

Genome-Wide Association Study for Grain Protein, Thousand Kernel Weight, and Normalized Difference Vegetation Index in Bread Wheat (Triticum aestivum L.)
 ,
,  , ,
, ,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Genotypes and Field Experiments
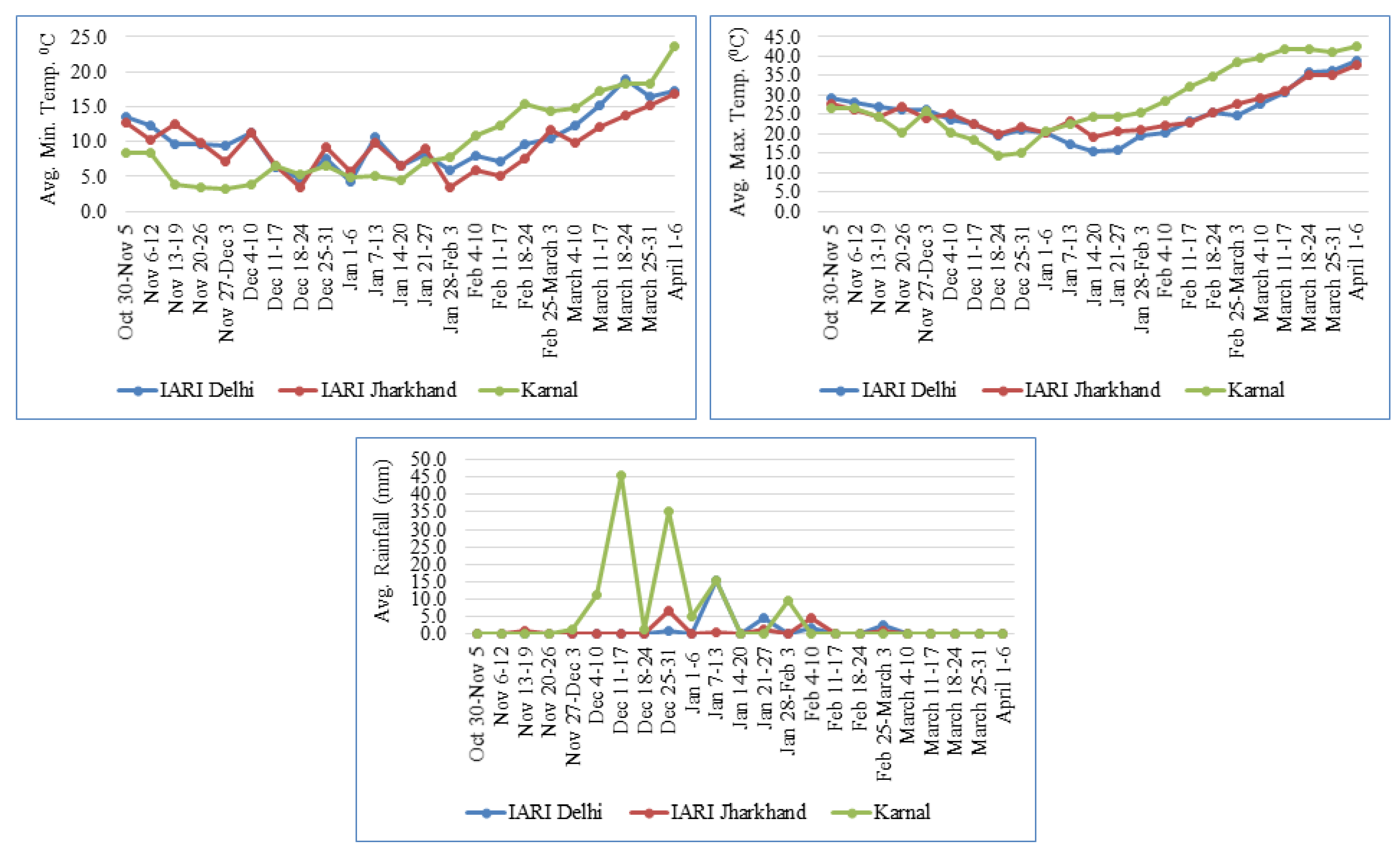
2.2. Data Recording and Statistical Analysis
2.3. Genotyping and Quality Control (QC)
2.4. Population Statistics and GWAS
2.5. In Silico Analysis
3. Results
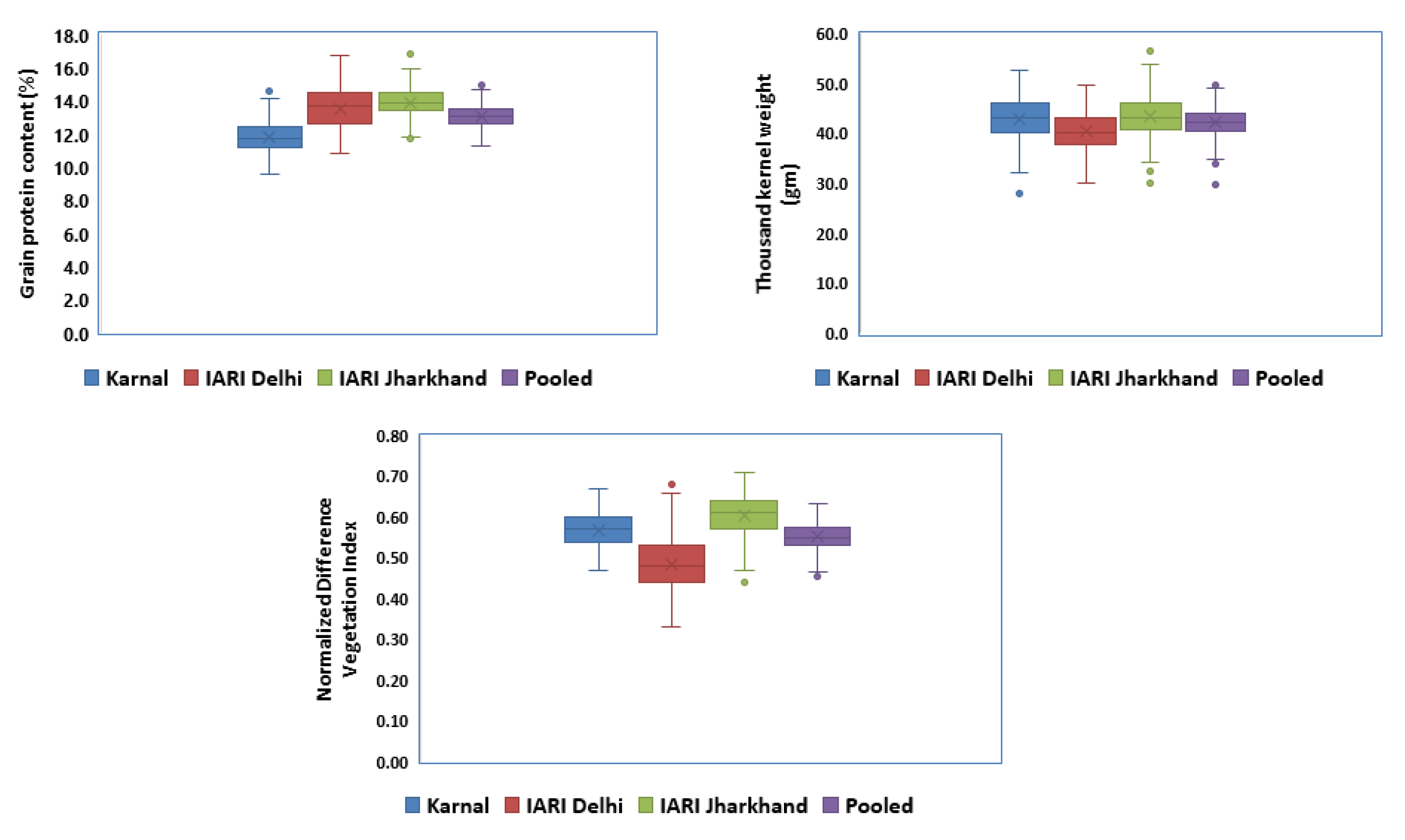
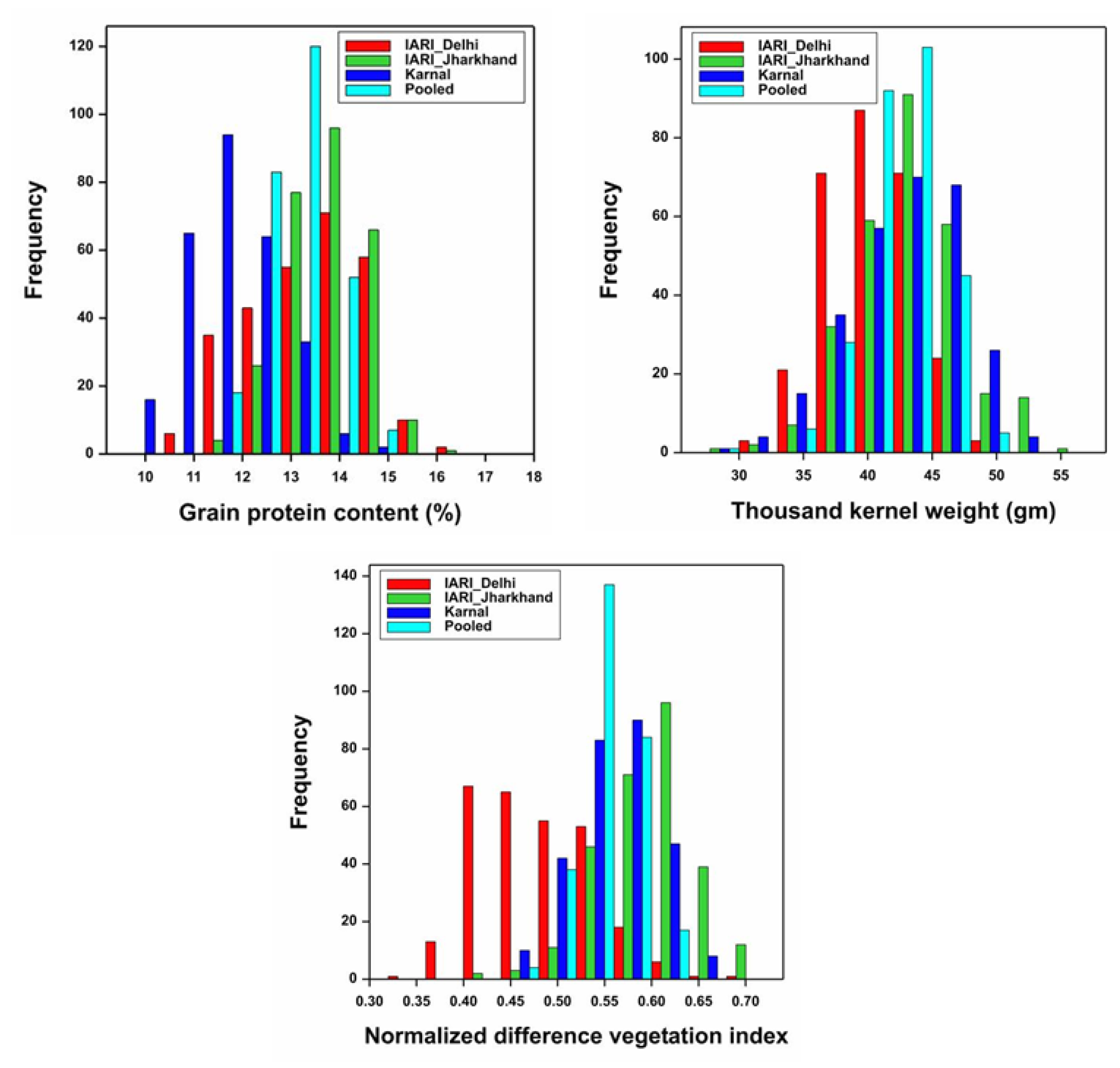
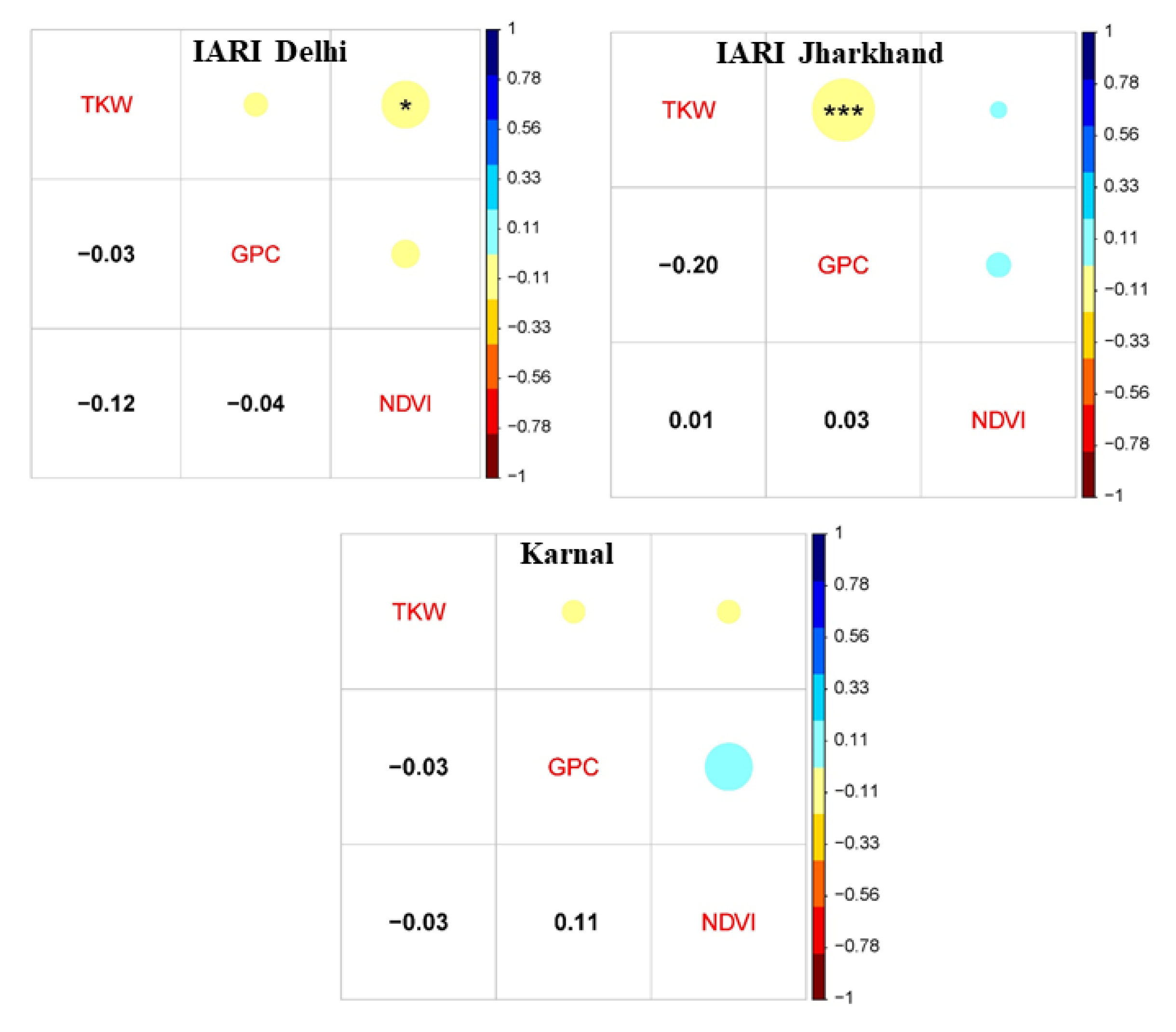
3.1. Variability, Heritability, and Correlation
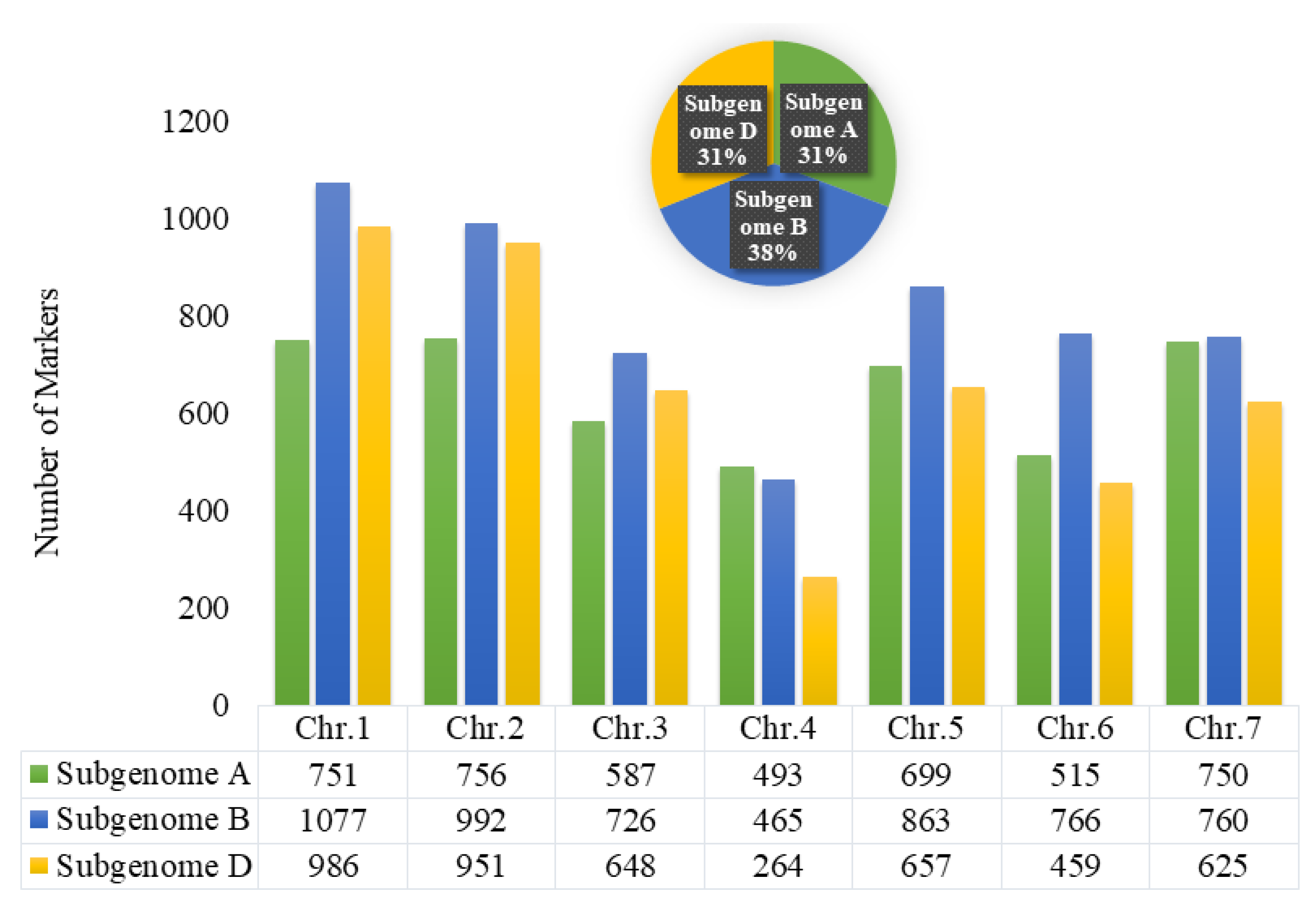
3.2. Marker Statistics
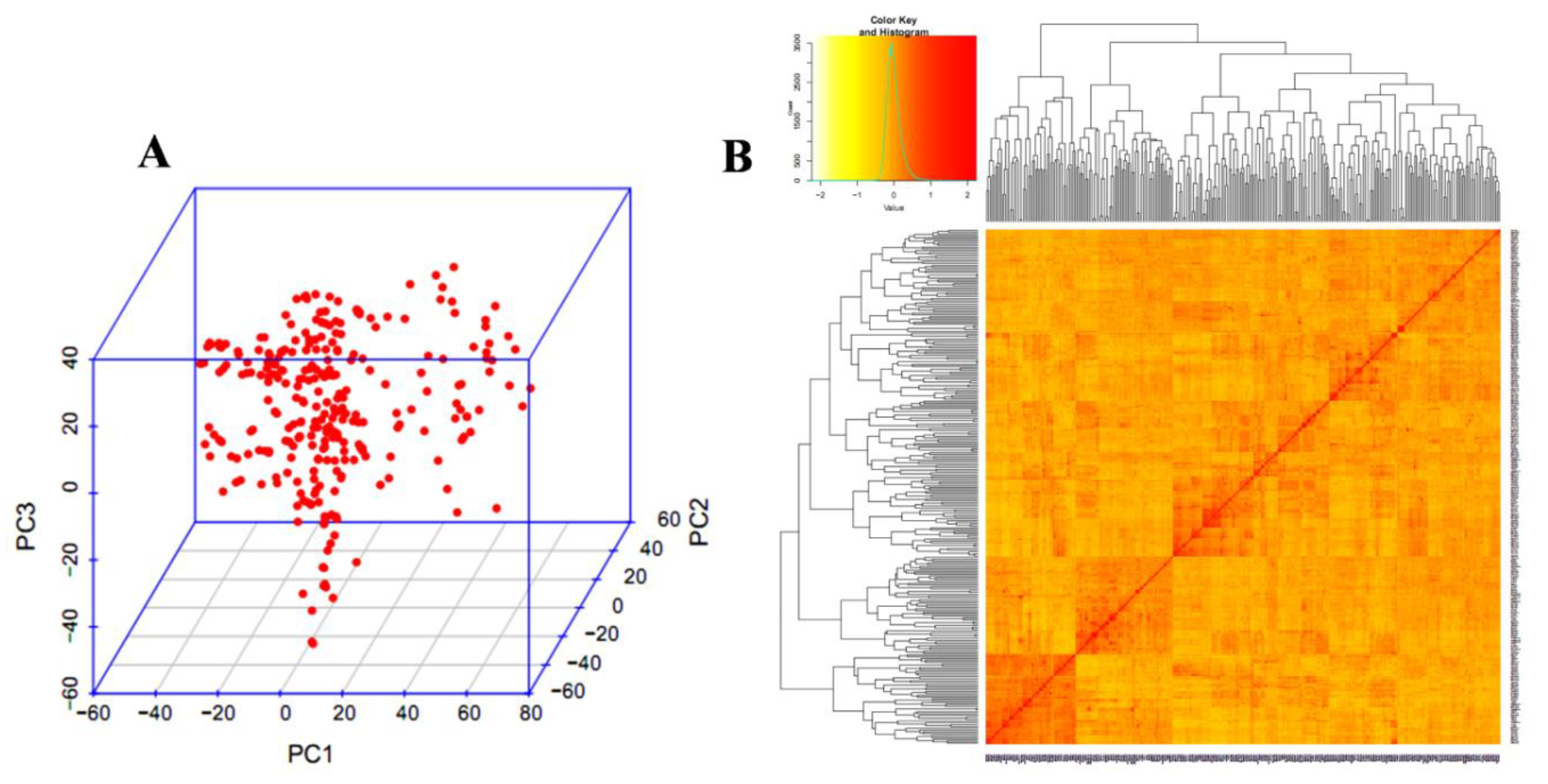
3.3. Population Structure and LD
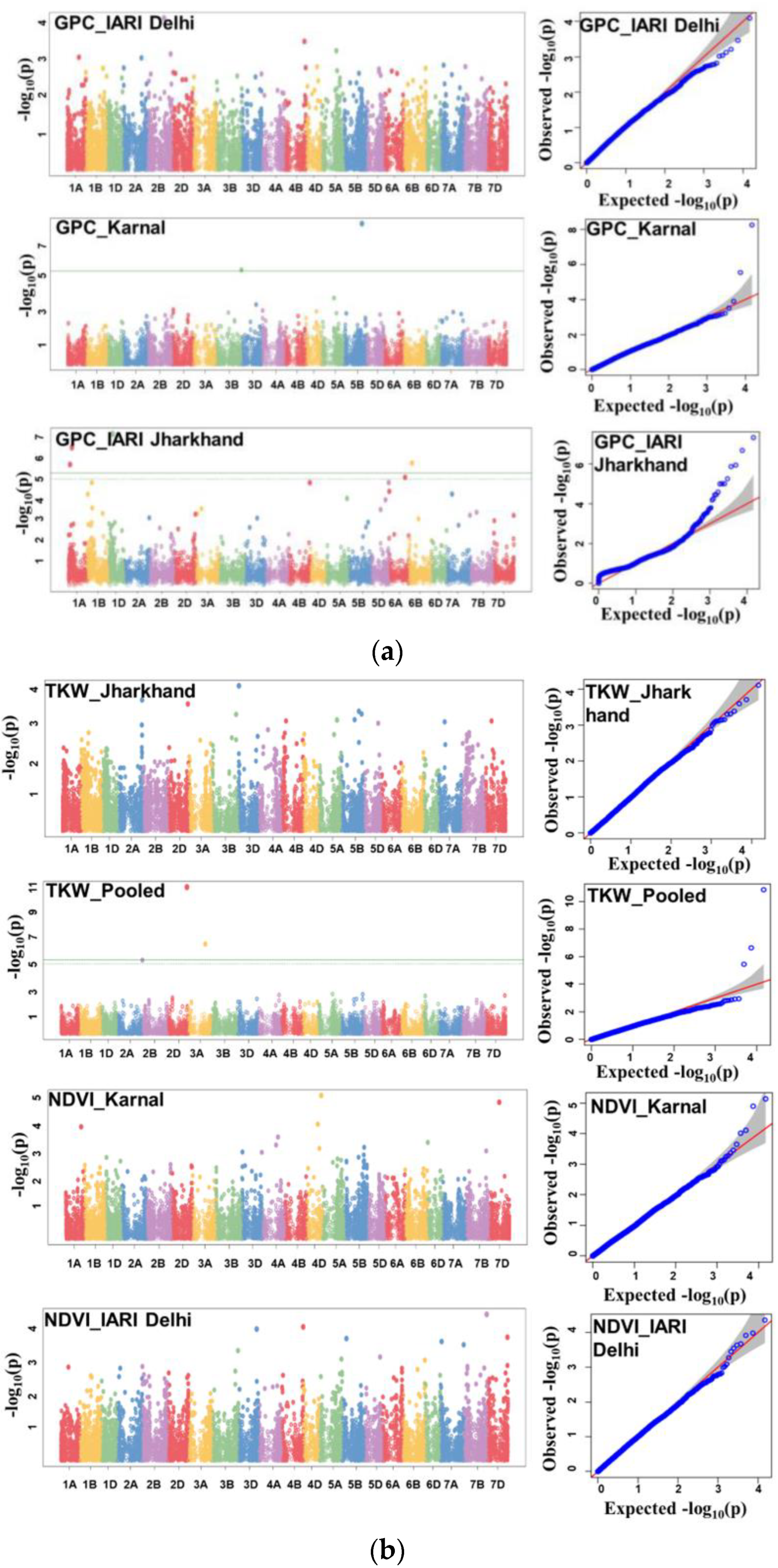
3.4. Genome-Wide Association Studies
3.5. Putative Candidate Genes Associated with MTAs
4. Discussion
4.1. Variability, Correlation, and GEI
4.2. Linkage Disequilibrium
4.3. MTAs
4.4. Putative Candidate Genes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schaible, U.E.; Kaufmann, S.H.E. Malnutrition and infection: Complex mechanisms and global impacts. PLoS Med. 2007, 4, e115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kar, B.R.; Rao, S.L.; Chandramouli, B.A. Cognitive development in children with chronic protein energy malnutrition. Behav. Brain Funct. 2008, 4, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shewry, P.R.; Halford, N.G. Cereal seed storage proteins: Structure, properties and role in grain utilization. J. Exp. Bot. 2002, 53, 947–958. [Google Scholar] [CrossRef] [Green Version]
- Delwiche, S.R. Chapter 11. In Cereal Grains: Assessing and Managing Quality; Wrigley, C.W., Batey, I.L., Eds.; Woodhead Publishing: Cambridge, UK, 2010; Volume 1, pp. 267–310. [Google Scholar]
- Karaman, M. Evaluation of the physiological and agricultural properties of some of the bread wheat (Triticum aestivum L.) genotypes registered in turkey using biplot analysis. Pak. J. Bot. 2020, 52, 1989–1997. [Google Scholar] [CrossRef]
- Krishnappa, G.; Ahlawat, A.K.; Shukla, R.B.; Singh, S.K.; Singh, S.K.; Singh, A.M.; Singh, G.P. Multi-environment analysis of grain quality traits in recombinant inbred lines of a biparental cross in bread wheat (Triticum aestivum L.). Cereal Res. Commun. 2019, 47, 334–344. [Google Scholar] [CrossRef]
- Hernandez-Espinosa, N.; Mondal, S.; Autrique, E.; Gonzalez-Santoyo, H.; Crossa, J.; Huerta-Espino, J.; Singh, R.P.; Guzman, C. Milling, processing and end-use quality traits of CIMMYT spring bread wheat germplasm under drought and heat stress. Field Crops Res. 2018, 215, 104–112. [Google Scholar] [CrossRef]
- Studnicki, M.; Wijata, M.; Sobczynski, G.; Samborski, S.; Gozdowski, D.; Rozbicki, J. Effect of genotype, environment and crop management on yield and quality traits in spring wheat. J. Cereal Sci. 2016, 72, 30–37. [Google Scholar] [CrossRef]
- Suhalia, A.; Sharma, A.; Kaur, S.; Sarlach, R.S.; Shokat, S.; Singh, S.; Rehman Arif, M.A.; Singh, S. Characterization of wheat Mexican landraces for drought and salt stress tolerance potential for future breeding. Cereal Res. Commun. 2022, 1–12. [Google Scholar] [CrossRef]
- Johnson, V.A.; Mattern, P.J.; Peterson, C.J.; Kuhr, S.L. Improvement of wheat protein by traditional breeding and genetic techniques. Cereal Chem. 1985, 62, 350–355. [Google Scholar]
- Avivi, L. High grain protein content in wild tetraploid wheat Triticum dicoccoides Korn. In Proceedings of the Fifth International Wheat Genetics Symposium, New Delhi, India, 23–28 February 1978; pp. 372–380. [Google Scholar]
- Distelfeld, A.; Uauy, C.; Fahima, T.; Dubcovsky, J. Physical map of the wheat high-grain protein content gene Gpc-B1 and development of a high-throughput molecular marker. New Phytol. 2006, 169, 753–763. [Google Scholar] [CrossRef] [Green Version]
- Distelfeld, A.; Cakmak, I.; Peleg, Z.; Ozturk, L.; Yazici, A.M.; Budak, H.; Saranga, Y.; Fahima, T. Multiple QTL-effects of wheat Gpc-B1 locus on grain protein and micronutrient concentrations. Plant Physiol. 2007, 129, 635–643. [Google Scholar] [CrossRef] [Green Version]
- Uauy, C.; Distelfeld, A.; Fahima, T.; Blechl, A.; Dubcovsky, J.A. NAC gene regulating senescence improves grain protein, zinc, and iron content in wheat. Science 2006, 314, 1298–1301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crain, J.; Ortiz-Monasterio, I.; Raun, B. Evaluation of a reduced cost active NDVI sensor for crop nutrient management. J. Sens. 2012, 2012, 582028. [Google Scholar] [CrossRef] [Green Version]
- Araus, J.L.; Slafer, G.A.; Royo, C.; Serret, M.D. Breeding for yield potential and stress. Adaptation in cereals. Crit. Rev. Plant Sci. 2008, 27, 377–412. [Google Scholar] [CrossRef]
- Poudel, R.; Bhinderwala, F.; Morton, M.; Powers, R.; Rose, D.J. Metabolic profiling of historical and modern wheat cultivars using proton nuclear magnetic resonance spectroscopy. Sci. Rep. 2021, 11, 3080. [Google Scholar] [CrossRef] [PubMed]
- Zorb, C.; Ludewig, U.; Hawkesford, M.J. Perspective on wheat yield and quality with reduced nitrogen supply. Trends Plant Sci. 2018, 23, 1029–1037. [Google Scholar] [CrossRef] [Green Version]
- Rehman Arif, M.A.; Shokat, S.; Plieske, J.; Lohwasser, U.; Chesnokov, Y.V.; Kumar, N.; Kulwal, P.; McGuire, P.; Sorrells, M.; Qualset, C.O.; et al. A SNP-based genetic dissection of versatile traits in bread wheat (Triticum aestivum L.). Plant J. 2021, 108, 960–976. [Google Scholar] [CrossRef]
- Akram, S.; Rehman Arif, M.A.; Hameed, A. A GBS-based GWAS analysis of adaptability and yield traits in bread wheat (Triticum aestivum L.). J. Appl. Genet. 2020, 62, 27–41. [Google Scholar] [CrossRef]
- Jadon, V.; Sharma, S.; Krishna, H.; Krishnappa, G.; Gajghate, R.; Devate, N.B.; Panda, K.K.; Jain, N.; Singh, P.K.; Singh, G.P. Molecular Mapping of Biofortification Traits in Bread Wheat (Triticum aestivum L.) Using a High-Density SNP Based Linkage Map. Genes 2023, 14, 221. [Google Scholar] [CrossRef]
- Krishnappa, G.; Rathan, N.D.; Sehgal, D.; Ahlawat, A.K.; Singh, S.K.; Singh, S.K.; Shukla, R.B.; Jaiswal, J.P.; Solanki, I.S.; Singh, G.P.; et al. Identification of novel genomic regions for biofortification traits using an SNP marker-enriched linkage map in wheat (Triticum aestivum L.). Front. Nutr. 2021, 8, 669444. [Google Scholar] [CrossRef]
- Chen, H.; Bemister, D.H.; Iqbal, M.; Strelkov, S.E.; Spaner, D.M. Mapping genomic regions controlling agronomic traits in spring wheat under conventional and organic managements. Crop Sci. 2020, 60, 2038–2052. [Google Scholar] [CrossRef]
- Marcotuli, I.; Gadaleta, A.; Mangini, M.; Signorile, A.M.; Zacheo, S.A.; Blanco, A.; Simeone, R.; Colasuonno, P. Development of a High-Density SNP-Based Linkage Map and Detection of QTL for β-Glucans, Protein Content, Grain Yield per Spike and Heading Time in Durum Wheat. Int. J. Mol. Sci. 2017, 18, 1329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, F.; Fan, X.; Chen, M.; Zhang, N.; Zhao, C.; Zhang, W.; Han, J.; Ji, J.; Zhao, X.; Yang, L.; et al. QTL detection for wheat kernel size and quality and the responses of these traits to low nitrogen stress. Theor. Appl. Genet. 2016, 129, 469–484. [Google Scholar] [CrossRef]
- Rathan, N.D.; Krishnappa, G.; Singh, A.-M.; Govindan, V. Mapping QTL for Phenological and Grain-Related Traits in a Mapping Population Derived from High-Zinc-Biofortified Wheat. Plants 2023, 12, 220. [Google Scholar] [CrossRef]
- Cabral, A.L.; Jordan, M.C.; Larson, G.; Somers, D.J.; Humphreys, D.G.; McCartney, C.A. Relationship between QTL for grain shape, grain weight, test weight, milling yield, and plant height in the spring wheat cross RL4452/‘AC Domain’. PLoS ONE 2018, 13, e0190681. [Google Scholar] [CrossRef] [Green Version]
- Krishnappa, G.; Singh, A.M.; Chaudhary, S.; Ahlawat, A.K.; Singh, S.K.; Shukla, R.B.; Jaiswal, J.P.; Singh, G.P.; Solanki, I.S. Molecular mapping of the grain iron and zinc concentration, protein content and thousand kernel weight in wheat (Triticum aestivum L.). PLoS ONE 2017, 12, e0174972. [Google Scholar] [CrossRef] [Green Version]
- Cuthbert, J.L.; Somers, D.J.; Brule-Babel, A.L.; Brown, P.D.; Crow, G.H. Molecular mapping of quantitative trait loci for yield and yield components in spring wheat (Triticum aestivum L.). Theor. Appl. Genet. 2008, 117, 595–608. [Google Scholar] [CrossRef]
- Sunil, H.; Upadhyay, D.; Gajghate, R.; Shashikumara, P.; Chouhan, D.; Singh, S.; Sunilkumar, V.P.; Manu, B.; Sinha, N.; Singh, S.; et al. QTL mapping for heat tolerance related traits using backcross inbred lines in wheat (Triticum aestivum L.). Indian J. Genet. 2020, 80, 242–249. [Google Scholar]
- Condorelli, G.E.; Maccaferri, M.; Newcomb, M.; Andrade-Sanchez, P.; White, J.W.; French, A.N.; Sciara, G.; Ward, R.; Tuberosa, R. Comparative Aerial and Ground Based High Throughput Phenotyping for the Genetic Dissection of NDVI as a Proxy for Drought Adaptive Traits in Durum Wheat. Front. Plant Sci. 2018, 9, 893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, F.; Wen, W.; Liu, J.; Rasheed, A.; Yin, G.; Xia, X.; Wu, X.; He, Z. Genome-Wide Linkage Mapping of QTL for Yield Components, Plant Height and Yield-Related Physiological Traits in the Chinese Wheat Cross Zhou 8425B/Chinese Spring. Front. Plant Sci. 2015, 6, 1099. [Google Scholar] [CrossRef] [Green Version]
- Flintgarcia, S.A.; Tornsberry, J.M.; And, E.S.; Buckler, I.V. Structure of linkage disequilibrium in plants. Annu. Rev. Plant Biol. 2003, 54, 357–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zondervan, K.T.; Cardon, L.R. The complex interplay among factors that influence allelic association. Nat. Rev. Genet. 2004, 5, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Korte, A.; Farlow, A. The advantages and limitations of trait analysis with GWAS: A review. Plant Methods 2013, 9, 29–38. [Google Scholar] [CrossRef] [Green Version]
- Pang, Y.; Liu, C.; Wang, D.; Amand, P.S.; Bernardo, A.; Li, W.; He, F.; Li, L.; Wang, L.; Yuan, X.; et al. High-resolution genome-wide association study identifies genomic regions and candidate genes for important agronomic traits in wheat. Mol. Plant 2020, 13, 1311–1327. [Google Scholar] [CrossRef] [PubMed]
- Brachi, B.; Morris, G.P.; Borevitz, J.O. Genome-wide association studies in plants: The missing heritability is in the field. Genome Biol. 2011, 12, 232. [Google Scholar] [CrossRef] [Green Version]
- Kaler, A.S.; Gillman, J.D.; Beissinger, T.; Purcell, L.C. Comparing different statistical models and multiple testing corrections for association mapping in soybean and maize. Front. Plant Sci. 2020, 25, 1794. [Google Scholar] [CrossRef] [Green Version]
- Huang, M.; Liu, X.; Zhou, Y.; Summers, R.M.; Zhang, Z. Blink: A package for the next level of genome-wide association studies with both individuals and markers in the millions. Gigascience 2019, 8, giy154. [Google Scholar] [CrossRef] [Green Version]
- Suliman, S.; Alemu, A.; Abdelmula, A.A.; Badawi, G.H.; Al-Abdallat, A.; Tadesse, W. Genome-wide association analysis uncovers stable QTLs for yield and quality traits of spring bread wheat (Triticum aestivum) across contrasting environments. Plant Gene 2021, 25, 100269. [Google Scholar] [CrossRef]
- Nigro, D.; Gadaleta, A.; Mangini, G.; Colasuonno, P.; Marcotuli, I.; Giancaspro, A.; Giove, S.L.; Simeone, R.; Blanco, A. Candidate genes and genome-wide association study of grain protein content and protein deviation in durum wheat. Planta 2019, 249, 1157–1175. [Google Scholar] [CrossRef]
- Kumar, J.; Saripalli, G.; Gahlaut, V.; Goel, N.; Meher, P.K.; Mishra, K.K.; Mishra, P.C.; Sehgal, D.; Vikram, P.; Sansaloni, C.; et al. Genetics of Fe, Zn, b-carotene, GPC and yield traits in bread wheat (Triticum aestivum L.) using multi-locus and multi-traits GWAS. Euphytica 2018, 214, 219. [Google Scholar] [CrossRef]
- Alemu, A.; Suliman, S.; Hagras, A.; Thabet, S.; Al-Abdallat, A.; Abdelmula, A.A.; Tadesse, W. Multi-model genome-wide association and genomic prediction analysis of 16 agronomic, physiological and quality related traits in ICARDA spring wheat. Euphytica 2021, 217, 205. [Google Scholar] [CrossRef]
- Rahimi, Y.; Bihamta, M.R.; Taleei, A.; Alipour, H.; Ingvarsson, P.K. Genome-wide association study of agronomic traits in bread wheat reveals novel putative alleles for future breeding programs. BMC Plant Biol. 2019, 19, 541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sukumaran, S.; Reynolds, M.P.; Sansaloni, C. Genome-wide association analyses identify QTL hotspots for yield and component traits in durum wheat grown under yield potential, drought, and heat stress environments. Front. Plant Sci. 2018, 9, 81. [Google Scholar] [CrossRef] [Green Version]
- Sun, C.; Zhang, F.; Yan, X.; Zhang, X.; Dong, Z.; Cui, D.; Chen, F. Genome-wide association study for 13 agronomic traits reveals distribution of superior alleles in bread wheat from the Yellow and Huai Valley of China. Plant Biotech. J. 2017, 15, 953–969. [Google Scholar] [CrossRef] [Green Version]
- Rufo, R.; Lopez, A.; Lopes, M.S.; Bellvert, J.; Soriano, J.M. Identification of Quantitative Trait Loci Hotspots Affecting Agronomic Traits and High-Throughput Vegetation Indices in Rainfed Wheat. Front. Plant Sci. 2021, 12, 735192. [Google Scholar] [CrossRef] [PubMed]
- Pradhan, S.; Babar, M.A.; Bai, G.; Khan, J.; Shahi, D.; Avci, M.; Guo, J.; McBreen, J.; Asseng, S.; Gezan, S.; et al. Genetic dissection of heat-responsive physiological traits to improve adaptation and increase yield potential in soft winter wheat. BMC Genom. 2020, 21, 315. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.; Azam, F.I.; Li, H.; Chang, X.; Li, B.; Jing, R. Mapping QTL for stay-green and agronomic traits in wheat under diverse water regimes. Euphytica 2017, 213, 246. [Google Scholar] [CrossRef] [Green Version]
- Hitz, K.; Clark, A.J.; Sanford, D.A.V. Identifying nitrogen-use efficient soft red winter wheat lines in high and low nitrogen environments. Field Crops Res. 2017, 200, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Pinto, R.S.; Reynolds, M.P.; Mathews, K.; McIntyre, C.L.; Olivares-Villegas, J.J.; Chapman, S.C. Heat and drought adaptive QTL in a wheat population designed to minimize confounding agronomic effects. Theor. Appl. Genet. 2010, 121, 1001–1021. [Google Scholar] [CrossRef] [Green Version]
- Singh, K.; Batra, R.; Sharma, S.; Saripalli, G.; Gautam, T.; Singh, R.; Pal, S.; Malik, P.; Kumar, M.; Jan, I.; et al. WheatQTLdb: A QTL database for wheat. Mol. Genet. Genom. 2021, 296, 1051–1056. [Google Scholar] [CrossRef]
- Zimin, A.V.; Puiu, D.; Hall, R.; Kingan, S.; Clavijo, B.J.; Salzberg, S.L. The first near-complete assembly of the hexaploid bread wheat genome, Triticum aestivum. Gigascience 2017, 6, gix097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aravind, J.; Mukesh Sankar, S.; Wankhede, D.P.; Kaur, V. AugmentedRCBD: Analysis of Augmented Randomised Complete Block Designs. R Package Version 0.1.5.9000. 2021. Available online: https://aravind-j.github.io/augmentedRCBD (accessed on 18 November 2022).
- Murray, M.G.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. Tassel: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef] [PubMed]
- Lipka, A.E.; Tian, F.; Wang, Q.; Peiffer, J.; Li, M.; Bradbury, P.J.; Gore, M.A.; Buckler, E.S.; Zhang, Z. GAPIT: Genome association and prediction integrated tool. Bioinformatics 2012, 28, 2397–2399. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Zhang, Z. GAPIT version 3: Boosting power and accuracy for genomic association and prediction. Genom. Proteom. Bioinform. 2021, 19, 629–640. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhang, R.; Cheng, Y.; Lei, P.; Song, W.; Zheng, W.; Nie, X. Genome-wide identification, evolution, and expression analysis of LBD transcription factor family in bread wheat (Triticum aestivum L.). Front. Plant Sci. 2021, 12, 721253. [Google Scholar] [CrossRef]
- Chen, Y.; Sun, A.; Wang, M.; Zhu, Z.; Ouwerkerk, P.B. Functions of the CCCH type zinc finger protein OsGZF1 in regulation of the seed storage protein GluB-1 from rice. Plant Mol. Biol. 2014, 84, 621–634. [Google Scholar] [CrossRef]
- Li, J.; Zhang, L.; Yuan, Y.; Wang, Q.; Elbaiomy, R.G.; Zhou, W.; Wu, H.; Soaud, S.A.; Abbas, M.; Chen, B.; et al. In Silico Functional Prediction and Expression Analysis of C2H2 Zinc-Finger Family Transcription Factor Revealed Regulatory Role of ZmZFP126 in Maize Growth. Front. Genet. 2021, 12, 770427. [Google Scholar] [CrossRef]
- Sharma, G.; Upadyay, A.K.; Biradar, H.; Hittalmani, S. OsNAC-like transcription factor involved in regulating seed-storage protein content at different stages of grain filling in rice under aerobic conditions. J. Genet. 2019, 98, 18. [Google Scholar] [CrossRef]
- Alptekin, B.; Mangel, D.; Pauli, D.; Blake, T.; Lachowiec, J.; Hoogland, T.; Fischer, A.; Sherman, J. Combined effects of a glycine-rich RNA-binding protein and a NAC transcription factor extend grain fill duration and improve malt barley agronomic performance. Theor. Appl. Genet. 2021, 134, 351–366. [Google Scholar] [CrossRef]
- Waters, B.M.; Uauy, C.; Dubcovsky, J.; Grusak, M.A. Wheat (Triticum aestivum) NAM proteins regulate the translocation of iron, zinc, and nitrogen compounds from vegetative tissues to grain. J. Exp. Bot. 2009, 60, 4263–4274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricachenevsky, F.K.; Menguer, P.K.; Sperotto, R.A. kNACking on heaven’s door: How important are NAC transcription factors for leaf senescence and Fe/Zn remobilization to seeds? Front. Plant Sci. 2013, 4, 226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, L.; Liu, Y.; Sun, H.; Han, Y.; Li, J.; Li, C.; Guo, W.; Meng, H.; Li, S.; Fan, Y.; et al. The mitochondrial folylpolyglutamate synthetase gene is required for nitrogen utilization during early seedling development in Arabidopsis. Plant Physiol. 2013, 161, 971–989. [Google Scholar] [CrossRef] [Green Version]
- Bleukx, W.; Delcour, J.A. A Second Aspartic Proteinase Associated with Wheat Gluten. J. Cereal Sci. 2000, 32, 31–42. [Google Scholar] [CrossRef]
- Jia, H.; Li, M.; Li, W.; Liu, L.; Jian, Y.; Yang, Z.; Shen, X.; Ning, Q.; Du, Y.; Zhao, R.; et al. A serine/threonine protein kinase encoding gene KERNEL NUMBER PER ROW6 regulates maize grain yield. Nat. Commun. 2020, 11, 988. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Lu, G.; Liu, L.; Raihan, M.S.; Xu, J.; Jian, L.; Zhao, L.; Tran, T.M.; Zhang, Q.; Liu, J.; et al. The Kernel Size-Related Quantitative Trait Locus qKW9 Encodes a Pentatricopeptide Repeat Protein That Affects Photosynthesis and Grain Filling. Plant Physiol. 2020, 183, 1696–1709. [Google Scholar] [CrossRef]
- Zhang, L.; Qi, Y.; Wu, M.; Zhao, L.; Zhao, Z.; Lei, C.; Hao, Y.; Yu, X.; Sun, Y.; Zhang, X.; et al. Mitochondrion-targeted PENTATRICOPEPTIDE REPEAT5 is required for cis-splicing of nad4 intron 3 and endosperm development in rice. Crop J. 2021, 9, 282–296. [Google Scholar] [CrossRef]
- Chen, L.; Li, Y.X.; Li, C.; Shi, Y.; Song, Y.; Zhang, D.; Li, Y.; Wang, T. Genome-wide analysis of the pentatricopeptide repeat gene family in different maize genomes and its important role in kernel development. BMC Plant Biol. 2018, 18, 366. [Google Scholar] [CrossRef]
- Ren, R.C.; Lu, X.; Zhao, Y.J.; Wei, Y.M.; Wang, L.L.; Zhang, L.; Zhang, W.T.; Zhang, C.; Zhang, X.S.; Zhao, X.Y. Pentatricopeptide repeat protein DEK40 is required for mitochondrial function and kernel development in maize. J. Exp. Bot. 2019, 70, 6163–6179. [Google Scholar] [CrossRef]
- Liu, R.; Cao, S.-K.; Sayyed, A.; Xu, C.; Sun, F.; Wang, F.; Tan, B.-C. The Mitochondrial Pentatricopeptide Repeat Protein PPR18 Is Required for the cis-Splicing of nad4 Intron 1 and Essential to Seed Development in Maize. Int. J. Mol. Sci. 2020, 21, 4047. [Google Scholar] [CrossRef]
- Pan, Z.; Liu, M.; Xiao, Z.; Ren, X.; Zhao, H.; Gong, D.; Liang, K.; Tan, Z.; Shao, Y.; Qiu, F. ZmSMK9, a pentatricopeptide repeat protein, is involved in the cis-splicing of nad5, kernel development and plant architecture in maize. Plant Sci. 2019, 288, 110205. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhu, Y.; Xu, X.; Sun, F.; Yang, J.; Cao, L.; Luo, X. OstMAPKKK5, a truncated mitogen-activated protein kinase kinasekinase 5, positively regulates plant height and yield in rice. Crop J. 2019, 7, 707–714. [Google Scholar] [CrossRef]
- Ur Rehman, S.; Wang, J.; Chang, X.; Zhang, X.; Mao, X.; Jing, R. A wheat protein kinase gene TaSnRK2.9-5A associated with yield contributing traits. Theor. Appl. Genet. 2019, 132, 907–919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, M.; Wang, Q.; Li, Z.; Cheng, H.; Li, Z.; Liu, X.; Song, W.; Appels, R.; Zhao, H. Expression of TaCYP78A3, a gene encoding cytochrome P450 CYP78A3 protein in wheat (Triticum aestivum L.), affects seed size. Plant J. 2015, 83, 312–325. [Google Scholar] [CrossRef]
- Chen, Y.; Han, Y.; Zhang, M.; Zhou, S.; Kong, X.; Wang, W. Overexpression of the wheat expansin gene TaEXPA2 improved seed production and drought tolerance in transgenic tobacco plants. PLoS ONE 2016, 11, e0153494. [Google Scholar] [CrossRef] [Green Version]
- Gopalareddy, K.; Singh, A.M.; Ahlawat, A.K.; Singh, G.P.; Jaiswal, J.P. Genotype-environment interaction for grain iron and zinc concentration in recombinant inbred lines of a bread wheat (Triticum aestivum L.) cross. Indian J. Genet. Plant Breed. 2015, 75, 307–313. [Google Scholar] [CrossRef]
- Goel, S.; Singh, K.; Singh, B.; Grewal, S.; Dwivedi, N.; Alqarawi, A.A.; Abd Allah, E.F.; Ahmad, P.; Singh, N.K. Analysis of genetic control and QTL mapping of essential wheat grain quality traits in a recombinant inbred population. PLoS ONE 2019, 14, e0200669. [Google Scholar] [CrossRef] [Green Version]
- Tan, C.; Zhou, X.; Zhang, P.; Wang, Z.; Wang, D.; Guo, W.; Yun, F. Predicting grain protein content of field-grown winter wheat with satellite images and partial least square algorithm. PLoS ONE 2020, 15, e0228500. [Google Scholar] [CrossRef]
- Yu, H.; Deng, Z.; Xiang, C.; Tian, J. Analysis of diversity and linkage disequilibrium mapping of agronomic traits on B-genome of wheat. J. Genom. 2014, 2, 20–30. [Google Scholar] [CrossRef] [Green Version]
- Dinesh, A.; Patil, A.; Zaidi, P.H.; Kuchanur, P.H.; Vinayan, M.T.; Seetharam, K. Genetic diversity, linkage disequilibrium and population structure among CIMMYT maize inbred lines, selected for heat tolerance study. Maydica 2016, 61, 1–7. [Google Scholar]
- Dadshani, S.; Mathew, B.; Ballvora, A.; Mason, A.S.; Leon, J. Detection of breeding signatures in wheat using a linkage disequilibrium-corrected mapping approach. Sci. Rep. 2021, 11, 5527. [Google Scholar] [CrossRef]
- Sheoran, S.; Jaiswal, S.; Kumar, D.; Raghav, N.; Sharma, R.; Pawar, S.; Paul, S.; Iquebal, M.A.; Jaiswar, A.; Sharma, P.; et al. Uncovering genomic regions associated with 36 agro-morphological traits in Indian spring wheat using GWAS. Front. Plant Sci. 2019, 10, 527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vos, P.G.; Paulo, M.J.; Voorrips, R.E.; Visser, R.G.; van Eck, H.J.; van Eeuwijk, F.A. Evaluation of LD decay and various LD-decay estimators in simulated and SNP-array data of tetraploid potato. Theor. Appl. Genet. 2017, 130, 123–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, P.K.; Rustgi, S.; Kulwal, P.L. Linkage disequilibrium and association studies in higher plants: Present status and future prospects. Plant Mol. Biol. 2005, 57, 461–485. [Google Scholar] [CrossRef]
- Fatiukha, A.; Filler, N.; Lupo, I.; Lidzbarsky, G.; Klymiuk, V.; Korol, A.B.; Pozniak, C.; Fahima, T.; Krugman, T. Grain protein content and thousand kernel weight QTLs identified in a durum × wild emmer wheat mapping population tested in five environments. Theor. Appl. Genet. 2020, 133, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Zhang, G.; Guo, B.; Qu, C.; Zhang, M.; Kong, F.; Zhao, Y.; Li, S. QTL mapping for quality traits using a high-density genetic map of wheat. PLoS ONE 2020, 15, e0230601. [Google Scholar] [CrossRef] [PubMed]
- Muqaddasi, Q.H.; Brassac, J.; Ebmeyer, E.; Kollers, S.; Korzun, V.; Argillier, O.; Stiewe, G.; Plieske, J.; Ganal, M.W.; Röder, M.S. Prospects of GWAS and predictive breeding for European winter wheat’s grain protein content, grain starch content, and grain hardness. Sci. Rep. 2020, 10, 12541. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Chen, J.; Li, R.; Deng, Z.; Zhang, K.; Liu, B.; Tian, J. Conditional QTL mapping of three yield components in common wheat (Triticum aestivum L.). Crop J. 2016, 4, 220–228. [Google Scholar] [CrossRef] [Green Version]
- Halder, J.; Gill, H.S.; Zhang, J.; Altameemi, R.; Olson, E.; Turnipseed, B.; Sehgal, S.K. Genome-wide association analysis of spike and kernel traits in the U.S. hard winter wheat. Plant Genome 2023, 13, e20300. [Google Scholar] [CrossRef]
- Huang, X.Q.; Kempf, H.; Ganal, M.W.; Roder, M.S. Advanced backcross QTL analysis in progenies derived from a cross between a German elite winter wheat variety and a synthetic wheat (Triticum aestivum L.). Theor. Appl. Genet. 2004, 109, 933–943. [Google Scholar] [CrossRef]
- Lv, D.; Zhang, C.; Yv, R.; Yao, J.; Wu, J.; Song, X.; Jian, J.; Song, P.; Zhang, Z.; Han, D.; et al. Utilization of a Wheat50K SNP Microarray-Derived High-Density Genetic Map for QTL Mapping of Plant Height and Grain Traits in Wheat. Plants 2021, 10, 1167. [Google Scholar] [CrossRef] [PubMed]
- Giancaspro, A.; Giove, S.L.; Zacheo, S.A.; Blanco, A.; Gadaleta, A. Genetic Variation for Protein Content and Yield-Related Traits in a Durum Population Derived from an Inter-Specific Cross between Hexaploid and Tetraploid Wheat Cultivars. Front. Plant Sci. 2019, 10, 1509. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Env. | Mean ± SD | Range | CV (%) | LSD | h2BS | GCV | ECV |
|---|---|---|---|---|---|---|---|---|
| GPC (%) | E1 | 13.5 ± 1.18 | 10.81–16.71 | 4.77 | 1.94 | 70.28 | 7.32 | 4.76 |
| E2 | 13.9 ± 0.84 | 11.88–16.62 | 5.47 | 2.29 | 68.53 | 6.61 | 5.47 | |
| E3 | 11.8 ± 0.91 | 09.59–14.81 | 3.42 | 1.23 | 72.80 | 6.96 | 3.46 | |
| TKW (gm) | E1 | 40.55 ± 0.21 | 31.01–50.41 | 3.14 | 3.83 | 86.56 | 7.95 | 3.13 |
| E2 | 43.36 ± 0.26 | 29.48–55.98 | 3.85 | 5.02 | 84.15 | 8.85 | 3.84 | |
| E3 | 42.81 ± 0.26 | 28.38–52.98 | 2.37 | 3.05 | 94.64 | 9.95 | 2.37 | |
| NDVI | E1 | 0.49 ± 0.06 | 0.32–0.69 | 6.58 | 0.10 | 72.69 | 9.75 | 6.59 |
| E2 | 0.60 ± 0.05 | 0.44–0.71 | 7.65 | 0.14 | 68.18 | 7.28 | 7.64 | |
| E3 | 0.57 ± 0.04 | 0.46–0.68 | 7.20 | 0.12 | 70.95 | 6.53 | 7.21 |
| Trait | Environment | SNPs | Chr. | Position | p Value | PVE (%) |
|---|---|---|---|---|---|---|
| Grain protein content (%) | E1 | AX-94714023 | 2B | 536316470 | 8.12 × 10−5 | 10.2 |
| E2 | AX-95107750 | 1A | 112941690 | 2.09 × 10−7 | 6.6 | |
| AX-94825050 | 1A | 53188500 | 1.35 × 10−6 | 7.7 | ||
| AX-95082115 | 1B | 144122241 | 1.00 × 10−5 | 7.7 | ||
| AX-94749397 | 1B | 16478742 | 3.53 × 10−5 | 6.2 | ||
| AX-94675928 | 1D | 112354107 | 4.71 × 10−8 | 7.2 | ||
| AX-94770504 | 4B | 667680308 | 9.97 × 10−6 | 7.0 | ||
| AX-94384140 | 5A | 659165855 | 5.65 × 10−5 | 6.9 | ||
| AX-94617912 | 5D | 450634975 | 6.54 × 10−5 | 6.3 | ||
| AX-94520919 | 5D | 550185848 | 9.88 × 10−6 | 10.1 | ||
| AX-94537786 | 6A | 501176793 | 5.53 × 10−6 | 7.7 | ||
| AX-95186193 | 6A | 3311006 | 2.59 × 10−5 | 7.0 | ||
| AX-94412218 | 6B | 100291191 | 1.14 × 10−6 | 7.9 | ||
| AX-95199688 | 7A | 171387994 | 3.47 × 10−5 | 6.9 | ||
| E3 | AX-94746929 | 3B | 800933346 | 2.88 × 10−6 | 10.9 | |
| AX-95248629 | 5B | 580431598 | 5.61 × 10−9 | 11.4 | ||
| Thousand kernel weight | E2 | AX-94651901 | 3D | 4012915 | 7.79 × 10−5 | 13.8 |
| Pooled | AX-95194336 | 2B | 9620943 | 3.54 × 10−6 | 8.7 | |
| AX-94454052 | 2D | 617073435 | 1.41 × 10−11 | 13.4 | ||
| AX-94861851 | 3A | 544385295 | 2.31 × 10−7 | 10.7 | ||
| Normalized difference vegetation index | E1 | AX-95111632 | 4B | 667859119 | 1.06 × 10−4 | 10.6 |
| AX-94826552 | 7B | 717202719 | 4.46 × 10−5 | 12.1 | ||
| E3 | AX-95006755 | 1A | 485355517 | 9.70 × 10−5 | 6.2 | |
| AX-94978133 | 4D | 465771817 | 7.36 × 10−6 | 10.1 | ||
| AX-94736370 | 4D | 359118968 | 7.80 × 10−5 | 11.7 | ||
| AX-94493107 | 7D | 306757146 | 1.28 × 10−5 | 11.5 |
| Trait | SNP ID | Position | Chr | Trace ID | Putative Candidate Genes | Function |
|---|---|---|---|---|---|---|
| GPC | AX-95107750 | 112941690 | 1A | TraesCS1A02G111700 | Lateral organ boundaries, LOB | Stress tolerance in wheat [59] |
| AX-94537786 | 501176793 | 6A | TraesCS6A02G274300 | P-loop containing nucleoside triphosphate hydrolase | – | |
| TraesCS6A02G274400 | Zinc finger, RING-H2-type | Regulates glutelin protein accumulation in Rice via controlling of Glu B-1 promoter [60]. Regulation of grain-related traits in maize [61] | ||||
| AX-94520919 | 550185848 | 5D | TraesCS5D02G537600 | NAC domain superfamily | Protein, iron, and zinc remobilization in wheat [14]. Regulation of seed-storage protein content in rice [62]. Controls percent grain protein in barley [63]. Remobilization of iron, zinc, and nitrogen from vegetative tissues to developing grains in wheat [64]. Iron and zinc remobilization to seeds in Rice [65] | |
| AX-94770504 | 667680308 | 4B | TraesCS4B02G392600 | Folylpolyglutamate synthase | Nitrogen utilization in Arabidopsis [66] | |
| AX-95199688 | 171387994 | 7A | TraesCS7A02G208600 | Aspartic peptidase domain | Gluten aspartic proteinase (GlAP 2) is associated with gluten breakdown in wheat [67] | |
| TraesCS7A02G208700 | Aluminum-activated malate transporter | – | ||||
| TKW | AX-94651901 | 4012915 | 3D | TraesCS3D02G011300 | Serine/threonine-protein kinase LRK10-like | Regulates kernel number and ear length in Maize [68] |
| TraesCS3D02G011200 | Pentatricopeptide repeat | Controls photosynthesis and grain filling in maize [69]. Endosperm development in Rice [70]. Maize kernel-related traits including thousand kernel weight [71]. Pentatricopeptide repeat protein DEK45 [72], PPR18 [73] and ZmSMK9 [74] are required for mitochondrial function and kernel development in maize | ||||
| AX-94454052 | 617073435 | 2D | TraesCS2D02G530900 | Protein kinase-like domain superfamily | OstMAPKKK5 controls plant height and yield in rice [75]. TaSnRK2.9-5A has a role in high TKW and grains per spike [76] | |
| NDVI | AX-95111632 | 667859119 | 4B | TraesCS4B02G393700 | Cytochrome P450 | Regulates grain size in wheat [77] |
| AX-94978133 | 465771817 | 4D | TraesCS4D02G296100 | Expansin | TaEXPA2 regulates drought responsiveness in transgenic tobacco [78] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krishnappa, G.; Khan, H.; Krishna, H.; Devate, N.B.; Kumar, S.; Mishra, C.N.; Parkash, O.; Kumar, S.; Kumar, M.; Mamrutha, H.M.; et al. Genome-Wide Association Study for Grain Protein, Thousand Kernel Weight, and Normalized Difference Vegetation Index in Bread Wheat (Triticum aestivum L.). Genes 2023, 14, 637. https://doi.org/10.3390/genes14030637
Krishnappa G, Khan H, Krishna H, Devate NB, Kumar S, Mishra CN, Parkash O, Kumar S, Kumar M, Mamrutha HM, et al. Genome-Wide Association Study for Grain Protein, Thousand Kernel Weight, and Normalized Difference Vegetation Index in Bread Wheat (Triticum aestivum L.). Genes. 2023; 14(3):637. https://doi.org/10.3390/genes14030637
Chicago/Turabian StyleKrishnappa, Gopalareddy, Hanif Khan, Hari Krishna, Narayana Bhat Devate, Satish Kumar, Chandra Nath Mishra, Om Parkash, Sachin Kumar, Monu Kumar, Harohalli Masthigowda Mamrutha, and et al. 2023. "Genome-Wide Association Study for Grain Protein, Thousand Kernel Weight, and Normalized Difference Vegetation Index in Bread Wheat (Triticum aestivum L.)" Genes 14, no. 3: 637. https://doi.org/10.3390/genes14030637