
Integration Analysis of circRNA–miRNA–mRNA and Identification of Critical Networks in Valgus-Varus Deformity (Gallus gallus)
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal and Tissue Collection
2.2. Total RNA Isolation and Construction of RNA-seq Libraries
2.3. Screening of DEmRNAs, DEmiRNAs, and DEcircRNAs
2.4. DEmRNA Functional Analysis
2.5. Construction of DEmRNA PPI Network and Modular Analysis
2.6. DEmiRNA-DEmRNA and DEcircRNA-DEmRNA Regulated Relationship
2.7. KEGG Enrichment Analysis of DEcircRNAs, and DEmiRNAs
2.8. Construction of the circRNA–miRNA–mRNA Network
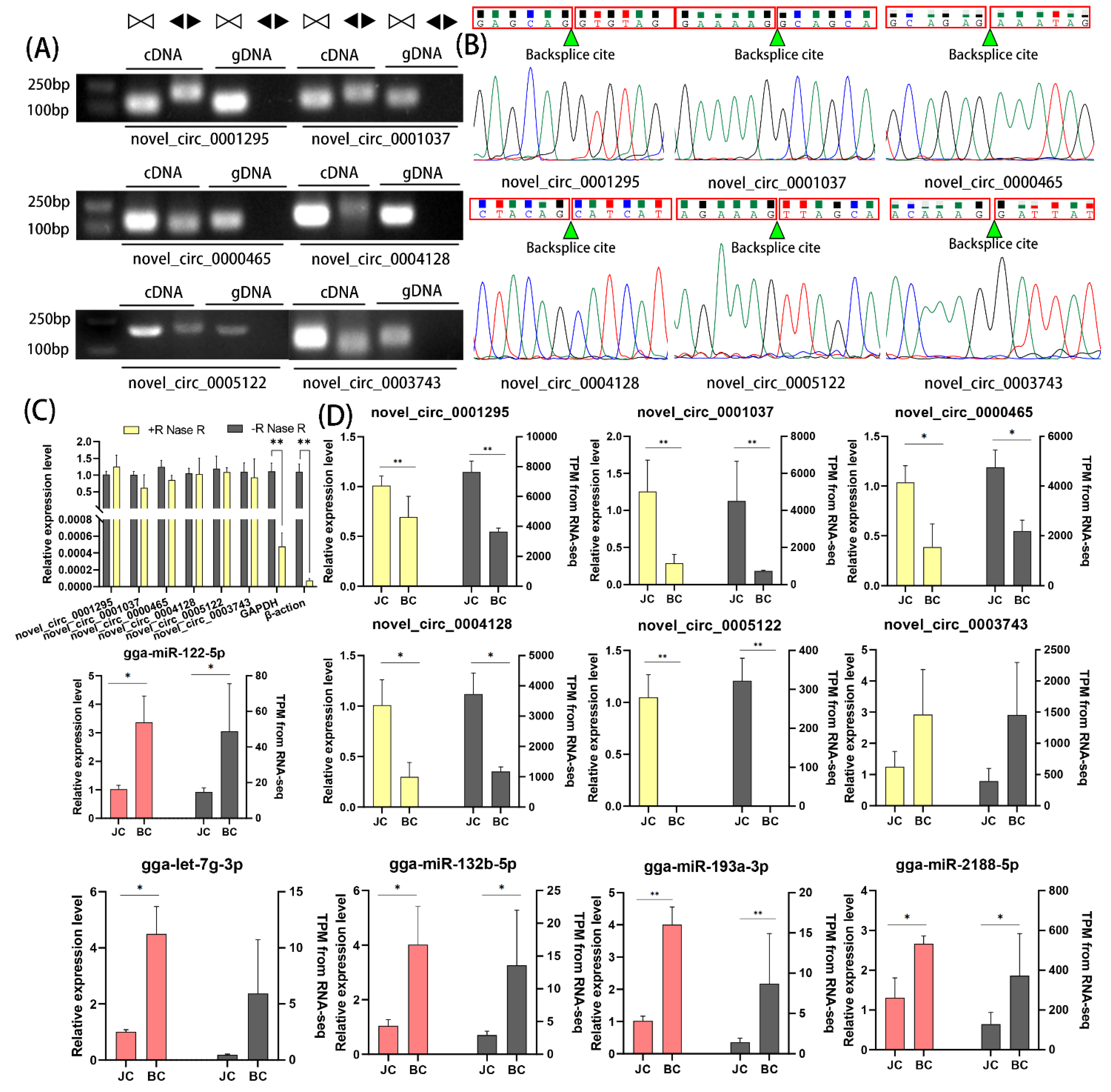
2.9. QPCR Validation of DEcircRNAs and DE miRNAs
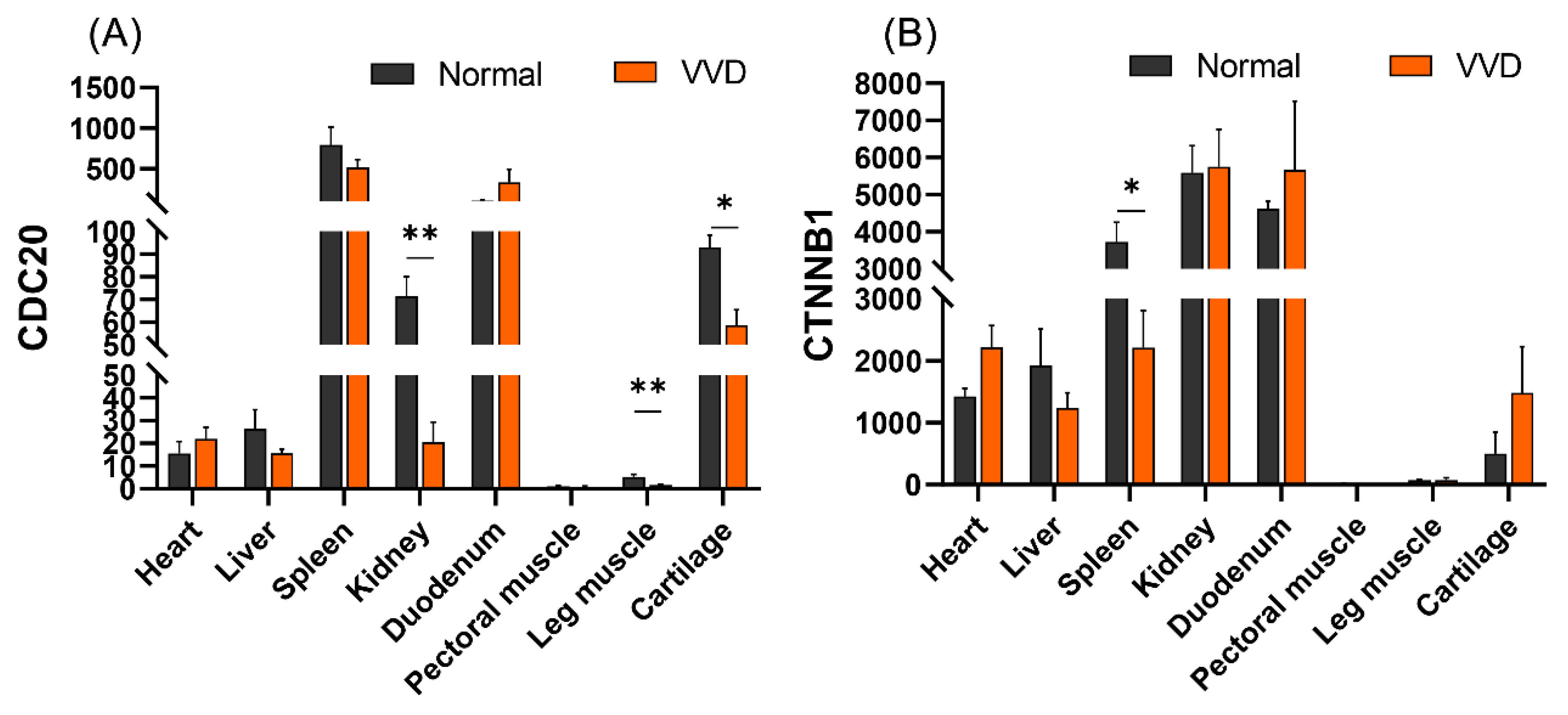
2.10. Expression profiles of CDC20 and CTNNB1
3. Results
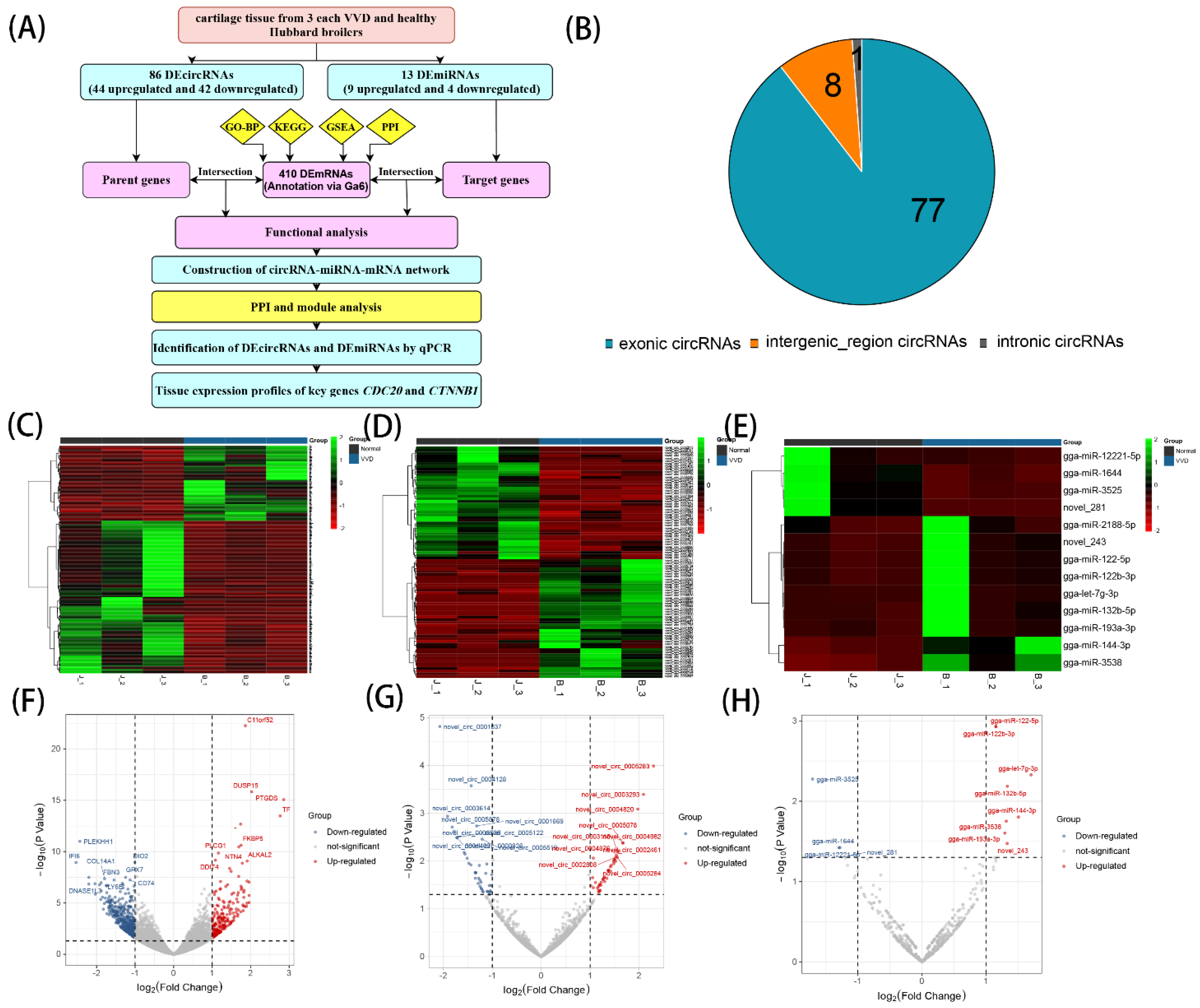
3.1. Identification of DEcircRNAs and DEmiRNAs in Chicken Leg Cartilage
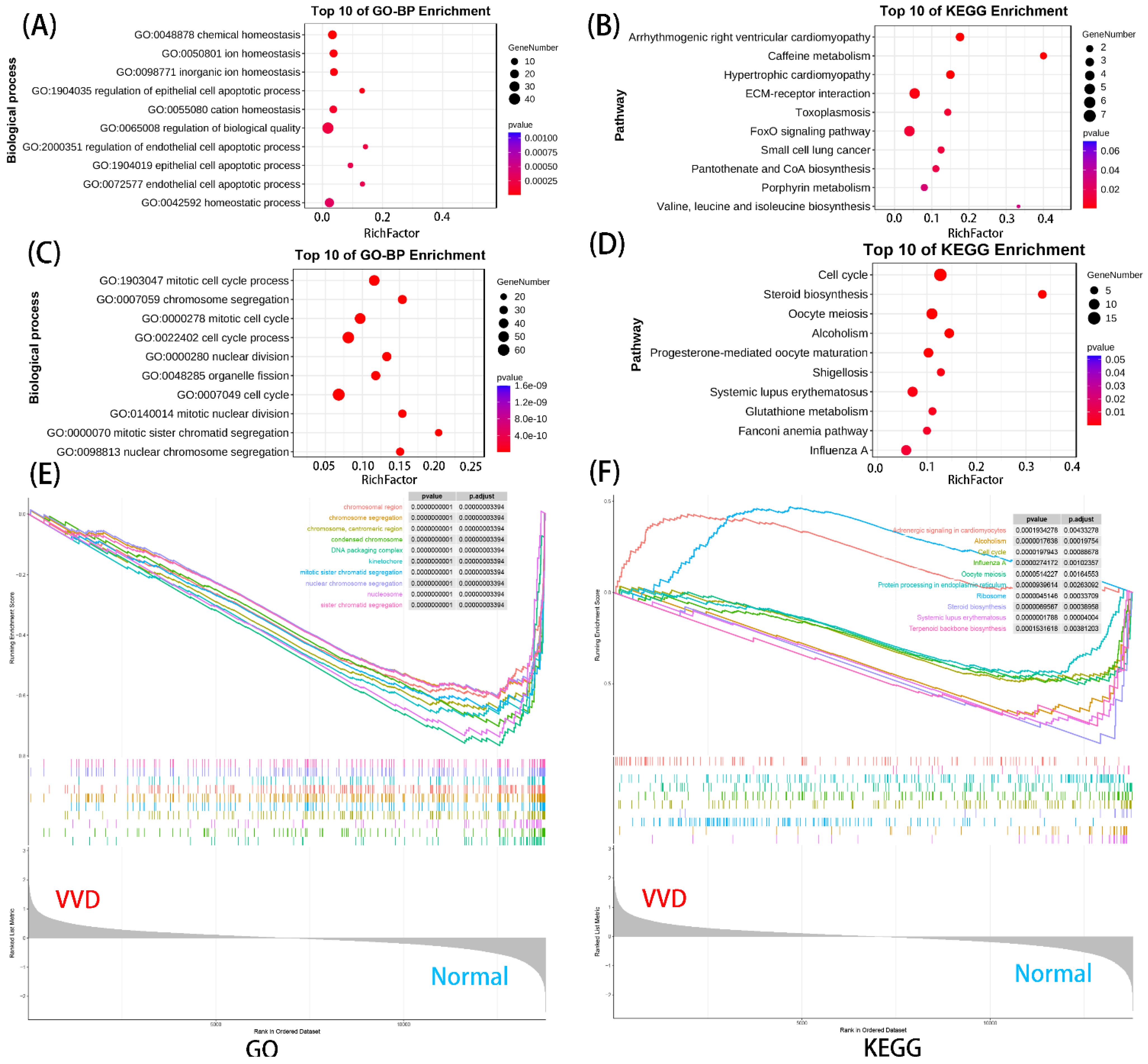
3.2. Functional Enrichment Analysis of DEmRNAs
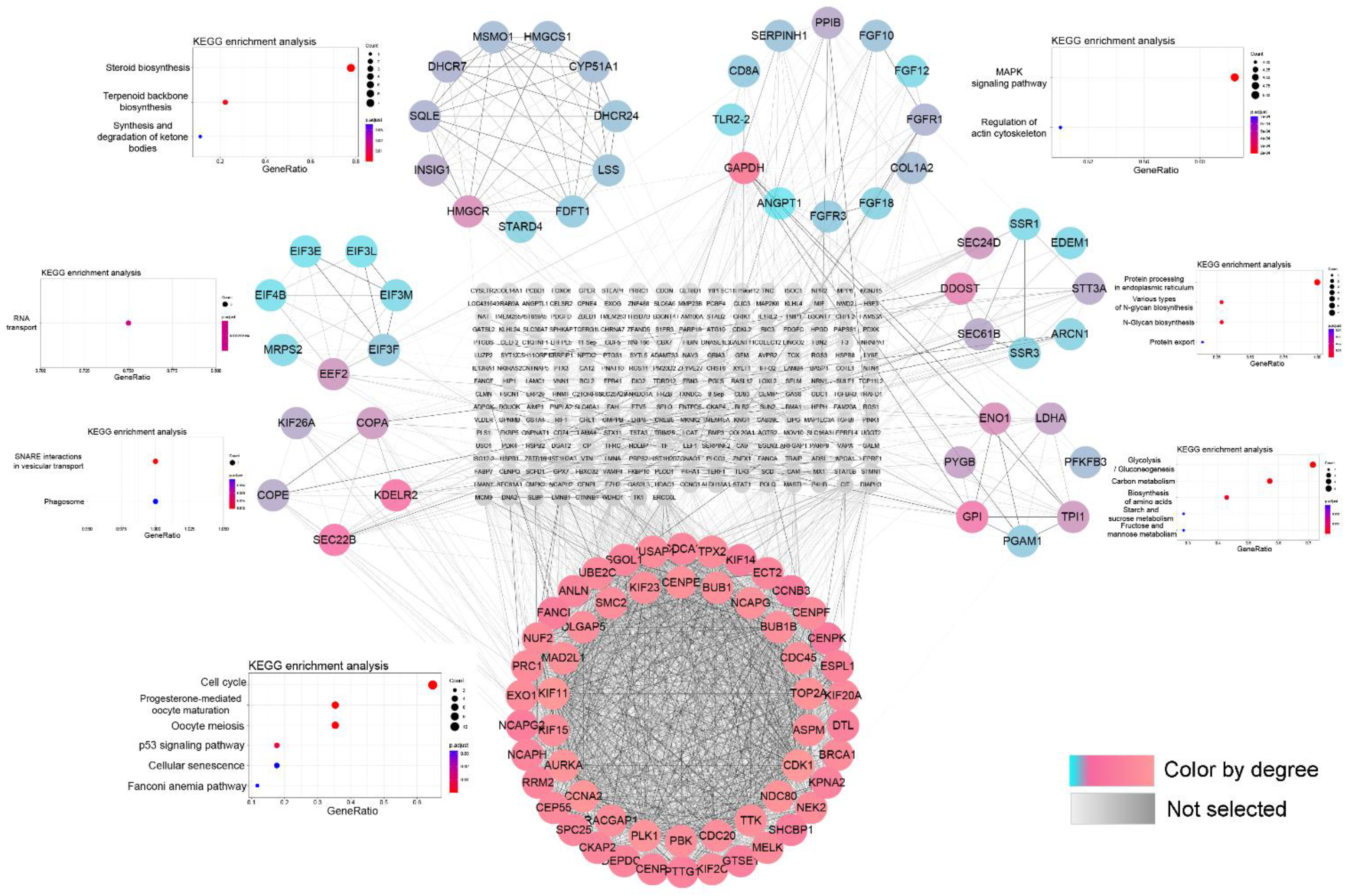
3.3. PPI Network and Module Function Analysis
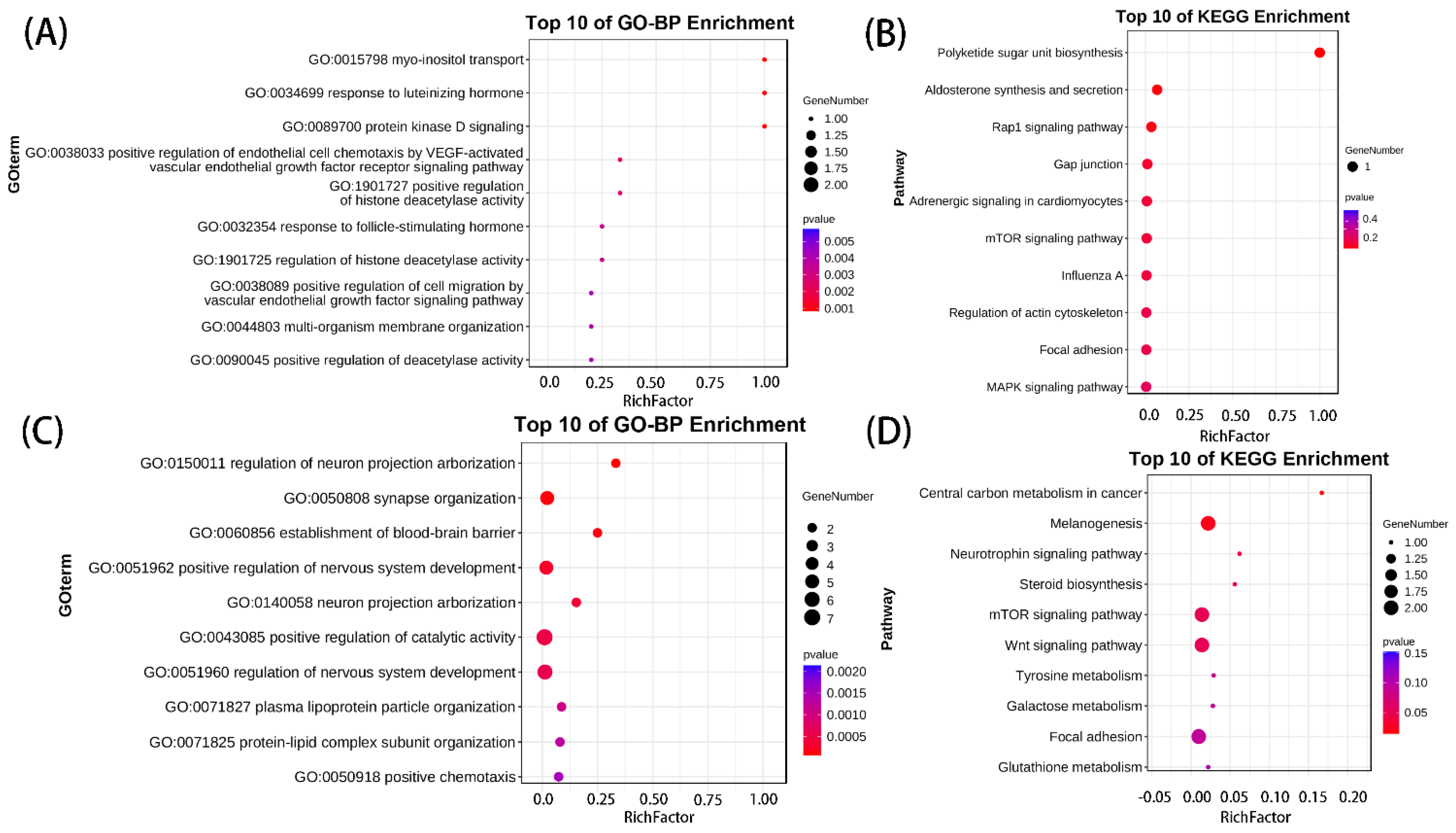
3.4. Enrichment Analysis on Overlapping Genes
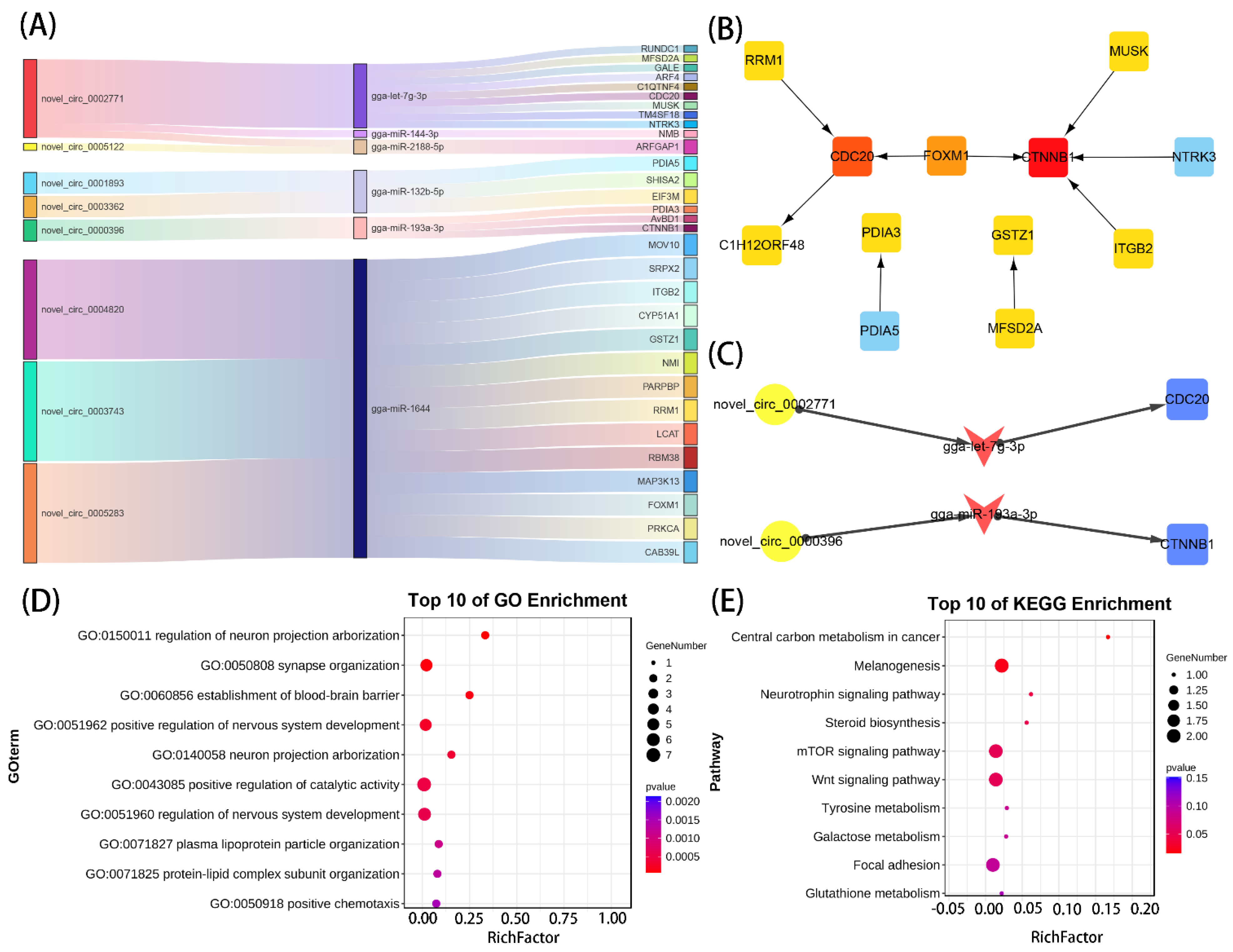
3.5. Construction of the circRNA–miRNA–mRNA Regulatory Network
3.6. qPCR Validation
3.7. Tissue Expression Profiles of CDC20 and CTNNB1
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Duff, S.R.; Thorp, B.H. Abnormal angulation/torsion of the pelvic appendicular skeleton in broiler fowl: Morphological and radiological findings. Res. Vet. Sci. 1985, 39, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Leterrier, C.; Nys, Y. Clinical and anatomical differences in varus and valgus deformities of chick limbs suggest different aetio-pathogenesis. Avian Pathol. 1992, 21, 429–442. [Google Scholar] [CrossRef] [PubMed]
- Randall, C.J.; Mills, C.P. Observations on leg deformity in broilers with particular reference to the intertarsal joint. Avian Pathol. 1981, 10, 407–431. [Google Scholar] [CrossRef] [PubMed]
- Shim, M.Y.; Karnuah, A.B.; Anthony, N.B.; Pesti, G.M.; Aggrey, S.E. The effects of broiler chicken growth rate on valgus, varus, and tibial dyschondroplasia. Poult Sci. 2012, 91, 62–65. [Google Scholar] [CrossRef]
- Cruickshank, J.J.; Sim, J.S. Morphometric and radiographic characteristics of tibial bone of broiler chickens with twisted leg disorders. Avian Dis. 1986, 30, 699–708. [Google Scholar] [CrossRef]
- Guo, Y.; Tang, H.; Wang, X.; Li, W.; Wang, Y.; Yan, F.; Kang, X.; Li, Z.; Han, R. Clinical assessment of growth performance, bone morphometry, bone quality, and serum indicators in broilers affected by valgus-varus deformity. Poult. Sci. 2019, 98, 4433–4440. [Google Scholar] [CrossRef]
- Newbrey, J.W.; Baksi, S.N.; Dhillon, A.S.; Zimmerman, N.G.; Truitt, S.G.; Riedinger, R. Histomorphometry and vitamin D metabolism of valgus-varus deformity in broiler chickens. Avian Dis. 1988, 32, 704–712. [Google Scholar] [CrossRef]
- Mattick, J.S.; Makunin, I.V. Non-coding RNA. Hum. Mol. Genet. 2006, 15, R17–R29. [Google Scholar] [CrossRef] [Green Version]
- Jeck, W.R.; Sharpless, N.E. Detecting and characterizing circular RNAs. Nat. Biotechnol. 2014, 32, 453–461. [Google Scholar] [CrossRef]
- Pasman, Z.; Been, M.D.; Garcia-Blanco, M.A. Exon circularization in mammalian nuclear extracts. RNA 1996, 2, 603–610. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, X.O.; Chen, T.; Xiang, J.F.; Yin, Q.F.; Xing, Y.H.; Zhu, S.; Yang, L.; Chen, L.L. Circular intronic long noncoding RNAs. Mol. Cell 2013, 51, 792–806. [Google Scholar] [CrossRef] [Green Version]
- Kloosterman, W.P.; Plasterk, R.H. The diverse functions of microRNAs in animal development and disease. Dev. Cell 2006, 11, 441–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krol, J.; Loedige, I.; Filipowicz, W. The widespread regulation of microRNA biogenesis, function and decay. Nat. Rev. Genet. 2010, 11, 597–610. [Google Scholar] [CrossRef] [PubMed]
- Salmena, L.; Poliseno, L.; Tay, Y.; Kats, L.; Pandolfi, P.P. A ceRNA hypothesis: The Rosetta Stone of a hidden RNA language? Cell 2011, 146, 353–358. [Google Scholar] [CrossRef] [Green Version]
- Kristensen, L.S.; Andersen, M.S.; Stagsted, L.V.W.; Ebbesen, K.K.; Hansen, T.B.; Kjems, J. The biogenesis, biology and characterization of circular RNAs. Nat. Rev. Genet. 2019, 20, 675–691. [Google Scholar] [CrossRef] [PubMed]
- Han, N.; Qian, F.; Niu, X.; Chen, G. Circ_0058792 regulates osteogenic differentiation through miR-181a-5p/Smad7 axis in steroid-induced osteonecrosis of the femoral head. Bioengineered 2022, 13, 12807–12822. [Google Scholar] [CrossRef] [PubMed]
- Hao, Y.; Lu, C.; Zhang, B.; Xu, Z.; Guo, H.; Zhang, G. CircPVT1 up-regulation attenuates steroid-induced osteonecrosis of the femoral head through regulating miR-21-5p-mediated Smad7/TGFβ signalling pathway. J. Cell Mol. Med. 2021, 25, 4608–4622. [Google Scholar] [CrossRef]
- Li, X.; Xie, C.; Xiao, F.; Su, H.; Li, Z.; Weng, J.; Huang, Y.; He, P. Circular RNA circ_0000423 regulates cartilage ECM synthesis via circ_0000423/miRNA-27b-3p/MMP-13 axis in osteoarthritis. Aging 2022, 14, 3400–3415. [Google Scholar] [CrossRef]
- Mao, G.; Xu, Y.; Long, D.; Sun, H.; Li, H.; Xin, R.; Zhang, Z.; Li, Z.; Yang, Z.; Kang, Y. Exosome-transported circRNA_0001236 enhances chondrogenesis and suppress cartilage degradation via the miR-3677-3p/Sox9 axis. Stem Cell Res. Ther. 2021, 12, 389. [Google Scholar] [CrossRef]
- Wu, S.; Huang, J.; Li, Y.; Liu, Z.; Zhao, L. Integrated analysis of lncrna and circrna mediated cerna regulatory networks in skin reveals innate immunity differences between wild-type and yellow mutant rainbow trout (oncorhynchus mykiss). Front. Immunol. 2022, 13, 802731. [Google Scholar] [CrossRef]
- Wahafu, P.; Xu, A.; Zhao, B.; Tuo, Y.; Yang, J. Circ_0005526 contributes to interleukin-1β-induced chondrocyte injury in osteoarthritis via upregulating transcription factor 4 by interacting with miR-142-5p. Bioengineered 2022, 13, 8407–8418. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Liu, T.; Yu, B.; Wang, B.; Peng, Q. CircRNA-UBE2G1 regulates LPS-induced osteoarthritis through miR-373/HIF-1a axis. Cell Cycle 2020, 19, 1696–1705. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Yang, B.; Zhou, S.; Wu, J. CircRNA circ_SEC24A upregulates DNMT3A expression by sponging miR-26b-5p to aggravate osteoarthritis progression. Int. Immunopharmacol. 2021, 99, 107957. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Luo, J.; Zeng, S. Circ-LRP1B functions as a competing endogenous RNA to regulate proliferation, apoptosis and oxidative stress of LPS-induced human C28/I2 chondrocytes. J. Bioenerg. Biomembr. 2022, 54, 93–108. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; He, P.; Pan, H.; Long, J.; Wang, J.; Li, Z.; Liu, H.; Jiang, W.; Zheng, Z. Circular RNA circ-4099 is induced by TNF-α and regulates ECM synthesis by blocking miR-616-5p inhibition of Sox9 in intervertebral disc degeneration. Exp. Mol. Med. 2018, 50, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Hu, S.; Ding, R.; Yuan, J.; Jia, J.; Wu, T.; Cheng, X. CircSNHG5 sponges Mir-495-3p and modulates CITED2 to protect cartilage endplate from degradation. Front. Cell Dev. Biol. 2021, 9, 668715. [Google Scholar] [CrossRef]
- Hansen, T.B.; Venø, M.T.; Damgaard, C.K.; Kjems, J. Comparison of circular RNA prediction tools. Nucleic Acids Res. 2016, 44, e58. [Google Scholar] [CrossRef] [Green Version]
- Memczak, S.; Jens, M.; Elefsinioti, A.; Torti, F.; Krueger, J.; Rybak, A.; Maier, L.; Mackowiak, S.D.; Gregersen, L.H.; Munschauer, M.; et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature 2013, 495, 333–338. [Google Scholar] [CrossRef]
- Gao, Y.; Wang, J.; Zhao, F. CIRI: An efficient and unbiased algorithm for de novo circular RNA identification. Genome Biol. 2015, 16, 4. [Google Scholar] [CrossRef] [Green Version]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein–protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2018, 47, D607–D613. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, V.; Bell, G.W.; Nam, J.-W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. eLife 2015, 4, e05005. [Google Scholar] [CrossRef] [PubMed]
- Bihan-Duval, E.L.; Beaumont, C.; Colleau, J.J. Estimation of the genetic correlations between twisted legs and growth or conformation traits in broiler chickens. J. Anim. Breed. Genet. 1997, 114, 239–259. [Google Scholar] [CrossRef] [PubMed]
- Rekaya, R.; Sapp, R.; Wing, T.; Aggrey, S. Genetic evaluation for growth, body composition, feed efficiency, and leg soundness. Poult. Sci. 2013, 92, 923–929. [Google Scholar] [CrossRef] [PubMed]
- Le Bihan-Duval, E.; Beaumont, C.; Colleau, J.J. Genetic parameters of the twisted legs syndrome in broiler chickens. Genet. Sel. Evol. 1996, 28, 177–195. [Google Scholar] [CrossRef]
- Yu, Y.; Wang, S.; Zhou, Z. Cartilage homeostasis affects femoral head necrosis induced by methylprednisolone in broilers. Int. J. Mol. Sci. 2020, 21, 4841. [Google Scholar] [CrossRef]
- Tian, W.-x.; Li, J.-k.; Qin, P.; Wang, R.; Ning, G.-b.; Qiao, J.-g.; Li, H.-q.; Bi, D.-r.; Pan, S.-y.; Guo, D.-z. Screening of differentially expressed genes in the growth plate of broiler chickens with Tibial Dyschondroplasia by microarray analysis. BMC Genomics. 2013, 14, 276. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Dong, J.; Zhang, P.; Zhou, D.; Zhang, Q. Transcriptome sequencing reveals key genes in three early phases of osteogenic, adipogenic, and chondrogenic differentiation of bone marrow mesenchymal stem cells in rats. Front. Mol. Biosci. 2021, 8, 782054. [Google Scholar] [CrossRef]
- Wang, X.; Ning, Y.; Zhang, P.; Poulet, B.; Huang, R.; Gong, Y.; Hu, M.; Li, C.; Zhou, R.; Lammi, M.J.; et al. Comparison of the major cell populations among osteoarthritis, Kashin–Beck disease and healthy chondrocytes by single-cell RNA-seq analysis. Cell Death Dis. 2021, 12, 551. [Google Scholar] [CrossRef]
- Cheng, Y.; Du, Y.; Zhang, X.; Zhang, P.; Liu, Y. Conditional knockout of Cdc20 attenuates osteogenesis in craniofacial bones. Tissue Cell 2022, 77, 101829. [Google Scholar] [CrossRef] [PubMed]
- Liang, F.; Peng, L.; Ma, Y.G.; Hu, W.; Zhang, W.B.; Deng, M.; Li, Y.M. Bioinformatics analysis and experimental validation of differentially expressed genes in mouse articular chondrocytes treated with IL-1β using microarray data. Exp. Ther. Med. 2022, 23, 6. [Google Scholar] [CrossRef]
- Xuan, F.; Yano, F.; Mori, D.; Chijimatsu, R.; Maenohara, Y.; Nakamoto, H.; Mori, Y.; Makii, Y.; Oichi, T.; Taketo, M.M.; et al. Wnt/β-catenin signaling contributes to articular cartilage homeostasis through lubricin induction in the superficial zone. Arthritis Res. Ther. 2019, 21, 247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golovchenko, S.; Hattori, T.; Hartmann, C.; Gebhardt, M.; Gebhard, S.; Hess, A.; Ausch, F.; Schlund, B.; von der Mark, K. Deletion of beta catenin in hypertrophic growth plate chondrocytes impairs trabecular bone formation. Bone 2013, 55, 102–112. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zou, L.; Lu, C.; He, D. Study on the mechanism of miR-330-3p in cartilage degeneration in temporomandibular joint osteoarthritis in rats. China J. Oral Maxillofac. Surg. 2022, 20, 320–327. [Google Scholar] [CrossRef]
- Wang, X.; Ning, Y.; Zhang, P.; Yang, L.; Wang, Y.; Guo, X. Chondrocytes damage induced by T-2 toxin via Wnt/β-catenin signaling pathway is involved in the pathogenesis of an endemic osteochondropathy, Kashin-Beck disease. Exp. Cell Res. 2017, 361, 141–148. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′-3′) | Amplification Length |
|---|---|---|
| novel_circ_0001295 Divergent primers | F: CCAGGCAATCGATGAGGAGT R: CATCCACGACGCTTGCAAAA | 197 bp |
| novel_circ_0001295 Convergent primers | F: TACATCGCAGAAGTTGCTCCAC R: ATCCATCCTTCACGAGGTAGC | 130 bp |
| novel_circ_0001037 Divergent primers | F: GCTCCAGAATGCATCAGAGGT R: TTCCGGTCTCCAATCACTGC | 175 bp |
| novel_circ_0001037 Convergent primers | F: ACCGGAACTCTGGGAAAAGC R: TACCCCTTTCCATTCCTGCG | 137 bp |
| novel_circ_0000465 Divergent primers | F: TACGTGGCTGGGCTTAAAGG R: TGGTCTGTAGGAGTCGATGC | 130 bp |
| novel_circ_0000465 Convergent primers | F: AAGAAACAGTCCTAGCGAGCA R: ATCACCTCTTCCGTACTCCAA | 146 bp |
| novel_circ_0004128 Divergent primers | F: TATGTCTTCGATGGTGCCTGT R: TTGCACTGTCATAGAGGGAGC | 160 bp |
| novel_circ_0004128 Convergent primers | F: AAGCAGAGCGATCTTCCGAC R: CACAGGCACCATCGAAGACA | 182 bp |
| novel_circ_0005122 Divergent primers | F: ATGTGATTGTGGATCCCGTCG R: TTCCCATCAACTGGTCTGCT | 199 bp |
| novel_circ_0005122 Convergent primers | F: CTTGGAGTGTGCGGTGTCT R: CCACTCATAGCACGTTGGGT | 190 bp |
| novel_circ_0003743 Divergent primers | F: GCCCGTACCAGACAAGGATA R: GTGGTAATCCTGTTGTGGGTCT | 139 bp |
| novel_circ_0003743 Convergent primers | F: GCATAACGGAGAGCACAGTGA R: CACCTGTGTCTTTGTTGGTCAG | 182 bp |
| gga-miR-122-5p | F: GCGTGGAGTGTGACAATGGT R: AGTGCAGGGTCCGAGGTATT | 67 bp |
| gga-let-7g-3p | F: CGCTGTACAGGCCACTGC R: GCAGGGTCCGAGGTATTCGC | 66 bp |
| gga-miR-132b-5p | F: CGCGACCATGGCTGTAGAC R: AGTGCAGGGTCCGAGGTATT | 65 bp |
| gga-miR-193a-3p | F: CGCGAACTGGCCTACAAAGT R: AGTGCAGGGTCCGAGGTATT | 66 bp |
| gga-miR-2188-5p | F: GGCGAAGGTCCAACCTCACA R: AGTGCAGGGTCCGAGGTATT | 66 bp |
| CDC20 | F: CTCTCCAGTGGGTCACGAAC | 108 bp |
| R: CTTGAGTCCGCACACCTCTT | ||
| CTNNB1 | F: TGCTGACTACCCAGTTGATGG | 199 bp |
| R: AGATACTAGCCCACCCCTCG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Ma, Y.; Cai, C.; Zhang, L.; Liu, X.; Jiang, R.; Li, D.; Li, Z.; Kang, X.; Tian, Y.; et al. Integration Analysis of circRNA–miRNA–mRNA and Identification of Critical Networks in Valgus-Varus Deformity (Gallus gallus). Genes 2023, 14, 622. https://doi.org/10.3390/genes14030622
Li J, Ma Y, Cai C, Zhang L, Liu X, Jiang R, Li D, Li Z, Kang X, Tian Y, et al. Integration Analysis of circRNA–miRNA–mRNA and Identification of Critical Networks in Valgus-Varus Deformity (Gallus gallus). Genes. 2023; 14(3):622. https://doi.org/10.3390/genes14030622
Chicago/Turabian StyleLi, Jianzeng, Yanchao Ma, Chunxia Cai, Lujie Zhang, Xinxin Liu, Ruirui Jiang, Donghua Li, Zhuanjian Li, Xiangtao Kang, Yadong Tian, and et al. 2023. "Integration Analysis of circRNA–miRNA–mRNA and Identification of Critical Networks in Valgus-Varus Deformity (Gallus gallus)" Genes 14, no. 3: 622. https://doi.org/10.3390/genes14030622