
Association of NEF2L2 Rs35652124 Polymorphism with Nrf2 Induction and Genotoxic Stress Biomarkers in Autism

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Cohorts
- Autism Mental Status Examination (AMSE) is an 8-item observational assessment that prompts the examiner to observe and document patients’ social, communicative, and behavioral functioning in the context of a routine clinical examination. The AMSE was developed by psychiatrists with autism expertise and is intended to guide clinical judgment in the context of diagnostic decision-making.
- Childhood Autism Rating Scale (CARS) is a scale for the quantification of the severity of autism pathology. The CARS assesses the child on a scale from 1 to 4 in each of 15 dimensions or symptoms (relating to people; emotional response; imitation; body use; object use; listening response; fear or nervousness; verbal communication; non-verbal communication; activity level; level and reliability of intellectual response; adaptation to change; visual response; taste, smell and touch response; and general impressions). Total scores of or above 30 strongly suggest the presence of autism. Children who have a score from 30 to 36 have mild to moderate autism, while those with scores ranging from 37 to 60 points have severe autism.
- Communication Questionnaire (SCQ) is a parent questionnaire designed for detecting risk for ASD. The SCQ was originally designed as a screening tool for children 4 years of age or older enrolled in epidemiological research or for studies comparing individuals with ASD and other clinical groups.
2.2. Blood Sampling
2.3. SNP Selection Criteria
2.4. Polymerase Chain Reaction
- Direct external (F1)—5′-GTCGCTGGAGTTCGGACGCTT-3′,
- Reverse external (R1)—5′-GCTTTGGTGGGAAGAGGTTCT-3′,
- Direct internal (F2)—5′-TCGCAGTCACCCTGAACGCCCT-3′,
- Reverse internal (R2)—5′-AGACACGTGGGAGTTCAGAGGG-3′.
2.5. Flow Cytometry
2.6. Evaluation of Gene Expression Using Real-Time PCR
2.7. Extraction of cfDNA Fragments from Plasma and cfDNA Quantification
2.8. Measurement of DNase1 Activity
2.9. Statistical Analysis
3. Results
3.1. Nrf2 rs35652124 Genotype Frequencies in Children with ASD and Healthy Controls
3.2. Adaptive Responses for Genotoxic Stress and Their Modulation with NRF2 Inducer in ASD Children Mononuclear Blood Cells
4. Discussion
4.1. Allelic Frequencies and Possible Clinical Significance
4.2. Molecular Markers
4.3. Nrf2 Induction in the Carriers of Different NFE2L2 Genotypes
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Lord, C. For Better or for Worse? Later Diagnoses of Autism Spectrum Disorder in Some Younger Siblings of Already Diagnosed Children. J. Am. Acad. Child Adolesc. Psychiatry 2018, 57, 822–823. [Google Scholar] [CrossRef]
- Chaste, P.; Leboyer, M. Autism risk factors: Genes, environment, and gene-environment interactions. Dialogues Clin. Neurosci. 2012, 14, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, N.; Adak, P. Birth related parameters are important contributors in autism spectrum disorders. Sci. Rep. 2022, 12, 14277. [Google Scholar] [CrossRef] [PubMed]
- Smaga, I.; Niedzielska, E.; Gawlik, M.; Moniczewski, A.; Krzek, J.; Przegaliński, E.; Pera, J.; Filip, M. Oxidative stress as an etiological factor and a potential treatment target of psychiatric disorders. Part Depression, anxiety, schizophrenia and autism. Pharmacol. Rep. 2015, 67, 569–580. [Google Scholar] [CrossRef] [PubMed]
- Bjørklund, G.; Meguid, N.A.; El-Bana, M.A.; Tinkov, A.A.; Saad, K.; Dadar, M.; Hemimi, M.; Skalny, A.V.; Hosnedlová, B.; Kizek, R.; et al. Oxidative Stress in Autism Spectrum Disorder. Mol. Neurobiol. 2020, 57, 2314–2332. [Google Scholar] [CrossRef]
- Manivasagam, T.; Arunadevi, S.; Essa, M.M.; Saravana Babu, C.; Borah, A.; Thenmozhi, A.J.; Qoronfleh, M.W. Role of Oxidative Stress and Antioxidants in Autism. Adv. Neurobiol. 2020, 24, 193–206. [Google Scholar]
- Markkanen, E.; Meyer, U.; Dianov, G.L. DNA Damage and Repair in Schizophrenia and Autism: Implications for Cancer Comorbidity and Beyond. Int. J. Mol. Sci. 2016, 17, 856. [Google Scholar] [CrossRef] [Green Version]
- Réus, G.Z.; Fries, G.R.; Stertz, L.; Badawy, M.; Passos, I.C.; Barichello, T.; Kapczinski, F.; Quevedo, J. The role of inflammation and microglial activation in the pathophysiology of psychiatric disorders. Neuroscience 2015, 300, 141–154. [Google Scholar] [CrossRef]
- Rossignol, D.A.; Frye, R.E. Evidence linking oxidative stress, mitochondrial dysfunction, and inflammation in the brain of individuals with autism. Front. Physiol. 2014, 5, 150. [Google Scholar] [CrossRef] [Green Version]
- Moi, P.; Chan, K.; Asunis, I.; Cao, A.; Kan, Y.W. Isolation of NF-E2-related factor 2 (Nrf2), a NF-E2-like basic leucine zipper transcriptional activator that binds to the tandem NF-E2/AP1 repeat of the β-globin locus control region. Proc. Natl. Acad. Sci. USA 1994, 91, 9926–9930. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Li, C.Y.; Kong, A.N. Induction of phase I, II and III drug metabolism/transport by xenobiotics. Arch. Pharm. Res. 2005, 28, 249–268. [Google Scholar] [CrossRef] [PubMed]
- Wakabayashi, N.; Slocum, S.L.; Skoko, J.J.; Shin, S.; Kensler, T.W. When NRF2 talks, who’s listening? Antioxid. Redox Signal. 2010, 13, 1649–1663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, H.Y.; Marzec, J.; Kleeberger, S.R. Functional polymorphisms in NRF2: Implications for human disease. Free Radic. Biol. Med. 2015, 88, 362–372. [Google Scholar] [CrossRef]
- Chumachenko, A.G.; Myazin, A.E.; Kuzovlev, A.N.; Gaponov, A.M.; Tutelyan, A.V.; Porokhovnik, L.N.; Golubev, A.M.; Pisarev, V.M. Allelic Variants of NRF2 and TLR9 Genes in Critical Illness. Gen. Reanimatol. 2016, 12, 8–23. [Google Scholar] [CrossRef]
- Yu, B.; Lin, H.; Yang, L.; Chen, K.; Luo, H.; Liu, J.; Gao, X.; Xia, X.; Huang, Z. Genetic variation in the Nrf2 promoter associates with defective spermatogenesis in humans. J. Mol. Med. 2012, 90, 1333–1342. [Google Scholar] [CrossRef] [PubMed]
- Figarska, S.M.; Vonk, J.M.; Boezen, H.M. NFE2L2 polymorphisms, mortality, and metabolism in the general population. Physiol. Genom. 2014, 46, 411–417. [Google Scholar] [CrossRef]
- Xu, X.; Sun, J.; Chang, X.; Wang, J.; Luo, M.; Wintergerst, K.A.; Miao, L.; Cai, L. Genetic variants of nuclear factor erythroid-derived 2-like 2 associated with the complications in Han descents with type 2 diabetes mellitus of Northeast China. J. Cell Mol. Med. 2016, 20, 2078–2088. [Google Scholar] [CrossRef] [Green Version]
- Arisawa, T.; Tahara, T.; Shibata, T.; Nagasaka, M.; Nakamura, M.; Kamiya, Y.; Fujita, H.; Yoshioka, D.; Okubo, M.; Sakata, M.; et al. Nrf2 gene promoter polymorphism is associated with ulcerative colitis in a Japanese population. Hepato-Gastroenterology 2008, 55, 394–397. [Google Scholar]
- von Otter, M.; Landgren, S.; Nilsson, S.; Celojevic, D.; Bergström, P.; Håkansson, A.; Nissbrandt, H.; Drozdzik, M.; Bialecka, M.; Kurzawski, M.; et al. Association of Nrf2-encoding NFE2L2 haplotypes with Parkinson’s disease. BMC Med. Genet. 2010, 11, 36. [Google Scholar] [CrossRef] [Green Version]
- von Otter, M.; Bergstrom, P.; Quattrone, A.; de Marco, E.V.; Annesi, G.; Söderkvist, P.; Wettinger, S.B.; Drozdzik, M.; Bialecka, M.; Nissbrandt, H.; et al. Genetic associations of Nrf2-encoding NFE2L2 variants with Parkinson disease—A multicenter study. BMC Med. Genet. 2014, 15, 131. [Google Scholar] [CrossRef] [Green Version]
- von Otter, M.; Landgren, S.; Nilsson, S.; Zetterberg, M.; Celojevic, D.; Bergström, P.; Minthon, L.; Bogdanovic, N.; Andreasen, N.; Gustafson, D.R.; et al. Nrf2-encoding NFE2L2 haplotypes influence disease progression but not risk in Alzheimer’s disease and age-related cataract. Mech. Ageing Dev. 2010, 131, 105–110. [Google Scholar] [CrossRef]
- Bergström, P.; von Otter, M.; Nilsson, S.; Nilsson, A.C.; Nilsson, M.; Andersen, P.M.; Hammarsten, O.; Zetterberg, H. Association of NFE2L2 and KEAP1 haplotypes with amyotrophic lateral sclerosis. Amyotroph. Lateral Scler. Front. Degener. 2014, 15, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.; Connors, S.L.; Macklin, E.A.; Smith, K.D.; Fahey, J.W.; Talalay, P.; Zimmerman, A.W. Sulforaphane treatment of autism spectrum disorder (ASD). Proc. Natl. Acad. Sci. USA 2014, 111, 15550–15555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campolo, M.; Casili, G.; Biundo, F.; Crupi, R.; Cordaro, M.; Cuzzocrea, S.; Esposito, E. The Neuroprotective Effect of Dimethyl Fumarate in an MPTP-Mouse Model of Parkinson’s Disease: Involvement of Reactive Oxygen Species/Nuclear Factor-κB/Nuclear Transcription Factor Related to NF-E. Antioxid. Redox Signal. 2017, 27, 453–471. [Google Scholar] [CrossRef] [Green Version]
- Scuderi, S.A.; Ardizzone, A.; Paterniti, I.; Esposito, E.; Campolo, M. Antioxidant and Anti-inflammatory Effect of Nrf2 Inducer Dimethyl Fumarate in Neurodegenerative Diseases. Antioxidants 2020, 9, 630. [Google Scholar] [CrossRef]
- Majkutewicz, I. Dimethyl fumarate: A review of preclinical efficacy in models of neurodegenerative diseases. Eur. J. Pharmacol. 2022, 926, 175025. [Google Scholar] [CrossRef]
- NCBI Website. Available online: http://www.ncbi.nlm.nih.gov/projects/SNP (accessed on 1 July 2021).
- Ran, C.; Wirdefeldt, K.; Brodin, L.; Ramezani, M.; Westerlund, M.; Xiang, F.; Anvret, A.; Willows, T.; Sydow, O.; Johansson, A.; et al. Genetic Variations and mRNA Expression of NRF2 in Parkinson’s Disease. Park. Dis. 2017, 2017, 4020198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimoyama, Y.; Mitsuda, Y.; Tsuruta, Y.; Hamajima, N.; Niwa, T. Polymorphism of Nrf2, an antioxidative gene, is associated with blood pressure and cardiovascular mortality in hemodialysis patients. Int. J. Med. Sci. 2014, 11, 726–731. [Google Scholar] [CrossRef] [Green Version]
- Korytina, G.F.; Akhmadishina, L.Z.; Aznabaeva, Y.G.; Kochetova, O.V.; Zagidullin, N.S.; Kzhyshkowska, J.G.; Zagidullin, S.Z.; Viktorova, T.V. Associations of the NRF2/KEAP1 pathway and antioxidant defense gene polymorphisms with chronic obstructive pulmonary disease. Gene 2019, 692, 102–112. [Google Scholar] [CrossRef]
- Marzec, J.M.; Christie, J.D.; Reddy, S.P.; Jedlicka, A.E.; Vuong, H.; Lanken, P.N.; Aplenc, R.; Yamamoto, T.; Yamamoto, M.; Cho, H.Y.; et al. Functional polymorphisms in the transcription factor NRF2 in humans increase the risk of acute lung injury. FASEB J. 2007, 21, 2237–2246. [Google Scholar] [CrossRef]
- Marczak, E.D.; Marzec, J.; Zeldin, D.C.; Kleeberger, S.R.; Brown, N.J.; Pretorius, M.; Lee, C.R. Polymorphisms in the transcription factor NRF2 and forearm vasodilator responses in humans. Pharm. Genom. 2012, 22, 620–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arisawa, T.; Tahara, T.; Shibata, T.; Nagasaka, M.; Nakamura, M.; Kamiya, Y.; Fujita, H.; Yoshioka, D.; Arima, Y.; Okubo, M.; et al. Association between promoter polymorphisms of nuclear factor-erythroid 2-related factor 2 gene and peptic ulcer diseases. Int. J. Mol. Med. 2007, 20, 849–853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, B.; Huang, Z. Variations in Antioxidant Genes and Male Infertility. Biomed. Res. Int. 2015, 2015, 513196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russia Infant Mortality Rate 1950-. Available online: https://www.macrotrends.net/countries/RUS/russia/infant-mortality-rate (accessed on 4 October 2022).
- Barbour, E.K.; Shaib, H.A.; Ahmadieh, D.M.; Kumosani, T.; Hamadeh, S.K.; Azhar, E.; Harakeh, S. A mini review of qRT-rtPCR technology application in uncovering the mechanism of food allergy and in the search for novel interventions. Antiinflamm. Antiallergy Agents Med. Chem. 2013, 12, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Ledderose, C.; Heyn, J.; Limbeck, E.; Kreth, S. Selection of reliable reference genes for quantitative real-time PCR in human T cells and neutrophils. BMC Res. Notes 2011, 4, 427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsaur, I.; Renninger, M.; Hennenlotter, J.; Oppermann, E.; Munz, M.; Kuehs, U.; Stenzl, A.; Schilling, D. Reliable housekeeping gene combination for quantitative PCR of lymph nodes in patients with prostate cancer. Anticancer Res. 2013, 33, 5243–5248. [Google Scholar]
- Invitrogen™ TRIzol™ Reagent User Guide. Available online: https://tools.thermofisher.com/content/sfs/manuals/trizol_reagent.pdf (accessed on 4 October 2022).
- Shmarina, G.V.; Ershova, E.S.; Simashkova, N.V.; Nikitina, S.G.; Chudakova, J.M.; Veiko, N.N.; Porokhovnik, L.N.; Basova, A.Y.; Shaposhnikova, A.F.; Pukhalskaya, D.A.; et al. Oxidized cell-free DNA as a stress-signaling factor activating the chronic inflammatory process in patients with autism spectrum disorders. J. Neuroinflamm. 2020, 17, 212. [Google Scholar] [CrossRef]
- Hammer, O.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Outlier Calculator. Available online: https://www.graphpad.com/quickcalcs/Grubbs1.cfm (accessed on 4 October 2022).
- Veĭko, N.N.; Bulycheva, N.V.; Roginko, O.A.; Veĭko, R.V.; Ershova, E.S.; Kozdoba, O.A.; Kuzmin, V.A.; Vinogradov, A.M.; Iudin, A.A.; Speranskiĭ, A.I. Ribosomal repeat in the cell free DNA as a marker for cell death. Biomed. Khim. 2008, 54, 78–93. [Google Scholar] [CrossRef]
- Korzeneva, I.B.; Kostyuk, S.V.; Ershova, L.S.; Osipov, A.N.; Zhuravleva, V.F.; Pankratova, G.V.; Porokhovnik, L.N.; Veiko, N.N. Human circulating plasma DNA significantly decreases while lymphocyte DNA damage increases under chronic occupational exposure to low-dose γ-neutron and tritium β-radiation. Mutat. Res. 2015, 779, 1–15. [Google Scholar] [CrossRef]
- SFARI Gene Database. Available online: https://gene.sfari.org/autdb/Welcome.do (accessed on 4 October 2022).
- Arisawa, T.; Tahara, T.; Shibata, T.; Nagasaka, M.; Nakamura, M.; Kamiya, Y.; Fujita, H.; Yoshioka, D.; Okubo, M.; Hirata, I.; et al. Nrf2 gene promoter polymorphism and gastric carcinogenesis. Hepato-Gastroenterology 2008, 55, 750–754. [Google Scholar] [PubMed]
- Arisawa, T.; Tahara, T.; Shibata, T.; Nagasaka, M.; Nakamura, M.; Kamiya, Y.; Fujita, H.; Yoshioka, D.; Arima, Y.; Okubo, M.; et al. The influence of promoter polymorphism of nuclear factor-erythroid 2-related factor 2 gene on the aberrant DNA methylation in gastric epithelium. Oncol. Rep. 2008, 19, 211–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.C.; Wu, Y.R.; Wu, Y.C.; Lee-Chen, G.J.; Chen, C.M. Genetic analysis of NFE2L2 promoter variation in Taiwanese Parkinson’s disease. Park. Relat. Disord. 2013, 19, 247–250. [Google Scholar] [CrossRef] [PubMed]
- Gruosso, T.; Mieulet, V.; Cardon, M.; Bourachot, B.; Kieffer, Y.; Devun, F.; Dubois, T.; Dutreix, M.; Vincent-Salomon, A.; Miller, K.M.; et al. Chronic oxidative stress promotes H2AX protein degradation and enhances chemosensitivity in breast cancer patients. EMBO Mol. Med. 2016, 8, 527–549. [Google Scholar] [CrossRef] [PubMed]
- Porokhovnik, L.N.; Passekov, V.P.; Gorbachevskaya, N.L.; Sorokin, A.B.; Veiko, N.N.; Lyapunova, N.A. Active Ribosomal Genes, Translational Homeostasis and Oxidative Stress in the Pathogenesis of Schizophrenia and Autism. Psychiatr. Genet. 2015, 25, 79–87. [Google Scholar] [CrossRef]
- Napoli, E.; Wong, S.; Hertz-Picciotto, I.; Giulivi, C. Deficits in bioenergetics and impaired immune response in granulocytes from children with autism. Pediatrics 2014, 133, e1405–e1410. [Google Scholar] [CrossRef] [Green Version]
- Singh, K.; Zimmerman, A.W. Sulforaphane treatment of young men with Autism Spectrum Disorder. In CNS & Neurological Disorders—Drug Targets; Bentham Science Publishers: Sharjah, United Arab Emirates, 2016; Volume 15, pp. 597–601. [Google Scholar]
- Momtazmanesh, S.; Amirimoghaddam-Yazdi, Z.; Moghaddam, H.S.; Mohammadi, M.R.; Akhondzadeh, S. Sulforaphane as an adjunctive treatment for irritability in Autism Spectrum Disorder: A randomized, double-blind, placebo-controlled clinical trial. Psychiatry Clin. Neurosci. 2020, 74, 398–405. [Google Scholar] [CrossRef]
- Nadeem, A.; Ahmad, S.F.; Al-Ayadhi, L.Y.; Attia, S.M.; Al-Harbi, N.O.; Alzahrani, K.S.; Bakheet, S.A. Differential regulation of Nrf2 is linked to elevated inflammation and nitrative stress in monocytes of children with autism. Psychoneuroendocrinology 2020, 113, 104554. [Google Scholar] [CrossRef]
- Ayhan, F.; Konopka, G. Regulatory genes and pathways disrupted in autism spectrum disorders. Prog. Neuropsychopharmacol. Biol. Psychiatry 2019, 89, 57–64. [Google Scholar] [CrossRef]


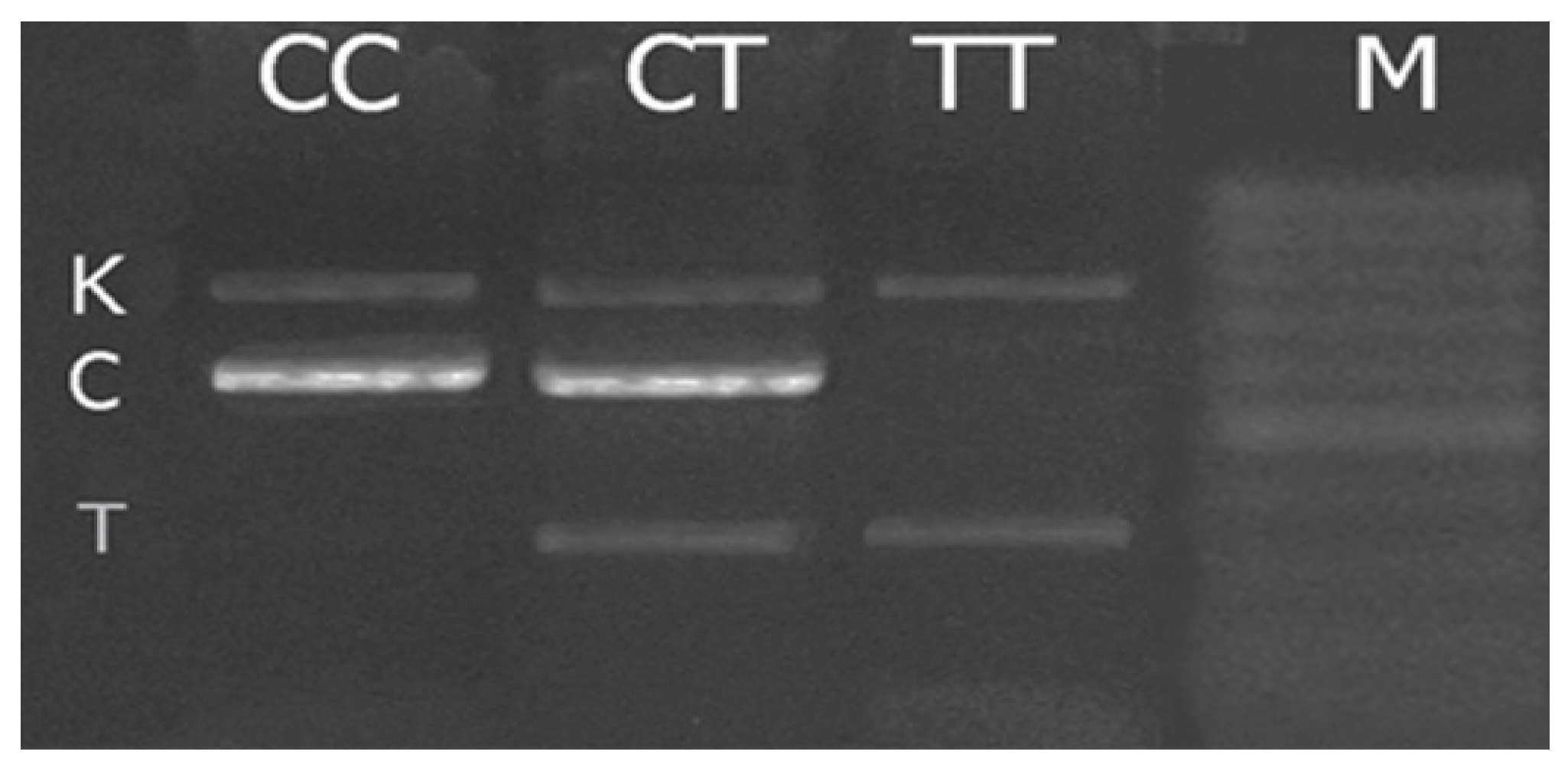{kind=link}
| Code | Chromosome 2 Map Position (GRCh38.106) * | Location | Clinical Significance (on the Base of Published Literature) | Population of Origin |
|---|---|---|---|---|
| rs35652124 | 177265345 T > C | Promotor -214 | Ulcerative colitis, gastritis, gastric ulcer | Japan [18,33] |
| Cardiovascular mortality in hemodialysis patients | Japan [29] | |||
| Arterial pressure, cardiovascular disease risk, and chronic obstructive pulmonary disease | Dutch [16] | |||
| Parkinson’s disease | Sweden and Poland—meta-analysis [19,20], European countries [28] | |||
| Oligoasthenozoospermia | China [15,34] |
| Genotype | rs35652124 | ||
| T/T | C/T | C/C | |
| Controls | 32 | 52 | 16 |
| Cases | 60 | 90 | 29 |
| Pcompar. (χ2) | p = 0.96 | ||
| Hardy-Weinberg, controls (χ2) | p = 0.501 | ||
| Hardy-Weinberg, cases (χ2) | p = 0.624 | ||
| cfDNA/Nuclease Activity | NRF2 | ph-NRF2 | Nrf2 DMF/Baseline | ph-Nrf2 DMF/Baseline | 8-oxo-G | mRNA Nrf2 DMF/Baseline | mRNA NQO1 DMF/Baseline | H2AX | |
| CC | 124.78 (92.95; 186.33) | 2.055 (1.37; 2.79) | 0.18 (0.14; 0.25) | 0.98 (0.95; 1.49) | 1.923 (1.653; 2.26) | 0.66 (0.57; 0.76) | 1.034 (0.63; 1.27) | 0.924 (0.88; 1.07) | 0.652 (0.51; 1.13) |
| CT | 61.687 (24.64; 91.11) | 1.832 (1.05; 2.74) | 0.149 (0.08; 0.39) | 1.389 (1.14; 1.54) | 0.853 (0.46; 1.47) | 1.08 (0.88; 1.20) | 0.789 (0.73; 1.13) | 1.107 (0.75; 1.28) | 0.499 (0.37; 0.64) |
| TT | 44.928 (29.38; 66.69) | 2.72 (1.52; 3.17) | 0.269 (0.14; 0.46) | 1.196 (0.95; 1.48) | 1.274 (0.69; 1.93) | 1.18 (0.99; 1.51) | 1.211 (0.88; 2.02) | 1.034 (0.91; 1.44) | 0.515 (0.47; 0.65) |
| C+ | 65.667 (35.97; 117.26) | 1.88 (1.16; 2.79) | 0.167 (0.09; 0.36) | 1.349 (0.97; 1.53) | 1.558 (0.85; 1.92) | 1.01 (0.78; 1.15) | 0.95 (0.72; 1.16) | 0.97 (0.87; 1.16) | 0.52 (0.41; 0.71) |
| T+ | 52.14 (26.08; 85.90) | 1.99 (1.42; 3.11) | 0.21 (0.09; 0.44) | 1.35 (0.97; 1.51) | 1.196 (0.68; 1.56) | 1.12 (0.91; 1.34) | 1.052 (0.75; 1.33) | 1.058 (0.88; 1.33) | 0.505 (0.41; 0.65) |
| CC vs. T+ | p = 0.022 * | p = 0.860 | p = 0.894 | p = 0.703 | p = 0.049 * | p = 0.018 * | p = 0.664 | p = 0.270 | p = 0.048 * |
| TT vs. C+ | p = 0.317 | p = 0.128 | p = 0.235 | p = 0.364 | p = 0.792 | p = 0.107 | p = 0.156 | p = 0.395 | p = 0.861 |
| Indices | Parameters | rs35652124 Genotype | |
| CC | CT + TT | ||
| NRF2 vs. H2AX | R | −0.075 | 0.479 * |
| P | 0.809 | 0.0058 | |
| N | 9 | 53 | |
| 8-oxo-G vs. pH2AX | R | 0.24 | 0.169 |
| P | 0.399 | 0.662 | |
| N | 14 | 8 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Porokhovnik, L.N.; Pisarev, V.M.; Chumachenko, A.G.; Chudakova, J.M.; Ershova, E.S.; Veiko, N.N.; Gorbachevskaya, N.L.; Mamokhina, U.A.; Sorokin, A.B.; Basova, A.Y.; et al. Association of NEF2L2 Rs35652124 Polymorphism with Nrf2 Induction and Genotoxic Stress Biomarkers in Autism. Genes 2023, 14, 718. https://doi.org/10.3390/genes14030718
Porokhovnik LN, Pisarev VM, Chumachenko AG, Chudakova JM, Ershova ES, Veiko NN, Gorbachevskaya NL, Mamokhina UA, Sorokin AB, Basova AY, et al. Association of NEF2L2 Rs35652124 Polymorphism with Nrf2 Induction and Genotoxic Stress Biomarkers in Autism. Genes. 2023; 14(3):718. https://doi.org/10.3390/genes14030718
Chicago/Turabian StylePorokhovnik, Lev N., Vladimir M. Pisarev, Anastasia G. Chumachenko, Julia M. Chudakova, Elizaveta S. Ershova, Natalia N. Veiko, Natalia L. Gorbachevskaya, Uliana A. Mamokhina, Alexander B. Sorokin, Anna Ya. Basova, and et al. 2023. "Association of NEF2L2 Rs35652124 Polymorphism with Nrf2 Induction and Genotoxic Stress Biomarkers in Autism" Genes 14, no. 3: 718. https://doi.org/10.3390/genes14030718