
Genetic Diversity, Conservation, and Utilization of Plant Genetic Resources


Abstract
:1. Introduction
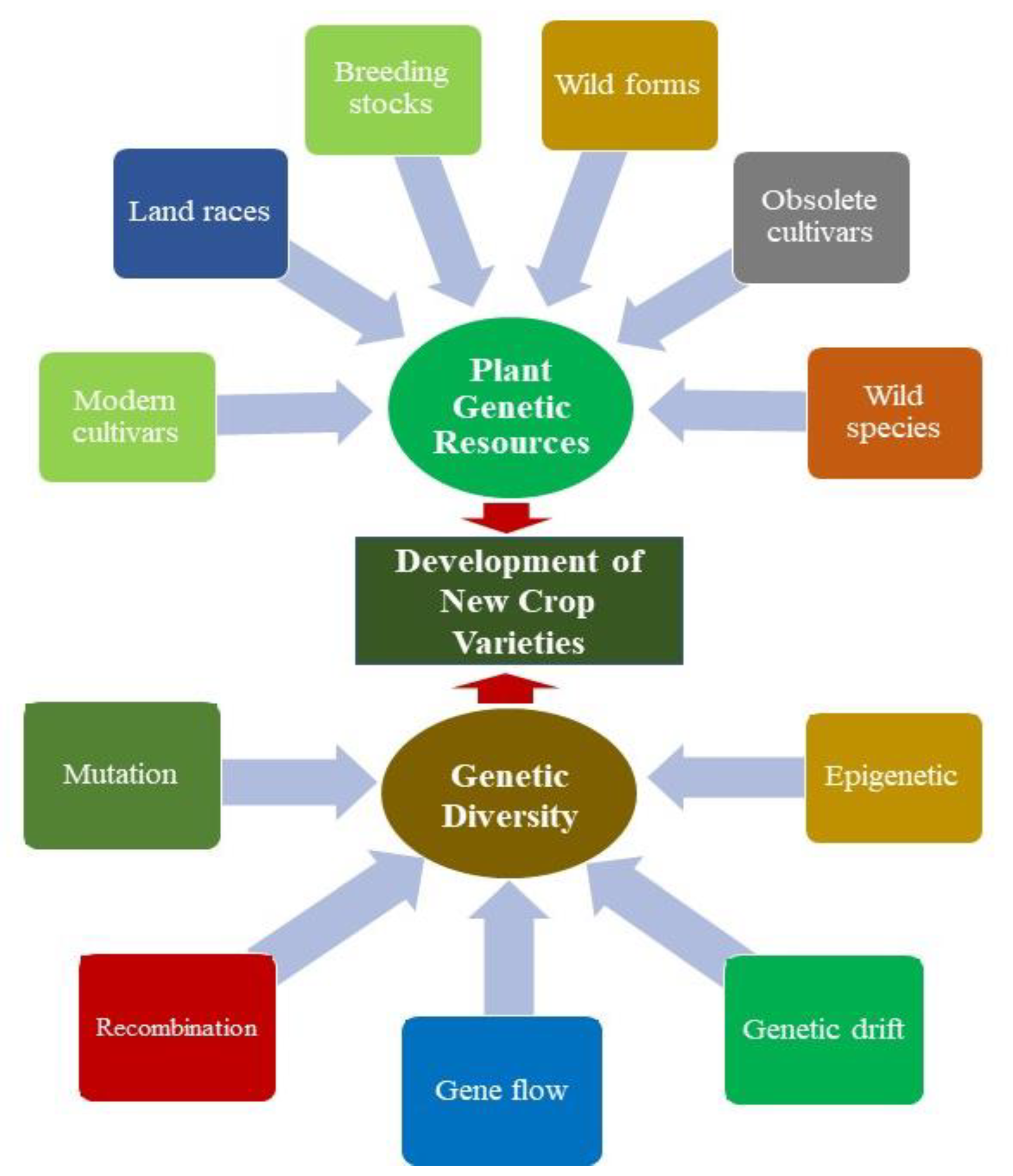
2. Importance of Genetic Diversity in Plant Genetic Resources
3. Factors Affecting Genetic Diversity
3.1. Mutation
3.2. Selection
3.3. Migration
3.4. Genetic Drift
4. Factors That Cause Genetic Vulnerability
4.1. Narrow Genetic Base of Crop Varieties
4.2. Wide Spread of Dominant Varieties
4.3. Unplanned Introduction of New Plant Species
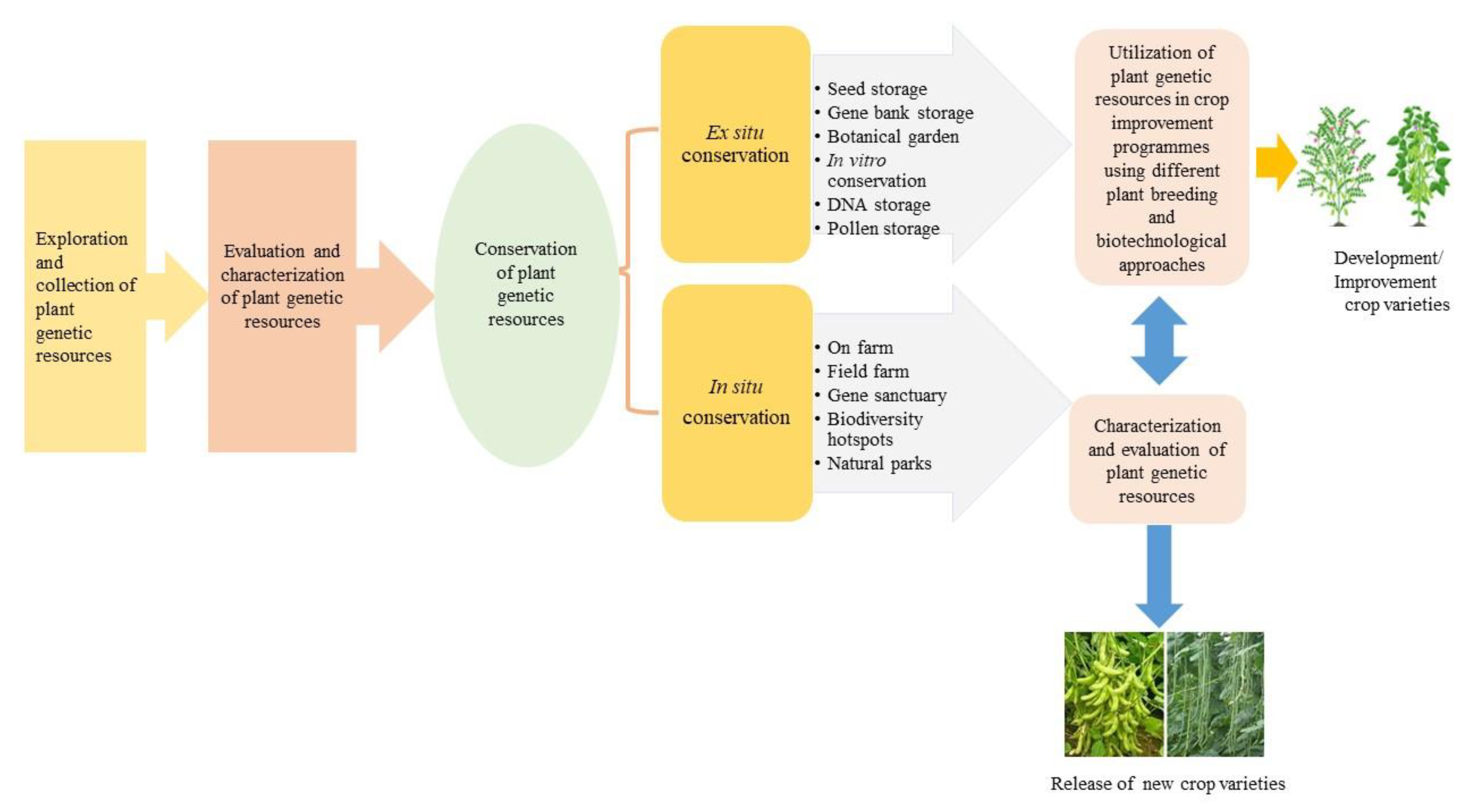
5. Conservation of Plant Genetic Resources
5.1. International Treaty on Plant Genetic Resources for Food and Agriculture
5.2. Nagoya Protocol
5.3. Svalbard Global Seed Vault
5.4. The Cartagena Protocol on Biosafety
5.5. In-Situ Conservation
5.6. Ex-Situ Conservation
5.7. Biotechnological Approaches
5.7.1. In Vitro Propagation
5.7.2. Cryopreservation
5.7.3. DNA Banks
5.7.4. Digital Sequence Information
6. Utilization of Plant Genetic Resources in Crop Improvement
Genomic Tools for Efficient Use of Plant Genetic Resources
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Brown, W.L. Genetic diversity and genetic vulnerability—An appraisal. Econ. Bot. 1983, 37, 4–12. [Google Scholar] [CrossRef]
- Nevo, E.; Beiles, A.; Ben-Shlomo, R. The evolutionary significance of genetic diversity: Ecological, demographic and life history correlates. Lect. Notes Biomath. 1984, 53, 13–21. [Google Scholar]
- Hasan, M.; Abdullah, H.M. Plant genetic resources and indigenous knowledge: An emerging needs for conservation. In Plant Genetic Resources and Traditional Knowledge for Food Security; Salgotra, R.K., Gupta, B.B., Eds.; Springer: Berlin/Heidelberg, Germany, 2015. [Google Scholar] [CrossRef]
- Ogwu, M.C.; Osawaru, M.E.; Ahana, C.M. Challenges in conserving and utilizing plant genetic resources (PGR). Int. J. Genet. Mol. Biol. 2014, 6, 16–22. [Google Scholar] [CrossRef] [Green Version]
- Ulukan, H. Plant genetic resources and breeding: Current scenario and future prospects. Int. J. Agric. Biol. 2011, 13, 447–454. [Google Scholar]
- Hyten, D.L.; Smith, J.R.; Frederick, R.D.; Tucker, M.L.; Song, Q.; Cregan, P.B. Bulked segregant analysis using the GoldenGate assay to locate the Rpp3 locus that confers resistance to soybean rust in soybean. Crop Sci. 2009, 49, 265–327. [Google Scholar] [CrossRef]
- Tanksley, S.D.; McCouch, S.R. Seed banks and molecular maps: Unlocking genetic potential from the wild. Science 1997, 277, 1063–1066. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, J.C.; Huber, C.D. The inflated significance of neutral genetic diversity in conservation genetics. Biol. Sci. 2021, 118, e2015096118. [Google Scholar] [CrossRef]
- Leipzig Declaration. International Technical Conference on Plant Genetic Resources, Leipzig. 1996. Available online: https://link.springer.com/chapter/10.1007/978-3-662-05212-9_2 (accessed on 7 June 2022).
- Varaprasad, K.S.; Sivaraj, N. Plant genetic resources conservation and use in light of recent policy developments. J. Plant Breed. 2010, 1, 1276–1293. [Google Scholar]
- Hammer, K.; Teklu, Y. Plant genetic resources: Selected issues from genetic erosion to genetic engineering. J. Agric. Rural Dev. Trop. Subtrop. 2010, 109, 15–50. [Google Scholar]
- IUPGR. International Undertaking on Plant Genetic Resources for Food and Agriculture. 1983. Available online: http://www.fao.org/ag/cgrfa/IU.htm (accessed on 7 June 2022).
- Bhandari, H.R.; Bhanu, A.N.; Srivastava, K.; Singh, M.N.; Shreya, H.A. Assessment of genetic diversity in crop plants—An overview. Adv. Plants Agric. Res. 2017, 7, 279–286. [Google Scholar] [CrossRef]
- Govindaraj, M.; Vetriventhan, M.; Srinivasan, M. Importance of genetic diversity assessment in crop plants and its recent advances: An overview of its analytical perspectives. Genet. Res. Int. 2015, 2015, 431487. [Google Scholar] [CrossRef] [Green Version]
- Wernberg, T.; Coleman, M.A.; Bennett, S.; Thomsen, M.S.; Tuya, F.; Kelaher, B.P. Genetic diversity and kelp forest vulnerability to climatic stress. Sci. Rep. 2018, 8, 1851. [Google Scholar] [CrossRef] [Green Version]
- Hoban, S.; Campbell, C.D.; da Silva, J.M.; Ekblom, R.; Funk, W.C.; Garner, B.A.; Godoy, J.A.; Kershaw, F.; MacDonald, A.J.; Mergeay, J.; et al. Genetic diversity is considered important but interpreted narrowly in country reports to the Convention on Biological Diversity: Current actions and indicators are insufficient. Biol. Conserv. 2021, 261, 109233. [Google Scholar] [CrossRef]
- Hughes, A.R.; Inouye, B.D.; Johnson, M.T.J.; Underwood, N.; Vellend, M. Ecological consequences of genetic diversity. Ecol. Lett. 2008, 11, 609–623. [Google Scholar] [CrossRef]
- Raffard, A.; Santoul, F.; Cucherousset, J.; Blanchet, S. The community and ecosystem consequences of intraspecific diversity: A meta-analysis. Biol. Rev. 2019, 94, 648–661. [Google Scholar] [CrossRef]
- Swingland, I.R. Biodiversity, Definition of. In Encyclopedia of Biodiversity, 2nd ed.; Levin, S.A., Ed.; Academic Press: Cambridge, MA, USA, 2013; pp. 399–410. [Google Scholar] [CrossRef]
- Primack, R.B. Essentials of Conservation Biology; Sinauer Associates, Inc.: Sunderland, MA, USA, 1993. [Google Scholar]
- Ashry, N.A. Plant Biodiversity and Biotechnology; Poltronieri, P., Burbulis, N., Fogher, C., Eds.; Woodhead Publishing Series in Biomedicine, From Plant Genomics to Plant Biotechnology; Woodhead Publishing: Sawston, UK, 2013; pp. 205–222. [Google Scholar] [CrossRef]
- Begna, T. Role and economic importance of crop genetic diversity in food security. J. Agric. Sci. Food Technol. 2021, 7, 164–169. [Google Scholar] [CrossRef]
- Ray, D.K.; Gerber, J.S.; MacDonald, G.K.; West, P.C. Climate variation explains a third of global crop yield variability. Nat. Commun. 2015, 6, 5989. [Google Scholar] [CrossRef] [Green Version]
- Acquaah, G. Principles of Plant Genetics and Breeding; John Wiley & Sons: Hoboken, NJ, USA, 2009. [Google Scholar]
- Hintum, T.V.; Jl, T. Hierarchical approaches to the analysis of genetic diversity in crop plants. In Core Collections of Plant Genetic Resources; Bioversity International: Rome, Italy, 1995. [Google Scholar]
- Ellstrand, N.C.; Prentice, H.C.; Hancock, J.F. Gene flow and introgression from domesticated plants into their wild relatives. Ann. Rev. Ecol. Syst. 1999, 30, 539–563. [Google Scholar] [CrossRef]
- Salgotra, R.K.; Thompson, M.; Chauhan, B.S. Unravelling the genetic potential of untapped crop wild genetic resources for crop improvement. Conserv. Genet. Res. 2021, 14, 109–124. [Google Scholar] [CrossRef]
- Ross-Ibarra, J.; Morrell, P.L.; Gaut, B.S. Plant domestication, a unique opportunity to identify the genetic basis of adaptation. Proc. Natl. Acad. Sci. USA 2007, 104, 8641–8648. [Google Scholar] [CrossRef] [Green Version]
- Begna, T. Importance and impact of ecological approaches to crop domestication. J. Biol. Agric. Healthc. 2020, 10, 32–37. [Google Scholar]
- Rauf, S.; da Silva, J.T.; Khan, A.A.; Naveed, A. Consequences of plant breeding on genetic diversity. Int. J. Plant Breed. 2010, 4, 1–21. [Google Scholar]
- McCouch, S. Diversifying selection in plant breeding. PLoS Biol. 2004, 2, e347. [Google Scholar] [CrossRef]
- Smith, B.D. Origins of agriculture in Eastern North America. Science 1989, 246, 1566–1571. [Google Scholar] [CrossRef]
- Oladosu, Y.; Rafii, M.Y.; Abdullah, N.; Hussin, G.; Ramli, A.; Rahim, H.A.; Miah, G.; Usman, M. Principle and application of plant mutagenesis in crop improvement: A review. Biotechnol. Biotechnol. Equip. 2016, 30, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Yilmaz, A.; Boydak, E. The effects of cobalt-60 applications on yield and yield components of cotton (Gossypium barbadense L.). Pak. J. Biol. Sci. 2006, 9, 2761–2769. [Google Scholar] [CrossRef] [Green Version]
- Martin, P. The taxonomy and ethology of the Afrixalus stuhlmanni complex (Anura: Hyperoliidae). Steenstrupia 2005, 29, 1–38. [Google Scholar]
- Parsons, P.A. Migration as a factor in natural selection. Genetica 1963, 33, 184–206. [Google Scholar] [CrossRef]
- Puffenberger, E.G. Recessive diseases and founder genetics. In Translational and Applied Genomics, Genomics of Rare Diseases; Gonzaga-Jauregui, C., Lupski, J.R., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 97–115. [Google Scholar] [CrossRef]
- Leigh, D.M.; Hendry, A.P.; Vázquez-Domínguez, E.; Friesen, V.L. Estimated six per cent loss of genetic variation in wild populations since the industrial revolution. Evol. Appl. 2019, 12, 1505–1512. [Google Scholar] [CrossRef] [Green Version]
- Ceballosa, G.; Paul, R.; Ehrlichb, L.; Dirzo, R. Biological annihilation via the ongoing sixth mass extinction signaled by vertebrate population losses and declines. Proc. Natl. Acad. Sci. USA 2017, 114, E6089–E6096. [Google Scholar] [CrossRef] [Green Version]
- Khoury, C.K.; Amariles, D.; Soto, J.S.; Diaz, M.V.; Sotelo, S.; Sosa, C.C.; Ramírez-Villegas, J.; Achicanoy, H.A.; Velásquez-Tibatá, J.; Guarino, L.; et al. Comprehensiveness of conservation of useful wild plants: An operational indicator for biodiversity and sustainable development targets. Ecol. Indic. 2019, 98, 420–429. [Google Scholar] [CrossRef]
- Hoban, S.; Bruford, M.; Jackson, J.D.; Lopes-Fernandes, M.; Heuertz, M.; Hohenlohe, P.A.; Paz-Vinas, I.; Sjögren-Gulve, P.; Segelbacher, G.; Vernesi, C.; et al. Genetic diversity targets and indicators in the CBD post-2020 Global Biodiversity Framework must be improved. Biol. Conserv. 2020, 248, 108654. [Google Scholar] [CrossRef]
- Schlaepfer, D.R.; Braschler, B.; Rusterholz, H.P.; Baur, B. Genetic effects of anthropogenic habitat fragmentation on remnant animal and plant populations: A meta-analysis. Ecosphere 2018, 9, e02488. [Google Scholar] [CrossRef]
- De Boef, W.S.; Berg, T.; Haverkort, B. Crop genetic resources. In Biotechnology; Building on Farmers’ Knowledge; Bunders, J., Haverkort, B., Hiemstra, W., Eds.; Macmillan: London, UK, 1996; pp. 103–128. [Google Scholar]
- De Oliveira, L.; Martins, E.R. A quantitative assessment of genetic erosion in ipecac (Psychotria ipecacuanha). Genet. Resour. Crop Evol. 2002, 49, 607–617. [Google Scholar] [CrossRef]
- Smolders, H.; Smolders, H. (Eds.) Enhancing Farmers’ Role in Crop Development: Framework Information for Participatory Plant Breeding in Farmer Field Schools; Centre for Genetic Resources: Wageningen, The Netherlands, 2006; p. 60. [Google Scholar]
- Panis, B.; Nagel, M.; Van den Houwe, I. Challenges and prospects for the conservation of crop genetic resources in field genebanks, in In Vitro Collections and/or in Liquid Nitrogen. Plants 2020, 9, 1634. [Google Scholar] [CrossRef]
- Building on Gender, Agrobiodiversity and Local Knowledge; Food and Agriculture Organization of the United Nations: Rome, Italy, 2004.
- Thrupp, L.A. Linking agricultural biodiversity and food security: The valuable role of agrobiodiversity for sustainable agriculture. Int. Aff. 2000, 76, 265–281. [Google Scholar] [CrossRef]
- The State of the World’s Biodiversity for Food and Agriculture; Food and Agriculture Organization of the United Nations: Rome, Italy, 2019.
- WIEWS—World Information and Early Warning System on Plant Genetic Resources for Food and Agriculture; Food and Agriculture Orgnization of the United Nations: Rome, Italy, 2020.
- Pandotra, P.; Gupta, S. Biotechnological Approaches for Conservation of Plant Genetic Resources and Traditional Knowledge. In Plant Genetic Resources and Traditional Knowledge for Food Security; Salgotra, R.K., Gupta, B.B., Eds.; Springer: Berlin/Heidelberg, Germany, 2015. [Google Scholar] [CrossRef]
- Malhotra, N.; Panatu, S.; Singh, B.; Negi, N.; Singh, D.; Singh, M.; Chandora, R. Genetic Resources: Collection, Conservation, Characterization and Maintenance. In Lentils; Singh, M., Ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 21–41. [Google Scholar] [CrossRef]
- Jose, D.M.; Lucía, D.L.R.; Isaura, M.; Luís, G.; Elena, C.M.; Cristina, M.; Joan, C.; Joan, S.; Ana, R.; German, A.; et al. Plant genebanks: Present situation and proposals for their improvement. the case of the Spanish network. Front. Plant Sci. 2018, 9, 1794. [Google Scholar] [CrossRef] [Green Version]
- Long, C.L.; Li, H.; Ouyang, Z.Q.; Yang, X.Y.; Li, Q.; Trangmar, B. Strategies for agrobiodiversity conservation and promotion: A case from Yunnan, China. Biodivers. Conserv. 2003, 12, 1145–1156. [Google Scholar] [CrossRef]
- Singh, K.; Gupta, K.; Tyagi, V.; Rajkumar, S. Plant genetic resources in India: Management and utilization. Vavilov J. Genet. Breed. 2020, 24, 306–314. [Google Scholar] [CrossRef]
- Rathore, D.S.; Srivastava, U.; Dhillon, B.S. Management of genetic resources of horticultural crops: Issues and strategies. In Plant Genetic Resources: Horticultural Crops; Dhillon, B.S., Tyagi, R.K., Saxena, S., Randhawa, G.J., Eds.; Narosa Publishing: New Delhi, India, 2005; pp. 1–18. [Google Scholar]
- Tao, K.L. Complementary conservation strategy for plant genetic resources. In Conservation Biotechnology of Plant Germplasm; Mandal, B.B., Chaudhury, R., Engelmann, F., Mal, B., Tao, K.L., Dhillon, B.S., Eds.; NBPGR, ICAR: New Delhi, India, 2003; p. 51. [Google Scholar]
- Cruz-Cruz, C.A.; González-Arnao, M.T.; Engelmann, F. Biotechnology and conservation of plant biodiversity. Resources 2013, 2, 73–95. [Google Scholar] [CrossRef]
- Paunesca, A. Biotechnology for endangered plant conservation: A critical overview. Rom. Biotechnol. Lett. 2009, 14, 4095–4104. [Google Scholar]
- Rajasekharan, P.E.; Sahijram, L. In vitro conservation of plant cermplasm. In Plant Biology and Biotechnology; Bahadur, B., Venkat Rajam, M., Sahijram, L., Krishnamurthy, K.V., Eds.; Springer: Berlin/Heidelberg, Germany, 2015; Volume II: Plant Genomics and Biotechnology. [Google Scholar] [CrossRef]
- Salgotra, R.K.; Sharma, M.; Pandotra, P. Biotechnological interventions for sustainable conservation of plant genetic resources in the scenario of climate change. Nat. Res. Conser. Res. 2018, 1, 1146. [Google Scholar] [CrossRef] [Green Version]
- Engelmann, F.; Drew, R.A. In vitro germplasm conservation. Acta Hortic 1998, 461, 41–47. [Google Scholar] [CrossRef]
- Ogbu, J.U.; Essien, B.A.; Essien, J.B.; Anaele, M.U. Conservation and management of genetic resources of horticultural crops in Nigeria: Issues and biotechnological strategies. Int. J. Agric. Res. Dev. 2010, 1, 91–98. [Google Scholar]
- Withers, L.A.; Engelmann, F. In vitro conservation of plant genetic resources. In Biotechnology in Agriculture; Altman, A., Ed.; Dekker: New York, NY, USA, 1997; pp. 57–88. [Google Scholar]
- Martin, C.; Iridono, J.M.; Benito-Gonzales, E.; Perez, C. The use of tissue culture techniques in the conservation of plant biodiversity. Agron. Food Ind. Hi. Tech. 1998, 9, 37–40. [Google Scholar]
- Salgotra, R.K.; Gupta, B.B.; Sood, M. Biotechnological interventions and their role in sustainable hill agriculture. J. Plant Sci. Res. 2015, 2, 118. [Google Scholar]
- Scowcroft, W.R. Genetic Variability in Tissue Culture: Impact on Germplasm Conservation and Utilization; IBPGR: Rome, Italy, 1984. [Google Scholar]
- Kaviani, B. Conservation of plant genetic resources by cryopreservation. Aust. J. Crop Sci. 2011, 5, 778–800. [Google Scholar]
- Fahy, G.M.; MacFarlane, D.R.; Angell, C.A.; Meryman, H.T. Vitrification as an approach to cryopreservation. Cryobiology 1984, 21, 407–426. [Google Scholar] [CrossRef]
- Dulloo, E.; Nagamura, Y.; Ryder, O. DNA storage as a complementary conservation strategy. In DNA Banks-Providing Novel Options for Gene Banks? Topical Reviews in Agricultural Biodiversit; De Vicente, M.C., Andersson, M.S., Eds.; International Plant Genetic Resources Institute: Rome, Italy, 2006. [Google Scholar]
- Duvick, D.N. Plant breeding: Past achievements and expectations for the future. Econ. Bot. 1986, 40, 289–297. [Google Scholar] [CrossRef]
- Wilkes, G. In situ conservation of agricultural systems. In Biodiversity: Culture, Conservations, and Eco-Development; Oldfield, M.L., Alcorn, J.B., Eds.; Westview Press: Boulder, CO, USA, 1991; pp. 86–101. [Google Scholar]
- Salgotra, R.K.; Gupta, B.B.; Ahmed, M.I. Characterization of thermo-sensitive genic male sterility (TGMS) rice genotypes (Oryza sativa L.) at different altitudes. Aus. J. Crop Sci. 2012, 6, 957–962. [Google Scholar]
- Cao, J.; Leng, G.; Yang, P.; Zhou, Q.; Wu, W. Variability in Crop Response to Spatiotemporal Variation in Climate in China, 1980–2014. Land 2022, 11, 1152. [Google Scholar] [CrossRef]
- Salgotra, R.K.; Stewart, N.C. Functional markers for precision plant breeding. Int. J. Mol. Sci. 2020, 21, 4792. [Google Scholar] [CrossRef] [PubMed]
- Richa, S.; Salgotra, R.K.; Bhat, J.A. Comparative diversity analysis in advanced breeding lines of basmati rice (Oryza sativa L.) using agro-morphological and SSR markers. Res. J. Biotech. 2017, 12, 85–93. [Google Scholar]
- Sharma, S.; Singh, A.K.; Kumar, B.; Salgotra, R.K. Microsatellite marker based genetic diversity analysis in basmati rice. Ecosacan 2015, 9, 389–392. [Google Scholar]
- Li, W.; Bian, C.; Wei, Y.M.; Liu, A.J.; Chen, G.Y.; Pu, G.C.; Liu, Y.X.; Zheng, Y.L. Evaluation of genetic diversity of Sichuan common wheat landraces in China by SSR markers. J. Integr. Agric. 2013, 12, 1501–1511. [Google Scholar] [CrossRef]
- Sokolkova, A.; Burlyaeva, M.; Valiannikova, T.; Cui, Y.; Vishnyakova, M.; Schafleitner, R.; Lee, C.R.; Ting, C.T.; Nair, R.M.; Nuzhdin, S.; et al. Genome-wide association study in accessions of the mini-core collection of mungbean (Vigna radiata) from the World Vegetable Gene Bank (Taiwan). BMC Plant Biol. 2020, 20, 363. [Google Scholar] [CrossRef]
- Jeong, N.; Kim, K.S.; Jeong, S.; Kim, J.Y.; Park, S.K.; Lee, J.S.; Jeong, S.C.; Kang, S.T.; Ha, B.K.; Kim, D.; et al. Korean soybean core collection: Genotypic and phenotypic diversity population structure and genome-wide association study. PLoS ONE 2019, 14, e0224074. [Google Scholar] [CrossRef]
- Mahajan, R.; Zargar, S.M.; Singh, R.; Salgotra, R.K.; Sufia, F.; Sonah, H. Population structure analysis and selection of core set among common bean genotypes from Jammu and Kashmir, India. Appl. Biochem. Biotechnol. 2017, 182, 16–28. [Google Scholar] [CrossRef]
- Özkan, G.; Haliloğlu, K.; Türkoğlu, A.; Özturk, H.I.; Elkoca, E.; Poczai, P. Determining genetic diversity and population structure of common bean (Phaseolus vulgaris L.) landraces from Türkiye using SSR markers. Genes 2022, 13, 1410. [Google Scholar] [CrossRef]
- Kimaro, D.; Melis, R.; Sibiya, J.; Shimelis, H.; Shayanowako, A. Analysis of genetic diversity and population structure of pigeonpea (Cajanus cajan L.) Accessions using SSR markers. Plants 2020, 9, 1643. [Google Scholar] [CrossRef]
- Aguilar-Meléndez, A.; Morrell, P.L.; Roose, M.L.; Kim, S.C. Genetic diversity and structure in semiwild and domesticated chilli (Capsicum annuum; Solanaceae) from Mexico. Am. J. Bot. 2009, 96, 1190–1202. [Google Scholar] [CrossRef] [PubMed]
- Akkale, C.; Yildirim, Z.; Yildirim, M.B.; Kaya, C.; Ozturk, G.; Tanyolaç, B. Assessing genetic diversity of some potato (Solanum tuberosum L.) genotypes grown in Turkey by using AFLP marker technique. Turk. J. Field Crops 2010, 15, 73–78. [Google Scholar]
- Baranski, R.; Maksylewicz-Kaul, A.; Nothnagel, T.; Cavagnaro, P.F.; Simon, P.W.; Grzebelus, D. Genetic diversity of carrot (Daucus carota L.) cultivars revealed by analysis of SSR loci. Genet. Resour. Crop Evol. 2012, 59, 163–170. [Google Scholar] [CrossRef] [Green Version]
- Rai, G.K.; Jamwal, D.; Singh, S.; Parveen, A.; Kumar, R.R.; Singh, M.; Rai, P.K.; Salgotra, R.K. assessment of genetic variation in tomato (Solanum Lycopersicum L.) based on quality traits and molecular markers. SABRAO J. Breed. Genet. 2016, 48, 80–89. [Google Scholar]
- Bakoumé, C.; Wickneswari, R.; Siju, S.; Rajanaidu, N.; Kushairi, A.; Billotte, N. Genetic diversity of the world’s largest oil palm (Elaeis guineensis Jacq.) field genebank accessions using microsatellite markers. Genet. Resour. Crop Evol. 2014, 62, 349–360. [Google Scholar] [CrossRef]
- Huang, C.; Nie, X.H.; Shen, C.; You, C.; Li, W.; Zhao, W.; Zhang, X.; Lin, Z. Population structure and genetic basis of the agronomic traits of upland cotton in China revealed by a genome-wide association study using high-density SNPs. Plant Biotechnol. J. 2017, 15, 1374–1386. [Google Scholar] [CrossRef] [PubMed]
- Kala, P.; Zargar, S.; Bali, R.; Gupta, N.; Salgotra, R.; Koul, A. Assessment of Genetic diversity in mulberry using morphological and molecular markers. Elect. J. Plant Breed. 2016, 7, 94–103. [Google Scholar] [CrossRef]
- Chauhan, B.S.; Kaur, V.; Salgotra, R.K. Genetic diversity and population structure analysis to study the evolution of herbicide resistance in Echinochloa colona ecotypes in Australia. Acta Physiol. Plant. 2022, 44, 32. [Google Scholar] [CrossRef]
- Salgotra, R.K.; Stewart, C.N., Jr. Genetic augmentation of legume crops using genomic resources and genotyping platforms for nutritional food security. Plants 2022, 11, 1866. [Google Scholar] [CrossRef]
- Nwosisi, S.; Dhakal, K.; Nandwani, D.; Raji, J.I.; Krishnan, S.; Beovides-García, Y. Genetic Diversity in Vegetable and Fruit Crops. In Genetic Diversity in Horticultural Plants; Nandwani, D., Ed.; Springer: Berlin/Heidelberg, Germany, 2019. [Google Scholar] [CrossRef]
- Bhat, R.; Singh, A.K.; Salgotra, R.K.; Sharma, M.; Mushtaq, M.; Bagati, S.; Hangloo, S.; Singh, A. Detection of QTL for panicle architecture in F2 population of rice. J. Genet. 2019, 98, 50. [Google Scholar] [CrossRef]
- Gupta, N.; Zargar, S.M.; Singh, R.; Nazir, M.; Mahajan, R.; Salgotra, R.K. Marker association study of yield attributing traits in common bean (Phaseolus vulgaris L.). Mol. Biol. Rep. 2020, 47, 6769–6783. [Google Scholar] [CrossRef]
- Bhat, R.; Singh, A.K.; Mushtaq, M.; Salgotra, R.K.; Sharma, M.; Bhat, B.A.; Basu, U.; Al-ashkar, I.; Hossain, M.A.; Ueda, A.; et al. Identification of QTLs for yield and associated traits in F2 population of rice Rohini. Phyton Int. J. Expl. Bot. 2022, 9, 2439–2459. [Google Scholar] [CrossRef]
- Ellstrand, N.C.; Prentice, H.C.; Hancock, J.F. Gene Flow and Introgression from Domesticated Plants into their Wild Relatives. In Adapted with permission from the Annual Review of Ecology and Systematics; Syvanen, M., Kado, C.I., Eds.; Horizontal Gene Transfer (Second Edition); Academic Press: Cambridge, MA, USA, 2002; pp. 217–236. [Google Scholar] [CrossRef]
- Sharma, M.; Dolkar, D.; Salgotra, R.K.; Sharma, D.; Punya, S. Molecular marker assisted confirmation of hybridity in Indian mustard (Brassica juncea L.). Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 894–900. [Google Scholar] [CrossRef]
- Michael, B.K.; Amber, R.N.; Justin, E.A.; Benjamin, K.B.; Loren, H.R. The genetics and genomics of plant domestication. BioScience 2017, 67, 971–982. [Google Scholar] [CrossRef] [Green Version]
- Swarup, S.; Cargill, E.J.; Crosby, K.; Flagel, L.; Kniskern, J.; Glenn, K.C. Genetic diversity is indispensable for plant breeding to improve crops. Crop Sci. 2020. [Google Scholar] [CrossRef]
- Chaudhary, J.; Deshmukh, R.; Sonah, H. Mutagenesis approaches and their role in crop improvement. Plants 2019, 8, 467. [Google Scholar] [CrossRef] [Green Version]
- Samal, P.; Pote, T.D.; Gopala Krishnan, S.; Singh, A.K.; Salgotra, R.K.; Rathour, R. Integrating marker-assisted selection and doubled haploidy for rapid introgression of semi-dwarfing and blast resistance genes into a Basmati rice variety ‘Ranbir Basmati’. Euphytica 2019, 215, 149. [Google Scholar] [CrossRef]
- Folusho, B.; Abebe, M.; Gbadebo, O.; Jose, C.; Sarah, H.; Nnanna, U.; Melaku, G. Genetic gains in yield and yield related traits under drought stress and favorable environments in a maize population improved using marker assisted recurrent selection. Front. Plant Sci. 2017, 8, 808. [Google Scholar] [CrossRef]
- Raina, M.; Salgotra, R.K.; Pandotra, P.; Rathour, R.; Singh, K. Genetic enhancement for semi-dwarf and bacterial blight resistance with enhanced grain quality characteristics in traditional Basmati rice through marker-assisted selection. Comptes Rendus Biol. 2019, 342, 142–153. [Google Scholar] [CrossRef]
- Salgotra, R.K.; Raina, M.; Rathore, R.; Bhat, J.A. Marker-assisted gene pyramiding (MAGP) for semi dwarfed bacterial blight resistance genes into traditional basmati variety “Ranbir Basmati”. Plant Gene 2021, 26, 100276. [Google Scholar] [CrossRef]
- Bhat, J.A.; Ali, S.; Salgotra, R.K.; Mir, Z.A.; Dutta, S.; Vasudha, J.; Tyagi, A.; Muntazir, M.; Jain, N.; Singh, P.K.; et al. Genomic selection in the era of next generation sequencing for complex traits in plant breeding. Front. Genet. 2016, 7, 221. [Google Scholar] [CrossRef] [Green Version]
- Bhat, J.A.; Salgotra, R.K.; Gupta, B.B.; Kaushik, R.P.; Kumar, B.; Sharma, M.; Razdan, V.K.; Rai, G.K.; Gupta, M. Development of bacterial blight resistance versions of basmati rice genotypes from Jammu, Northern Himalaya using marker-assisted selection. Indian J. Biochem. Biophys. 2015, 52, 341–348. [Google Scholar]
- Kohli, S.; Gupta, B.B.; Kumar, B.; Kumar, R.; Chaturvedi, V.D.; Singh, H.; Priyatham, K.; Salgotra, R.K.; Samnotra, R.K. Characterization of basmati germplasm for bacterial leaf blight resistance and grain quality traits under North-Western Himalayan region of J&K. Agric. Mech. Asia 2022, 53, 7429–7438. [Google Scholar]
- Hu, T.; Zhong, X.; Yang, Q.; Zhou, X.; Li, X.; Yang, S.; Hou, L.; Yao, Q.; Guo, Q.; Kang, Z. Introgression of two quantitative trait loci for stripe rust resistance into three chinese wheat cultivars. Agronomy 2020, 10, 483. [Google Scholar] [CrossRef] [Green Version]
- Yadav, P.S.; Mishra, V.K.; Arun, B.; Chand, R.; Vishwakarma, M.K.; Vasistha, N.K.; Mishra, A.N.; Kalappanavar, I.K.; Joshi, A.K. Enhanced resistance in wheat against stem rust achieved by marker assisted backcrossing involving three independent Sr genes. Curr. Plant Biol. 2015, 2, 25–33. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.; Srivastava, P.; Mavi, G.S.; Kaur, S.; Kaur, J.; Bala, B.; Singh, T.P.; Sohu, V.S.; Chhuneja, P.; Bains, N.S.; et al. Resurrection of wheat cultivar PBG343 using marker-assisted gene pyramiding for rust resistance. Front. Plant Sci. 2021, 12, 570408. [Google Scholar] [CrossRef] [PubMed]
- Slikova, S.; GREGOVÁ, E.; BARTOŠ, P.; KRAIC, J. Marker-Assisted Selection for leaf rust resistance in wheat by transfer of gene Lr19. Plant Protect. Sci. 2003, 39, 13–17. [Google Scholar] [CrossRef] [Green Version]
- Chandran, S.; Pukalenthy, B.; Adhimoolam, K.; Manickam, D.; Sampathrajan, V.; Chocklingam, V.; Eswaran, K.; Arunachalam, K.; Joikumar Meetei, L.; Rajasekaran, R.; et al. Marker-assisted selection to pyramid the opaque-2 (o2) and β-carotene (crtrb1) genes in maize. Front. Genet. 2019, 10, 859. [Google Scholar] [CrossRef] [Green Version]
- Dinesh, H.B.; Chandappa, L.; Vishwanath, K.P.; Singh, P.; Manjunatha, L.; Ambika, D.S.; Kumar, M.P.K. Genetic analysis and marker assisted backcrossing for transfer of mosaic virus resistance in cowpea [Vigna unguiculata (L.) Walp.]. Legume Res. 2018, 41, 663–668. [Google Scholar] [CrossRef]
- Zargar, S.M.; Nazir, M.; Gupta, M.; Farhat, S.; Mahajan, R.; Salgotra, R.K.; Mir, R.A. Molecular marker assisted approaches (MMAA) for enhancing low water stress tolerance in common bean: An update. Mol. Plant Breed. 2014, 5, 1–12. [Google Scholar] [CrossRef]
- Garzon, L.N.; Ligarreto, G.A.; Blair, M.W. Molecular marker-assisted backcrossing of anthracnose resistance into andean climbing beans (Phaseolus vulgaris L.). Crop Sci. 2008, 48, 562–570. [Google Scholar] [CrossRef]
- Kim, M.; Hyten, D.L.; Niblack, T.L.; Diers, B.W. Stacking resistance alleles from wild and domestic soybean sources improves soybean cyst nematode resistance. Crop Sci. 2011, 51, 934–943. [Google Scholar] [CrossRef] [Green Version]
- Ramalingam, J.; Alagarasan, G.; Savitha, P.; Lydia, K.; Pothiraj, G.; Vijayakumar, E.; Sudhagar, R.; Singh, A.; Vedna, K.; Vanniarajan, C. Improved host-plant resistance to Phytophthora rot and powdery mildew in soybean (Glycine max (L.) Merr.). Sci. Rep. 2020, 10, 13928. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.G.; Lin, Z.; Kai, L.; Ying, M.; Wang, L.Q.; Yang, Y.Q.; Yang, Y.H.; Zhi, H.J. Marker-assisted pyramiding of soybean resistance genes RSC4, RSC8, and RSC14Q to soybean mosaic virus. J. Integr. Agric. 2017, 16, 2413–2420. [Google Scholar] [CrossRef]
- Maranna, S.; Verma, K.; Talukdar, A.; Lal, S.K.; Kumar, A.; Mukherjee, K. Introgression of null allele of kunitz trypsin inhibitor through marker-assisted backcross breeding in soybean (Glycine max L. Merr.). BMC Genet. 2016, 12, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rawal, R.; Kumar, V.; Rani, A.; Gokhale, S.M. Genetic elimination of off-flavour generating lipoxygenase-2 gene of soybean through marker assisted backcrossing and its effect on seed longevity. Plant Breed. Biotech. 2020, 8, 163–173. [Google Scholar] [CrossRef]
- Varshney, R.K.; Pandey, M.K.; Janila, P.; Nigam, S.N.; Sudini, H.; Gowda, M.V.; Sriswathi, M.; Radhakrishnan, T.; Manohar, S.S.; Nagesh, P. Marker-assisted introgression of a QTL region to improve rust resistance in three elite and popular varieties of peanut (Arachis hypogaea L.). Theor. Appl. Genet. 2014, 127, 1771–1781. [Google Scholar] [CrossRef] [Green Version]
- Chu, Y.; Gill, R.; Clevenger, J.; Timper, P.; Holbrook, C.C.; Ozias-Akins, P. Identification of rare recombinants leads to tightly linked markers for nematode resistance in peanut. Peanut Sci. 2016, 43, 88–93. [Google Scholar] [CrossRef] [Green Version]
- Chu, Y.; Wu, C.L.; Holbrook, C.C.; Tillman, B.L.; Person, G.; Ozias-Akins, P. Marker-assisted selection to pyramid nematode resistance and the high oleic trait in peanut. Plant Genome 2011, 4, 110–117. [Google Scholar] [CrossRef] [Green Version]
- Janila, P.; Pandey, M.K.; Manohar, S.S.; Variath, M.T.; Nallathambi, P.; Nadaf, H.L.; Sudini, H.; Varshney, R.K. Foliar fungal disease-resistant introgression lines of groundnut (Arachis hypogaea L.) record higher pod and haulm yield in multilocation testing. Plant Breed. 2016, 135, 355–366. [Google Scholar]
- Shasidhar, Y.; Variath, M.T.; Vishwakarma, M.K.; Manohar, S.S.; Gangurde, S.S.; Sriswathi, M.; Sudini, H.K.; Dobariya, K.L.; Bera, S.K.; Radhakrishnan, T.; et al. Improvement of three Indian popular groundnut varieties for foliar disease resistance and high oleic acid using SSR markers and SNP array in marker-assisted backcrossing. Crop J. 2020, 8, 1–15. [Google Scholar] [CrossRef]
- Varshney, R.K.; Mohan, S.M.; Gaur, P.M.; Chamarthi, S.K.; Singh, V.K.; Srinivasan, S.; Swapna, N.; Sharma, M.; Pande, S.; Singh, S.; et al. Marker-assisted backcrossing to introgress resistance to fusarium wilt race 1 and ascochyta blight in C 214, an elite cultivar of chickpea. Plant Genome 2014, 7, 35. [Google Scholar] [CrossRef] [Green Version]
- Varshney, R.K.; Gaur, P.M.; Chamarthi, S.K.; Krishnamurthy, L.; Tripathi, S.; Kashiwagi, J.; Samineni, S.; Singh, V.K.; Thudi, M.; Jaganathan, D. Fast-track introgression of “QTL-hotspot” for root traits and other drought tolerance traits in JG 11, an elite and leading variety of chickpea. Plant Genome 2013, 6, 1–9. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| S. No. | International Research Institute | Mandate/Crops |
|---|---|---|
| 1. | International Rice Research Institute (IRRI), Los Banos, Philippines | Rice |
| 2. | Centre International de-Mejoramients de Maize (CIMMYT), Trigo, El Baton, Mexico | Maize and wheat (triticale, barely, sorghum) |
| 3. | Center International de-agricultural Tropical (CIAT), Palmira, Columbia | Cassava and beans (also maize and rice), in collaboration with CIMMYT and IRRI |
| 4. | International Institute of Tropical Agriculture (IITA), Ibadan, Nigeria | Grain legumes, roots and tubers, farming systems, cassava, banana, yam |
| 5. | Centre International de la Papa (CIP), Lima. Peru | Potato, Andean root, and tubers |
| 6. | International Crops Research Institute, for Semi-Arid Tropics (ICRISAT), Hyderabad, India | Sorghum, groundnut, pearl millet, Bengal gram, red gram |
| 7. | West African Rice Development Association (WARDA), Monrovia, Liberia | Regional cooperative rice research in collaboration with IITA and IRRI |
| 8. | International Plant Genetic Research Institute (IPGRI), Rome Italy | Genetic conservation |
| 9. | National Bureau of Plant Genetic Resources, New Delhi, India | Fruits, tubers, medicinal and aromatic crops, spices, bulbous crops |
| 10. | The Asian Vegetable Research and Development Center (AVRDC), Taiwan | Tomato, onion, peppers, Chinese cabbage |
| 11. | International Center for Tropical Agriculture (CIAT) Columbia | Cassava |
| 12. | The New Zealand Institute for Plant and Food Research Limited, New Zealand | Kiwifruit (Actinidia spp.) |
| 13. | Svalbard Global Seed Vault, Norway | All crops from different countries |
| Crop | Molecular Breeding Approaches | Trait(s) Improved | Reference |
|---|---|---|---|
| Rice | MABB | Bacterial blight resistance | [107] |
| MABB | Semi-dwarf and bacterial blight resistance | [104,108] | |
| MABB | Blast resistance | [102] | |
| Wheat | MABB | Stripe rust resistance | [109] |
| MABB | Stem rust resistance | [110,111] | |
| MAS | Leaf rust resistance | [112] | |
| Maize | MABB | Quality improvement | [113] |
| Cowpea | MABC | Mosaic virus (CpMV) resistance | [114] |
| Common bean | MABB | Improved drought adaptation | [115] |
| MABC | Anthracnose resistance | [116] | |
| Soybean | MAS and MABC | Several soybean cyst nematodes and multiple disease-resistant genotypes | [117] |
| MABB | Powdery mildew diseases resistance | [118] | |
| MABC | Soybean mosaic virus (SMV) resistance | [119] | |
| MABC | Free Kunitz trypsin inhibitor | [120] | |
| MABC | Elimination of lipoxygenase-2, | [121] | |
| Peanut | MABC | Introgression lines showing higher yield and increased rust resistance | [122] |
| MABC | Resistance to nematode | [123,124] | |
| MABC | Enhanced oleic acid | [125,126] | |
| Chickpea | MABC | Resistance to fusarium wilt | [127] |
| MABC | Resistance to Ascochyta blight | [127] | |
| MABC | Drought tolerance | [128] | |
| MABC | Elimination of lipoxygenase-2, | [121] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salgotra, R.K.; Chauhan, B.S. Genetic Diversity, Conservation, and Utilization of Plant Genetic Resources. Genes 2023, 14, 174. https://doi.org/10.3390/genes14010174
Salgotra RK, Chauhan BS. Genetic Diversity, Conservation, and Utilization of Plant Genetic Resources. Genes. 2023; 14(1):174. https://doi.org/10.3390/genes14010174
Chicago/Turabian StyleSalgotra, Romesh Kumar, and Bhagirath Singh Chauhan. 2023. "Genetic Diversity, Conservation, and Utilization of Plant Genetic Resources" Genes 14, no. 1: 174. https://doi.org/10.3390/genes14010174