
RNA-Seq Reveals the Underlying Molecular Mechanism of First Cleavage Time Affecting Porcine Embryo Development



 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Porcine Oocytes
2.2. In Vitro Maturation (IVM) of Porcine Oocytes
2.3. Parthenogenetic Activation (PA)
2.4. In Vitro Fertilization (IVF)
2.5. Cultivation and Statistics of Embryos Cleaved at Different Time
2.6. Assessments of Nucleus Number in Blastocysts of PA Embryos
2.7. RNA Sequencing of Embryos
2.8. Quantitative Real-Time PCR
2.9. Statistical Analysis
3. Results
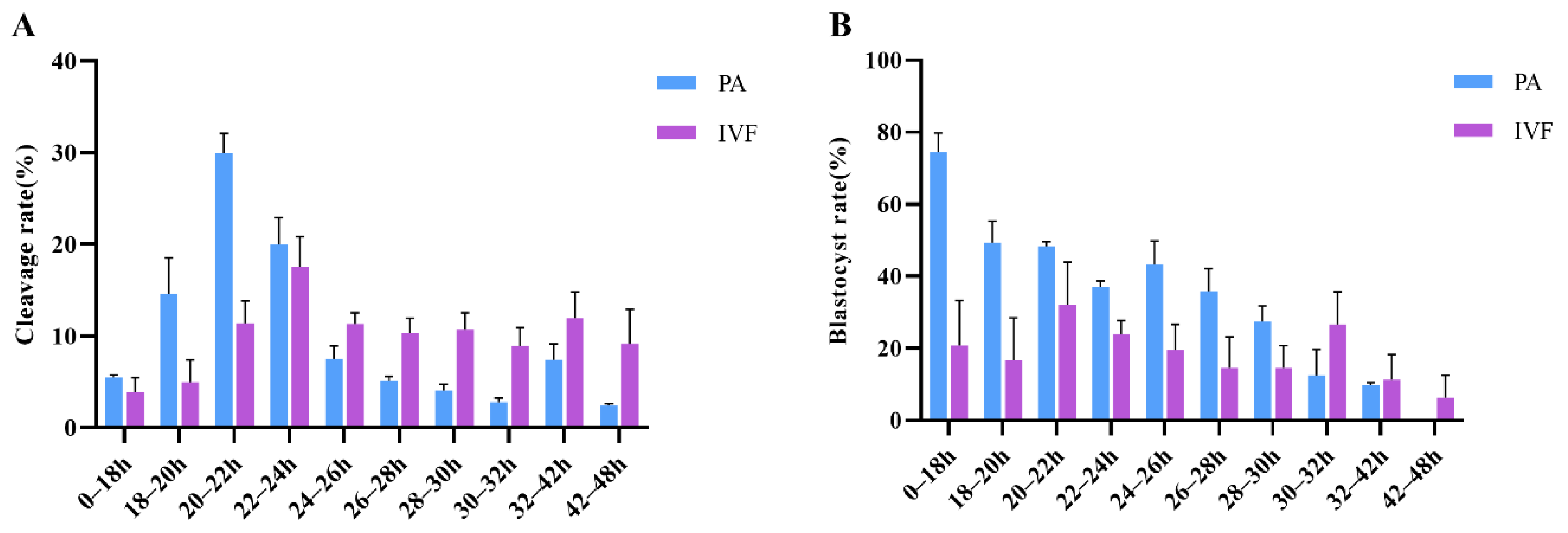
3.1. Relationship between the Timing of First Cleavage and Embryo Developmental Potential of PA
3.2. First Cleavage Time and Developmental Competence of IVF Embryos
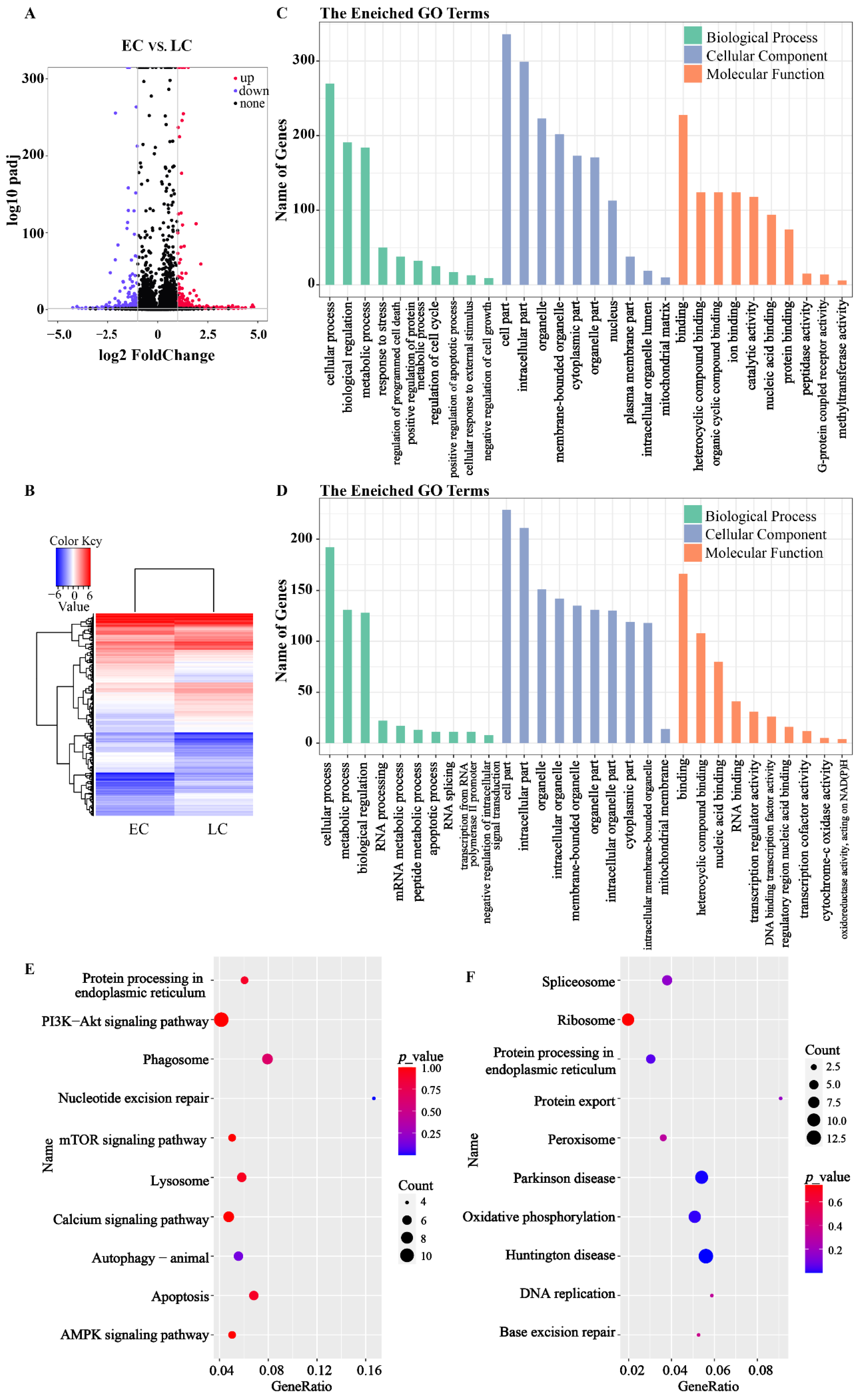
3.3. RNA-Seq of the PA Embryos
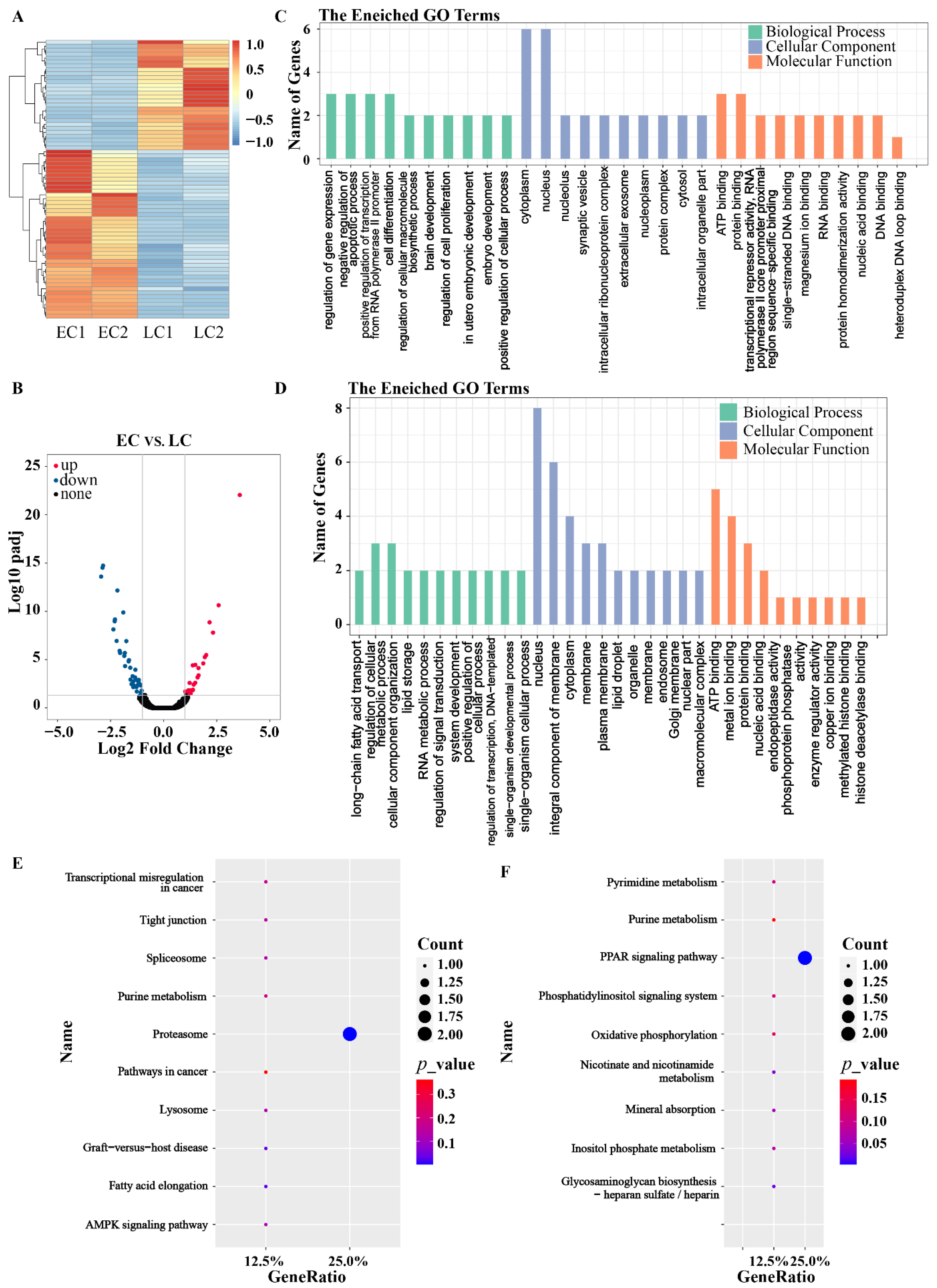
3.4. RNA-Seq of the IVF Embryos
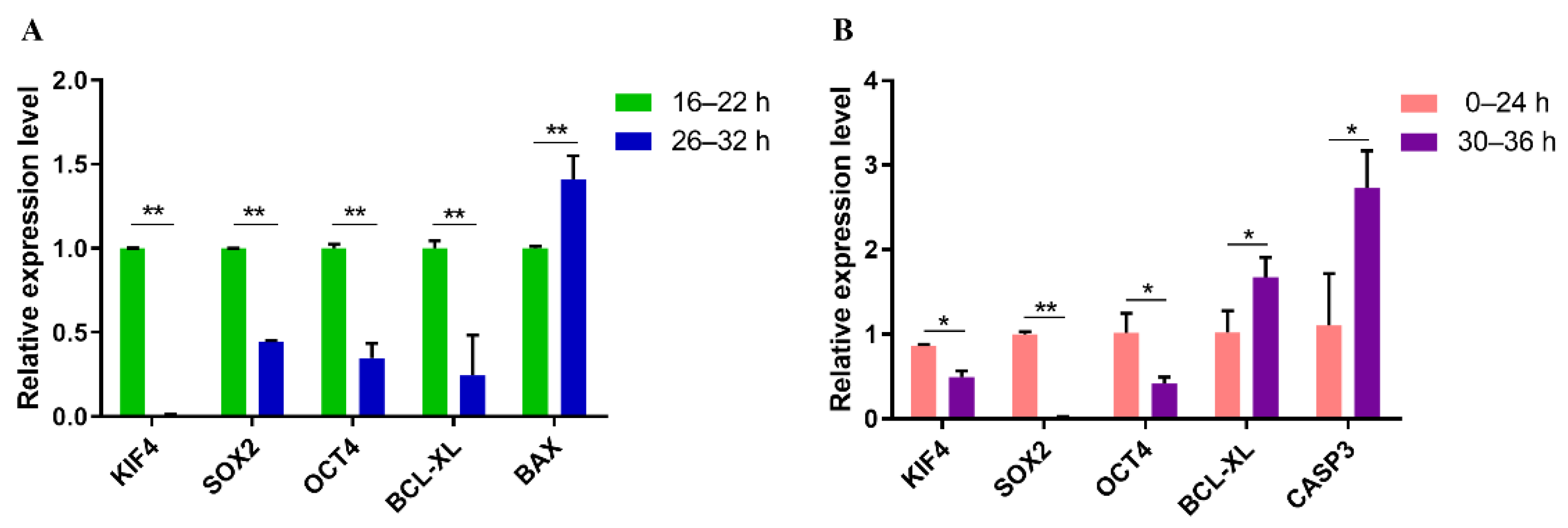
3.5. Quantitative Real-Time PCR Analysis of Genes Related to Apoptosis and Pluripotent in PA and IVF Embryos
3.6. Molecular Mechanism Underlying the Retardation of Development in Late Cleavage Embryos
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Loutradis, D.; Drakakis, P.; Kallianidis, K.; Milingos, S.; Dendrinos, S.; Michalas, S. Oocyte morphology correlates with embryo quality and pregnancy rate after intracytoplasmic sperm injection. Fertil. Steril. 1999, 72, 240–244. [Google Scholar] [CrossRef]
- Baczkowski, T.; Kurzawa, R.; Glabowski, W. Methods of embryo scoring in in vitro fertilization. Reprod. Biol. 2004, 4, 5–22. [Google Scholar] [PubMed]
- Lechniak, D.; Pers-Kamczyc, E.; Pawlak, P. Timing of the first zygotic cleavage as a marker of developmental potential of mammalian embryos. Reprod. Biol. 2008, 8, 23–42. [Google Scholar] [CrossRef]
- Machtinger, R.; Racowsky, C. Morphological systems of human embryo assessment and clinical evidence. Reprod. Biomed. Online 2013, 26, 210–221. [Google Scholar] [CrossRef] [Green Version]
- Montag, M.; Toth, B.; Strowitzki, T. New approaches to embryo selection. Reprod. Biomed. Online 2013, 27, 539–546. [Google Scholar] [CrossRef]
- Bavister, B. Timing of Embryo Development. In Assessment of Embryo Quality: Invasive and Non-invasive Techniques; Springer Science + Business Media: Dordrecht, The Netherlands, 2002; pp. 139–156. [Google Scholar]
- Soom, A.V.; Ysebaert, M.-T.R.S.; Kruif, A.D. Relationship Between Timing of Development, Morula Morphology, and Cell Allocation to Inner Cell Mass and Trophectoderm in In Vitro-Produced Bovine Embryos. Mol. Reprod. Dev. 1997, 47, 47–56. [Google Scholar] [CrossRef]
- Vandaele, L.; Mateusen, B.; Maes, D.; de Kruif, A.; van Soom, A. Is apoptosis in bovine in vitro produced embryos related to early developmental kinetics and in vivo bull fertility? Theriogenology. 2006, 65, 1691–1703. [Google Scholar] [CrossRef]
- Velker, B.A.M.; Denomme, M.M.; Mann, A.M.R.W. Loss of Genomic Imprinting in Mouse Embryos with Fast Rates of Preimplantation Development in Culture. Biol. Reprod. 2012, 86, 143. [Google Scholar]
- Gutierrez-Adan, A.; White, C.R.; Soom, A.V.; Mann, M.R.W. Why we should not select the faster embryo: Lessons from mice and cattle. Reprod. Fertil. Dev. 2014, 27, 765–775. [Google Scholar] [CrossRef]
- Dang-Nguyen, T.Q.; Kikuchi, K.; Somfai, T.; Ozawa, M.; Nakai, M.; Viet-Linh, N.; Kanai, Y.; Nguyen, B.X.; Nagai, T. Evaluation of developmental competence of in vitro-produced porcine embryos based on the timing, pattern and evenness of the first cleavage and onset of the second cleavage. J. Reprod. Dev. 2010, 56, 593–600. [Google Scholar] [CrossRef] [Green Version]
- Isom, S.C.; Li, R.F.; Whitworth, K.M.; Prather, S.R. Timing of first embryonic cleavage is a positive indicator of the in vitro developmental potential of porcine embryos derived from in vitro fertilization, somatic cell nuclear transfer and parthenogenesis. Mol. Reprod. Dev. 2012, 79, 197–207. [Google Scholar] [CrossRef]
- Lee, M.J.; Lee, R.K.; Lin, M.H.; Hwu, Y.M. Cleavage speed and implantation potential of early-cleavage embryos in IVF or ICSI cycles. J. Assist. Reprod. Genet. 2012, 29, 745–750. [Google Scholar] [CrossRef] [Green Version]
- Kaith, S.; Saini, M.; Raja, A.K.; Sahare, A.A.; Jyotsana, B.; Chauhan, M.S.; Manik, R.S.; Singla, S.K. Early Cleavage of Handmade Cloned Buffalo (Bubalus bubalis) Embryos is an Indicator of Their Developmental Competence and Quality. Reprod. Domest. Anim. 2015, 50, 214–220. [Google Scholar] [CrossRef]
- Milazzotto, M.P.; Goissis, M.D.; Chitwood, J.L.; Annes, K.; Soares, C.A.; Assumpção, M.E.O.Á.; Ross, P.J. Early cleavages influence the molecular and the metabolic pattern of individually cultured bovine blastocysts. Mol. Reprod. Dev. 2016, 83, 324–336. [Google Scholar] [CrossRef]
- Leoni, G.G.; Succu, S.; Berlinguer, F.; Rosati, I.; Bebbere, D.; Bogliolo, L.; Ledda, S.C.; Naitana, S. Delay on the in vitro kinetic development of prepubertal ovine embryos. Anim. Reprod. Sci. 2006, 92, 373–383. [Google Scholar] [CrossRef]
- Gonzales, D.S.; Pinheiro, J.C.; Bavister, B.D. Prediction of the developmental potential of hamster embryos in vitro by precise timing of the third cell cycle. J. Reprod. Fertil. 1995, 105, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Karagenc, L.; Sertkaya, Z.; Ciray, N.; Ulug, U.; Bahceci, M. Impact of oxygen concentration on embryonic development of mouse zygotes. Reprod. Biomed. Online 2004, 9, 409–417. [Google Scholar] [CrossRef]
- Liu, Z.; Xiang, G.; Xu, K.; Che, J.; Xu, C.; Li, K.; Wang, B.; Mu, Y. Transcriptome Analyses Reveal Differential Transcriptional Profiles in Early- and Late-Dividing Porcine Somatic Cell Nuclear Transfer Embryos. Genes 2020, 11, 1499. [Google Scholar] [CrossRef]
- Milewski, R.; Szpila, M.; Ajduk, A. Dynamics of cytoplasm and cleavage divisions correlates with preimplantation embryo development. Soc. Reprod. Fertil. 2018, 155, 1–14. [Google Scholar] [CrossRef]
- Vandaele, L.; Mateusen, B.; Maes, D.G.D.; Kruif, A.D.; Soom, A.V. Temporal detection of caspase-3 and -7 in bovine in vitro produced embryos of different developmental capacity. Soc. Reprod. Fertil. 2007, 133, 709–718. [Google Scholar] [CrossRef] [Green Version]
- Jiao, Y.; Gao, B.; Wang, G.; Li, H.; Ahmed, J.Z.; Zhang, D.; Ye, S.; Liu, S.; Li, M.; Shi, D.; et al. The key long non-coding RNA screening and validation between germinal vesicle and metaphase II of porcine oocyte in vitro maturation. Reprod. Domest. Anim. 2020, 55, 351–363. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Y.; Zhu, S.; Li, J.; Ahmed, J.Z.; Li, M.; Huang, B. PS48 promotes in vitro maturation and developmental competence of porcine oocytes through activating PI3K/Akt signalling pathway. Reprod. Domest. Anim. 2020, 55, 1678–1687. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Wang, B.; Wang, K.; Fan, Y.; Chen, Y.; Ye, C.; Feng, M.; Li, L.; Zhang, S.; Wei, H. Investigation into the Relationship Between Sperm Cysteine-Rich Secretory Protein 2 (CRISP2) and Sperm Fertilizing Ability and Fertility of Boars. Front. Vet. Sci. 2021, 8, 653413. [Google Scholar] [CrossRef] [PubMed]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650–1667. [Google Scholar] [CrossRef] [PubMed]
- Basak Balaban, B.U. Embryo culture as a diagnostic tool. RBMOnline 2003, 6, 671–682. [Google Scholar] [CrossRef]
- Nagy, Z.P.; Dozortsev, D.; Diamond, M.; Rienzi, L.; Ubaldi, F.; Greco, E. Pronuclear morphology evaluation with subsequent evaluation of embryo morphology significantly increases implantation rates. Fertil. Steril. 2003, 80, 67–74. [Google Scholar] [CrossRef]
- Arce, J.C.; Ziebe, S.; Lundin, K.; Janssens, R.; Helmgaard, L.; Sørensen, P. Interobserver agreement and intraobserver reproducibility of embryo quality assessments. Hum. Reprod. 2006, 21, 2141–2148. [Google Scholar] [CrossRef] [Green Version]
- Baxter Bendus, A.E.; Mayer, J.F.; Shipley, S.K.; Catherino, W.H. Interobserver and intraobserver variation in day 3 embryo grading. Fertil. Steril. 2006, 86, 1608–1615. [Google Scholar] [CrossRef]
- Paternot, G.; Devroe, J.; Debrock, S.; D’Hooghe, T.M.; Spiessens, C. Intra- and inter-observer analysis in the morphological assessment of early-stage embryos. Reprod. Biol. Endocrinol. 2009, 7, 105. [Google Scholar] [CrossRef] [Green Version]
- Lemmen, J.; Agerholm, S.Z. Kinetic markers of human embryo quality using time-lapse recordings of IVF/ICSI-fertilized oocytes. Rbmonline 2008, 17, 385–391. [Google Scholar] [CrossRef]
- Wong, C.C.; Loewke, K.E.; Bossert, N.L.; Behr, B.; Jonge, C.J.D.; Pera, R.A.R. Non-invasive imaging of human embryos before embryonic genome activation predicts development to the blastocyst stage. Nat. Biotechnol. 2010, 28, 1115–1121. [Google Scholar] [CrossRef]
- Racowsky, C.; Kovacs, P.; Martins, W.P. A critical appraisal of time-lapse imaging for embryo selection: Where are we and where do we need to go? J. Assist. Reprod. Genet. 2015, 32, 1025–1030. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.H.; Day, M.L. Egg timers: How is developmental time measured in the early vertebrate embryo? Bioessays 2000, 22, 57–63. [Google Scholar] [CrossRef]
- Yuan, F.; Xie, Q.; Wu, J.; Bai, Y.; Mao, B.; Dong, Y.; Bi, W.; Ji, G.; Tao, W.; Wang, Y.; et al. MST1 promotes apoptosis through regulating Sirt1-dependent p53 deacetylation. J. Biol. Chem. 2011, 286, 6940–6945. [Google Scholar] [CrossRef] [Green Version]
- Platonova, O.; Akey, I.V.; Head, J.F.; Akey, C.W. Crystal structure and function of human nucleoplasmin (npm2): A histone chaperone in oocytes and embryos. Biochemistry 2011, 50, 8078–8089. [Google Scholar] [CrossRef] [Green Version]
- Pan, W.; Wang, L.; Zhang, X.F.; Zhang, H.; Zhang, J.; Xu, P.; Zhang, Y.; Hu, P.; Zhang, X.-D.; Du, R.-L.; et al. Hypoxia-induced microRNA-191 contributes to hepatic ischemia/reperfusion injury through the ZONAB/Cyclin D1 axis. Cell Death Differ. 2019, 26, 291–305. [Google Scholar] [CrossRef]
- Zheng, Q.; Zheng, X.; Zhang, L.; Luo, H.; Qian, L.; Liu, Y.; Gao, Y.; Niu, M.; Meng, J.; Zhang, M.; et al. The Neuron-Specific Protein TMEM59L Mediates Oxidative Stress-Induced Cell Death. Mol. Neurobiol. 2017, 54, 4189–4200. [Google Scholar] [CrossRef] [Green Version]
- Fan, Z.; Beresford, P.J.; Oh, D.Y.; Zhang, D.; Lieberman, J. Tumor Suppressor NM23-H1 Is a Granzyme A-Activated DNase during CTL-Mediated Apoptosis, and the Nucleosome Assembly Protein SET Is Its Inhibitor. Cell 2003, 112, 659–672. [Google Scholar] [CrossRef] [Green Version]
- Ono, T.; Losada, A.; Hirano, M.; Myers, M.P.; Neuwald, A.F.; Hirano, T. Differential Contributions of Condensin I and Condensin II to Mitotic Chromosome Architecture in Vertebrate Cells. Cell 2003, 115, 109–121. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cleavage Time | Cleavage Number | Blastocyst Number | Cleavage Rate (%) * | Blastocyst Rate (%) * |
|---|---|---|---|---|
| 0–16 h | 2.75 ± 0.25 | 1.75 ± 0.25 | 1.82 ± 0.23 | 62.50 ± 4.17 |
| 16–18 h | 7.00 ± 1.35 | 5.75 ± 1.31 | 4.42 ± 0.77 | 79.52 ± 5.30 |
| 18–20 h | 22.50 ± 5.52 | 10.50 ± 2.47 | 14.58 ± 3.93 | 49.30 ± 6.02 |
| 20–22 h | 48.25 ± 9.76 | 23.00 ± 4.36 | 29.95 ± 2.15 | 48.20 ± 1.39 |
| 22–24 h | 32.50 ± 8.27 | 12.00 ± 2.94 | 20.02 ± 2.88 | 37.15 ± 1.53 |
| 24–26 h | 13.00 ± 4.24 | 5.50 ± 1.85 | 7.50 ± 1.43 | 43.31 ± 6.46 |
| 26–28 h | 8.50 ± 2.02 | 3.25 ± 1.11 | 5.14 ± 0.44 | 35.83 ± 6.29 |
| 28–30 h | 6.50 ± 1.85 | 1.75 ± 0.48 | 4.04 ± 0.68 | 27.50 ± 4.33 |
| 30–32 h | 4.00 ± 0.00 | 0.50 ± 0.29 | 2.74 ± 0.47 | 12.50 ± 7.22 |
| 32–42 h | 10.50 ± 0.87 | 1.00 ± 0.00 | 7.37 ± 1.77 | 9.70 ± 0.72 |
| 42–48 h | 3.75 ± 0.48 | 0.00 ± 0.00 | 2.43 ± 0.19 | 0.00 ± 0.00 |
| Content | PA | IVF | ||
|---|---|---|---|---|
| EC | LC | EC | LC | |
| Number of cleavage embryos | 77.8 ± 14.0 a | 19.0 ± 3.5 b | 20.3 ± 3.5 | 11.3 ± 0.9 |
| Cleavage rate (%) | 48.9 ± 4.2 a | 11.9 ± 0.7 b | 41.7 ± 4.2 | 28.6 ± 4.7 |
| Number of blastocysts | 39.3 ± 6.0 a | 5.5 ± 1.8 b | 5.3 ± 1.9 | 1.7 ± 0.7 |
| Blastocyst rate (%) | 51.2 ± 2.2 a | 27.2 ± 6.0 b | 28.9 ± 3.9 a | 10.7 ± 1.9 b |
| Primary blastocyst cell number | 44.5 ± 4.2 a | 33.5 ± 2.7 b | ||
| Initial blastocyst cell number | 49.7 ± 4.1 a | 40.3 ± 3.3 b | ||
| Expanded blastocyst cell number | 74.8 ± 6.4 a | 50.7 ± 3.7 b | ||
| Hatching blastocyst cell number | 65.0 ± 5.6 | — | ||
| Cleavage Time | Cleavage Number | Blastocyst Number | Cleavage Rate (%) | Blastocyst Rate (%) |
|---|---|---|---|---|
| 0–18 h | 2.00 ± 0.71 | 0.50 ± 0.29 | 3.85 ± 1.61 | 20.83 ± 12.50 |
| 18–20 h | 2.25 ± 1.31 | 0.50 ± 0.29 | 4.94 ± 2.45 | 16.67 ± 11.79 |
| 20–22 h | 5.50 ± 1.26 | 2.00 ± 1.08 | 11.35 ± 2.46 | 32.22 ± 11.71 |
| 22–24 h | 8.75 ± 1.84 | 2.00 ± 0.41 | 17.54 ± 3.31 | 23.98 ± 3.71 |
| 24–26 h | 6.00 ± 1.58 | 1.00 ± 0.41 | 11.33 ± 1.20 | 19.58 ± 7.08 |
| 26–28 h | 5.75 ± 1.93 | 0.75 ± 0.48 | 10.34 ± 1.60 | 14.58 ± 8.59 |
| 28–30 h | 5.75 ± 2.10 | 0.75 ± 0.25 | 10.67 ± 1.85 | 14.58 ± 6.25 |
| 30–32 h | 4.25 ± 0.75 | 1.25 ± 0.48 | 8.92 ± 1.98 | 18.33 ± 10.67 |
| 32–42 h | 5.50 ± 0.87 | 0.75 ± 0.48 | 11.94 ± 2.84 | 15.48 ± 5.87 |
| 42–48 h | 5.75 ± 3.22 | 0.25 ± 0.25 | 9.13 ± 3.77 | 6.25 ± 6.25 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, X.; Li, T.; Xiong, X.; Shan, H.; Feng, T.; Cui, K.; Shi, D.; Liu, Q.; Li, Z. RNA-Seq Reveals the Underlying Molecular Mechanism of First Cleavage Time Affecting Porcine Embryo Development. Genes 2022, 13, 1251. https://doi.org/10.3390/genes13071251
Song X, Li T, Xiong X, Shan H, Feng T, Cui K, Shi D, Liu Q, Li Z. RNA-Seq Reveals the Underlying Molecular Mechanism of First Cleavage Time Affecting Porcine Embryo Development. Genes. 2022; 13(7):1251. https://doi.org/10.3390/genes13071251
Chicago/Turabian StyleSong, Xinhui, Tiantian Li, Xin Xiong, Huiquan Shan, Tong Feng, Kuiqing Cui, Deshun Shi, Qingyou Liu, and Zhipeng Li. 2022. "RNA-Seq Reveals the Underlying Molecular Mechanism of First Cleavage Time Affecting Porcine Embryo Development" Genes 13, no. 7: 1251. https://doi.org/10.3390/genes13071251