell, M.C.; Ranciaro, A. Human adaptation, demography and cattle domestication: An overview of the complexity of lactase persistence in Africa. Hum. Mol. Genet. 2021, 30, R98–R109. [Google Scholar] [CrossRef]
- Vicente, M.; Priehodová, E.; Diallo, I.; Podgorná, E.; Poloni, E.S.; Černý, V.; Schlebusch, C.M. Population history and genetic adaptation of the Fulani nomads: Inferences from genome-wide data and the lactase persistence trait. BMC Genom. 2019, 20, 915. [Google Scholar] [CrossRef] [Green Version]
- Enattah, N.S.; Jensen, T.G.; Nielsen, M.; Lewinski, R.; Kuokkanen, M.; Rasinpera, H.; El-Shanti, H.; Seo, J.K.; Alifrangis, M.; Khalil, I.F.; et al. Independent introduction of two lactase-persistence alleles into human populations reflects different history of adaptation to milk culture. Am. J. Hum. Genet. 2008, 82, 57–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Černý, V.; Kulichová, I.; Poloni, E.S.; Nunes, J.M.; Pereira, L.; Mayor, A.; Sanchez-Mazas, A. Genetic history of the African Sahelian populations. Hla 2018, 91, 153–166. [Google Scholar] [CrossRef] [PubMed]
- Tříska, P.; Soares, P.; Patin, E.; Fernandes, V.; Černý, V.; Pereira, L. Extensive admixture and selective pressure across the Sahel Belt. Genome Biol. Evol. 2015, 7, 484–3495. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Mazas, A.; Černý, V.; Da, D.; Buhler, S.; Podgorná, E.; Chevallier, E.; Brunet, L.; Weber, S.; Kervaire, B.; Testi, M.; et al. The HLA-B landscape of Africa: Signatures of pathogen-driven selection and molecular identification of candidate alleles to malaria protection. Mol. Ecol. 2017, 26, 6238–6252. [Google Scholar] [CrossRef] [Green Version]
- Luoni, G.; Verra, F.; Arca, B.; Sirima, B.S.; Troye-Blomberg, M.; Coluzzi, M.; Kwiatkowski, D.; Modiano, D. Antimalarial antibody levels and IL4 polymorphism in the Fulani of West Africa. Genes Immun. 2001, 2, 411–414. [Google Scholar] [CrossRef] [Green Version]
- Modiano, D.; Luoni, G.; Sirima, B.S.; Lanfrancotti, A.; Petrarca, V.; Cruciani, F.; Simpore, J.; Ciminelli, B.M.; Foglietta, E.; Grisanti, P.; et al. The lower susceptibility to Plasmodium falciparum malaria of Fulani of Burkina Faso (west Africa) is associated with low frequencies of classic malaria-resistance genes. Trans. R. Soc. Trop. Med. Hyg. 2001, 95, 149–152. [Google Scholar] [CrossRef]
- Boström, S.; Giusti, P.; Arama, C.; Persson, J.O.; Dara, V.; Traore, B.; Dolo, A.; Doumbo, O.; Troye-Blomberg, M. Changes in the levels of cytokines, chemokines and malaria-specific antibodies in response to Plasmodium falciparum infection in children living in sympatry in Mali. Malar. J. 2012, 11, 109. [Google Scholar] [CrossRef] [Green Version]
- Quin, J.E.; Bujila, I.; Cherif, M.; Sanou, G.S.; Qu, Y.; Vafa Homann, M.; Rolicka, A.; Sirima, S.B.; O’Connell, M.A.; Lennartsson, A.; et al. Major transcriptional changes observed in the Fulani, an ethnic group less susceptible to malaria. Elife 2017, 6, 29156. [Google Scholar] [CrossRef]
- Heyer, E.; Chaix, R.; Pavard, S.; Austerlitz, F. Sex-specific demographic behaviours that shape human genomic variation. Mol. Ecol. 2012, 21, 597–612. [Google Scholar] [CrossRef]
- Garcia-Fernandez, C.; Font-Porterias, N.; Kucinskas, V.; Sukarova-Stefanovska, E.; Pamjav, H.; Makukh, H.; Dobon, B.; Bertranpetit, J.; Netea, M.G.; Calafell, F.; et al. Sex-biased patterns shaped the genetic history of Roma. Sci. Rep. 2020, 10, 14464. [Google Scholar] [CrossRef]
- Silva, M.; Oliveira, M.; Vieira, D.; Brandao, A.; Rito, T.; Pereira, J.B.; Fraser, R.M.; Hudson, B.; Gandini, F.; Edwards, C.; et al. A genetic chronology for the Indian Subcontinent points to heavily sex-biased dispersals. BMC Evol. Biol. 2017, 17, 88. [Google Scholar] [CrossRef] [Green Version]
- Shriner, D.; Rotimi, C.N. Genetic history of Chad. Am. J. Phys. Anthropol. 2018, 167, 804–812. [Google Scholar] [CrossRef]
- Čížková, M.; Munclinger, P.; Diallo, M.Y.; Kulichová, I.; Mokhtar, M.G.; Dème, A.; Pereira, L.; Černý, V. Genetic Structure of the Western and Eastern African Sahel/Savannah Belt and the Role of Nomadic Pastoralists as Inferred from the Variation of D-Loop Mitochondrial DNA Sequences. Hum. Biol. 2017, 89, 281–302. [Google Scholar] [CrossRef]
- Bryc, K.; Auton, A.; Nelson, M.R.; Oksenberg, J.R.; Hauser, S.L.; Williams, S.; Froment, A.; Bodo, J.M.; Wambebe, C.; Tishkoff SA et, a.l. Genome-wide patterns of population structure and admixture in West Africans and African Americans. Proc. Natl. Acad. Sci. USA 2010, 107, 786–791. [Google Scholar] [CrossRef] [Green Version]
- Korunes, K.L.; Soares-Souza, G.B.; Bobrek, K.; Tang, H.; Araújo, I.I.; Goldberg, A.; Beleza, S. Sex-biased admixture and assortative mating shape genetic variation and influence demographic inference in admixed Cabo Verdeans. G3 2022, 12, jkac183. [Google Scholar] [CrossRef]
- Fortes-Lima, C.; Gessain, A.; Ruiz-Linares, A.; Bortolini, M.C.; Migot-Nabias, F.; Bellis, G.; Moreno-Mayar, J.V.; Restrepo, B.N.; Rojas, W.; Avendano-Tamayo, E.; et al. Genome-wide Ancestry and Demographic History of African-Descendant Maroon Communities from French Guiana and Suriname. Am. J. Hum. Genet. 2017, 101, 725–736. [Google Scholar] [CrossRef] [Green Version]
- Fortes-Lima, C.; Tříska, P.; Čížková, M.; Podgorná, E.; Diallo, M.Y.; Schlebusch, C.M.; Černý, V. Demographic and Selection Histories of Populations Across the Sahel/Savannah Belt. Mol. Biol. Evol. 2022, 39, msac209. [Google Scholar] [CrossRef]
- Alexander, D.H.; Lange, K. Enhancements to the ADMIXTURE algorithm for individual ancestry estimation. BMC Bioinform. 2011, 12, 246. [Google Scholar] [CrossRef] [Green Version]
- Diaz-Papkovich, A.; Anderson-Trocme, L.; Ben-Eghan, C.; Gravel, S. UMAP reveals cryptic population structure and phenotype heterogeneity in large genomic cohorts. PLoS Genet. 2019, 15, e1008432. [Google Scholar] [CrossRef] [Green Version]
- Auton, A.; Brooks, L.D.; Durbin, R.M.; Garrison, E.P.; Kang, H.M.; Korbel, J.O.; Marchini, J.L.; McCarthy, S.; McVean, G.A.; Abecasis, G.R. A global reference for human genetic variation. Nature 2015, 526, 68–74. [Google Scholar]
- Gurdasani, D.; Carstensen, T.; Tekola-Ayele, F.; Pagani, L.; Tachmazidou, I.; Hatzikotoulas, K.; Karthikeyan, S.; Iles, L.; Pollard, M.O.; Choudhury, A.; et al. The African Genome Variation Project shapes medical genetics in Africa. Nature 2015, 517, 327–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haber, M.; Mezzavilla, M.; Bergstrom, A.; Prado-Martinez, J.; Hallast, P.; Saif-Ali, R.; Al-Habori, M.; Dedoussis, G.; Zeggini, E.; Blue-Smith, J.; et al. Chad Genetic Diversity Reveals an African History Marked by Multiple Holocene Eurasian Migrations. Am. J. Hum. Genet. 2016, 99, 1316–1324. [Google Scholar] [CrossRef] [Green Version]
- Hollfelder, N.; Schlebusch, C.M.; Gunther, T.; Babiker, H.; Hassan, H.Y.; Jakobsson, M. Northeast African genomic variation shaped by the continuity of indigenous groups and Eurasian migrations. PLoS Genet. 2017, 13, e1006976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, V.; Brucato, N.; Ferreira, J.C.; Pedro, N.; Cavadas, B.; Ricaut, F.X.; Alshamali, F.; Pereira, L. Genome-Wide Characterization of Arabian Peninsula Populations: Shedding Light on the History of a Fundamental Bridge between Continents. Mol. Biol. Evol. 2019, 36, 575–586. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, C.L.; Pita, G.; Cavadas, B.; Lopez, S.; Sanchez-Martinez, L.J.; Dugoujon, J.M.; Novelletto, A.; Cuesta, P.; Pereira, L.; Calderon, R. Human Genomic Diversity Where the Mediterranean Joins the Atlantic. Mol. Biol. Evol. 2020, 37, 1041–1055. [Google Scholar] [CrossRef]
- Patterson, N.; Moorjani, P.; Luo, Y.; Mallick, S.; Rohland, N.; Zhan, Y.; Genschoreck, T.; Webster, T.; Reich, D. Ancient admixture in human history. Genetics 2012, 192, 1065–1093. [Google Scholar] [CrossRef] [Green Version]
- Diallo, M.Y.; Čížková, M.; Kulichová, I.; Podgorná, E.; Priehodová, E.; Nováčková, J.; Fernandes, V.; Pereira, L.; Černý, V. Circum-Saharan Prehistory through the Lens of mtDNA Diversity. Genes 2022, 13, 533. [Google Scholar] [CrossRef]
- Young, W.C. The Rashaayda Bedouin: Arab Pastoralists of Eastern Sudan; Harcourt Brace College Publishers: Fort Worth, UK, 1996. [Google Scholar]
- Lipson, M.; Ribot, I.; Mallick, S.; Rohland, N.; Olalde, I.; Adamski, N.; Broomandkhoshbacht, N.; Lawson, A.M.; Lopez, S.; Oppenheimer, J.; et al. Ancient West African foragers in the context of African population history. Nature 2020, 577, 665–670. [Google Scholar] [CrossRef]
- Fu, Q.; Hajdinjak, M.; Moldovan, O.T.; Constantin, S.; Mallick, S.; Skoglund, P.; Patterson, N.; Rohland, N.; Lazaridis, I.; Nickel, B.; et al. An early modern human from Romania with a recent Neanderthal ancestor. Nature 2015, 524, 216–219. [Google Scholar] [CrossRef] [Green Version]
- Duckworth, C.N.; Cuénod, A.; Mattingly, D.J. Mobile Technologies in the Ancient Sahara and Beyond; Cambridge University Press: Cambridge, UK, 2020. [Google Scholar]
- Maley, J.; Vernet, R. Populations and Climatic Evolution in North Tropical Africa from the End of the Neolithic to the Dawn of the Modern Era. Afr. Archaeol. Rev. 2015, 32, 179–232. [Google Scholar] [CrossRef]
- Lovejoy, P.E. Salt of the Desert Sun: A History of Salt Production and Trade in the Central Sudan; Cambridge University Press: Cambridge, UK, 1986. [Google Scholar]
- McDougall, E.A. Salts of the Western Sahara: Myths, Mysteries, and Historical Significance. Int. J. Afr. Hist. Stud. 1990, 23, 231–257. [Google Scholar] [CrossRef]
- McIntosh, S.K. Long-Distance Exchange and Urban Trajectories in the First Millennium AD. Case Studies from the Middle Niger and Middle Senegal River Valleys. In Urbanisation and State Formation in the Ancient Sahara and Beyond; Sterry, M., Mattingly, D.J., Eds.; Cambridge University Press: Cambridge, UK, 2020; pp. 521–563. [Google Scholar]
- Sherratt, A. Plough and pastoralism: Aspects of the secondary products revolution. In Pattern of the Past: Studies in Honour of David Clarke; Hodder, I., Isaac, G., Hammond, N., Eds.; Cambridge University Press: Cambridge, UK, 1981; pp. 261–305. [Google Scholar]
- Retsö, J. The domestication of the camel and the establisment of the frankincence road from South Arabia. Orient. Suec. 1991, 40, 187–219. [Google Scholar]
- Braukämper, U. Notes on the origin of Baggara Arab culture with special reference to the Shuwa. In Arabs and Arabic in the Lake Chad Region; Ownes, J., Ed.; Rüdiger Köppe Verlag: Köln, German, 1994; Volume Sprache und Geschichte in Africa 14, pp. 13–46. [Google Scholar]
- Schultz, E. From Pagan to Pullo: Ethnic Identity Change in Northern Cameroon. Afr. J. Int. Afr. Inst. 1984, 54, 46–64. [Google Scholar] [CrossRef]
- Barth, F. Ethnic groups and boundaries. The Social Organization of Culture Difference; Universitetsforlaget: Oslo, Norway, 1969. [Google Scholar]
- Mohammadou, E. Traditions Historiques des Peuples du Cameroun Central, Mbere et Mboum Tikar; Ilcaa: Tokyo, Japan, 1990; Volume 1. [Google Scholar]
- de Haan, C.; Dubern, E.; Garancher, B.; Quintero, C. Pastoralism Development in the Sahel A Road to Stability? International Bank for Reconstruction and Development: Washington, WA, USA, 2016. [Google Scholar]
- Ickowicz, A.; Ancey, V.; Corniaux, C.; Duteurtre, G.; Poccard-Chappuis, R.; Touré, I.; Vall, E.; Wane, A. Crop–livestock production systems in the Sahel–increasing resilience for adaptation to climate change and preserving food security. In Building Resilience for Adaptation to Climate Change in the Agriculture Sector; Meybeck, A., Lankoski, J., Redfern, S., Azzu, N., Gitz, V., Eds.; FAO: Rome, Italy, 2012. [Google Scholar]
- Swift, J. Sahelian Pastoralists: Underdevelopment, Desertification, and Famine. Annu. Rev. Anthropol. 1977, 6, 457–478. [Google Scholar] [CrossRef]
- Simonet, C.; Traoré, S.M.; Brunelin, S.; Royer, L. Livestick Markets in the Sahel: Market Integration and the role of Climate and Cconflict in Price Formation; BRACED: London, UK, 2020. [Google Scholar]
- Pereira, L.; Mutesa, L.; Tindana, P.; Ramsay, M. African genetic diversity and adaptation inform a precision medicine agenda. Nat. Rev. Genet. 2021, 22, 284–306. [Google Scholar] [CrossRef]
- Scheinfeldt, L.B.; Soi, S.; Tishkoff, S.A. Colloquium paper: Working toward a synthesis of archaeological, linguistic, and genetic data for inferring African population history. Proc. Natl. Acad. Sci. USA 2010, 107 (Suppl. 2), 8931–8938. [Google Scholar] [CrossRef] [Green Version]
- Hammer, M.F.; Woerner, A.E.; Mendez, F.L.; Watkins, J.C.; Wall, J.D. Genetic evidence for archaic admixture in Africa. Proc. Natl. Acad. Sci. USA 2011, 108, 15123–15128. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.; Pavlidis, P.; Taskent, R.O.; Alachiotis, N.; Flanagan, C.; DeGiorgio, M.; Blekhman, R.; Ruhl, S.; Gokcumen, O. Archaic Hominin Introgression in Africa Contributes to Functional Salivary MUC7 Genetic Variation. Mol. Biol. Evol. 2017, 34, 2704–2715. [Google Scholar] [CrossRef] [Green Version]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Černý, V.; Priehodová, E.; Fortes-Lima, C. A Population Genetic Perspective on Subsistence Systems in the Sahel/Savannah Belt of Africa and the Historical Role of Pastoralism. Genes 2023, 14, 758. https://doi.org/10.3390/genes14030758
Černý V, Priehodová E, Fortes-Lima C. A Population Genetic Perspective on Subsistence Systems in the Sahel/Savannah Belt of Africa and the Historical Role of Pastoralism. Genes. 2023; 14(3):758. https://doi.org/10.3390/genes14030758
Chicago/Turabian StyleČerný, Viktor, Edita Priehodová, and Cesar Fortes-Lima. 2023. "A Population Genetic Perspective on Subsistence Systems in the Sahel/Savannah Belt of Africa and the Historical Role of Pastoralism" Genes 14, no. 3: 758. https://doi.org/10.3390/genes14030758