
Walking Training Increases microRNA-126 Expression and Muscle Capillarization in Patients with Peripheral Artery Disease
 , , , , ,
, , , , ,  , and
, and
Abstract
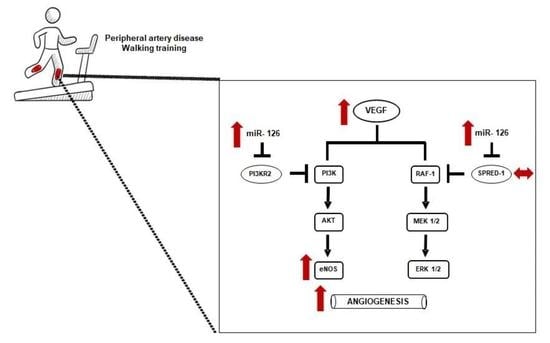
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Patients and Procedures
2.3. Muscle Analyses
2.3.1. Angiogenesis and Muscle Fiber Type Distribution
2.3.2. RNA Extraction
2.3.3. mRNA and miRNA Analysis by Real-Time Polymerase Chain Reaction
2.4. Statistical Analysis
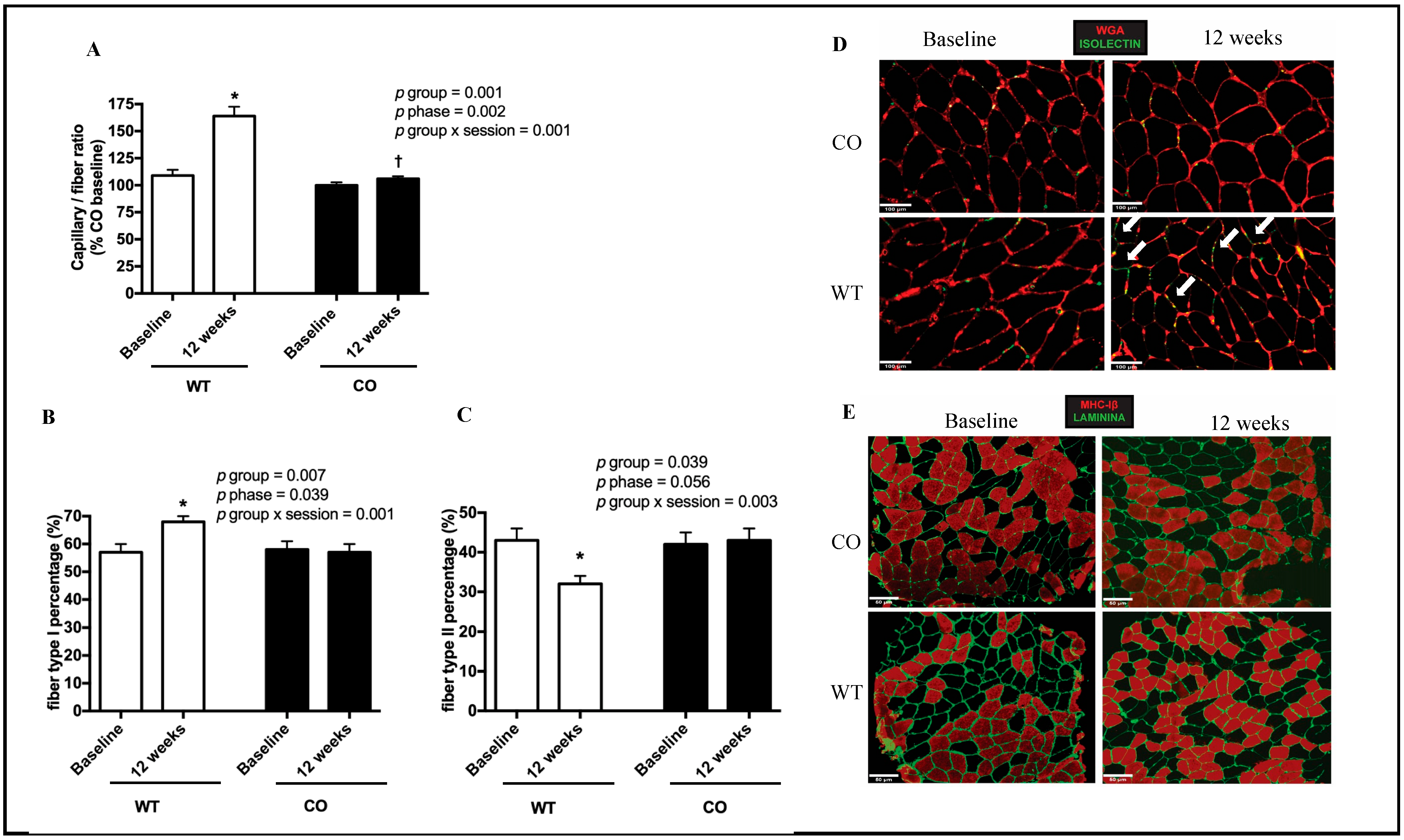
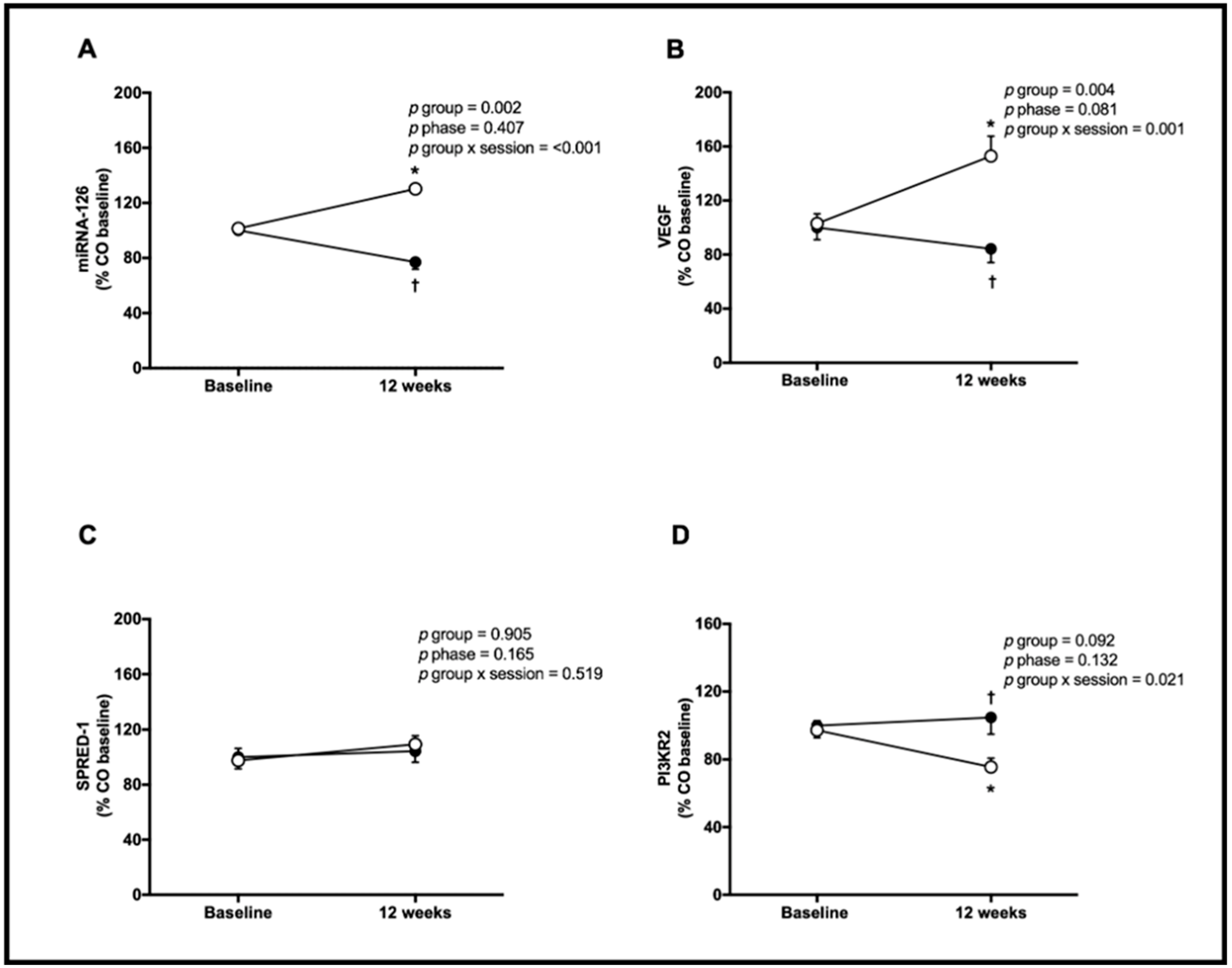
3. Results
4. Discussion
Study Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Norgren, L.; Hiatt, W.R.; Dormandy, J.A.; Nehler, M.R.; Harris, K.A.; Fowkes, F.G.; Group, T.I.W. Inter-Society Consensus for the Management of Peripheral Arterial Disease (TASC II). J. Vasc. Surg. 2007, 45, S5–S67. [Google Scholar] [CrossRef] [Green Version]
- Fowkes, F.G.; Aboyans, V.; Fowkes, F.J.; McDermott, M.M.; Sampson, U.K.; Criqui, M.H. Peripheral artery disease: Epidemiology and global perspectives. Nat. Rev. Cardiol. 2017, 14, 156–170. [Google Scholar] [CrossRef] [PubMed]
- Myers, S.A.; Johanning, J.M.; Stergiou, N.; Lynch, T.G.; Longo, G.M.; Pipinos, I.I. Claudication distances and the Walking Impairment Questionnaire best describe the ambulatory limitations in patients with symptomatic peripheral arterial disease. J. Vasc. Surg. 2008, 47, 550–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clyne, C.A.; Mears, H.; Weller, R.O.; O’Donnell, T.F. Calf muscle adaptation to peripheral vascular disease. Cardiovasc. Res. 1985, 19, 507–512. [Google Scholar] [CrossRef] [PubMed]
- Askew, C.D.; Green, S.; Walker, P.J.; Kerr, G.K.; Green, A.A.; Williams, A.D.; Febbraio, M.A. Skeletal muscle phenotype is associated with exercise tolerance in patients with peripheral arterial disease. J. Vasc. Surg. 2005, 41, 802–807. [Google Scholar] [CrossRef] [Green Version]
- McGuigan, M.R.; Bronks, R.; Newton, R.U.; Sharman, M.J.; Graham, J.C.; Cody, D.V.; Kraemer, W.J. Muscle fiber characteristics in patients with peripheral arterial disease. Med. Sci. Sport. Exerc. 2001, 33, 2016–2021. [Google Scholar] [CrossRef]
- Robbins, J.L.; Jones, W.S.; Duscha, B.D.; Allen, J.D.; Kraus, W.E.; Regensteiner, J.G.; Hiatt, W.R.; Annex, B.H. Relationship between leg muscle capillary density and peak hyperemic blood flow with endurance capacity in peripheral artery disease. J. Appl. Physiol. 2011, 111, 81–86. [Google Scholar] [CrossRef] [Green Version]
- Gerhard-Herman, M.D.; Gornik, H.L.; Barrett, C.; Barshes, N.R.; Corriere, M.A.; Drachman, D.E.; Fleisher, L.A.; Fowkes, F.G.; Hamburg, N.M.; Kinlay, S.; et al. 2016 AHA/ACC Guideline on the Management of Patients With Lower Extremity Peripheral Artery Disease: Executive Summary: A Report of the American College of Cardiology/American Heart Association Task Force on Clinical Practice Guidelines. Circulation 2017, 135, e686–e725. [Google Scholar] [CrossRef]
- Duscha, B.D.; Robbins, J.L.; Jones, W.S.; Kraus, W.E.; Lye, R.J.; Sanders, J.M.; Allen, J.D.; Regensteiner, J.G.; Hiatt, W.R.; Annex, B.H. Angiogenesis in skeletal muscle precede improvements in peak oxygen uptake in peripheral artery disease patients. Arter. Thromb. Vasc. Biol. 2011, 31, 2742–2748. [Google Scholar] [CrossRef] [Green Version]
- Andrade-Lima, A.; Silva Junior, N.; Chehuen, M.; Miyasato, R.; Souza, R.W.A.; Leicht, A.S.; Brum, P.C.; de Oliveira, E.M.; Wolosker, N.; Forjaz, C.L.M. Walking Training Improves Systemic and Local Pathophysiological Processes in Intermittent Claudication. Eur. J. Vasc. Endovasc. Surg. 2021, 61, 954–963. [Google Scholar] [CrossRef]
- Milkiewicz, M.; Hudlicka, O.; Brown, M.D.; Silgram, H. Nitric oxide, VEGF, and VEGFR-2: Interactions in activity-induced angiogenesis in rat skeletal muscle. Am. J. Physiol. Heart Circ. Physiol. 2005, 289, H336–H343. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.J.; Yang, D.D.; Na, S.; Sandusky, G.E.; Zhang, Q.; Zhao, G. Dicer is required for embryonic angiogenesis during mouse development. J. Biol. Chem. 2005, 280, 9330–9335. [Google Scholar] [CrossRef] [Green Version]
- Kim, V.N. MicroRNA biogenesis: Coordinated cropping and dicing. Nat. Rev. Mol. Cell Biol. 2005, 6, 376–385. [Google Scholar] [CrossRef]
- Fish, J.E.; Santoro, M.M.; Morton, S.U.; Yu, S.; Yeh, R.F.; Wythe, J.D.; Ivey, K.N.; Bruneau, B.G.; Stainier, D.Y.; Srivastava, D. miR-126 regulates angiogenic signaling and vascular integrity. Dev. Cell 2008, 15, 272–284. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, N.D., Jr.; Roseguini, B.T.; Chehuen, M.; Fernandes, T.; Mota, G.F.; Martin, P.K.; Han, S.W.; Forjaz, C.L.; Wolosker, N.; de Oliveira, E.M. Effects of oral N-acetylcysteine on walking capacity, leg reactive hyperemia, and inflammatory and angiogenic mediators in patients with intermittent claudication. Am. J. Physiol. Heart Circ. Physiol. 2015, 309, H897–H905. [Google Scholar] [CrossRef] [Green Version]
- Luttrell, M.J.; Halliwill, J.R. Recovery from exercise: Vulnerable state, window of opportunity, or crystal ball? Front. Physiol. 2015, 6, 204. [Google Scholar] [CrossRef] [Green Version]
- Gardner, A.W.; Parker, D.E.; Montgomery, P.S. Changes in vascular and inflammatory biomarkers after exercise rehabilitation in patients with symptomatic peripheral artery disease. J. Vasc. Surg. 2019, 70, 1280–1290. [Google Scholar] [CrossRef]
- Gardner, A.W.; Skinner, J.S.; Cantwell, B.W.; Smith, L.K. Progressive vs. single-stage treadmill tests for evaluation of claudication. Med. Sci. Sport. Exerc. 1991, 23, 402–408. [Google Scholar] [CrossRef]
- Da Rocha Chehuen, M.; Cucato, G.G.; Saes, G.F.; Costa, L.A.; Leicht, A.S.; Ritti-Dias, R.M.; Wolosker, N.; de Moraes Forjaz, C.L. Reproducibility of Anaerobic and Pain Thresholds in Male Patients With Intermittent Claudication. J. Cardiopulm. Rehabil. Prev. 2016, 36, 358–367. [Google Scholar] [CrossRef]
- Jain, A.; Liu, K.; Ferrucci, L.; Criqui, M.H.; Tian, L.; Guralnik, J.M.; Tao, H.; McDermott, M.M. Declining walking impairment questionnaire scores are associated with subsequent increased mortality in peripheral artery disease. J. Am. Coll. Cardiol. 2013, 61, 1820–1829. [Google Scholar] [CrossRef]
- Treat-Jacobson, D.; McDermott, M.M.; Bronas, U.G.; Campia, U.; Collins, T.C.; Criqui, M.H.; Gardner, A.W.; Hiatt, W.R.; Regensteiner, J.G.; Rich, K.; et al. Optimal Exercise Programs for Patients With Peripheral Artery Disease: A Scientific Statement From the American Heart Association. Circulation 2019, 139, e10–e33. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, S.; Yokoyama, N.; Tamori, Y.; Akutsu, K.; Hashimoto, H.; Takeshita, S. Patients with peripheral artery disease who complete 12-week supervised exercise training program show reduced cardiovascular mortality and morbidity. Circ. J. 2009, 73, 167–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, W.C.; Mohler, E., 3rd; Ratcliffe, S.J.; Wehrli, F.W.; Detre, J.A.; Floyd, T.F. Skeletal muscle microvascular flow in progressive peripheral artery disease: Assessment with continuous arterial spin-labeling perfusion magnetic resonance imaging. J. Am. Coll. Cardiol. 2009, 53, 2372–2377. [Google Scholar] [CrossRef] [Green Version]
- Lindner, J.R.; Womack, L.; Barrett, E.J.; Weltman, J.; Price, W.; Harthun, N.L.; Kaul, S.; Patrie, J.T. Limb stress-rest perfusion imaging with contrast ultrasound for the assessment of peripheral arterial disease severity. JACC Cardiovasc. Imaging 2008, 1, 343–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beckitt, T.A.; Day, J.; Morgan, M.; Lamont, P.M. Skeletal muscle adaptation in response to supervised exercise training for intermittent claudication. Eur. J. Vasc. Endovasc. Surg. 2012, 44, 313–317. [Google Scholar] [CrossRef] [Green Version]
- Palmer-Kazen, U.; Religa, P.; Wahlberg, E. Exercise in patients with intermittent claudication elicits signs of inflammation and angiogenesis. Eur. J. Vasc. Endovasc. Surg. 2009, 38, 689–696. [Google Scholar] [CrossRef] [Green Version]
- Wood, R.E.; Sanderson, B.E.; Askew, C.D.; Walker, P.J.; Green, S.; Stewart, I.B. Effect of training on the response of plasma vascular endothelial growth factor to exercise in patients with peripheral arterial disease. Clin. Sci. (Lond.) 2006, 111, 401–409. [Google Scholar] [CrossRef]
- Wang, S.; Aurora, A.B.; Johnson, B.A.; Qi, X.; McAnally, J.; Hill, J.A.; Richardson, J.A.; Bassel-Duby, R.; Olson, E.N. The endothelial-specific microRNA miR-126 governs vascular integrity and angiogenesis. Dev. Cell 2008, 15, 261–271. [Google Scholar] [CrossRef] [Green Version]
- Donghui, T.; Shuang, B.; Xulong, L.; Meng, Y.; Yujing, G.; Yujie, H.; Juan, L.; Dongsheng, Y. Improvement of microvascular endothelial dysfunction induced by exercise and diet is associated with microRNA-126 in obese adolescents. Microvasc. Res. 2019, 123, 86–91. [Google Scholar] [CrossRef]
- Silva, N.D., Jr.; Fernandes, T.; Soci, U.P.; Monteiro, A.W.; Phillips, M.I.; EM, D.E.O. Swimming training in rats increases cardiac MicroRNA-126 expression and angiogenesis. Med. Sci. Sport. Exerc. 2012, 44, 1453–1462. [Google Scholar] [CrossRef]
- Gomes, J.L.; Fernandes, T.; Soci, U.P.; Silveira, A.C.; Barretti, D.L.; Negrao, C.E.; Oliveira, E.M. Obesity Downregulates MicroRNA-126 Inducing Capillary Rarefaction in Skeletal Muscle: Effects of Aerobic Exercise Training. Oxid. Med. Cell Longev. 2017, 2017, 2415246. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, T.; Magalhaes, F.C.; Roque, F.R.; Phillips, M.I.; Oliveira, E.M. Exercise training prevents the microvascular rarefaction in hypertension balancing angiogenic and apoptotic factors: Role of microRNAs-16, -21, and -126. Hypertension 2012, 59, 513–520. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
da Silva, N.D., Jr.; Andrade-Lima, A.; Chehuen, M.R.; Leicht, A.S.; Brum, P.C.; Oliveira, E.M.; Wolosker, N.; Pelozin, B.R.A.; Fernandes, T.; Forjaz, C.L.M. Walking Training Increases microRNA-126 Expression and Muscle Capillarization in Patients with Peripheral Artery Disease. Genes 2023, 14, 101. https://doi.org/10.3390/genes14010101
da Silva ND Jr., Andrade-Lima A, Chehuen MR, Leicht AS, Brum PC, Oliveira EM, Wolosker N, Pelozin BRA, Fernandes T, Forjaz CLM. Walking Training Increases microRNA-126 Expression and Muscle Capillarization in Patients with Peripheral Artery Disease. Genes. 2023; 14(1):101. https://doi.org/10.3390/genes14010101
Chicago/Turabian Styleda Silva, Natan D., Jr., Aluisio Andrade-Lima, Marcel R. Chehuen, Anthony S. Leicht, Patricia C. Brum, Edilamar M. Oliveira, Nelson Wolosker, Bruno R. A. Pelozin, Tiago Fernandes, and Cláudia L. M. Forjaz. 2023. "Walking Training Increases microRNA-126 Expression and Muscle Capillarization in Patients with Peripheral Artery Disease" Genes 14, no. 1: 101. https://doi.org/10.3390/genes14010101