

Common Genetic Factors and Pathways in Alzheimer’s Disease and Ischemic Stroke: Evidences from GWAS


Abstract
:1. Introduction
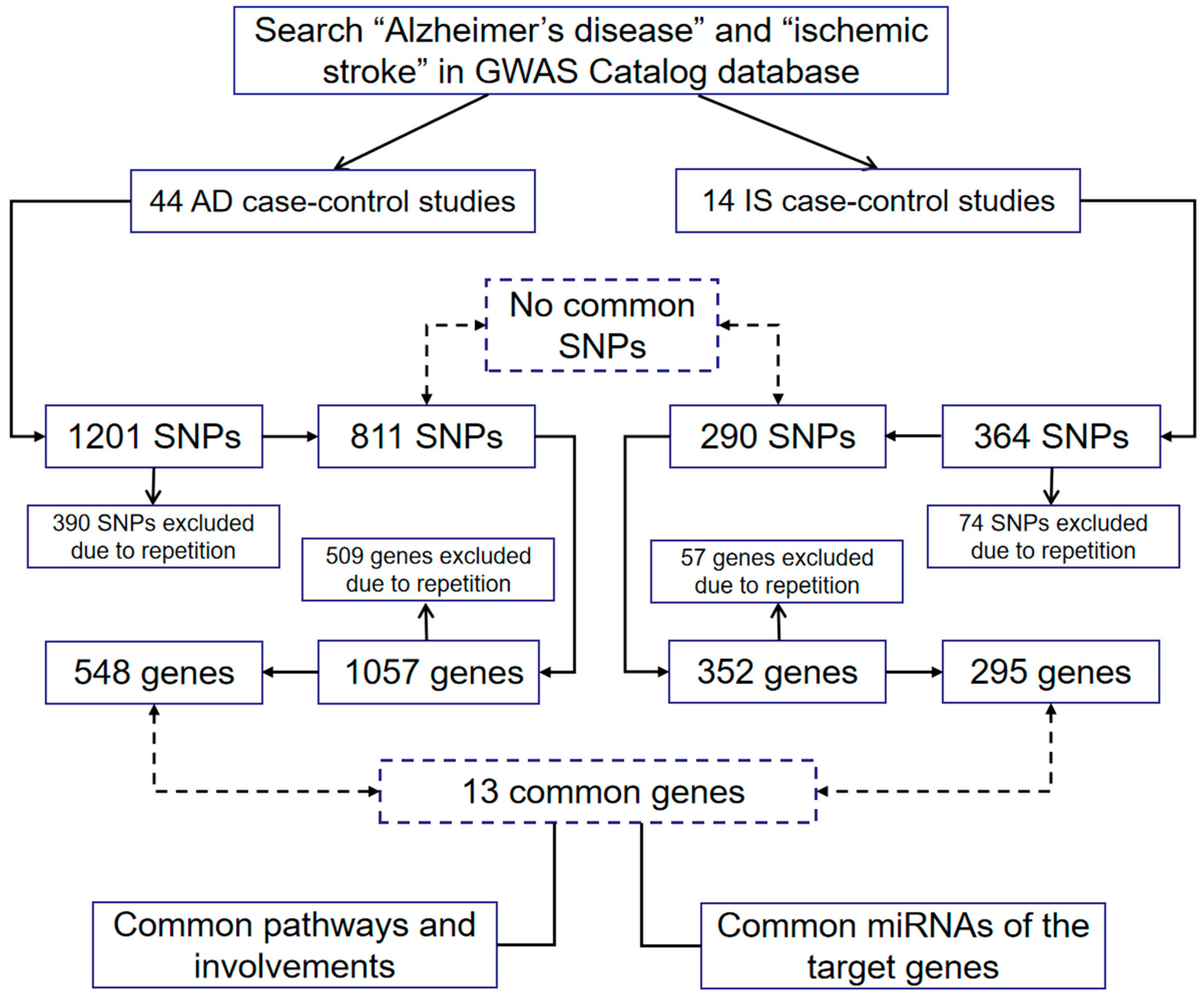
2. Searching Methodology
3. The Roles of AD or IS Causative Genes in Its Counterpart
3.1. AD Causative Genes in IS
3.2. IS Causative Genes in AD
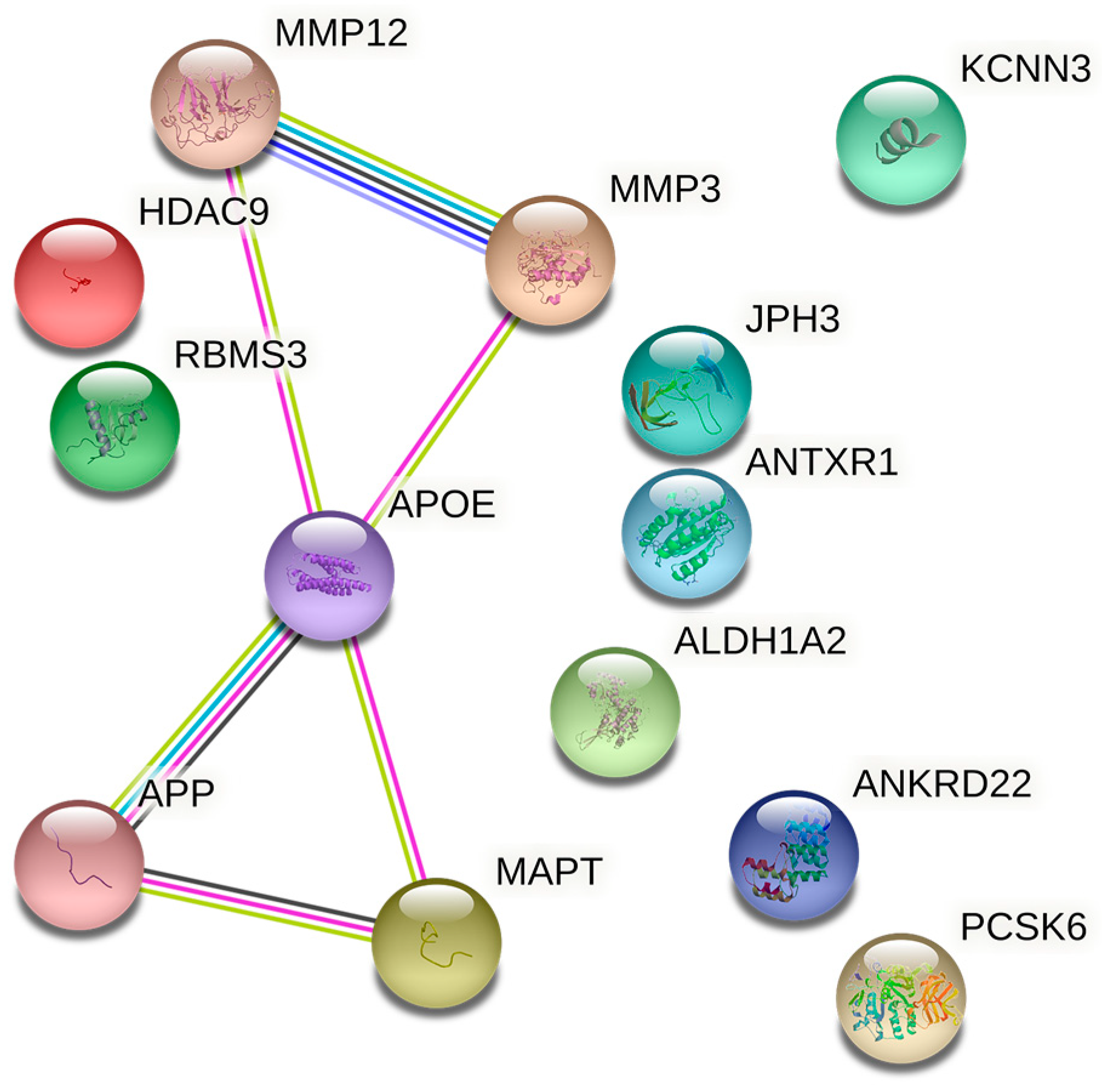
4. Common Susceptibility Genes of AD and IS
4.1. HDAC9
4.2. MMP3 and MMP12
4.3. Other Genes
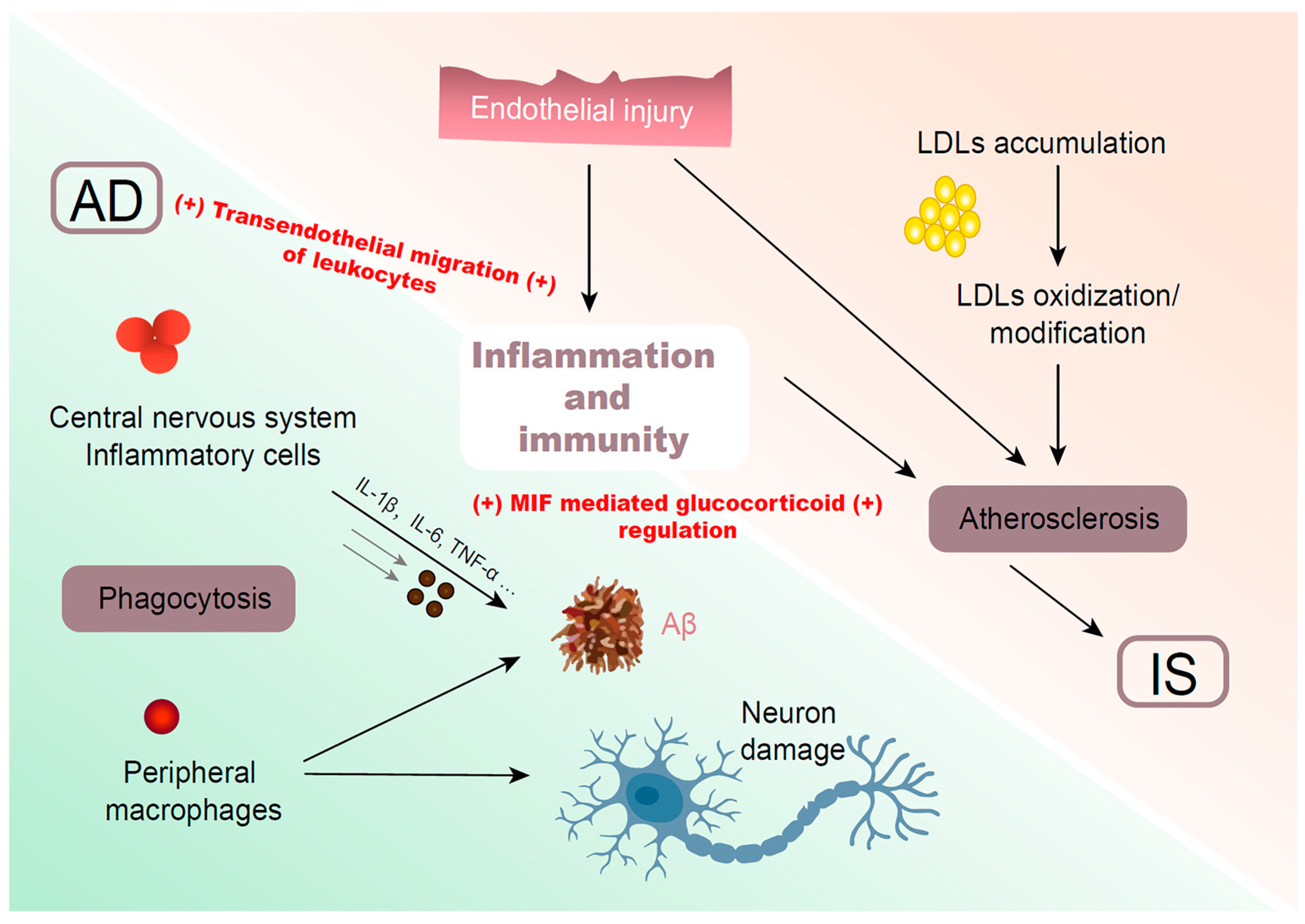
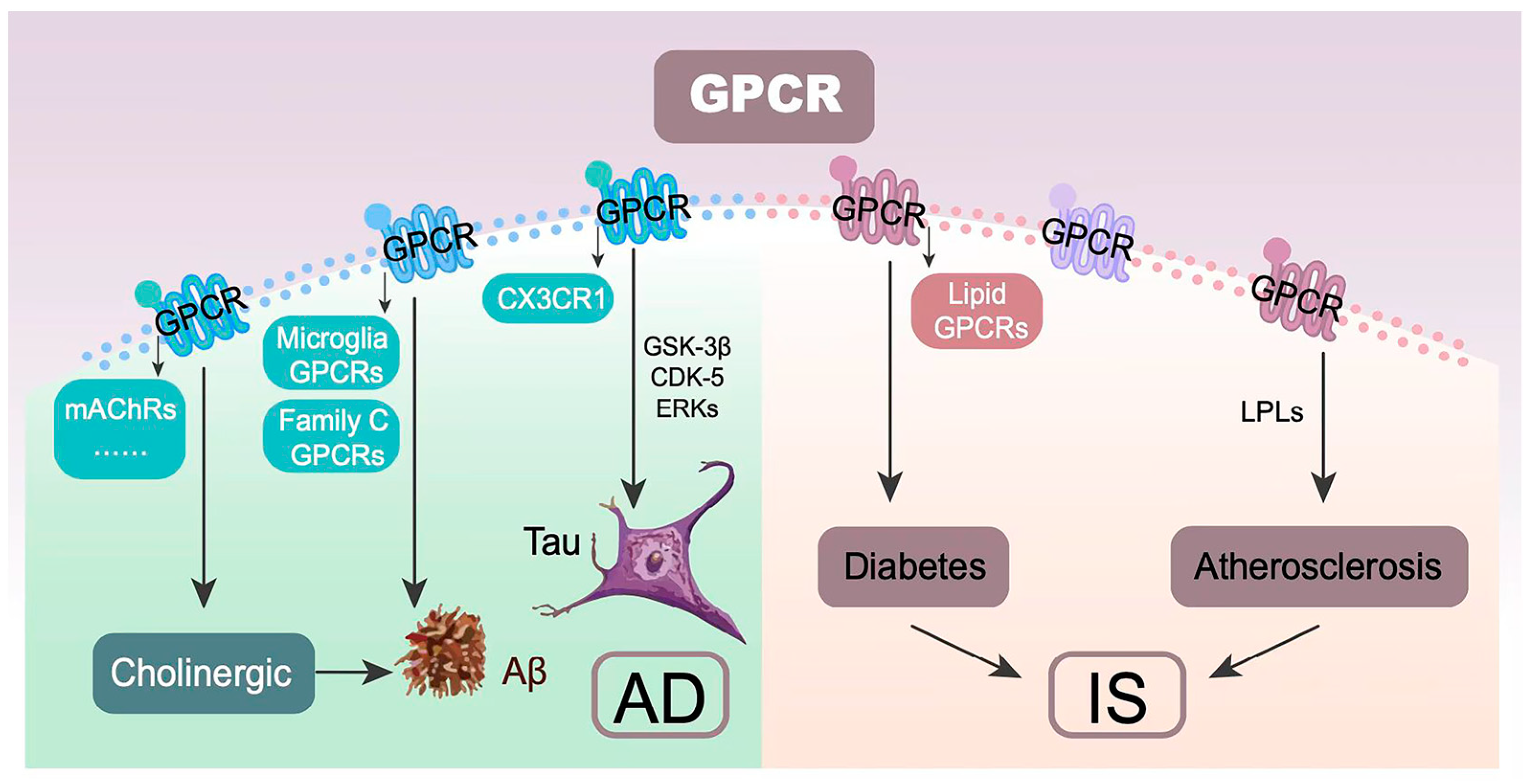
5. Common Molecular Pathways
5.1. Inflammation and Immunity
5.2. GPCR
5.3. Signal Transduction
6. Common miRNAs
7. Advantages and Limitations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Release notice—Dementia and stroke comorbidity among Canadians aged 65 years and older: Highlights from the Canadian Chronic Disease Surveillance System. Health Promot. Chronic. Dis. Prev. Can. 2020, 40, 324. [CrossRef] [PubMed]
- GBD 2016 Stroke Collaborators. Global, regional, and national burden of stroke, 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Neurol. 2019, 18, 439–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, J.A.; Arvanitakis, Z.; Bang, W.; Bennett, D.A. Mixed brain pathologies account for most dementia cases in community-dwelling older persons. Neurology 2007, 69, 2197–2204. [Google Scholar] [CrossRef] [PubMed]
- Traylor, M.; Adib-Samii, P.; Harold, D.; Alzheimer’s Disease Neuroimaging Initiative, The International Stroke Genetics Consortium (ISGC), UK Young Lacunar Stroke DNA resource; Dichgans, M.; Williams, J.; Lewis, C.M.; Markus, H.S.; Fornage, M.; Holliday, E.G.; et al. Shared genetic contribution to Ischaemic Stroke and Alzheimer’s Disease. Ann. Neurol. 2016, 79, 739–747. [Google Scholar] [CrossRef] [Green Version]
- Cui, P.; Ma, X.; Li, H.; Lang, W.; Hao, J. Shared Biological Pathways Between Alzheimer’s Disease and Ischemic Stroke. Front. Neurosci. 2018, 12, 605. [Google Scholar] [CrossRef]
- Wei, C.J.; Cui, P.; Li, H.; Lang, W.J.; Liu, G.Y.; Ma, X.F. Shared genes between Alzheimer’s disease and ischemic stroke. CNS Neurosci. Ther. 2019, 25, 855–864. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.R.; Islam, T.; Shahjaman, M.; Zaman, T.; Faruquee, H.M.; Jamal, M.; Huq, F.; Quinn, J.M.W.; Moni, M.A. Discovering biomarkers and pathways shared by Alzheimer’s disease and ischemic stroke to identify novel therapeutic targets. Medicina 2019, 55, 191. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Ni, Q.B.; Wang, K.; Han, Z.; Sun, B.L. Stroke and Alzheimer’s disease: A mendelian randomization study. Front. Genet. 2020, 11, 581. [Google Scholar] [CrossRef]
- Feske, S.K. Ischemic stroke. Am. J. Med. 2021, 134, 1457–1464. [Google Scholar] [CrossRef]
- Jack, C.R., Jr.; Bennett, D.A.; Blennow, K.; Carrillo, M.C.; Dunn, B.; Haeberlein, S.B.; Holtzman, D.M.; Jagust, W.; Jessen, F.; Karlawish, J.; et al. NIA-AA Research Framework: Toward a biological definition of Alzheimer’s disease. Alzheimers Dement. 2018, 14, 535–562. [Google Scholar] [CrossRef]
- Vemuri, P.; Lesnick, T.G.; Przybelski, S.A.; Knopman, D.S.; Lowe, V.J.; Graff-Radford, J.; Roberts, R.O.; Mielke, M.M.; Machulda, M.M.; Petersen, R.C.; et al. Age, vascular health, and Alzheimer disease biomarkers in an elderly sample. Ann. Neurol. 2017, 82, 706–718. [Google Scholar] [CrossRef] [PubMed]
- Govindpani, K.; McNamara, L.G.; Smith, N.R.; Vinnakota, C.; Waldvogel, H.J.; Faull, R.L.; Kwakowsky, A. Vascular dysfunction in Alzheimer’s iisease: A prelude to the pathological process or a consequence of it? J. Clin. Med. 2019, 8, 651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zupanic, E.; von Euler, M.; Winblad, B.; Xu, H.; Secnik, J.; Kramberger, M.G.; Religa, D.; Norrving, B.; Garcia-Ptacek, S. Mortality after ischemic stroke in patients with Alzheimer’s disease dementia and other dementia disorders. J. Alzheimers Dis. 2021, 81, 1253–1261. [Google Scholar] [CrossRef]
- Hoyer, S. Age as risk factor for sporadic dementia of the Alzheimer type? Ann. N. Y. Acad. Sci. 1994, 719, 248–256. [Google Scholar] [CrossRef] [PubMed]
- Hunter, E.; Kelleher, J.D. Age specific models to capture the change in risk factor contribution by age to short term primary ischemic stroke risk. Front. Neurol. 2022, 13, 803749. [Google Scholar] [CrossRef] [PubMed]
- Cummings, J.; Ortiz, A.; Castellino, J.; Kinney, J. Diabetes: Risk factor and translational therapeutic implications for Alzheimer’s disease. Eur. J. Neurosci. 2022, 56, 5727–5757. [Google Scholar] [CrossRef]
- Khoury, J.C.; Kleindorfer, D.; Alwell, K.; Moomaw, C.J.; Woo, D.; Adeoye, O.; Flaherty, M.L.; Khatri, P.; Ferioli, S.; Broderick, J.P.; et al. Diabetes mellitus: A risk factor for ischemic stroke in a large biracial population. Stroke 2013, 44, 1500–1504. [Google Scholar] [CrossRef] [Green Version]
- Lloret, A.; Monllor, P.; Esteve, D.; Cervera-Ferri, A.; Lloret, M.A. Obesity as a risk factor for Alzheimer’s disease: Implication of leptin and glutamate. Front. Neurosci. 2019, 13, 508. [Google Scholar] [CrossRef]
- Cong, X.; Liu, S.; Wang, W.; Ma, J.; Li, J. Combined consideration of body mass index and waist circumference identifies obesity patterns associated with risk of stroke in a Chinese prospective cohort study. BMC Public Health 2022, 22, 347. [Google Scholar] [CrossRef]
- Wang, R.; Qiu, C.; Dintica, C.S.; Shang, Y.; Calderon Larranaga, A.; Wang, H.X.; Xu, W. Shared risk and protective factors between Alzheimer’s disease and ischemic stroke: A population-based longitudinal study. Alzheimers Dement. 2021, 17, 191–204. [Google Scholar] [CrossRef]
- Wightman, D.P.; Jansen, I.E.; Savage, J.E.; Shadrin, A.A.; Bahrami, S.; Holland, D.; Rongve, A.; Borte, S.; Winsvold, B.S.; Drange, O.K.; et al. A genome-wide association study with 1,126,563 individuals identifies new risk loci for Alzheimer’s disease. Nat. Genet. 2021, 53, 1276–1282. [Google Scholar] [CrossRef] [PubMed]
- Malik, R.; Chauhan, G.; Traylor, M.; Sargurupremraj, M.; Okada, Y.; Mishra, A.; Rutten-Jacobs, L.; Giese, A.K.; van der Laan, S.W.; Gretarsdottir, S.; et al. Multiancestry genome-wide association study of 520,000 subjects identifies 32 loci associated with stroke and stroke subtypes. Nat. Genet. 2018, 50, 524–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armato, U.; Chiarini, A.; Chakravarthy, B.; Chioffi, F.; Pacchiana, R.; Colarusso, E.; Whitfield, J.F.; Dal Pra, I. Calcium-sensing receptor antagonist (calcilytic) NPS 2143 specifically blocks the increased secretion of endogenous Abeta42 prompted by exogenous fibrillary or soluble Abeta25-35 in human cortical astrocytes and neurons-therapeutic relevance to Alzheimer’s disease. Biochim. Biophys. Acta 2013, 1832, 1634–1652. [Google Scholar] [CrossRef] [PubMed]
- Mohmmad Abdul, H.; Wenk, G.L.; Gramling, M.; Hauss-Wegrzyniak, B.; Butterfield, D.A. APP and PS-1 mutations induce brain oxidative stress independent of dietary cholesterol: Implications for Alzheimer’s disease. Neurosci. Lett. 2004, 368, 148–150. [Google Scholar] [CrossRef]
- Ito, F.; Sono, Y.; Ito, T. Measurement and clinical significance of lipid peroxidation as a biomarker of oxidative stress: Oxidative stress in diabetes, atherosclerosis, and chronic inflammation. Antioxidants 2019, 8, 72. [Google Scholar] [CrossRef] [Green Version]
- Langness, V.F.; van der Kant, R.; Das, U.; Wang, L.; Chaves, R.D.S.; Goldstein, L.S.B. Cholesterol-lowering drugs reduce APP processing to Abeta by inducing APP dimerization. Mol. Biol. Cell 2021, 32, 247–259. [Google Scholar] [CrossRef]
- Fekih-Mrissa, N.; Klai, S.; Mrad, M.; Mansour, M.; Zaouali, J.; Gritli, N.; Mrissa, R. Apolipoprotein E genotypes associated with Alzheimer disease and concomitant stroke. J. Stroke Cerebrovasc. Dis. 2014, 23, 681–685. [Google Scholar] [CrossRef]
- Mahajan, A.; Taliun, D.; Thurner, M.; Robertson, N.R.; Torres, J.M.; Rayner, N.W.; Payne, A.J.; Steinthorsdottir, V.; Scott, R.A.; Grarup, N.; et al. Fine-mapping type 2 diabetes loci to single-variant resolution using high-density imputation and islet-specific epigenome maps. Nat. Genet. 2018, 50, 1505–1513. [Google Scholar] [CrossRef] [Green Version]
- Chitnis, T.; Banwell, B.; Kappos, L.; Arnold, D.L.; Gucuyener, K.; Deiva, K.; Skripchenko, N.; Cui, L.Y.; Saubadu, S.; Hu, W.; et al. Safety and efficacy of teriflunomide in paediatric multiple sclerosis (TERIKIDS): A multicentre, double-blind, phase 3, randomised, placebo-controlled trial. Lancet Neurol. 2021, 20, 1001–1011. [Google Scholar] [CrossRef]
- Patel, D.; Mez, J.; Vardarajan, B.N.; Staley, L.; Chung, J.; Zhang, X.; Farrell, J.J.; Rynkiewicz, M.J.; Cannon-Albright, L.A.; Teerlink, C.C.; et al. Association of Rare Coding Mutations with Alzheimer Disease and Other Dementias Among Adults of European Ancestry. JAMA Netw. Open 2019, 2, e191350. [Google Scholar] [CrossRef]
- Sassi, C.; Nalls, M.A.; Ridge, P.G.; Gibbs, J.R.; Lupton, M.K.; Troakes, C.; Lunnon, K.; Al-Sarraj, S.; Brown, K.S.; Medway, C.; et al. Mendelian adult-onset leukodystrophy genes in Alzheimer’s disease: Critical influence of CSF1R and NOTCH3. Neurobiol. Aging 2018, 66, 179.e17–179.e29. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Zhang, H.; Liu, B.; Ji, X. Rs2293871 regulates HTRA1 expression and affects cerebral small vessel stroke and Alzheimer’s disease. Brain 2019, 142, e61. [Google Scholar] [CrossRef] [PubMed]
- Schwartzentruber, J.; Cooper, S.; Liu, J.Z.; Barrio-Hernandez, I.; Bello, E.; Kumasaka, N.; Young, A.M.H.; Franklin, R.J.M.; Johnson, T.; Estrada, K.; et al. Genome-wide meta-analysis, fine-mapping and integrative prioritization implicate new Alzheimer’s disease risk genes. Nat. Genet. 2021, 53, 392–402. [Google Scholar] [CrossRef] [PubMed]
- Carty, C.L.; Keene, K.L.; Cheng, Y.C.; Meschia, J.F.; Chen, W.M.; Nalls, M.; Bis, J.C.; Kittner, S.J.; Rich, S.S.; Tajuddin, S.; et al. Meta-Analysis of Genome-Wide Association Studies Identifies Genetic Risk Factors for Stroke in African Americans. Stroke 2015, 46, 2063–2068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mez, J.; Chung, J.; Jun, G.; Kriegel, J.; Bourlas, A.P.; Sherva, R.; Logue, M.W.; Barnes, L.L.; Bennett, D.A.; Buxbaum, J.D.; et al. Two novel loci, COBL and SLC10A2, for Alzheimer’s disease in African Americans. Alzheimers Dement. 2017, 13, 119–129. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Chauhan, G.; Sharma, S.; Dabla, S.; Sylaja, P.N.; Chaudhary, N.; Gupta, S.; Agrawal, C.S.; Anand, K.S.; Srivastava, A.K.; et al. Association of SUMOylation Pathway Genes With Stroke in a Genome-Wide Association Study in India. Neurology 2021, 97, e345–e356. [Google Scholar] [CrossRef]
- Nazarian, A.; Yashin, A.I.; Kulminski, A.M. Genome-wide analysis of genetic predisposition to Alzheimer’s disease and related sex disparities. Alzheimers Res. Ther. 2019, 11, 5. [Google Scholar] [CrossRef] [Green Version]
- Chung, J.; Das, A.; Sun, X.; Sobreira, D.R.; Leung, Y.Y.; Igartua, C.; Mozaffari, S.; Chou, Y.F.; Thiagalingam, S.; Mez, J.; et al. Genome-wide association and multi-omics studies identify MGMT as a novel risk gene for Alzheimer’s disease among women. Alzheimers Dement. 2022. [Google Scholar] [CrossRef]
- Lee, T.H.; Ko, T.M.; Chen, C.H.; Chang, Y.J.; Lu, L.S.; Chang, C.H.; Huang, K.L.; Chang, T.Y.; Lee, J.D.; Chang, K.C.; et al. A genome-wide association study links small-vessel ischemic stroke to autophagy. Sci. Rep. 2017, 7, 15229. [Google Scholar] [CrossRef] [Green Version]
- Network, N.S.G.; International Stroke Genetics, C. Loci associated with ischaemic stroke and its subtypes (SiGN): A genome-wide association study. Lancet Neurol. 2016, 15, 174–184. [Google Scholar] [CrossRef]
- Malik, R.; Dau, T.; Gonik, M.; Sivakumar, A.; Deredge, D.J.; Edeleva, E.V.; Gotzfried, J.; van der Laan, S.W.; Pasterkamp, G.; Beaufort, N.; et al. Common coding variant in SERPINA1 increases the risk for large artery stroke. Proc. Natl. Acad. Sci. USA 2017, 114, 3613–3618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Traylor, M.; Malik, R.; Nalls, M.A.; Cotlarciuc, I.; Radmanesh, F.; Thorleifsson, G.; Hanscombe, K.B.; Langefeld, C.; Saleheen, D.; Rost, N.S.; et al. Genetic variation at 16q24.2 is associated with small vessel stroke. Ann. Neurol. 2017, 81, 383–394. [Google Scholar] [CrossRef] [Green Version]
- Traylor, M.; Persyn, E.; Tomppo, L.; Klasson, S.; Abedi, V.; Bakker, M.K.; Torres, N.; Li, L.; Bell, S.; Rutten-Jacobs, L.; et al. Genetic basis of lacunar stroke: A pooled analysis of individual patient data and genome-wide association studies. Lancet Neurol. 2021, 20, 351–361. [Google Scholar] [CrossRef]
- Jun, G.; Ibrahim-Verbaas, C.A.; Vronskaya, M.; Lambert, J.C.; Chung, J.; Naj, A.C.; Kunkle, B.W.; Wang, L.S.; Bis, J.C.; Bellenguez, C.; et al. A novel Alzheimer disease locus located near the gene encoding tau protein. Mol. Psychiatry 2016, 21, 108–117. [Google Scholar] [CrossRef] [Green Version]
- Kamboh, M.I.; Demirci, F.Y.; Wang, X.; Minster, R.L.; Carrasquillo, M.M.; Pankratz, V.S.; Younkin, S.G.; Saykin, A.J.; Alzheimer’s Disease Neuroimaging, I.; Jun, G.; et al. Genome-wide association study of Alzheimer’s disease. Transl. Psychiatry 2012, 2, e117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, J.; Zhang, X.; Allen, M.; Wang, X.; Ma, Y.; Beecham, G.; Montine, T.J.; Younkin, S.G.; Dickson, D.W.; Golde, T.E.; et al. Genome-wide pleiotropy analysis of neuropathological traits related to Alzheimer’s disease. Alzheimers Res. Ther. 2018, 10, 22. [Google Scholar] [CrossRef] [PubMed]
- D’Addario, C.; Candia, S.B.; Arosio, B.; Di Bartolomeo, M.; Abbate, C.; Case, A.; Candeletti, S.; Romualdi, P.; Damanti, S.; Maccarrone, M.; et al. Transcriptional and epigenetic phenomena in peripheral blood cells of monozygotic twins discordant for alzheimer’s disease, a case report. J. Neurol. Sci. 2017, 372, 211–216. [Google Scholar] [CrossRef]
- Lu, Y.; Tan, L.; Wang, X. Circular HDAC9/microRNA-138/Sirtuin-1 pathway mediates synaptic and amyloid precursor protein processing deficits in Alzheimer’s disease. Neurosci. Bull. 2019, 35, 877–888. [Google Scholar] [CrossRef]
- Lu, S.; Li, H.; Li, K.; Fan, X.D. HDAC9 promotes brain ischemic injury by provoking IkappaBalpha/NF-kappaB and MAPKs signaling pathways. Biochem. Biophys. Res. Commun. 2018, 503, 1322–1329. [Google Scholar] [CrossRef]
- Shi, J.; Liu, H.; Wang, H.; Kong, X. MicroRNA expression signature in degenerative aortic stenosis. Biomed. Res. Int. 2016, 2016, 4682172. [Google Scholar] [CrossRef]
- Zhang, H.; Mo, X.; Wang, A.; Peng, H.; Guo, D.; Zhong, C.; Zhu, Z.; Xu, T.; Zhang, Y. Association of DNA methylation in blood pressure-related genes with ischemic stroke risk and prognosis. Front. Cardiovasc. Med. 2022, 9, 796245. [Google Scholar] [CrossRef] [PubMed]
- Prestel, M.; Prell-Schicker, C.; Webb, T.; Malik, R.; Lindner, B.; Ziesch, N.; Rex-Haffner, M.; Roh, S.; Viturawong, T.; Lehm, M.; et al. The atherosclerosis risk variant rs2107595 mediates allele-specific transcriptional regulation of HDAC9 via E2F3 and Rb1. Stroke 2019, 50, 2651–2660. [Google Scholar] [CrossRef] [PubMed]
- Shroff, N.; Ander, B.P.; Zhan, X.; Stamova, B.; Liu, D.; Hull, H.; Hamade, F.R.; Dykstra-Aiello, C.; Ng, K.; Sharp, F.R.; et al. HDAC9 Polymorphism Alters Blood Gene Expression in Patients with Large Vessel Atherosclerotic Stroke. Transl. Stroke Res. 2019, 10, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Ito, S.; Kimura, K.; Haneda, M.; Ishida, Y.; Sawada, M.; Isobe, K. Induction of matrix metalloproteinases (MMP3, MMP12 and MMP13) expression in the microglia by amyloid-β stimulation via the PI3K/Akt pathway. Exp. Gerontol. 2007, 42, 532–537. [Google Scholar] [CrossRef]
- Asciutto, G.; Dias, N.V.; Edsfeldt, A.; Nitulescu, M.; Persson, A.; Nilsson, M.; Duner, P.; Nilsson, J.; Goncalves, I. Low elastin content of carotid plaques is associated with increased risk of ipsilateral stroke. PLoS ONE 2015, 10, e0121086. [Google Scholar] [CrossRef] [Green Version]
- Carcel-Marquez, J.; Cullell, N.; Muino, E.; Gallego-Fabrega, C.; Lledos, M.; Ibanez, L.; Krupinski, J.; Montaner, J.; Cruchaga, C.; Lee, J.M.; et al. Causal effect of MMP-1 (matrix metalloproteinase-1), MMP-8, and MMP-12 levels on ischemic stroke: A mendelian randomization study. Stroke 2021, 52, e316–e320. [Google Scholar] [CrossRef]
- Kuo, P.C.; Weng, W.T.; Scofield, B.A.; Furnas, D.; Paraiso, H.C.; Intriago, A.J.; Bosi, K.D.; Yu, I.C.; Yen, J.H. Interferon-β alleviates delayed tPA-induced adverse effects via modulation of MMP3/9 production in ischemic stroke. Blood Adv. 2020, 4, 4366–4381. [Google Scholar] [CrossRef]
- San Segundo-Acosta, P.; Montero-Calle, A.; Fuentes, M.; Rabano, A.; Villalba, M.; Barderas, R. Identification of Alzheimer’s Disease Autoantibodies and Their Target Biomarkers by Phage Microarrays. J. Proteome Res. 2019, 18, 2940–2953. [Google Scholar] [CrossRef]
- Han, J.; Feng, G.H.; Liu, H.W.; Yi, J.P.; Wu, J.B.; Yao, X.X. Classifying mild cognitive impairment and Alzheimer’s disease by constructing a 14-gene diagnostic model. Am. J. Transl. Res. 2022, 14, 4477–4492. [Google Scholar]
- Perisic, L.; Hedin, E.; Razuvaev, A.; Lengquist, M.; Osterholm, C.; Folkersen, L.; Gillgren, P.; Paulsson-Berne, G.; Ponten, F.; Odeberg, J.; et al. Profiling of atherosclerotic lesions by gene and tissue microarrays reveals PCSK6 as a novel protease in unstable carotid atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 2432–2443. [Google Scholar] [CrossRef]
- Alderton, G.; Scanlon, S.T. Inflammation. Science 2021, 374, 1068–1069. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Gaiteri, C.; Bodea, L.G.; Wang, Z.; McElwee, J.; Podtelezhnikov, A.A.; Zhang, C.; Xie, T.; Tran, L.; Dobrin, R.; et al. Integrated systems approach identifies genetic nodes and networks in late-onset Alzheimer’s disease. Cell 2013, 153, 707–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holmes, C.; Cunningham, C.; Zotova, E.; Woolford, J.; Dean, C.; Kerr, S.; Culliford, D.; Perry, V.H. Systemic inflammation and disease progression in Alzheimer disease. Neurology 2009, 73, 768–774. [Google Scholar] [CrossRef] [Green Version]
- Kinney, J.W.; Bemiller, S.M.; Murtishaw, A.S.; Leisgang, A.M.; Salazar, A.M.; Lamb, B.T. Inflammation as a central mechanism in Alzheimer’s disease. Alzheimers Dement. 2018, 4, 575–590. [Google Scholar] [CrossRef] [PubMed]
- Baik, S.H.; Kang, S.; Son, S.M.; Mook-Jung, I. Microglia contributes to plaque growth by cell death due to uptake of amyloid β in the brain of Alzheimer’s disease mouse model. Glia 2016, 64, 2274–2290. [Google Scholar] [CrossRef] [PubMed]
- Shippy, D.C.; Watters, J.J.; Ulland, T.K. Transcriptional response of murine microglia in Alzheimer’s disease and inflammation. BMC Genom. 2022, 23, 183. [Google Scholar] [CrossRef]
- Bostanciklioglu, M. An update on the interactions between Alzheimer’s disease, autophagy and inflammation. Gene 2019, 705, 157–166. [Google Scholar] [CrossRef]
- Yao Mattisson, I.; Rattik, S.; Bjorkbacka, H.; Ljungcrantz, I.; Terrinoni, M.; Lebens, M.; Holmgren, J.; Fredrikson, G.N.; Gullstrand, B.; Bengtsson, A.A.; et al. Immune responses against oxidized LDL as possible targets for prevention of atherosclerosis in systemic lupus erythematosus. Vascul. Pharmacol. 2021, 140, 106863. [Google Scholar] [CrossRef]
- Xu, K.; Saaoud, F.; Yu, S.; Drummer, C.t.; Shao, Y.; Sun, Y.; Lu, Y.; Sun, J.; Yu, J.; Jiang, X.; et al. Monocyte adhesion assays for detecting endothelial cell activation in vascular inflammation and atherosclerosis. Methods Mol. Biol. 2022, 2419, 169–182. [Google Scholar] [CrossRef]
- Esenwa, C.C.; Elkind, M.S. Inflammatory risk factors, biomarkers and associated therapy in ischaemic stroke. Nat. Rev. Neurol. 2016, 12, 594–604. [Google Scholar] [CrossRef]
- Calandra, T.; Bucala, R. Macrophage Migration Inhibitory Factor (MIF): A Glucocorticoid Counter-Regulator within the Immune System. Crit. Rev. Immunol. 2017, 37, 359–370. [Google Scholar] [CrossRef] [PubMed]
- Wei, N.; Xu, Y.; Li, Y.; Shi, J.; Zhang, X.; You, Y.; Sun, Q.; Zhai, H.; Hu, Y. A bibliometric analysis of T cell and atherosclerosis. Front. Immunol. 2022, 13, 948314. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Lu, Y.; Cao, Y.; Dang, C.; Wang, N.; Tian, K.; Luo, Q.; Guo, E.; Luo, S.; Wang, L.; et al. Identification of diagnostic signatures associated with immune infiltration in Alzheimer’s disease by integrating bioinformatic analysis and machine-learning strategies. Front. Aging Neurosci. 2022, 14, 919614. [Google Scholar] [CrossRef]
- Matoba, K. Monitoring Leukocyte Migration During Atherosclerosis In Vivo. Methods Mol. Biol. 2022, 2419, 475–479. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Li, J.; Cai, Y.; Liu, W.; Chen, M.; Xu, X.; Deng, M.; Sun, J.; Zhou, L.; Huang, Y.; et al. The Novel lncRNA ENST00000530525 Affects ANO1, Contributing to Blood-Brain Barrier Injury in Cultured hCMEC/D3 Cells Under OGD/R Conditions. Front. Genet. 2022, 13, 873230. [Google Scholar] [CrossRef]
- Lefkowitz, R.J. A brief history of G-protein coupled receptors (Nobel Lecture). Angew. Chem. Int. Ed. Engl. 2013, 52, 6366–6378. [Google Scholar] [CrossRef]
- Zhao, J.; Deng, Y.; Jiang, Z.; Qing, H. G protein-coupled receptors (GPCRs) in Alzheimer’s disease: A focus on BACE1 related GPCRs. Front. Aging Neurosci. 2016, 8, 58. [Google Scholar] [CrossRef] [Green Version]
- Haque, M.E.; Kim, I.S.; Jakaria, M.; Akther, M.; Choi, D.K. Importance of GPCR-Mediated Microglial Activation in Alzheimer’s Disease. Front. Cell Neurosci. 2018, 12, 258. [Google Scholar] [CrossRef] [Green Version]
- Dal Prà, I.; Armato, U.; Chiarini, A. Family C G-protein-coupled receptors in Alzheimer’s disease and therapeutic implications. Front. Pharmacol. 2019, 10, 1282. [Google Scholar] [CrossRef]
- Liao, X.; Zhang, Y.; Wang, Y.; Wang, J. The effect of cdk-5 overexpression on tau phosphorylation and spatial memory of rat. Sci. China C Life Sci. 2004, 47, 251–257. [Google Scholar] [CrossRef]
- Siano, G.; Caiazza, M.C.; Olla, I.; Varisco, M.; Madaro, G.; Quercioli, V.; Calvello, M.; Cattaneo, A.; Di Primio, C. Identification of an ERK inhibitor as a therapeutic drug against tau aggregation in a new cell-based assay. Front. Cell Neurosci. 2019, 13, 386. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, C.H.; Betts, J.C.; Blackstock, W.P.; Nebreda, A.R.; Anderton, B.H. Phosphorylation sites on tau identified by nanoelectrospray mass spectrometry: Differences in vitro between the mitogen-activated protein kinases ERK2, c-Jun N-terminal kinase and P38, and glycogen synthase kinase-3beta. J. Neurochem. 2000, 74, 1587–1595. [Google Scholar] [CrossRef] [PubMed]
- Bolos, M.; Llorens-Martin, M.; Perea, J.R.; Jurado-Arjona, J.; Rabano, A.; Hernandez, F.; Avila, J. Absence of CX3CR1 impairs the internalization of Tau by microglia. Mol. Neurodegener. 2017, 12, 59. [Google Scholar] [CrossRef] [Green Version]
- Thathiah, A.; De Strooper, B. G protein-coupled receptors, cholinergic dysfunction, and Abeta toxicity in Alzheimer’s disease. Sci. Signal. 2009, 2, re8. [Google Scholar] [CrossRef]
- Li, Y.F.; Li, R.S.; Samuel, S.B.; Cueto, R.; Li, X.Y.; Wang, H.; Yang, X.F. Lysophospholipids and their G protein-coupled receptors in atherosclerosis. Front. Biosci. 2016, 21, 70–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghislain, J.; Poitout, V. Targeting lipid GPCRs to treat type 2 diabetes mellitus—Progress and challenges. Nat. Rev. Endocrinol. 2021, 17, 162–175. [Google Scholar] [CrossRef] [PubMed]
- Jia, W.; Wang, C.; Zheng, J.; Li, Y.; Yang, C.; Wan, Q.L.; Shen, J. Pioglitazone Hydrochloride Extends the Lifespan of Caenorhabditis elegans by Activating DAF-16/FOXO- and SKN-1/NRF2-Related Signaling Pathways. Oxid. Med. Cell Longev. 2022, 2022, 8496063. [Google Scholar] [CrossRef] [PubMed]
- Blondrath, K.; Steel, J.H.; Katsouri, L.; Ries, M.; Parker, M.G.; Christian, M.; Sastre, M. The nuclear cofactor receptor interacting protein-140 (RIP140) regulates the expression of genes involved in Abeta generation. Neurobiol. Aging 2016, 47, 180–191. [Google Scholar] [CrossRef] [Green Version]
- Yi, Z.J.; Gong, J.P.; Zhang, W. Transcriptional co-regulator RIP140: An important mediator of the inflammatory response and its associated diseases (Review). Mol. Med. Rep. 2017, 16, 994–1000. [Google Scholar] [CrossRef] [Green Version]
- Das, B.C.; Dasgupta, S.; Ray, S.K. Potential therapeutic roles of retinoids for prevention of neuroinflammation and neurodegeneration in Alzheimer’s disease. Neural. Regen. Res. 2019, 14, 1880–1892. [Google Scholar] [CrossRef]
- Unsworth, A.J.; Flora, G.D.; Sasikumar, P.; Bye, A.P.; Sage, T.; Kriek, N.; Crescente, M.; Gibbins, J.M. RXR Ligands Negatively Regulate Thrombosis and Hemostasis. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 812–822. [Google Scholar] [CrossRef]
- Natunen, T.; Martiskainen, H.; Sarajarvi, T.; Helisalmi, S.; Pursiheimo, J.P.; Viswanathan, J.; Laitinen, M.; Makinen, P.; Kauppinen, T.; Rauramaa, T.; et al. Effects of NR1H3 genetic variation on the expression of liver X receptor α and the progression of Alzheimer’s disease. PLoS ONE 2013, 8, e80700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandoval-Hernandez, A.G.; Restrepo, A.; Cardona-Gomez, G.P.; Arboleda, G. LXR activation protects hippocampal microvasculature in very old triple transgenic mouse model of Alzheimer’s disease. Neurosci. Lett. 2016, 621, 15–21. [Google Scholar] [CrossRef]
- Grbic, E.; Gorkic, N.; Pleskovic, A.; Ljuca, F.; Gasparini, M.; Mrda, B.; Cilensek, I.; Fras, Z.; Petrovic, D. Association between the rs2279238 of the Liver X receptor α gene polymorphism and advanced carotid atherosclerosis in the Slovenian cohort. Gene 2022, 840, 146764. [Google Scholar] [CrossRef] [PubMed]
- Rasheed, A.; Shawky, S.A.; Tsai, R.; Jung, R.G.; Simard, T.; Saikali, M.F.; Hibbert, B.; Rayner, K.J.; Cummins, C.L. The secretome of liver X receptor agonist-treated early outgrowth cells decreases atherosclerosis in Ldlr-/- mice. Stem Cells Transl. Med. 2021, 10, 479–491. [Google Scholar] [CrossRef] [PubMed]
- Endo-Umeda, K.; Kim, E.; Thomas, D.G.; Liu, W.; Dou, H.; Yalcinkaya, M.; Abramowicz, S.; Xiao, T.; Antonson, P.; Gustafsson, J.A.; et al. Myeloid LXR (Liver X Receptor) Deficiency Induces Inflammatory Gene Expression in Foamy Macrophages and Accelerates Atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2022, 42, 719–731. [Google Scholar] [CrossRef]
- Chang, K.L.; Wong, L.R.; Pee, H.N.; Yang, S.; Ho, P.C. Reverting Metabolic Dysfunction in Cortex and Cerebellum of APP/PS1 Mice, a Model for Alzheimer’s Disease by Pioglitazone, a Peroxisome Proliferator-Activated Receptor γ (PPARgamma) Agonist. Mol. Neurobiol. 2019, 56, 7267–7283. [Google Scholar] [CrossRef]
- Cheng, F.; Si, X.M.; Yang, G.L.; Zhou, L. Relationship between PPAR-γ gene polymorphisms and ischemic stroke risk: A meta-analysis. Brain Behav. 2021, 11, e2434. [Google Scholar] [CrossRef]
- Sabry, M.M.; Dawood, A.F.; Rashed, L.A.; Sayed, S.M.; Hassan, S.; Younes, S.F. Relation between resistin, PPAR-γ, obesity and atherosclerosis in male albino rats. Arch. Physiol. Biochem. 2020, 126, 389–398. [Google Scholar] [CrossRef]
- Moon, M.; Jung, E.S.; Jeon, S.G.; Cha, M.Y.; Jang, Y.; Kim, W.; Lopes, C.; Mook-Jung, I.; Kim, K.S. Nurr1 (NR4A2) regulates Alzheimer’s disease-related pathogenesis and cognitive function in the 5XFAD mouse model. Aging Cell 2019, 18, e12866. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.G.; Tang, Y.; Tan, J.Z.; Wang, J.W.; Chen, G.J.; Zhu, B.L. The effect of NR4A1 on APP metabolism and tau phosphorylation. Genes Dis. 2018, 5, 342–348. [Google Scholar] [CrossRef] [PubMed]
- Hanna, R.N.; Shaked, I.; Hubbeling, H.G.; Punt, J.A.; Wu, R.; Herrley, E.; Zaugg, C.; Pei, H.; Geissmann, F.; Ley, K.; et al. NR4A1 (Nur77) deletion polarizes macrophages toward an inflammatory phenotype and increases atherosclerosis. Circ. Res. 2012, 110, 416–427. [Google Scholar] [CrossRef] [Green Version]
- Ishunina, T.A.; Bogolepova, I.N.; Swaab, D.F. Increased Neuronal Nuclear and Perikaryal Size in the Medial Mamillary Nucleus of Vascular Dementia and Alzheimer’s Disease Patients: Relation to Nuclear Estrogen Receptor α. Dement. Geriatr. Cogn. Disord. 2019, 47, 274–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Zhang, F.; Jiang, S.; Siedlak, S.L.; Shen, L.; Perry, G.; Wang, X.; Tang, B.; Zhu, X. Estrogen receptor-α is localized to neurofibrillary tangles in Alzheimer’s disease. Sci. Rep. 2016, 6, 20352. [Google Scholar] [CrossRef] [Green Version]
- Meng, Q.; Li, Y.; Ji, T.; Chao, Y.; Li, J.; Fu, Y.; Wang, S.; Chen, Q.; Chen, W.; Huang, F.; et al. Estrogen prevent atherosclerosis by attenuating endothelial cell pyroptosis via activation of estrogen receptor α-mediated autophagy. J. Adv. Res. 2021, 28, 149–164. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Chao, Y.; Zhang, W.; Zhang, Y.; Bi, Y.; Fu, Y.; Cai, D.; Meng, Q.; Li, Y.; Bian, H. Activation of estrogen receptor α (ERalpha) is required for Alisol B23-acetate to prevent post-menopausal atherosclerosis and reduced lipid accumulation. Life Sci. 2020, 258, 118030. [Google Scholar] [CrossRef]
- Min, J.; Weitian, Z.; Peng, C.; Yan, P.; Bo, Z.; Yan, W.; Yun, B.; Xukai, W. Correlation between insulin-induced estrogen receptor methylation and atherosclerosis. Cardiovasc. Diabetol. 2016, 15, 156. [Google Scholar] [CrossRef] [Green Version]
- Ji, X.; Jia, L.; Jia, J.; Qi, L. Genetic association of urokinase-type plasminogen activator gene rs2227564 site polymorphism with sporadic Alzheimer’s disease in the Han Chinese population. Neural. Regen. Res. 2012, 7, 2377–2383. [Google Scholar] [CrossRef]
- Diaz, A.; Merino, P.; Guo, J.D.; Yepes, M.A.; McCann, P.; Katta, T.; Tong, E.M.; Torre, E.; Rangaraju, S.; Yepes, M. Urokinase-Type Plasminogen Activator Protects Cerebral Cortical Neurons from Soluble Abeta-Induced Synaptic Damage. J. Neurosci. 2020, 40, 4251–4263. [Google Scholar] [CrossRef]
- Svensson, P.A.; Olson, F.J.; Hagg, D.A.; Ryndel, M.; Wiklund, O.; Karlstrom, L.; Hulthe, J.; Carlsson, L.M.; Fagerberg, B. Urokinase-type plasminogen activator receptor is associated with macrophages and plaque rupture in symptomatic carotid atherosclerosis. Int. J. Mol. Med. 2008, 22, 459–464. [Google Scholar]
- Paland, N.; Aharoni, S.; Fuhrman, B. Urokinase-type plasminogen activator (uPA) modulates monocyte-to-macrophage differentiation and prevents Ox-LDL-induced macrophage apoptosis. Atherosclerosis 2013, 231, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.D.; Kim, M.S. Exposure to a mixture of heavy metals induces cognitive impairment: Genes and microRNAs involved. Toxicology 2022, 471, 153164. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.D.; Kim, M.S. The role of mixed B vitamin intakes on cognitive performance: Modeling, genes and miRNAs involved. J. Psychiatr. Res. 2022, 152, 38–56. [Google Scholar] [CrossRef]
- Wen, Q.; Verheijen, M.; Wittens, M.M.J.; Czurylo, J.; Engelborghs, S.; Hauser, D.; van Herwijnen, M.H.M.; Lundh, T.; Bergdahl, I.A.; Kyrtopoulos, S.A.; et al. Lead-exposure associated miRNAs in humans and Alzheimer’s disease as potential biomarkers of the disease and disease processes. Sci. Rep. 2022, 12, 15966. [Google Scholar] [CrossRef]
- Nguyen, D.D.N.; Zain, S.M.; Kamarulzaman, M.H.; Low, T.Y.; Chilian, W.M.; Pan, Y.; Ting, K.N.; Hamid, A.; Abdul Kadir, A.; Pung, Y.F. Intracellular and exosomal microRNAome profiling of human vascular smooth muscle cells during replicative senescence. Am. J. Physiol. Heart Circ. Physiol. 2021, 321, H770–H783. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.D. Resveratrol, Endocrine Disrupting Chemicals, Neurodegenerative Diseases and Depression: Genes, Transcription Factors, microRNAs, and Sponges Involved. Neurochem. Res. 2022. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Li, L. circKMT2E Protect Retina from Early Diabetic Retinopathy through SIRT1 Signaling Pathway via Sponging miR-204-5p. Comput. Math Methods Med. 2022, 2022, 7188193. [Google Scholar] [CrossRef]
- Liu, X.; Wu, J.; Nie, H.; Zhu, X.; Song, G.; Han, L.; Qin, W. Comprehensive Analysis of circRNAs, miRNAs, and mRNAs Expression Profiles and ceRNA Networks in Decidua of Unexplained Recurrent Spontaneous Abortion. Front. Genet. 2022, 13, 858641. [Google Scholar] [CrossRef]
- Tang, Y.F.; Wu, W.J.; Zhang, J.Y.; Zhang, J. Reconstruction and analysis of the aberrant lncRNA-miRNA-mRNA network based on competitive endogenous RNA in adenoid cystic carcinoma of the salivary gland. Transl. Cancer Res. 2021, 10, 5133–5149. [Google Scholar] [CrossRef]
- Yang, S.; Zheng, W.; Yang, C.; Zu, R.; Ran, S.; Wu, H.; Mu, M.; Sun, S.; Zhang, N.; Thorne, R.F.; et al. Integrated Analysis of Hub Genes and MicroRNAs in Human Placental Tissues from In Vitro Fertilization-Embryo Transfer. Front. Endocrinol. (Lausanne) 2021, 12, 774997. [Google Scholar] [CrossRef]
- Zhang, F.; Yu, X.; Lin, Z.; Wang, X.; Gao, T.; Teng, D.; Teng, W. Using Tumor-Infiltrating Immune Cells and a ceRNA Network Model to Construct a Prognostic Analysis Model of Thyroid Carcinoma. Front. Oncol. 2021, 11, 658165. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, P.; Teng, X.; Zhang, Z.; Song, S. Comprehensive Analysis of Expression Regulation for RNA m6A Regulators With Clinical Significance in Human Cancers. Front. Oncol. 2021, 11, 624395. [Google Scholar] [CrossRef] [PubMed]
- Hao, M.; Liu, W.; Ding, C.; Peng, X.; Zhang, Y.; Chen, H.; Dong, L.; Liu, X.; Zhao, Y.; Chen, X.; et al. Identification of hub genes and small molecule therapeutic drugs related to breast cancer with comprehensive bioinformatics analysis. PeerJ 2020, 8, e9946. [Google Scholar] [CrossRef] [PubMed]
- Zhai, M.; Zhu, Y.; Yang, M.; Mao, C. Human Mesenchymal Stem Cell Derived Exosomes Enhance Cell-Free Bone Regeneration by Altering Their miRNAs Profiles. Adv. Sci. (Weinh) 2020, 7, 2001334. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Chen, D.; Xie, G.; Li, J.; Tang, J.; Tang, L. LncRNA-mediated ceRNA network was identified as a crucial determinant of differential effects in periodontitis and periimplantitis by high-throughput sequencing. Clin. Implant Dent. Relat. Res. 2020, 22, 424–450. [Google Scholar] [CrossRef]
- Ma, T.; Liu, A.; Xu, D.; Zhang, T. Mechanisms underlying the promotion of osteosarcoma cell proliferation and invasion by lncRNA PBB12. Oncol. Rep. 2020, 43, 736–746. [Google Scholar] [CrossRef]
- Kondo, Y.; Kogure, T.; Ninomiya, M.; Fukuda, R.; Monma, N.; Ikeo, K.; Tanaka, Y. The reduction of miR146b-5p in monocytes and T cells could contribute to the immunopathogenesis of hepatitis C virus infection. Sci. Rep. 2019, 9, 13393. [Google Scholar] [CrossRef] [Green Version]
- Fan, C.; Liu, X.; Li, W.; Wang, H.; Teng, Y.; Ren, J.; Huang, Y. Circular RNA circ KMT2E is up-regulated in diabetic cataract lenses and is associated with miR-204-5p sponge function. Gene 2019, 710, 170–177. [Google Scholar] [CrossRef]
- Gumus, G.; Giray, D.; Bobusoglu, O.; Tamer, L.; Karpuz, D.; Hallioglu, O. MicroRNA values in children with rheumatic carditis: A preliminary study. Rheumatol. Int. 2018, 38, 1199–1205. [Google Scholar] [CrossRef]
- Su, L.N.; Song, X.Q.; Xue, Z.X.; Zheng, C.Q.; Yin, H.F.; Wei, H.P. Network analysis of microRNAs, transcription factors, and target genes involved in axon regeneration. J. Zhejiang Univ. Sci. B 2018, 19, 293–304. [Google Scholar] [CrossRef]
- Gao, W.; Wu, Y.; He, X.; Zhang, C.; Zhu, M.; Chen, B.; Liu, Q.; Qu, X.; Li, W.; Wen, S.; et al. MicroRNA-204-5p inhibits invasion and metastasis of laryngeal squamous cell carcinoma by suppressing forkhead box C1. J. Cancer 2017, 8, 2356–2368. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Liu, C.; Huang, S.; Ma, Y.; Fang, J.; Chen, Y. A Downmodulated MicroRNA Profiling in Patients with Gastric Cancer. Gastroenterol. Res. Pract. 2017, 2017, 1526981. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Ma, X.; Zhang, Y.; Ni, D.; Ai, Q.; Li, H.; Zhang, X. Establishment of a miRNA-mRNA regulatory network in metastatic renal cell carcinoma and screening of potential therapeutic targets. Tumour. Biol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Hubens, W.H.G.; Krauskopf, J.; Beckers, H.J.M.; Kleinjans, J.C.S.; Webers, C.A.B.; Gorgels, T. Small RNA Sequencing of Aqueous Humor and Plasma in Patients With Primary Open-Angle Glaucoma. Investig. Ophthalmol. Vis. Sci. 2021, 62, 24. [Google Scholar] [CrossRef]
- Liu, S.; Wang, H.; Mu, J.; Wang, H.; Peng, Y.; Li, Q.; Mao, D.; Guo, L. MiRNA-211 triggers an autophagy-dependent apoptosis in cervical cancer cells: Regulation of Bcl-2. Naunyn. Schmiedebergs Arch. Pharmacol. 2020, 393, 359–370. [Google Scholar] [CrossRef]
- Zhong, Y.; Zhu, F.; Ding, Y. Differential microRNA expression profile in the plasma of preeclampsia and normal pregnancies. Exp. Ther. Med. 2019, 18, 826–832. [Google Scholar] [CrossRef]
- Shinawi, T.; Nasser, K.K.; Moradi, F.A.; Mujalli, A.; Albaqami, W.F.; Almukadi, H.S.; Elango, R.; Shaik, N.A.; Banaganapalli, B. A comparative mRNA- and miRNA transcriptomics reveals novel molecular signatures associated with metastatic prostate cancers. Front. Genet. 2022, 13, 1066118. [Google Scholar] [CrossRef]
- Dandare, A.; Rabia, G.; Khan, M.J. In silico analysis of non-coding RNAs and putative target genes implicated in metabolic syndrome. Comput. Biol. Med. 2021, 130, 104229. [Google Scholar] [CrossRef]
- Feng, C.; Li, Y.; Lin, Y.; Cao, X.; Li, D.; Zhang, H.; He, X. CircRNA-associated ceRNA network reveals ErbB and Hippo signaling pathways in hypopharyngeal cancer. Int. J. Mol. Med. 2019, 43, 127–142. [Google Scholar] [CrossRef] [Green Version]
- Jing, L.; Jin, C.; Lu, Y.; Huo, P.; Zhou, L.; Wang, Y.; Tian, Y. Investigation of microRNA expression profiles associated with human alcoholic cardiomyopathy. Cardiology 2015, 130, 223–233. [Google Scholar] [CrossRef]
- Li, S.; Song, F.; Lei, X.; Li, J.; Li, F.; Tan, H. hsa_circ_0004018 suppresses the progression of liver fibrosis through regulating the hsa-miR-660-3p/TEP1 axis. Aging (Albany N.Y.) 2020, 12, 11517–11529. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Zhang, J.; Zou, C.; Xie, X.; Wang, Y.; Wang, B.; Zhao, Z.; Tu, J.; Wang, X.; Li, H.; et al. Microarray Expression Profile and Functional Analysis of Circular RNAs in Osteosarcoma. Cell Physiol. Biochem. 2017, 43, 969–985. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Dou, J.; Cao, J.; Wang, Y.; Gao, Q.; Zeng, Q.; Liu, W.; Liu, B.; Cui, Z.; Teng, L.; et al. Uncovering the potential differentially expressed miRNAs as diagnostic biomarkers for hepatocellular carcinoma based on machine learning in The Cancer Genome Atlas database. Oncol. Rep. 2020, 43, 1771–1784. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.; Zhang, Y.; Yang, J. A Five-microRNA Signature as Prognostic Biomarker in Colorectal Cancer by Bioinformatics Analysis. Front. Oncol. 2019, 9, 1207. [Google Scholar] [CrossRef] [Green Version]
- Plata-Bello, J.; Farina-Jeronimo, H.; Betancor, I.; Salido, E. High Expression of FOXP2 Is Associated with Worse Prognosis in Glioblastoma. World Neurosurg. 2021, 150, e253–e278. [Google Scholar] [CrossRef]
- Wang, Y.; Li, Y.; Jia, D.; Zheng, J.; Wang, G. Correlation between single nucleotide polymorphisms in CXCR4 microRNA binding site and the susceptibility to knee osteoarthritis in Han Chinese population. J. Clin. Lab. Anal. 2021, 35, e23600. [Google Scholar] [CrossRef]
- Ekmekci, C.G.; Coskunpinar, E.; Avci, H.; Farooqi, A.A.; Orhan, K.S.; Akbas, F. Integrative analysis of mRNA and microRNA expression profiles in laryngeal squamous cell carcinoma. J. Cell Biochem. 2019, 120, 3415–3422. [Google Scholar] [CrossRef]
- Peng, Z.; Duan, Y.; Zhong, S.; Chen, J.; Li, J.; He, Z. RNA-seq analysis of extracellular vesicles from hyperphosphatemia-stimulated endothelial cells provides insight into the mechanism underlying vascular calcification. BMC Nephrol. 2022, 23, 192. [Google Scholar] [CrossRef]
- Chen, N.; Wan, G.; Zeng, X. Integrated Whole-Transcriptome Profiling and Bioinformatics Analysis of the Polypharmacological Effects of Ganoderic Acid Me in Colorectal Cancer Treatment. Front. Oncol. 2022, 12, 833375. [Google Scholar] [CrossRef] [PubMed]
- Baker, M.A.; Davis, S.J.; Liu, P.; Pan, X.; Williams, A.M.; Iczkowski, K.A.; Gallagher, S.T.; Bishop, K.; Regner, K.R.; Liu, Y.; et al. Tissue-Specific MicroRNA Expression Patterns in Four Types of Kidney Disease. J. Am. Soc. Nephrol. 2017, 28, 2985–2992. [Google Scholar] [CrossRef] [Green Version]
- Ju, L.; Han, M.; Li, X.; Zhao, C. MicroRNA Signature of Lung Adenocarcinoma with EGFR Exon 19 Deletion. J. Cancer 2017, 8, 1311–1318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoen, C.; Glennon, J.C.; Abghari, S.; Bloemen, M.; Aschrafi, A.; Carels, C.E.L.; Von den Hoff, J.W. Differential microRNA expression in cultured palatal fibroblasts from infants with cleft palate and controls. Eur. J. Orthod. 2018, 40, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Pehserl, A.M.; Ress, A.L.; Stanzer, S.; Resel, M.; Karbiener, M.; Stadelmeyer, E.; Stiegelbauer, V.; Gerger, A.; Mayr, C.; Scheideler, M.; et al. Comprehensive Analysis of miRNome Alterations in Response to Sorafenib Treatment in Colorectal Cancer Cells. Int. J. Mol. Sci. 2016, 17, 2011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, M.; Lu, L.; Dai, T.; Li, A.; Yu, Y.; Li, Y.; Xu, Z.; Chen, Y. Construction of a lncRNA-mediated ceRNA network and a genomic-clinicopathologic nomogram to predict survival for breast cancer patients. Cancer Biomark. 2022. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Zhou, S.; Guo, W. Construction of the coexpression network involved in the pathogenesis of thyroid eye disease via bioinformatics analysis. Hum. Genom. 2022, 16, 38. [Google Scholar] [CrossRef]
- Li, S.; Zhang, M.; Xu, F.; Wang, Y.; Leng, D. Detection significance of miR-3662, miR-146a, and miR-1290 in serum exosomes of breast cancer patients. J. Cancer Res. Ther. 2021, 17, 749–755. [Google Scholar] [CrossRef]
- Liu, A.; Zhou, Y.; Zhao, T.; Tang, X.; Zhou, B.; Xu, J. MiRNA-3662 reverses the gemcitabine resistance in pancreatic cancer through regulating the tumor metabolism. Cancer Chemother. Pharmacol. 2021, 88, 343–357. [Google Scholar] [CrossRef]
- Liu, A.; Xu, J. Circ_03955 promotes pancreatic cancer tumorigenesis and Warburg effect by targeting the miR-3662/HIF-1alpha axis. Clin. Transl. Oncol. 2021, 23, 1905–1914. [Google Scholar] [CrossRef]
- Byun, Y.J.; Piao, X.M.; Jeong, P.; Kang, H.W.; Seo, S.P.; Moon, S.K.; Lee, J.Y.; Choi, Y.H.; Lee, H.Y.; Kim, W.T.; et al. Urinary microRNA-1913 to microRNA-3659 expression ratio as a non-invasive diagnostic biomarker for prostate cancer. Investig. Clin. Urol. 2021, 62, 340–348. [Google Scholar] [CrossRef]
- Ye, J.; Xiao, X.; Han, Y.; Fan, D.; Zhu, Y.; Yang, L. MiR-3662 suppresses cell growth, invasion and glucose metabolism by targeting HK2 in hepatocellular carcinoma cells. Neoplasma 2020, 67, 773–781. [Google Scholar] [CrossRef]
- Niveditha, D.; Jasoria, M.; Narayan, J.; Majumder, S.; Mukherjee, S.; Chowdhury, R.; Chowdhury, S. Common and Unique microRNAs in Multiple Carcinomas Regulate Similar Network of Pathways to Mediate Cancer Progression. Sci. Rep. 2020, 10, 2331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Zuo, X.; Zhang, Y.; Han, G.; Zhang, L.; Wu, J.; Wang, X. MiR-3662 suppresses hepatocellular carcinoma growth through inhibition of HIF-1alpha-mediated Warburg effect. Cell Death Dis. 2018, 9, 549. [Google Scholar] [CrossRef] [PubMed]
- Yasui, K.; Izumida, M.; Nakagawa, T.; Kubo, Y.; Hayashi, H.; Ito, T.; Ikeda, H.; Matsuyama, T. MicroRNA-3662 expression correlates with antiviral drug resistance in adult T-cell leukemia/lymphoma cells. Biochem. Biophys. Res. Commun. 2018, 501, 833–837. [Google Scholar] [CrossRef] [PubMed]
- Powrozek, T.; Kuznar-Kaminska, B.; Dziedzic, M.; Mlak, R.; Batura-Gabryel, H.; Sagan, D.; Krawczyk, P.; Milanowski, J.; Malecka-Massalska, T. The diagnostic role of plasma circulating precursors of miRNA-944 and miRNA-3662 for non-small cell lung cancer detection. Pathol. Res. Pract. 2017, 213, 1384–1387. [Google Scholar] [CrossRef]
- Powrozek, T.; Mlak, R.; Dziedzic, M.; Malecka-Massalska, T.; Sagan, D. Analysis of primary-miRNA-3662 and its mature form may improve detection of the lung adenocarcinoma. J. Cancer Res. Clin. Oncol. 2017, 143, 1941–1946. [Google Scholar] [CrossRef]
- Zhao, W.; Shen, W.W.; Cao, X.M.; Ding, W.Y.; Yan, L.P.; Gao, L.J.; Li, X.L.; Zhong, T.Y. Novel mechanism of miRNA-365-regulated trophoblast apoptosis in recurrent miscarriage. J. Cell Mol. Med. 2017, 21, 2412–2425. [Google Scholar] [CrossRef]
- Maharry, S.E.; Walker, C.J.; Liyanarachchi, S.; Mehta, S.; Patel, M.; Bainazar, M.A.; Huang, X.; Lankenau, M.A.; Hoag, K.W.; Ranganathan, P.; et al. Dissection of the Major Hematopoietic Quantitative Trait Locus in Chromosome 6q23.3 Identifies miR-3662 as a Player in Hematopoiesis and Acute Myeloid Leukemia. Cancer Discov. 2016, 6, 1036–1051. [Google Scholar] [CrossRef] [Green Version]
- Powrozek, T.; Krawczyk, P.; Kowalski, D.M.; Winiarczyk, K.; Olszyna-Serementa, M.; Milanowski, J. Plasma circulating microRNA-944 and microRNA-3662 as potential histologic type-specific early lung cancer biomarkers. Transl. Res. 2015, 166, 315–323. [Google Scholar] [CrossRef]
- Li, Z.; Sun, Y.; He, M.; Liu, J. Differentially-expressed mRNAs, microRNAs and long noncoding RNAs in intervertebral disc degeneration identified by RNA-sequencing. Bioengineered 2021, 12, 1026–1039. [Google Scholar] [CrossRef]
- Chen, W.; Tang, D.; Dai, Y.; Diao, H. Establishment of microRNA, transcript and protein regulatory networks in Alport syndrome induced pluripotent stem cells. Mol. Med. Rep. 2019, 19, 238–250. [Google Scholar] [CrossRef] [Green Version]
- Lan, X.; Han, J.; Wang, B.; Sun, M. Integrated analysis of transcriptome profiling of lncRNAs and mRNAs in livers of type 2 diabetes mellitus. Physiol. Genom. 2022, 54, 86–97. [Google Scholar] [CrossRef] [PubMed]
- Tarek, M.M.; Yahia, A.; El-Nakib, M.M.; Elhefnawi, M. Integrative assessment of CIP2A overexpression and mutational effects in human malignancies identifies possible deleterious variants. Comput. Biol. Med. 2021, 139, 104986. [Google Scholar] [CrossRef] [PubMed]
- Morenikeji, O.B.; Adegbaju, M.S.; Okoh, O.S.; Babalola, A.E.; Grytsay, A.; Braimah, O.A.; Akinyemi, M.O.; Thomas, B.N. Deciphering inhibitory mechanism of coronavirus replication through host miRNAs-RNA-dependent RNA polymerase interactome. Front. Genet. 2022, 13, 973252. [Google Scholar] [CrossRef] [PubMed]
- Barbier, R.H.; McCrea, E.M.; Lee, K.Y.; Strope, J.D.; Risdon, E.N.; Price, D.K.; Chau, C.H.; Figg, W.D. Abiraterone induces SLCO1B3 expression in prostate cancer via microRNA-579-3p. Sci. Rep. 2021, 11, 10765. [Google Scholar] [CrossRef]
- Hommers, L.G.; Richter, J.; Yang, Y.; Raab, A.; Baumann, C.; Lang, K.; Schiele, M.A.; Weber, H.; Wittmann, A.; Wolf, C.; et al. A functional genetic variation of SLC6A2 repressor hsa-miR-579-3p upregulates sympathetic noradrenergic processes of fear and anxiety. Transl. Psychiatry 2018, 8, 226. [Google Scholar] [CrossRef] [Green Version]
- Kasimanickam, V.; Kastelic, J. MicroRNA in sperm from Duroc, Landrace and Yorkshire boars. Sci. Rep. 2016, 6, 32954. [Google Scholar] [CrossRef] [Green Version]
- Yilmaz, S.G.; Geyik, S.; Neyal, A.M.; Soko, N.D.; Bozkurt, H.; Dandara, C. Hypothesis: Do miRNAs Targeting the Leucine-Rich Repeat Kinase 2 Gene (LRRK2) Influence Parkinson’s Disease Susceptibility? OMICS 2016, 20, 224–228. [Google Scholar] [CrossRef]
- Ulusan Bagci, O.; Caner, A. [miRNA Expression Profile in Ileocecal Adenocarcinoma Cells Infected with Cryptosporidium]. Mikrobiyol. Bul. 2022, 56, 449–465. [Google Scholar] [CrossRef]
- Tripathi, S.K.; Mathaiyan, J.; Kayal, S.; Nachiappa Ganesh, R. Identification of Differentially Expressed Mirna by Next Generation Sequencing in Locally Advanced Breast Cancer Patients of South Indian Origin. Asian Pac. J. Cancer Prev. 2022, 23, 2255–2261. [Google Scholar] [CrossRef]
- Jeong, S.; Kim, S.A.; Ahn, S.G. HOXC6-Mediated miR-188-5p Expression Induces Cell Migration through the Inhibition of the Tumor Suppressor FOXN2. Int. J. Mol. Sci. 2021, 23, 9. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mapped Gene | Region | SNP | Position Relative to Gene | Risk Allele | p Value | Risk to | Reference |
|---|---|---|---|---|---|---|---|
| ALDH1A2 | 15q21.3 | rs4775044 | intron variant | N.A | 4.0 × 10−6 | AD | Schwartzentruber et al., 2021 [33] |
| rs4471613 | intron variant | A | 5.0 × 10−7 | IS | Carty et al., 2015 [34] | ||
| ANKRD22 | 10q23.31 | rs147285445 | intron variant | T | 4.0 × 10−6 | AD | Mez et al., 2017 [35] |
| rs11202867 | intron variant | N.A | 1.0 × 10−5 | IS | Kumar et al., 2021 [36] | ||
| ANTXR1 | 2p13.3 | rs7561207 | intron variant | N.A | 4.0 × 10−6 | AD | Nazarian et al., 2019 [37] |
| rs149587156 | intergenic variant | N.A | 5.0 × 10−6 | IS | Kumar et al., 2021 [36] | ||
| DIO2-AS1 | 14q31.1 | rs7155666 | intron variant | A | 8.0 × 10−10 | AD | Chung et al., 2022 [38] |
| rs11846182 | intron variant | T | 9.0 × 10−6 | IS | Lee et al., 2017 [39] | ||
| HDAC9 | 7p21.1 | rs117756856 | intron variant | A | 9.0 × 10−6 | AD | Mez et al., 2017 [35] |
| rs11984041 | intron variant | T | 5.0 × 10−9 | IS | Network NSG. 2016 [40] | ||
| rs2023938 | 3’prime UTR variant | G | 8.0 × 10−7 | IS | Malik et al., 2017 [41] | ||
| rs71524263 | intron variant | N.A | 2.0 × 10−12 | IS | Traylor et al., 2017 [42] | ||
| rs2107595 | regulatory region variant | N.A | 4.0 × 10−15 | IS | Malik et al., 2018 [22] | ||
| JPH3 | 16q24.2 | rs117760708 | non coding transcript exon variant | T | 3.0 × 10−6 | AD | Mez et al., 2017 [35] |
| rs12445022 | intergenic variant | A | 9.0 × 10−8 | IS | Traylor et al., 2021 [43] | ||
| KCNN3 | 1q21.3 | rs16830122 | intron variant | A | 2.0 × 10−6 | AD | Jun et al., 2016 [44] |
| rs114812453 | intergenic variant | N.A | 3.0 × 10−7 | IS | Kumar et al., 2021 [36] | ||
| LNC-LBCS | 6p22.3 | rs62402815 | intron variant | N.A | 2.0 × 10−6 | AD | Nazarian et al., 2019 [37] |
| rs9348394 | intron variant | N.A | 5.0 × 10−6 | IS | Kumar et al., 2021 [36] | ||
| MMP12 | 11q22.2 | rs12808148 | intergenic variant | N.A | 1.0 × 10−6 | AD | Kamboh et al., 2012 [45] |
| rs72983521 | intergenic variant | N.A | 3.0 × 10−8 | IS | Malik et al., 2018 [22] | ||
| MMP3 | 11q22.2 | rs12808148 | intergenic variant | N.A | 1.0 × 10−6 | AD | Kamboh et al., 2012 [45] |
| rs72983521 | intergenic variant | N.A | 5.0 × 10−8 | IS | Malik et al., 2018 [22] | ||
| PCSK6 | 15q26.3 | rs146322114 | intron variant | A | 2.0 × 10−6 | AD | Mez et al., 2017 [35] |
| rs528002287 | intron variant | N.A | 6.0 × 10−6 | IS | Kumar et al., 2021 [36] | ||
| RBMS3 | 3p24.1 | rs17022021 | intron variant | T | 8.0 × 10−6 | AD | Mez et al., 2017 [35] |
| rs115182009 | intron variant | N.A | 9.0 × 10−7 | IS | Kumar et al., 2021 [36] | ||
| RNU6-909P | 5p14.1 | rs150631144 | intron variant | T | 1.0 × 10−7 | AD | Mez et al., 2017 [35] |
| rs13354619 | intron variant | N.A | 2.0 × 10−6 | IS | Kumar et al., 2021 [36] |
| No. | Reference | AD and IS | Common Molecular Pathways Involved | Others | |
|---|---|---|---|---|---|
| NoSG | NoSS | ||||
| 1 | Traylor et al. 2016 [4] | 0 | 0 | Phospholipid efflux, cholesterol efflux, reverse cholesterol transport, negative regulation of nuclear factor kappa B (NF-κB) transcription factor activity (AD and small vessel disease) | One region (ATP5H/KCTD2/ICT1) associated with both AD and small vessel disease |
| 2 | Cui et al. 2018 [5] | / | / | Glioma, toll-like receptor signaling pathway, non-small cell lung cancer, natural killer cell mediated cytotoxicity, phospholipase D signaling pathway, hepatitis B, cadherin signaling pathway, wnt signaling pathway, immunoregulatory interactions between a lymphoid and a non-lymphoid cell, synthesis of PIPs at the plasma membrane, cooperation of PDCL (PhLP1) and TRiC/CCT in G-protein beta folding, PI metabolism, signaling pathways in glioblastoma | 56 biological processes, 95 cellular components, and 28 molecular functions |
| 3 | Wei et al. 2019 [6] | 16 | / | Immune system | / |
| 4 | Rahman MR et al. 2019 [7] | 22 | / | Alcoholism, MAPK signaling, glycine metabolism, serine metabolism, threonine metabolism | Transcriptional regulator: SPIB, SMAD3, and SOX2 |
| 5 | Wang T, et al.2020 [8] | 0 | 0 | Different types of stroke, including any stroke, any ischemic stroke, large artery stroke, and cardio-embolic stroke would not be causally associated with AD risk | / |
| 6 | This study | 13 | 0 | MIF mediated glucocorticoid regulation, transendothelial migration of leukocytes, GPCR pathway, signal transduction, signaling by nuclear receptors, ESR-mediated signaling, ethanol effects on histone modifications, urokinase-type plasminogen activator (uPA) and uPAR-mediated signaling, UPA-UPAR pathway | 23 miRNAs regulate more than seven common risk genes |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, W.; Huang, Y. Common Genetic Factors and Pathways in Alzheimer’s Disease and Ischemic Stroke: Evidences from GWAS. Genes 2023, 14, 353. https://doi.org/10.3390/genes14020353
Dong W, Huang Y. Common Genetic Factors and Pathways in Alzheimer’s Disease and Ischemic Stroke: Evidences from GWAS. Genes. 2023; 14(2):353. https://doi.org/10.3390/genes14020353
Chicago/Turabian StyleDong, Wei, and Yue Huang. 2023. "Common Genetic Factors and Pathways in Alzheimer’s Disease and Ischemic Stroke: Evidences from GWAS" Genes 14, no. 2: 353. https://doi.org/10.3390/genes14020353