
Using Next-Generation Sequencing to Disentangle the Diet and Incidence of Intestinal Parasites of Falkland Flightless Steamer Duck Tachyeres brachypterus and Patagonian Crested Duck Lophonetta specularioides Sharing a South Atlantic Island

Abstract
:1. Introduction
2. Materials and Methods
2.1. Studied Species
2.2. Sample Collection
2.3. DNA Isolation
2.4. Construction of Sequencing Library
2.5. Bioinformatics Analyses of Sequences
2.6. Statistical Analyses
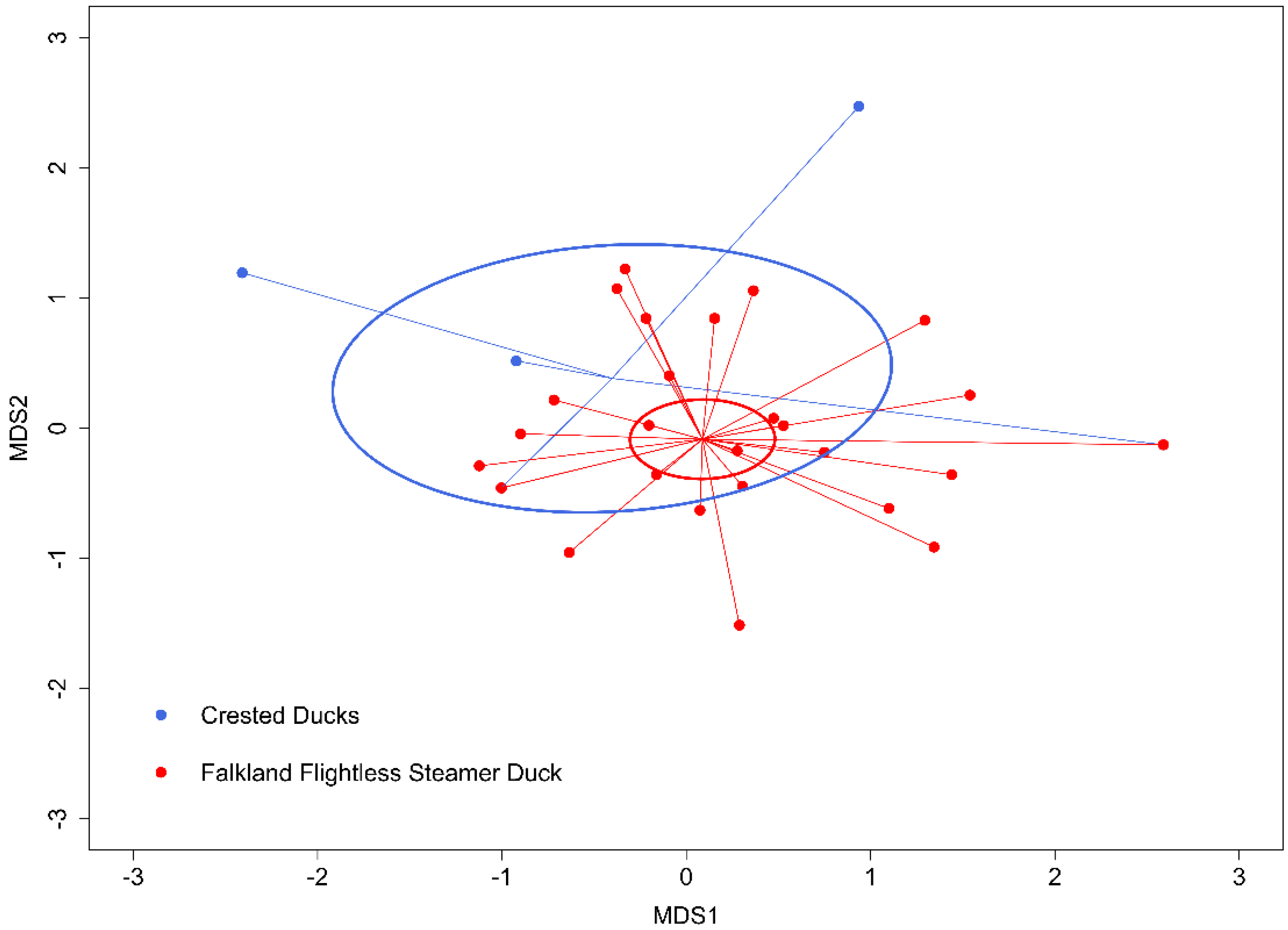
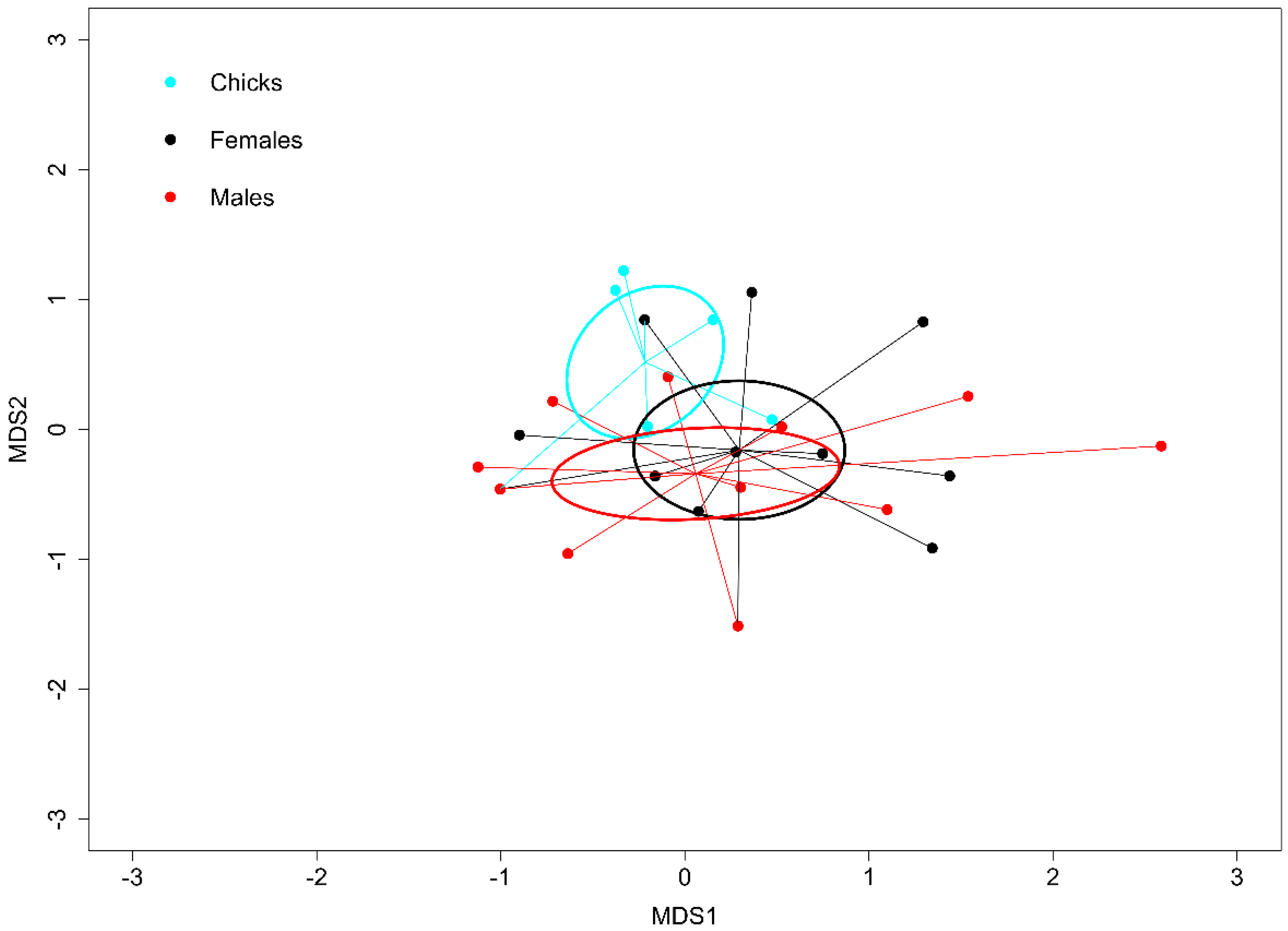
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gause, G.F. The Struggle for Existence; Williams and Wilkins: Baltimore, MD, USA, 1934. [Google Scholar]
- Hardin, G. The competitive exclusion principle. Science 1960, 131, 1292–1297. [Google Scholar] [CrossRef] [Green Version]
- MacArthur, R.H. Population ecology of some warblers of northeastern coniferous forests. Ecology 1958, 39, 599–619. [Google Scholar] [CrossRef]
- Pianka, E.R. Sympatry of desert lizards, and spatial heterogeneity. Ecology 1969, 50, 1012–1030. [Google Scholar] [CrossRef]
- Wakefield, E.D.; Phillips, R.A.; Matthiopoulos, J. Quantifying habitat use and preferences of pelagic seabirds using individual movement data: A review. Mar. Ecol. Prog. Ser. 2009, 391, 165–182. [Google Scholar] [CrossRef] [Green Version]
- Wilson, R.P. Resource partitioning and niche hyper-volume overlap in free-living Pygoscelid penguins. Funct. Ecol. 2010, 24, 646–657. [Google Scholar] [CrossRef]
- Masello, J.F.; Mundry, R.; Poisbleau, M.; Demongin, L.; Voigt, C.C.; Wikelski, M.; Quillfeldt, P. Diving seabirds share foraging space and time within and among species. Ecosphere 2010, 1, 1–28. [Google Scholar] [CrossRef]
- Buckton, S.T.; Ormerod, S.J. Niche segregation of Himalayan river birds. J. Field Ornithol. 2008, 79, 176–185. [Google Scholar] [CrossRef]
- Mansor, M.S.; Rozali, F.Z.; Davies, S.; Nor, S.M.; Ramli, R. High-throughput sequencing reveals dietary segregation in Malaysian babblers. Curr. Zool. 2022, 68, 381–389. [Google Scholar] [CrossRef]
- Pompanon, F.; Deagle, B.E.; Symondson, W.O.C.; Brown, D.S.; Jarman, S.N.; Taberlet, P. Who is eating what: Diet assessment using next generation sequencing. Mol. Ecol. 2012, 21, 1931–1950. [Google Scholar] [CrossRef] [Green Version]
- Masello, J.F.; Barbosa, A.; Kato, A.; Mattern, T.; Medeiros, R.; Stockdale, J.E.; Kümmel, M.N.; Bustamante, P.; Belliure, J.; Benzal, J.; et al. How animals distribute themselves in space: Energy landscapes of Antarctic avian predators. Mov. Ecol. 2021, 9, 24. [Google Scholar] [CrossRef]
- Sutton, G.J.; Bost, C.A.; Kouzani, A.Z.; Adams, S.D.; Mitchell, K.; Arnould, J.P.Y. Fine-scale foraging effort and efficiency of Macaroni penguins is influenced by prey type, patch density and temporal dynamics. Mar. Biol. 2021, 168, 3. [Google Scholar] [CrossRef]
- Estes, J.A.; Terborgh, J.; Brashares, J.S.; Power, M.E.; Berger, J.; Bond, W.J.; Carpenter, S.R.; Essington, T.E.; Holt, R.D.; Jackson, J.B.C.; et al. Trophic Downgrading of Planet Earth. Science 2011, 333, 301–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sydeman, W.J.; Poloczanska, E.; Reed, T.E.; Thompson, S.A. Climate change and marine vertebrates. Science 2015, 350, 772–777. [Google Scholar] [CrossRef] [Green Version]
- Cabodevilla, X.; Mougeot, F.; Bota, G.; Mañosa, S.; Cuscó, F.; Martínez-García, J.; Arroyo, B.; Madeira, M.J. Metabarcoding insights into the diet and trophic diversity of six declining farmland birds. Sci. Rep. 2021, 11, 21131. [Google Scholar] [CrossRef] [PubMed]
- Traugott, M.; Kamenova, S.; Ruess, L.; Seeber, J.; Plantegenest, M. Empirically characterising trophic networks: What emerging DNA-based methods, stable isotope and fatty acid analyses can offer. Adv. Ecol. Res. 2013, 49, 177–224. [Google Scholar]
- Gaglio, D.; Cook, T.R.; Connan, M.; Ryan, P.G.; Sherley, R.B. Dietary studies in birds: Testing a non-invasive method using digital photography in seabirds. Methods Ecol. Evol. 2017, 8, 214–222. [Google Scholar] [CrossRef] [Green Version]
- Deagle, B.E.; Gales, N.J.; Evans, K.; Jarman, S.N.; Robinson, S.; Trebilco, R.; Hindell, M.A. Studying Seabird Diet through Genetic Analysis of Faeces: A Case Study on Macaroni Penguins (Eudyptes chrysolophus). PLoS ONE 2007, 2, e831. [Google Scholar] [CrossRef] [Green Version]
- Michalik, A.; van Noordwijk, H.; Brickle, P.; Eggers, T.; Quillfeldt, P. The diet of the Imperial Shag Phalacrocorax atriceps at a colony on New Island, Falkland Islands using three different sampling techniques. Polar Biol. 2010, 33, 1537–1546. [Google Scholar] [CrossRef]
- Swift, J.F.; Lance, R.F.; Guan, X.; Britzke, E.R.; Lindsay, D.L.; Edwards, C.E. Multifaceted DNA metabarcoding: Validation of a noninvasive, next-generation approach to studying bat populations. Evol. Appl. 2018, 11, 1120–1138. [Google Scholar] [CrossRef]
- Alberdi, A.; Aizpurua, O.; Bohmann, K.; Gopalakrishnan, S.; Lynggaard, C.; Nielsen, M.; Gilbert, M.T.P. Promises and pitfalls of using high-throughput sequencing for diet analysis. Mol. Ecol. Resour. 2019, 19, 327–348. [Google Scholar] [CrossRef]
- Deagle, B.E.; Thomas, A.C.; McInnes, J.C.; Clarke, L.J.; Vesterinen, E.J.; Clare, E.L.; Kartzinel, T.R.; Eveson, J.P. Counting with DNA in metabarcoding studies: How should we convert sequence reads to dietary data? Mol. Ecol. 2019, 28, 391–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowser, A.K.; Diamond, A.W.; Addison, J.A. From Puffins to Plankton: A DNA-Based Analysis of a Seabird Food Chain in the Northern Gulf of Maine. PLoS ONE 2013, 8, e83152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, M.; Ning, Y.; Han, M.; Zhao, C.; Tian, J.; Li, L.; Xiao, H.; Liu, G. A comparison of next-generation sequencing with clone sequencing in the diet analysis of Asian great bustard. Conserv. Genet. Resour. 2019, 11, 15–17. [Google Scholar] [CrossRef]
- Dunn, J.C.; Stockdale, J.E.; Moorhouse-Gann, R.J.; McCubbin, A.; Hipperson, H.; Morris, A.J.; Grice, P.V.; Symondson, W.O.C. The decline of the Turtle Dove: Dietary associations with body condition and competition with other columbids analysed using high-throughput sequencing. Mol. Ecol. 2018, 27, 3386–3407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleinschmidt, B.; Burger, C.; Dorsch, M.; Nehls, G.; Heinänen, S.; Morkūnas, J.; Žydelis, R.; Moorhouse-Gann, R.J.; Hipperson, H.; Symondson, W.O.C.; et al. The diet of red-throated divers (Gavia stellata) overwintering in the German Bight (North Sea) analysed using molecular diagnostics. Mar. Biol. 2019, 166, e77. [Google Scholar] [CrossRef]
- Deagle, B.E.; Chiaradia, A.; McInnes, J.; Jarman, S.N. Pyrosequencing faecal DNA to determine diet of little penguins: Is what goes in what comes out? Conserv. Genet. 2010, 11, 2039–2048. [Google Scholar] [CrossRef]
- Rytkönen, S.; Vesterinen, E.J.; Westerduin, C.; Leviäkangas, T.; Vatka, E.; Mutanen, M.; Välimäki, P.; Hukkanen, M.; Suokas, M.; Orell, M. From feces to data: A metabarcoding method for analyzing consumed and available prey in a bird-insect food web. Ecol. Evol. 2019, 9, 631–639. [Google Scholar] [CrossRef] [Green Version]
- McInnes, J.C.; Jarman, S.N.; Lea, M.-A.; Raymond, B.; Deagle, B.E.; Phillips, R.A.; Catry, P.; Stanworth, A.; Weimerskirch, H.; Kusch, A.; et al. DNA Metabarcoding as a Marine Conservation and Management Tool: A Circumpolar Examination of Fishery Discards in the Diet of Threatened Albatrosses. Front. Mar. Sci. 2017, 4, 277. [Google Scholar] [CrossRef]
- McCosker, C.; Flanders, K.; Ono, K.; Dufault, M.; Mellone, D.; Olson, Z. Metabarcoding Fecal DNA Reveals Extent of Halichoerus grypus (Gray Seal) Foraging on Invertebrates and Incidence of Parasite Exposure. Northeast. Nat. 2020, 27, 681–700. [Google Scholar] [CrossRef]
- Montalva, F.; Perez-Venegas, D.J.; Gutiérrez, J.; Seguel, M. The contrasting hidden consequences of parasitism: Effects of a hematophagous nematode (Uncinaria sp.) in the development of a marine mammal swimming behavior. Ecol. Evol. 2019, 9, 3689–3699. [Google Scholar] [CrossRef]
- Baylis, A.M.M.; de Lecea, A.M.; Tierney, M.; Orben, R.A.; Ratcliffe, N.; Wakefield, E.; Catry, P.; Campioni, L.; Costa, M.; Boersma, P.D.; et al. Overlap between marine predators and proposed Marine Managed Areas on the Patagonian Shelf. Ecol. Appl. 2021, 31, e02426. [Google Scholar] [CrossRef] [PubMed]
- Woods, R.W.; Woods, A. Atlas of Breeding Birds of the Falkland Islands; Anthony Nelson: Oswestry, UK, 1997. [Google Scholar]
- Augé, A.A.; Dias, M.P.; Lascelles, B.; Baylis, A.M.M.; Black, A.; Boersma, P.D.; Catry, P.; Crofts, S.; Galimberti, F.; Granadeiro, J.P.; et al. Framework for mapping key areas for marine megafauna to inform Marine Spatial Planning: The Falkland Islands case study. Mar. Policy 2018, 92, 61–72. [Google Scholar] [CrossRef] [Green Version]
- Strange, I.; Catry, P.; Strange, G.; Quillfeldt, P. New Island, Falkland Islands. A South Atlantic Wildlife Sanctuary for Conservation Management; Strange, G., Strange, M., Eds.; New Island Conservation Trust: Stanley, Falkland Islands, 2007. [Google Scholar]
- Agnew, D.J. Critical aspects of the Falkland Islands pelagic ecosystem: Distribution, spawning and migration of pelagic animals in relation to oil exploration. Aquat. Conserv. Mar. Freshw. Ecosyst. 2002, 12, 39–50. [Google Scholar] [CrossRef]
- Barlow, K.E.; Boyd, I.L.; Croxall, J.P.; Reid, K.; Staniland, I.J.; Brierley, A.S. Are penguins and seals in competition for Antarctic krill at South Georgia? Mar. Biol. 2002, 140, 205–213. [Google Scholar]
- Neely, K.; Brickle, P. Marine Life of the Falkland Islands; Shallow Marine Surveys Group: Stanley, Falkland Islands, 2013; p. 106. [Google Scholar]
- Quillfeldt, P.; Michalik, A.; Veit-Köhler, G.; Strange, I.J.; Masello, J.F. Inter-annual changes in diet and foraging trip lengths in a small pelagic seabird, the Thin-billed prion Pachyptila belcheri. Mar. Biol. 2010, 157, 2043–2050. [Google Scholar] [CrossRef]
- Masello, J.F.; Wikelski, M.; Voigt, C.C.; Quillfeldt, P. Distribution patterns predict individual specialization in the diet of Dolphin Gulls. PLoS ONE 2013, 8, e67714. [Google Scholar] [CrossRef]
- Strange, I.; Campos, A.; Catry, P.; Masello, J.F.; Quillfeldt, P. Area description, breeding bird status and conservation notes of the ‘New Island Group’ in section ‘FK011: Falkland Islands’ in chapter ‘Falkland Islands’. In Important Bird Areas in the United Kingdom Overseas Territories; Sanders, S., Ed.; RSPB: Sandy, UK, 2006; pp. 131–133. [Google Scholar]
- Weller, M.W. Ecology and behaviour of steamer ducks. Wildfowl J. 1976, 27, 45–53. [Google Scholar]
- Woods, R.W. The Birds of the Falkland Islands; Anthony Nelson: Oswestry, UK, 1975; p. 240. [Google Scholar]
- Weller, M.W. Ecological studies of Falkland Islands’ waterfowl. Wildfowl J. 1972, 23, 25–44. [Google Scholar]
- Jarman, S.N.; Deagle, B.E.; Gales, N.J. Group-specific polymerase chain reaction for DNA-based analysis of species diversity and identity in dietary samples. Mol. Ecol. 2004, 13, 1313–1322. [Google Scholar] [CrossRef]
- Palumbi, S.; Martin, A.; Romano, S.; McMillian, W.O.; Stice, L.; Grabowski, G. The Simple Fool’s Guide to PCR; University of Hawaii: Honolulu, HI, USA, 1991. [Google Scholar]
- Xavier, J.C.; Cherel, Y.; Medeiros, R.; Velez, N.; Dewar, M.; Ratcliffe, N.; Carreiro, A.R.; Trathan, P.N. Conventional and molecular analysis of the diet of gentoo penguins: Contributions to assess scats for non-invasive penguin diet monitoring. Polar Biol. 2018, 41, 2275–2287. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Afgan, E.; Baker, D.; Batut, B.; van den Beek, M.; Bouvier, D.; Čech, M.; Chilton, J.; Clements, D.; Coraor, N.; Grüning, B.A.; et al. The Galaxy platform for accessible, reproducible and collaborative biomedical analyses: 2018 update. Nucleic Acids Res. 2018, 46, W537–W544. [Google Scholar] [CrossRef] [Green Version]
- Deagle, B.E.; Kirkwood, R.; Jarman, S.N. Analysis of Australian fur seal diet by pyrosequencing prey DNA in faeces. Mol. Ecol. 2009, 18, 2022–2038. [Google Scholar] [CrossRef]
- Vesterinen, E.J.; Lilley, T.; Laine, V.N.; Wahlberg, N. Next generation sequencing of fecal DNA reveals the dietary diversity of the widespread insectivorous predator Daubenton’s bat (Myotis daubentonii) in Southwestern Finland. PLoS ONE 2013, 8, e82168. [Google Scholar] [CrossRef] [Green Version]
- Barrett, R.T.; Camphuysen, K.; Anker-Nilssen, T.; Chardine, J.W.; Furness, R.W.; Garthe, S.; Hüppop, O.; Leopold, M.F.; Montevecchi, W.A.; Veit, R.R. Diet studies of seabirds: A review and recommendations. ICES J. Mar. Sci. 2007, 64, 1675–1691. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.; Simpson, G.L.; Solymos, P.; Stevens, M.; Wagner, H. vegan: Community Ecology Package. R Package Version 2.0-2. 2011. 2012. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 13 March 2023).
- Faith, D.P.; Minchin, P.R.; Belbin, L. Compositional dissimilarity as a robust measure of ecological distance. Vegetatio 1987, 69, 57–68. [Google Scholar] [CrossRef]
- Minchin, P.R. An evaluation of the relative robustness of techniques for ecological ordination. Vegetatio 1987, 69, 89–107. [Google Scholar] [CrossRef]
- Davidson, S.C.; Bohrer, G.; Gurarie, E.; LaPoint, S.; Mahoney, P.J.; Boelman, N.T.; Eitel, J.U.H.; Prugh, L.R.; Vierling, L.A.; Jennewein, J.; et al. Ecological insights from three decades of animal movement tracking across a changing Arctic. Science 2020, 370, 712–715. [Google Scholar] [CrossRef]
- Jetz, W.; Tertitski, G.; Kays, R.; Mueller, U.; Wikelski, M.; Åkesson, S.; Anisimov, Y.; Antonov, A.; Arnold, W.; Bairlein, F.; et al. Biological earth observation with animal sensors. Trends Ecol. Evol. 2022, 37, 293–298. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Deng, J.; Leen, T.; Li, G.; Klaassen, M. Continuous on-board behaviour classification using accelerometry: A case study with a new GPS-3G-Bluetooth system in Pacific black ducks. Methods Ecol. Evol. 2022, 13, 1429–1435. [Google Scholar] [CrossRef]
- Croxall, J.P.; Naito, Y.; Kato, A.; Rothery, P.; Briggs, D.R. Diving patterns and performance in the Antarctic Blue-eyed Shag Phalacrocorax atriceps. J. Zool. 1991, 225, 177–199. [Google Scholar] [CrossRef]
- Kato, A.; Nishiumi, I.; Naito, Y. Sexual difference in the diet of King Cormorants at Macquarie Island. Polar Biol. 1996, 16, 75–77. [Google Scholar] [CrossRef]
- Cook, T.R.; Cherel, Y.; Bost, C.A.; Tremblayz, M. Chick-rearing Crozet shags (Phalacrocorax melanogenis) display sex-specific foraging behaviour. Antarct. Sci. 2007, 19, 55–63. [Google Scholar] [CrossRef] [Green Version]
- Weimerskirch, H.; Shaffer, S.A.; Tremblay, Y.; Costa, D.P.; Gadenne, H.; Kato, A.; Ropert-Coudert, Y.; Sato, K.; Aurioles, D. Species- and sex-specific differences in foraging behaviour and foraging zones in blue-footed and brown boobies in the Gulf of California. Mar. Ecol. Prog. Ser. 2009, 391, 267–278. [Google Scholar] [CrossRef] [Green Version]
- Quillfeldt, P.; Schroff, S.; van Noordwijk, H.J.; Michalik, A.; Ludynia, K.; Masello, J.F. Flexible foraging behavior of a sexually dimorphic seabird: Large males do not always dive deep. Mar. Ecol. Prog. Ser. 2011, 428, 271–287. [Google Scholar] [CrossRef] [Green Version]
- Diaz, J.I.; Fusaro, B.; Longarzo, L.; Coria, N.R.; Vidal, V.; Jerez, S.; Ortiz, J.; Barbosa, A. Gastrointestinal helminths of Gentoo penguins (Pygoscelis papua) from Stranger Point, 25 de Mayo/King George Island, Antarctica. Parasitol. Res. 2013, 112, 1877–1881. [Google Scholar] [CrossRef]
- Claar, D.C.; Kuris, A.; Leslie, K.L.; Welicky, R.L.; Williams, M.A.; Wood, C.L. Parasite Biodiversity. Lessons Conserv. 2021, 11, 39–57. [Google Scholar]
- Galaktionov, K.; Bustnes, J.O. Species composition and prevalence of seabird trematode larvae in periwinkles at two littoral sites in North-Norway. Sarsia 1995, 80, 187–191. [Google Scholar] [CrossRef]
- Galaktionov, K.V.; Bustnes, J.O.; Bårdsen, B.-J.; Wilson, J.G.; Nikolaev, K.E.; Sukhotin, A.A.; Skírnisson, K.; Saville, D.H.; Ivanov, M.V.; Regel, K.V. Factors influencing the distribution of trematode larvae in blue mussels Mytilus edulis in the North Atlantic and Arctic Oceans. Mar. Biol. 2015, 162, 193–206. [Google Scholar] [CrossRef]
- Galaktionov, K.V.; Węsławski, J.M.; Stempniewicz, L. Food chain, parasites and climate changes in the high Arctic: A case study on trophically transmitted parasites of common eider Somateria mollissima at Franz Josef Land. Polar Biol. 2021, 44, 1321–1342. [Google Scholar] [CrossRef]
- Vestbo, S.; Hindberg, C.; Forbes, M.R.; Mallory, M.L.; Merkel, F.; Steenweg, R.J.; Funch, P.; Gilchrist, H.G.; Robertson, G.J.; Provencher, J.F. Helminths in common eiders (Somateria mollissima): Sex, age, and migration have differential effects on parasite loads. Int. J. Parasitol. Parasites Wildl. 2019, 9, 184–194. [Google Scholar] [CrossRef]
- Møller, A.P. Parasites and sexual selection: Current status of the Hamilton and Zuk hypothesis. J. Evol. Biol. 1990, 3, 319–328. [Google Scholar] [CrossRef] [Green Version]
- De Lope, F.; Gonzalez, G.; Pérez, J.J.; Møller, A.P. Increased detrimental effects of ectoparasites on their bird hosts during adverse environmental conditions. Oecologia 1993, 95, 234–240. [Google Scholar] [CrossRef]
- Hestetun, J.T.; Bye-Ingebrigtsen, E.; Nilsson, R.H.; Glover, A.G.; Johansen, P.-O.; Dahlgren, T.G. Significant taxon sampling gaps in DNA databases limit the operational use of marine macrofauna metabarcoding. Mar. Biodivers. 2020, 50, 70. [Google Scholar] [CrossRef]
- Leite, B.R.; Vieira, P.E.; Teixeira, M.A.L.; Lobo-Arteaga, J.; Hollatz, C.; Borges, L.M.S.; Duarte, S.; Troncoso, J.S.; Costa, F.O. Gap-analysis and annotated reference library for supporting macroinvertebrate metabarcoding in Atlantic Iberia. Reg. Stud. Mar. Sci. 2020, 36, 101307. [Google Scholar] [CrossRef]
- Specchia, V.; Tzafesta, E.; Marini, G.; Scarcella, S.; D’Attis, S.; Pinna, M. Gap Analysis for DNA Barcode Reference Libraries for Aquatic Macroinvertebrate Species in the Apulia Region (Southeast of Italy). J. Mar. Sci. Eng. 2020, 8, 538. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Prey Target | Gene | Primer Name | Sequence 5′-3′ | Annealing Temperature (°C) | Amplicon Size (bp) | Reference |
|---|---|---|---|---|---|---|
| Bilateralia | Nuclear 18S | BilSSU110f | AGAGGTGAAATTSTTGGAYCG | 60 | ∼245 | [45] |
| BilSSU1300r | CCTTTAAGTTTCAGCTTTGCA | |||||
| Mollusca | 16S rRNA | L2510 | CGCCTGTTTATCAAAAACAT | 50 | ~350 | [46] |
| H3059 | TTTCCCCGCGGTCGCCCC | |||||
| Osteichthyes | Mitochondrial 12S | FishF1 | CGGTAAAACTCGTGCC | 56 | ~300 | [47] |
| FishR1 | CCGCCAAGTCCTTTGGG |
| Phylum | Class | Order | Family | Species | Common Name | Falkland Flightless Steamer Duck | Patagonian Crested Duck | |||
|---|---|---|---|---|---|---|---|---|---|---|
| Females | Males | Chicks | Adults | Chicks | ||||||
| Annelida | Polychaeta | Sabelida | Sabellidae | feather duster worms | - | - | - | 14 | - | |
| Terebellida | Terebelldae | bristle worms | 7 | 20 | 10 | - | - | |||
| Arthropoda | Copepoda | Calanoida | - | - | 18 | 43 | 75 | |||
| Diaptomidae | - | - | - | 14 | - | |||||
| Harpacticoida | - | 7 | - | - | - | |||||
| Branchiopoda | Anostraca | fairy shrimps | 14 | 7 | - | - | - | |||
| Diplostraca | clam shrimps | 7 | 27 | 9 | - | 25 | ||||
| Daphniidae | water fleas | 7 | 27 | 9 | - | - | ||||
| Lynceidae | - | - | - | - | 25 | |||||
| Malacostraca | Decapoda | Munididae | lobster krill | 7 | 13 | - | 14 | - | ||
| Paguridae | hermit crabs | - | 7 | 9 | - | - | ||||
| Portunidae | 29 | 20 | 18 | - | - | |||||
| Euphausiacea | Euphausiidae | krill | - | - | - | 14 | 25 | |||
| Isopoda | isopods | 43 | 67 | 18 | 43 | 25 | ||||
| Serolidae | Serolidae marine isopods | 21 | 13 | - | - | - | ||||
| Sphaeromatidae | marine pill bugs | 36 | 53 | 18 | 43 | 25 | ||||
| Arachnida | Oribatida | moss mites | 7 | - | - | - | - | |||
| Trombidiformes | Halacaridae | meiobenthic mites | - | 7 | - | - | - | |||
| Insecta | Coleoptera | beetles | 21 | 20 | - | - | - | |||
| Dytiscidae | predaceous diving beetles | - | 7 | - | - | - | ||||
| Diptera | flies | - | - | - | 14 | - | ||||
| Mollusca | Polyplacophora | sea cradles | 7 | 13 | - | - | - | |||
| Bivalvia | bivalves | 21 | 7 | 9 | 29 | - | ||||
| Mytilida | Mytilidae | mussels | 7 | 7 | - | - | - | |||
| Venerida | - | - | - | 14 | - | |||||
| Gastropoda | Nacellidae | Nacella sp. | 57 | 33 | 36 | - | - | |||
| Neogastropoda | Cominellidae | Pareuthria plumbea | leaden whelk | 29 | 7 | 18 | - | - | ||
| Fissurelloidea | Fissurellidae | keyhole limpets | - | 7 | - | - | - | |||
| Trochida | Turbinidae | star snails | 21 | 20 | 45 | 14 | - | |||
| Echinodermata | Holothuroidea | sea cucumbers | - | 7 | - | - | - | |||
| Asteroidea | sea stars | 7 | 13 | - | - | - | ||||
| Chordata | Ascidiacea | Stolidobranchia | Molgulidae | ascidians | 7 | - | - | 14 | - | |
| Order | Family | Species | Common Name | Falkland Flightless Steamer Duck | Patagonian Crested Duck | |||
|---|---|---|---|---|---|---|---|---|
| Females | Males | Chicks | Adults | Chicks | ||||
| fishes | 43 | 20 | 45 | - | - | |||
| Clupeiformes | Clupeidae | herrings | - | 7 | - | - | - | |
| Engraulidae | anchovies | - | 13 | - | - | - | ||
| Myctophiformes | Myctophidae | Gymnoscopelus braueri | Brauer’s lanternfish | 7 | - | - | - | - |
| Scorpaeniformes | Agonidae | alligatorfishes | 7 | - | - | - | - | |
| Perciformes | 43 | 20 | 9 | - | - | |||
| Eleginopsidae | Eleginops sp. | 7 | - | - | - | - | ||
| Eleginops maclovinus | Patagonian blennie | 7 | - | - | - | - | ||
| Channichthyidae | crocodile icefishes | 7 | - | - | - | - | ||
| Carangidae | Jacks | - | 7 | - | - | - | ||
| Sparidae | sea bream fishes | 29 | 20 | 9 | - | - | ||
| Scombridae | Scomber sp. | mackerels | - | 7 | - | - | - | |
| Nototheniidae | Patagonotothen sp. | 38 | 20 | 9 | - | - | ||
| Patagonotothen longipes | Antarctic blennies | 7 | 13 | 9 | - | - | ||
| Patagonotothen ramsayi | cod icefish | 21 | 13 | 9 | - | - | ||
| Patagonotothen sima | humped rockcod | 29 | 20 | 9 | - | - | ||
| Class | Order | Family | Common name | Falkland Flightless Steamer Duck | Patagonian Crested Duck | |||
|---|---|---|---|---|---|---|---|---|
| Females | Males | Chicks | Adults | Chicks | ||||
| Cestoda | parasitic tapeworm | 29 | 53 | 55 | 29 | 25 | ||
| Bothriocephalidea | Bothriocephalidae | 7 | - | - | - | - | ||
| Cyclophyllidea | - | - | - | 14 | - | |||
| Hymenolepididae | - | 7 | 9 | - | - | |||
| Trematoda | parasitic flatworms/flukes | 64 | 20 | 18 | - | 25 | ||
| Plagiorchiida | 29 | 7 | 18 | - | 25 | |||
| Notocotylidae | 14 | 7 | - | - | 25 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Masello, J.F.; Schumm, Y.R.; Griep, S.; Quillfeldt, P. Using Next-Generation Sequencing to Disentangle the Diet and Incidence of Intestinal Parasites of Falkland Flightless Steamer Duck Tachyeres brachypterus and Patagonian Crested Duck Lophonetta specularioides Sharing a South Atlantic Island. Genes 2023, 14, 731. https://doi.org/10.3390/genes14030731
Masello JF, Schumm YR, Griep S, Quillfeldt P. Using Next-Generation Sequencing to Disentangle the Diet and Incidence of Intestinal Parasites of Falkland Flightless Steamer Duck Tachyeres brachypterus and Patagonian Crested Duck Lophonetta specularioides Sharing a South Atlantic Island. Genes. 2023; 14(3):731. https://doi.org/10.3390/genes14030731
Chicago/Turabian StyleMasello, Juan F., Yvonne R. Schumm, Sven Griep, and Petra Quillfeldt. 2023. "Using Next-Generation Sequencing to Disentangle the Diet and Incidence of Intestinal Parasites of Falkland Flightless Steamer Duck Tachyeres brachypterus and Patagonian Crested Duck Lophonetta specularioides Sharing a South Atlantic Island" Genes 14, no. 3: 731. https://doi.org/10.3390/genes14030731