
Mutation Rate and Spectrum of the Silkworm in Normal and Temperature Stress Conditions

 , , and
, , and
Abstract
:- Significance Statement:
1. Introduction
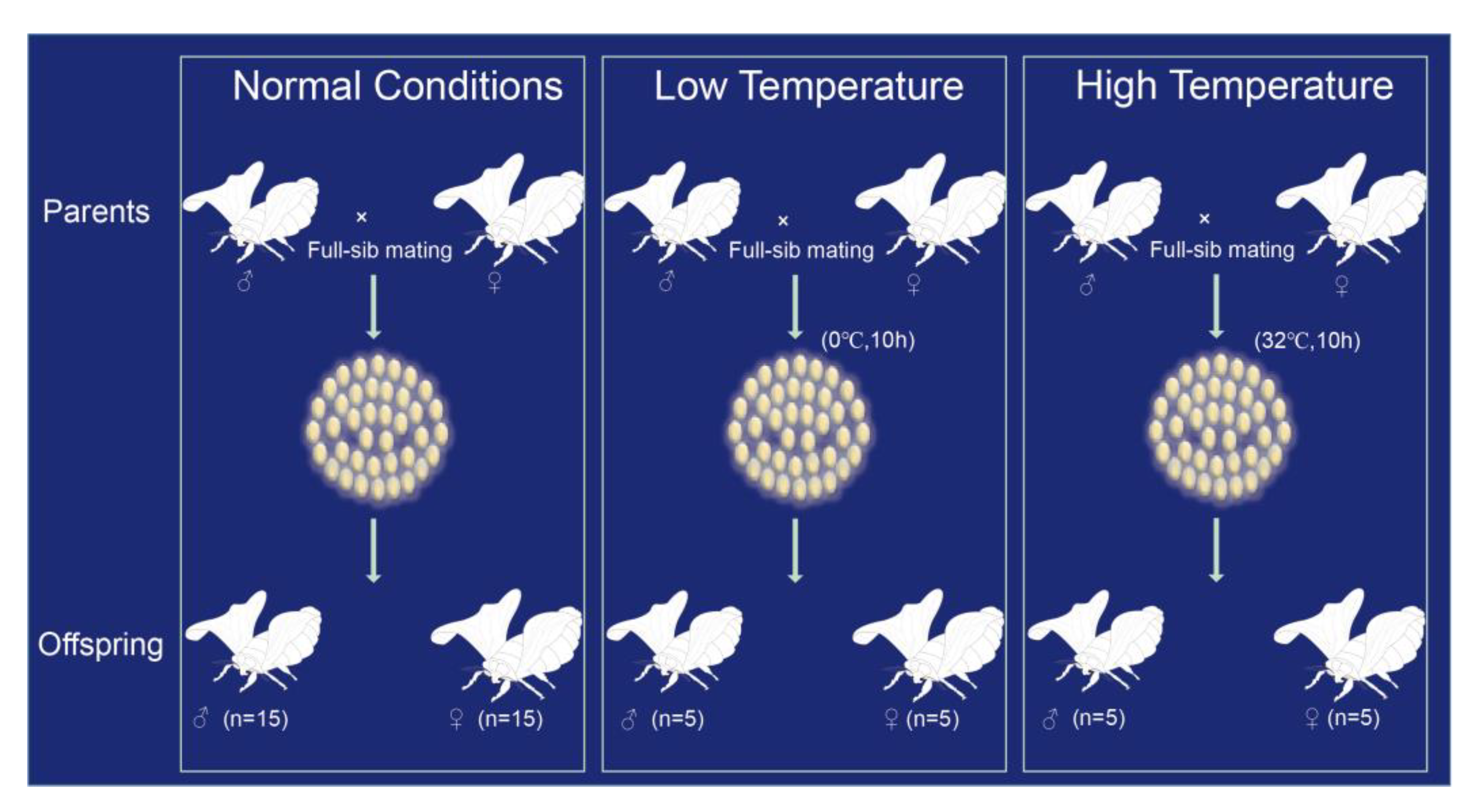
2. Materials and Methods
2.1. Sample Source and DNA Extraction
2.2. Genome Sequencing
2.3. Identification of the Mutations
2.4. Estimation of False Positives
2.5. Estimation of False Negatives
2.6. Statistical Analysis and Silkworm Domestication Time Estimation
2.7. Data Access
3. Results
3.1. Identification of the de Novo Mutations
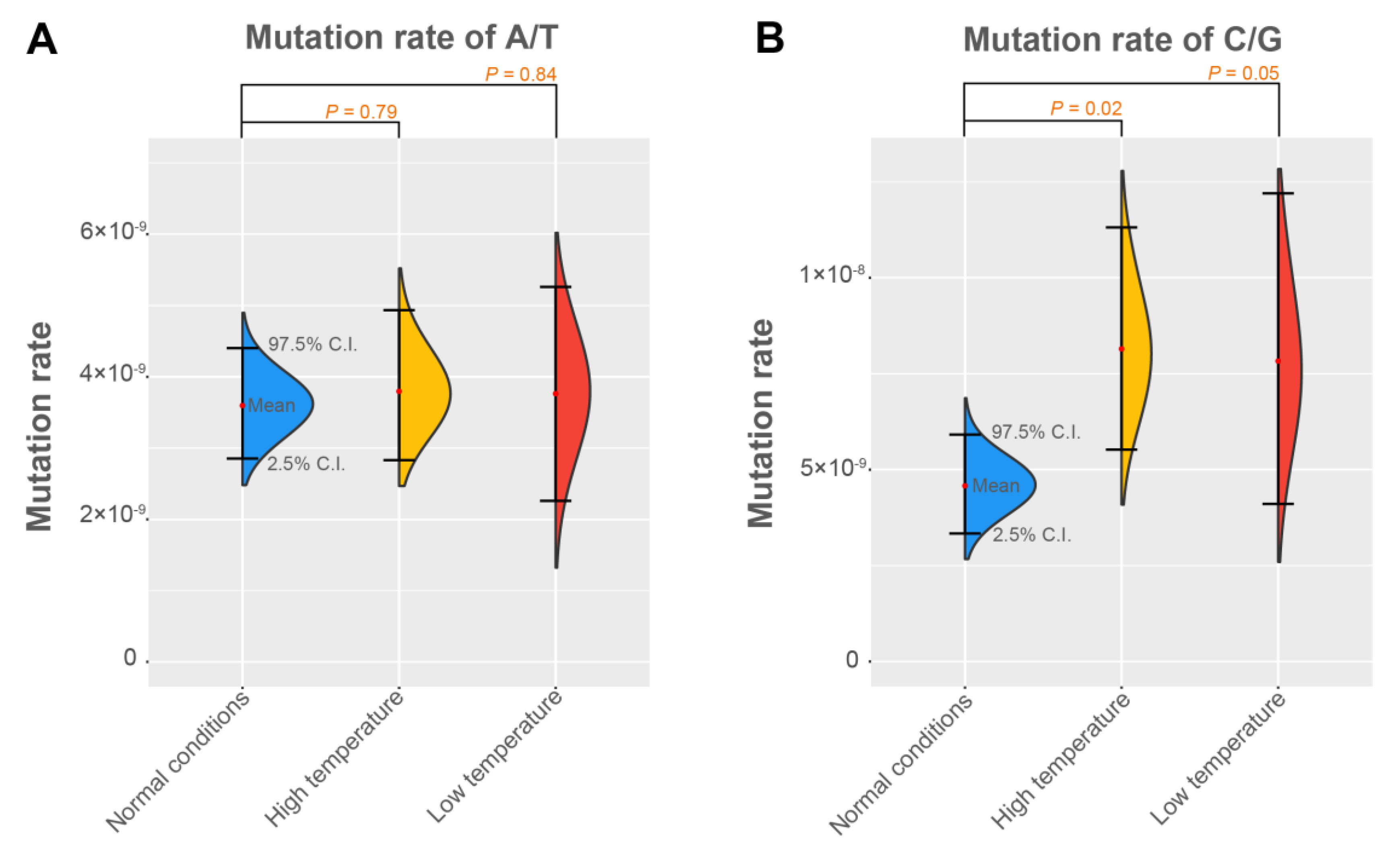
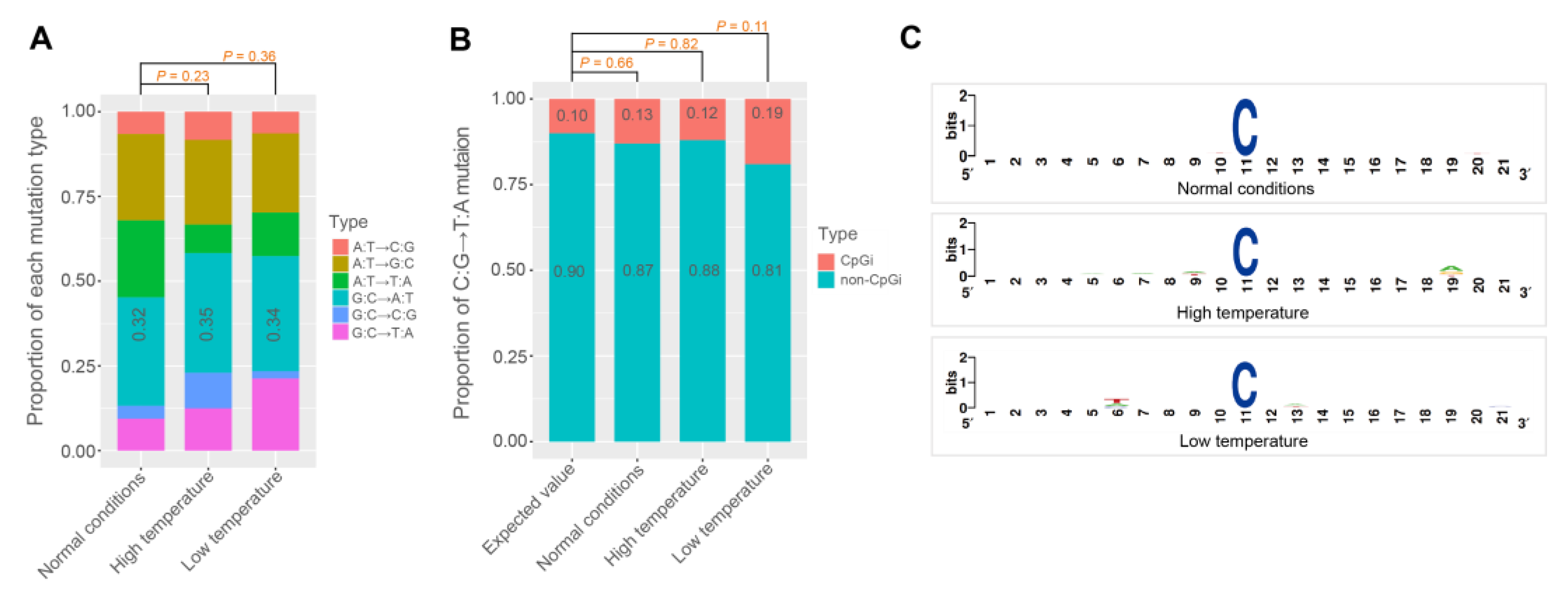
3.2. The Base Mutation Bias
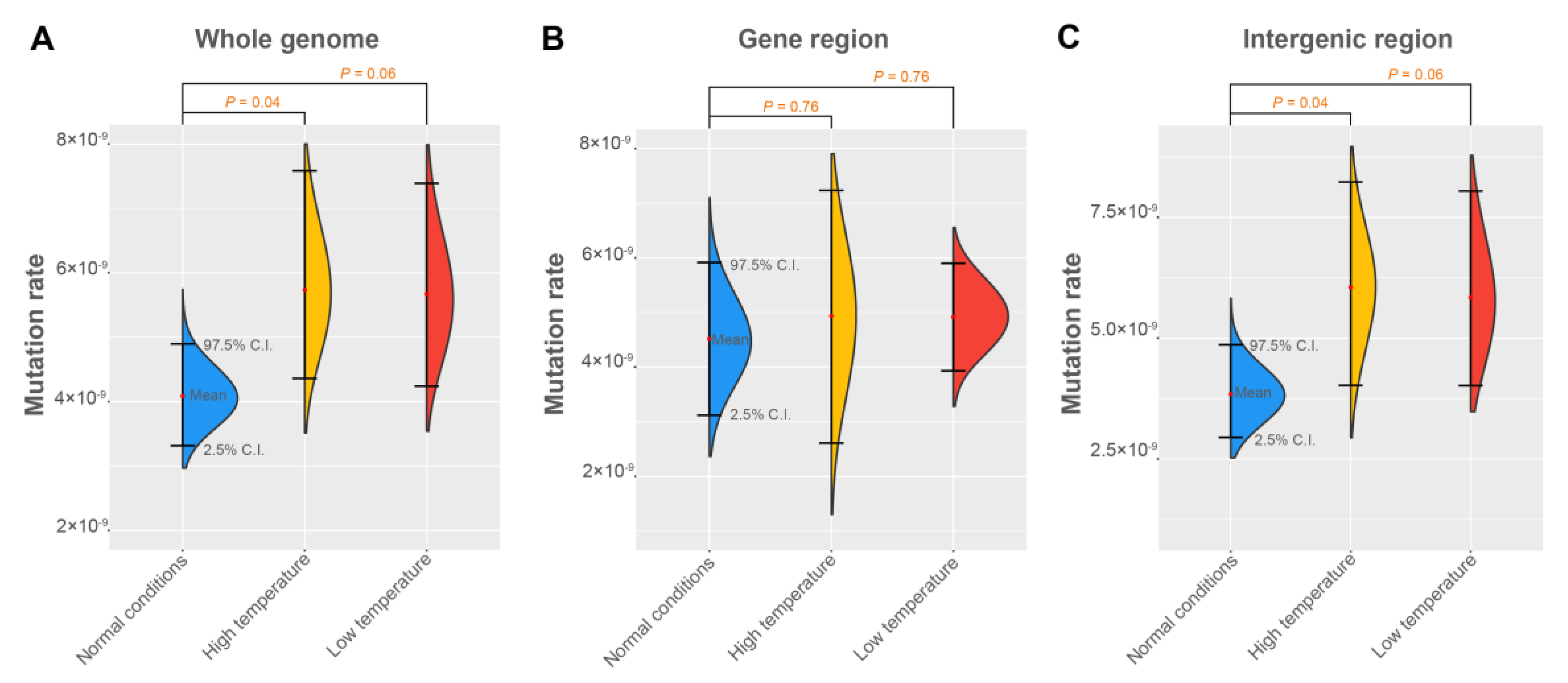
3.3. The Mutation Rates under Different Environments
4. Discussion
4.1. The Mutation Rate Varies in Different Species
4.2. The Mutation Rate Varies in Different Environments
4.3. The Mutation Rate Is Constant in Gene Regions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lynch, M. Evolution of the mutation rate. Trends Genet. 2010, 26, 345–352. [Google Scholar] [CrossRef] [Green Version]
- Maharjan, R.P.; Ferenci, T. Escherichia coli mutation rates and spectra with combinations of environmental limitations. Microbiology 2018, 164, 1491–1502. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, J. Yeast spontaneous mutation rate and spectrum vary with environment. Curr. Biol. 2019, 29, 1584–1591. [Google Scholar] [CrossRef]
- Denver, D.R.; Wilhelm, L.J.; Howe, D.K.; Gafner, K.; Dolan, P.C.; Baer, C.F. Variation in base-substitution mutation in experimental and natural lineages of Caenorhabditis nematodes. Genome Biol. Evol. 2012, 4, 513–522. [Google Scholar] [CrossRef] [Green Version]
- Konrad, A.; Brady, M.J.; Bergthorsson, U.; Katju, V. Mutational landscape of spontaneous base substitutions and small indels in experimental Caenorhabditis elegans populations of differing size. Genetics 2019, 212, 837–854. [Google Scholar] [CrossRef] [PubMed]
- Saxena, A.S.; Salomon, M.P.; Matsuba, C.; Yeh, S.D.; Baer, C.F. Evolution of the mutational process under relaxed selection in Caenorhabditis elegans. Mol. Biol. Evol. 2019, 36, 239–251. [Google Scholar] [CrossRef] [PubMed]
- Keightley, P.D.; Ness, R.W.; Halligan, D.L.; Haddrill, P.R. Estimation of the spontaneous mutation rate per nucleotide site in a Drosophila melanogaster full-sib family. Genetics 2014, 196, 313–320. [Google Scholar] [CrossRef] [Green Version]
- Uchimura, A.; Higuchi, M.; Minakuchi, Y.; Ohno, M.; Toyoda, A.; Fujiyama, A.; Miura, I.; Wakana, S.; Nishino, J.; Yagi, T. Germline mutation rates and the long-term phenotypic effects of mutation accumulation in wild-type laboratory mice and mutator mice. Genome Res. 2015, 25, 1125–1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conrad, D.F.; Keebler, J.E.M.; DePristo, M.A.; Lindsay, S.J.; Zhang, Y.J.; Casals, F.; Idaghdour, Y.; Hartl, C.L.; Torroja, C.; Garimella, K.V.; et al. Variation in genome-wide mutation rates within and between human families. Nat. Genet. 2011, 43, 712–714. [Google Scholar]
- Xia, Q.Y.; Guo, Y.R.; Zhang, Z.; Li, D.; Xuan, Z.L.; Li, Z.; Dai, F.Y.; Li, Y.R.; Cheng, D.J.; Li, R.Q.; et al. Complete resequencing of 40 genomes reveals domestication events and genes in silkworm (Bombyx). Science 2009, 326, 433–436. [Google Scholar] [CrossRef] [Green Version]
- Sun, W.; Yu, H.; Shen, Y.; Banno, Y.; Xiang, Z.; Zhang, Z. Phylogeny and evolutionary history of the silkworm. Sci. China Life Sci. 2012, 55, 483–496. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.Y.; Han, M.J.; Kang, L.F.; Li, Z.W.; Shen, Y.H.; Zhang, Z. Demographic history and gene flow during silkworm domestication. BMC Evol. Biol. 2014, 14, 185. [Google Scholar] [CrossRef] [Green Version]
- Xiang, H.; Liu, X.J.; Li, M.W.; Zhu, Y.N.; Wang, L.Z.; Cui, Y.; Liu, L.Y.; Fang, G.Q.; Qian, H.Y.; Xu, A.Y.; et al. The evolutionary road from wild moth to domestic silkworm. Nat. Ecol. Evol. 2018, 2, 1268–1279. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.B.; Zhang, R.S.; Bian, H.X.; Li, Q.; Xia, R.X.; Li, Y.P.; Liu, Y.Q.; Lu, C. Comparative mitochondrial genomes provide new insights into the true wild progenitor and origin of domestic silkworm Bombyx mori. Int. J. Biol. Macromol. 2019, 131, 176–183. [Google Scholar] [CrossRef]
- Li, W.H. Molecular Evolution; Sinauer Associates: Sunderland, MA, USA, 1997. [Google Scholar]
- Keightley, P.D.; Pinharanda, A.; Ness, R.W.; Simpson, F.; Dasmahapatra, K.K.; Mallet, J.; Davey, J.W.; Jiggins, C.D. Estimation of the spontaneous mutation rate in Heliconius melpomene. Mol. Biol. Evol. 2015, 32, 239–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segurel, L.; Wyman, M.J.; Przeworski, M. Determinants of mutation rate variation in the human germline. Annu. Rev. Genom. Hum. Genet. 2014, 15, 47–70. [Google Scholar] [CrossRef]
- Besenbacher, S.; Liu, S.Y.; Izarzugaza, J.M.G.; Grove, J.; Belling, K.; Bork-Jensen, J.; Huang, S.; Als, T.D.; Li, S.; Yadav, R.; et al. Novel variation and de novo mutation rates in population-wide de novo assembled Danish trios. Nat. Commun. 2015, 6, 5969. [Google Scholar] [CrossRef] [Green Version]
- Lynch, M. Mutation and human exceptionalism: Our future genetic load. Genetics 2016, 202, 869–875. [Google Scholar] [CrossRef] [PubMed]
- Rahbari, R.; Wuster, A.; Lindsay, S.J.; Hardwick, R.J.; Alexandrov, L.B.; Turki, S.A.; Dominiczak, A.; Morris, A.; Porteous, D.; Smith, B.; et al. Timing, rates and spectra of human germline mutation. Nat. Genet. 2016, 48, 126–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, W.S.W.; Solomon, B.D.; Bodian, D.L.; Kothiyal, P.; Eley, G.; Huddleston, K.C.; Baker, R.; Thach, D.C.; Iyer, R.K.; Vockley, J.G.; et al. New observations on maternal age effect on germline de novo mutations. Nat. Commun. 2016, 7, 10486. [Google Scholar] [CrossRef] [Green Version]
- Jonsson, H.; Sulem, P.; Kehr, B.; Kristmundsdottir, S.; Zink, F.; Hjartarson, E.; Hardarson, M.T.; Hjorleifsson, K.E.; Eggertsson, H.P.; Gudjonsson, S.A.; et al. Parental influence on human germline de novo mutations in 1548 trios from Iceland. Nature 2017, 549, 519–522. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Jia, Y.; Sun, X.; Tian, D.; Hurst, L.D.; Yang, S. Direct determination of the mutation rate in the Bumblebee reveals evidence for weak recombination-associated mutation and an approximate rate constancy in insects. Mol. Biol. Evol. 2017, 34, 119–130. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Wang, L.; Huang, J.; Zhang, X.; Yuan, Y.; Chen, J.Q.; Hurst, L.D.; Tian, D.C. Parent-progeny sequencing indicates higher mutation rates in heterozygotes. Nature 2015, 523, 463–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yosef, I.; Edgar, R.; Levy, A.; Amitai, G.; Sorek, R.; Munitz, A.; Qimron, U. Natural selection underlies apparent stress-induced mutagenesis in a bacteriophage infection model. Nat. Microbiol. 2016, 1, 16047. [Google Scholar] [CrossRef]
- Jiang, C.; Mithani, A.; Belfield, E.J.; Mott, R.; Hurst, L.D.; Harberd, N.P. Environmentally responsive genome-wide accumulation of de novo Arabidopsis thaliana mutations and epimutations. Genome Res. 2014, 24, 1821–1829. [Google Scholar] [CrossRef] [Green Version]
- Long, H.; Miller, S.F.; Strauss, C.; Zhao, C.; Cheng, L.; Ye, Z.; Griffin, K.; Te, R.; Lee, H.; Chen, C.C.; et al. Antibiotic treatment enhances the genome-wide mutation rate of target cells. Proc. Natl. Acad. Sci. USA 2016, 113, E2498–E2505. [Google Scholar] [CrossRef] [Green Version]
- Williams, A.B.; Foster, P.L. Stress-Induced Mutagenesis. EcoSal Plus 2012, 5, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maharjan, R.P.; Ferenci, T. A shifting mutational landscape in 6 nutritional states: Stress-induced mutagenesis as a series of distinct stress input-mutation output relationships. PLoS Biol. 2017, 15, e2001477. [Google Scholar] [CrossRef] [Green Version]
- Shewaramani, S.; Finn, T.J.; Leahy, S.C.; Kassen, R.; Rainey, P.B.; Moon, C.D. Anaerobically grown Escherichia coli has an enhanced mutation rate and distinct mutational spectra. PLoS Genet. 2017, 13, e1006570. [Google Scholar] [CrossRef] [Green Version]
- Chu, X.L.; Zhang, B.W.; Zhang, Q.G.; Zhu, B.R.; Lin, K.; Zhang, D.Y. Temperature responses of mutation rate and mutational spectrum in an Escherichia coli strain and the correlation with metabolic rate. BMC Evol. Biol. 2018, 18, 126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shor, E.; Fox, C.A.; Broach, J.R. The yeast environmental stress response regulates mutagenesis induced by proteotoxic stress. PLoS Genet. 2013, 9, e1003680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agrawal, A.F.; Wang, A.D. Increased transmission of mutations by low-condition females: Evidence for condition-dependent DNA repair. PLoS Biol. 2008, 6, e30. [Google Scholar] [CrossRef] [Green Version]
- Sharp, N.P.; Agrawal, A.F. Evidence for elevated mutation rates in low-quality genotypes. Proc. Natl. Acad. Sci. USA 2012, 109, 6142–6146. [Google Scholar] [CrossRef] [Green Version]
- Wang, A.D.; Agrawal, A.F. DNA repair pathway choice is influenced by the health of Drosophila melanogaster. Genetics 2012, 192, 361–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Z.; Cui, J.; Wang, L.; Teng, N.; Zhang, S.; Lam, H.M.; Zhu, Y.; Xiao, S.; Ke, W.; Lin, J.; et al. Genome-wide DNA mutations in Arabidopsis plants after multigenerational exposure to high temperatures. Genome Biol. 2021, 22, 160. [Google Scholar] [CrossRef]
- Belfield, E.J.; Brown, C.; Ding, Z.J.; Chapman, L.; Luo, M.; Hinde, E.; van Es, S.W.; Johnson, S.; Ning, Y.; Zheng, S.J.; et al. Thermal stress accelerates Arabidopsis thaliana mutation rate. Genome Res. 2021, 31, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Xia, Q.Y.; Zhou, Z.Y.; Lu, C.; Cheng, D.J.; Dai, F.Y.; Li, B.; Zhao, P.; Zha, X.F.; Cheng, T.C.; Chai, C.L.; et al. A draft sequence for the genome of the domesticated silkworm (Bombyx mori). Science 2004, 306, 1937–1940. [Google Scholar]
- Kawamoto, M.; Jouraku, A.; Toyoda, A.; Yokoi, K.; Minakuchi, Y.; Katsuma, S.; Fujiyama, A.; Kiuchi, T.; Yamamoto, K.; Shimada, T. High-quality genome assembly of the silkworm, Bombyx mori. Insect Biochem. Mol. Biol. 2019, 107, 53–62. [Google Scholar] [CrossRef]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [Green Version]
- DePristo, M.A.; Banks, E.; Poplin, R.; Garimella, K.V.; Maguire, J.R.; Hartl, C.; Philippakis, A.A.; del Angel, G.; Rivas, M.A.; Hanna, M.; et al. A framework for variation discovery and genotyping using next-generation DNA sequencing data. Nat. Genet. 2011, 43, 491–498. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; 1000 Genome Project Data Processing Subgroup. The sequence alignment/map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Thorvaldsdottir, H.; Robinson, J.T.; Mesirov, J.P. Integrative Genomics Viewer (IGV): High-performance genomics data visualization and exploration. Brief. Bioinform. 2013, 14, 178–192. [Google Scholar] [CrossRef] [Green Version]
- Colot, V.; Rossignol, J.L. Eukaryotic DNA methylation as an evolutionary device. Bioessays 1999, 21, 402–411. [Google Scholar] [CrossRef]
- Zhao, Z.; Boerwinkle, E. Neighboring-nucleotide effects on single nucleotide polymorphisms: A study of 2.6 million polymorphisms across the human genome. Genome Res. 2002, 12, 1679–1686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, F.; Zhao, Z. The influence of neighboring-nucleotide composition on single nucleotide polymorphisms (SNPs) in the mouse genome and its comparison with human SNPs. Genomics 2004, 84, 785–795. [Google Scholar] [CrossRef] [PubMed]
- Keller, I.; Bensasson, D.; Nichols, R.A. Transition-transversion bias is not universal: A counter example from grasshopper pseudogenes. PLoS Genet. 2007, 3, e22. [Google Scholar] [CrossRef]
- Xiang, H.; Zhu, J.D.; Chen, Q.A.; Dai, F.Y.; Li, X.; Li, M.W.; Zhang, H.Y.; Zhang, G.J.; Li, D.; Dong, Y.; et al. Single base-resolution methylome of the silkworm reveals a sparse epigenomic map. Nat. Biotechnol. 2010, 28, 756. [Google Scholar] [CrossRef] [Green Version]
- Keith, N.; Tucker, A.E.; Jackson, C.E.; Sung, W.; Lucas Lledo, J.I.; Schrider, D.R.; Schaack, S.; Dudycha, J.L.; Ackerman, M.; Younge, A.J.; et al. High mutational rates of large-scale duplication and deletion in Daphnia pulex. Genome Res. 2016, 26, 60–69. [Google Scholar] [CrossRef] [Green Version]
- Koch, E.; Schweizer, R.M.; Schweizer, T.M.; Stahler, D.R.; Smith, D.W.; Wayne, R.K.; Novembre, J. De novo mutation rate estimation in wolves of known pedigree. Mol. Biol. Evol. 2019, 36, 2536–2547. [Google Scholar] [CrossRef] [Green Version]
- Smeds, L.; Qvarnstrom, A.; Ellegren, H. Direct estimate of the rate of germline mutation in a bird. Genome Res. 2016, 26, 1211–1218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamatoyannopoulos, J.A.; Adzhubei, I.; Thurman, R.E.; Kryukov, G.V.; Mirkin, S.M.; Sunyaev, S.R. Human mutation rate associated with DNA replication timing. Nat. Genet. 2009, 41, 393–395. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.Y.; Wyman, M.J.; Sella, G.; Przeworski, M. Interpreting the dependence of mutation rates on age and time. PLoS Biol. 2016, 14, e1002355. [Google Scholar] [CrossRef]
- Rahmathulla, V.K. Management of Climatic Factors for Successful Silkworm (Bombyx mori L.) Crop and Higher Silk Production: A Review. Psyche 2012, 2012, 121234. [Google Scholar]
- Rhind, N.; Gilbert, D.M. DNA Replication Timing. Cold Spring Harb. Perspect. Biol. 2013, 5, a010132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sogo, J.M.; Lopes, M.; Foiani, M. Fork reversal and ssDNA accumulation at stalled replication forks owing to checkpoint defects. Science 2002, 297, 599–602. [Google Scholar] [CrossRef]
- Mirkin, E.V.; Mirkin, S.M. Replication fork stalling at natural impediments. Microbiol. Mol. Biol. Rev. 2007, 71, 13–35. [Google Scholar] [CrossRef] [Green Version]
- Zeman, M.K.; Cimprich, K.A. Causes and consequences of replication stress. Nat. Cell Biol. 2014, 16, 2–9. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Regions and Specific Bases | Treatments | Mutation Rate (×10−9) | ||
|---|---|---|---|---|
| Mean | 2.5% C.I. | 97.5% C.I. | ||
| Whole genome | Normal conditions | 4.09 | 3.31 | 4.90 |
| High temperature | 5.80 | 4.35 | 7.44 | |
| Low temperature | 5.68 | 4.13 | 7.38 | |
| Gene region | Normal conditions | 4.52 | 3.23 | 5.92 |
| High temperature | 4.95 | 2.65 | 7.23 | |
| Low temperature | 4.92 | 3.94 | 5.90 | |
| Intergenic region | Normal conditions | 3.84 | 2.94 | 4.81 |
| High temperature | 6.05 | 4.04 | 8.06 | |
| Low temperature | 5.85 | 4.01 | 7.87 | |
| A/T | Normal conditions | 3.60 | 2.91 | 4.34 |
| High temperature | 3.80 | 3.20 | 4.44 | |
| Low temperature | 3.76 | 2.88 | 4.69 | |
| C/G | Normal conditions | 4.58 | 3.34 | 6.01 |
| High temperature | 8.14 | 6.41 | 9.87 | |
| Low temperature | 7.85 | 5.81 | 10.1 | |
| Treatments | Mutation Types | Mutation Rate (×10−9) | ||
|---|---|---|---|---|
| Mean | 2.5% C.I. | 97.5% C.I. | ||
| Normal conditions | A:T→C:G | 0.43 | 0.12 | 0.81 |
| A:T→G:C | 1.67 | 1.05 | 2.42 | |
| A:T→T:A | 1.49 | 0.99 | 1.98 | |
| G:C→A:T | 3.24 | 2.19 | 4.39 | |
| G:C→C:G | 0.38 | 0.00 | 0.86 | |
| G:C→T:A | 0.95 | 0.38 | 1.53 | |
| High temperature | A:T→C:G | 0.76 | 0.38 | 1.20 |
| A:T→G:C | 2.28 | 1.77 | 2.79 | |
| A:T→T:A | 0.76 | 0.44 | 1.08 | |
| G:C→A:T | 4.95 | 3.59 | 6.51 | |
| G:C→C:G | 1.45 | 0.77 | 2.13 | |
| G:C→T:A | 1.75 | 0.88 | 2.53 | |
| Low temperature | A:T→C:G | 0.56 | 0.25 | 0.88 |
| A:T→G:C | 2.07 | 1.44 | 2.75 | |
| A:T→T:A | 1.13 | 0.63 | 1.69 | |
| G:C→A:T | 4.63 | 3.18 | 6.27 | |
| G:C→C:G | 0.29 | 0.00 | 0.58 | |
| G:C→T:A | 2.93 | 1.75 | 4.19 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, M.; Ren, J.; Guo, H.; Tong, X.; Hu, H.; Lu, K.; Dai, Z.; Dai, F. Mutation Rate and Spectrum of the Silkworm in Normal and Temperature Stress Conditions. Genes 2023, 14, 649. https://doi.org/10.3390/genes14030649
Han M, Ren J, Guo H, Tong X, Hu H, Lu K, Dai Z, Dai F. Mutation Rate and Spectrum of the Silkworm in Normal and Temperature Stress Conditions. Genes. 2023; 14(3):649. https://doi.org/10.3390/genes14030649
Chicago/Turabian StyleHan, Minjin, Jianyu Ren, Haipeng Guo, Xiaoling Tong, Hai Hu, Kunpeng Lu, Zongrui Dai, and Fangyin Dai. 2023. "Mutation Rate and Spectrum of the Silkworm in Normal and Temperature Stress Conditions" Genes 14, no. 3: 649. https://doi.org/10.3390/genes14030649