
Genome-Wide Analysis of the WRKY Gene Family in Malus domestica and the Role of MdWRKY70L in Response to Drought and Salt Stresses

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification and Comprehensive Analysis of the WRKY Family Members in the Apple Genome
2.2. Plant Materials and Stress Treatments
2.3. Subcellular Localization Analysis of MdWRKYs
2.4. Genetic Transformation of MdWRKY70L
2.5. Resistance Analysis of Transgenic Tobacco
2.6. Quantitative PCR (qPCR)
2.7. Statistical Analysis
3. Results
3.1. Identification of WRKY Genes in Apple Genome
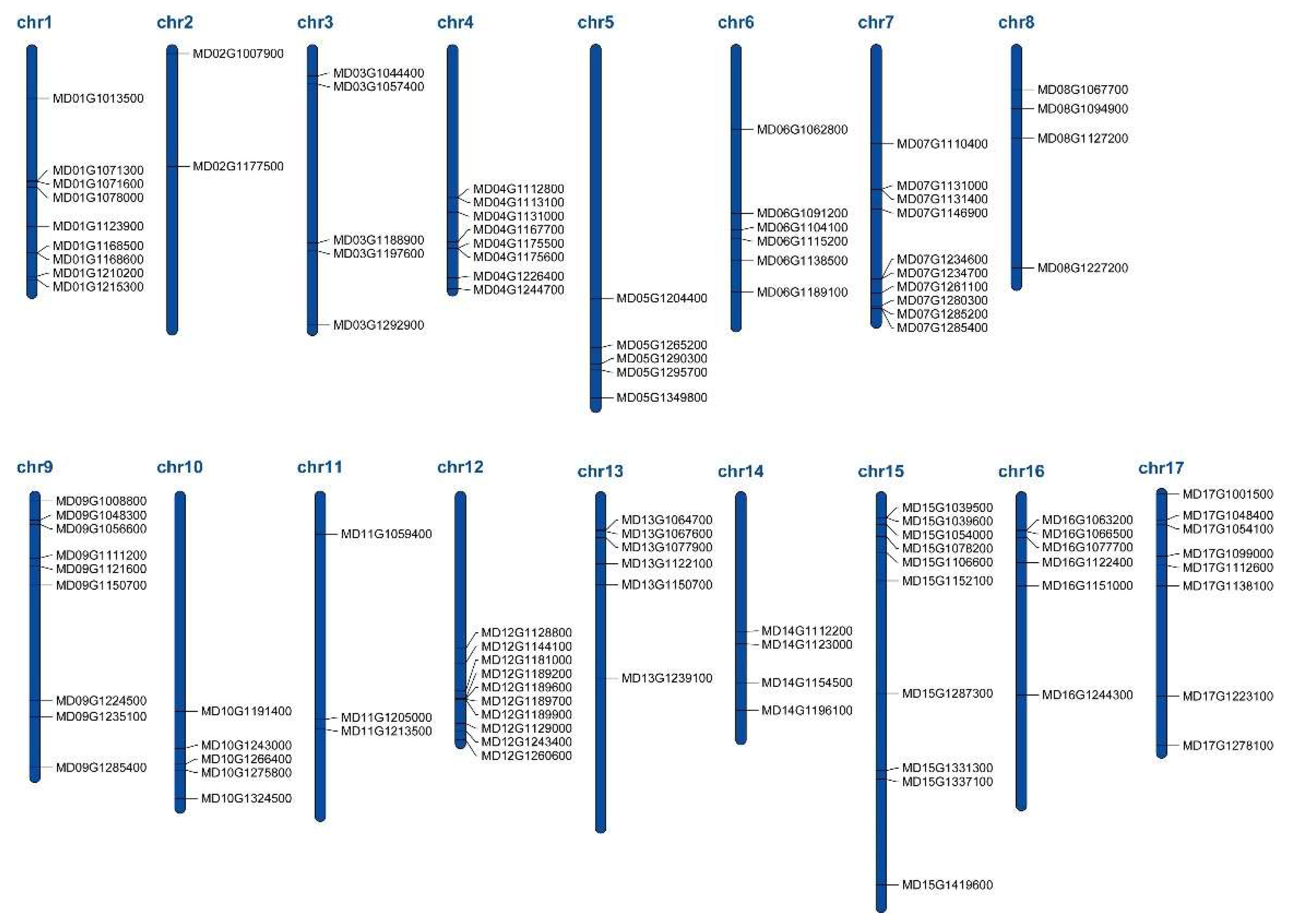
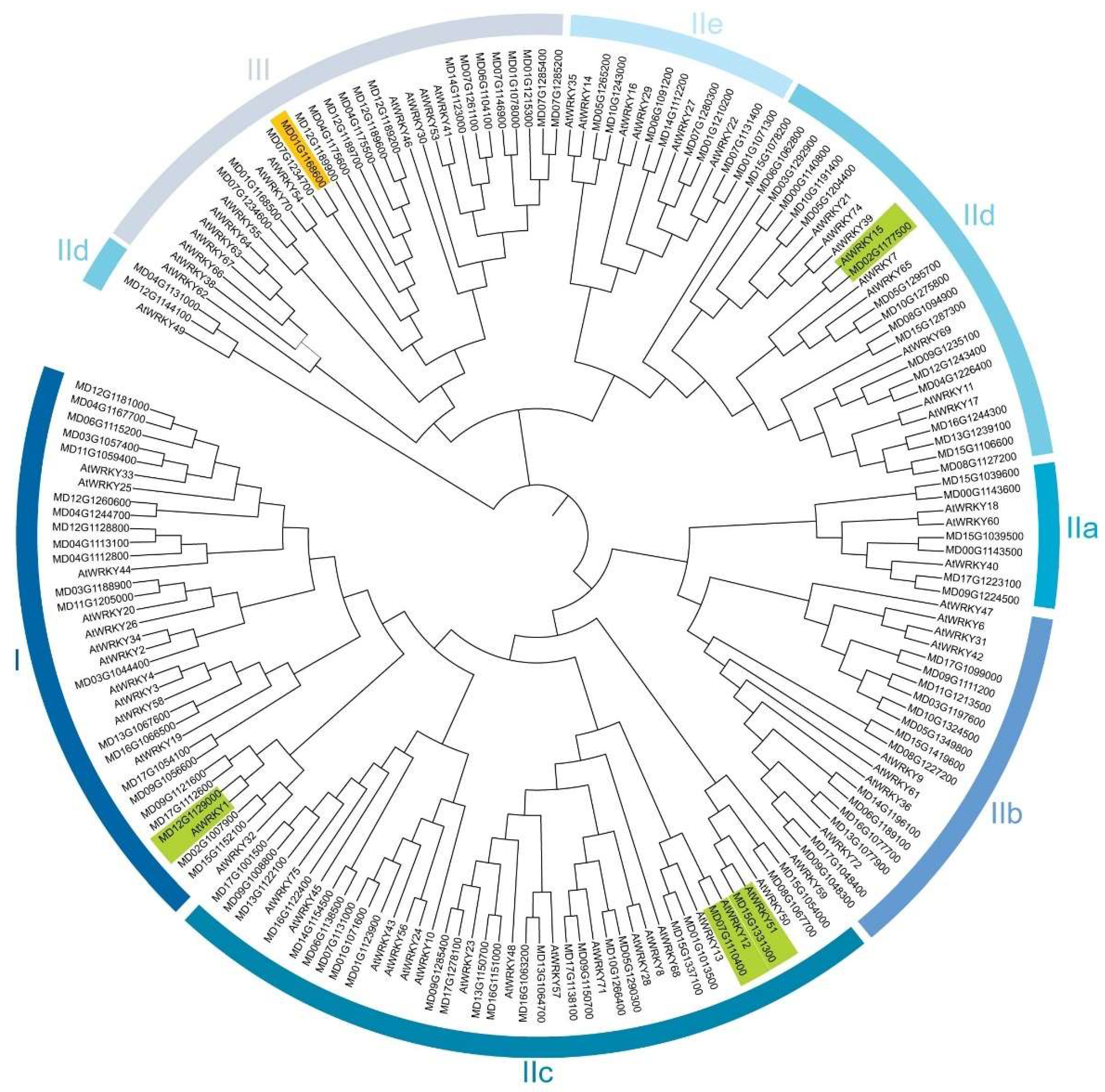
3.2. Chromosome Distribution and Evolutionary Analysis of the WRKY Sequences
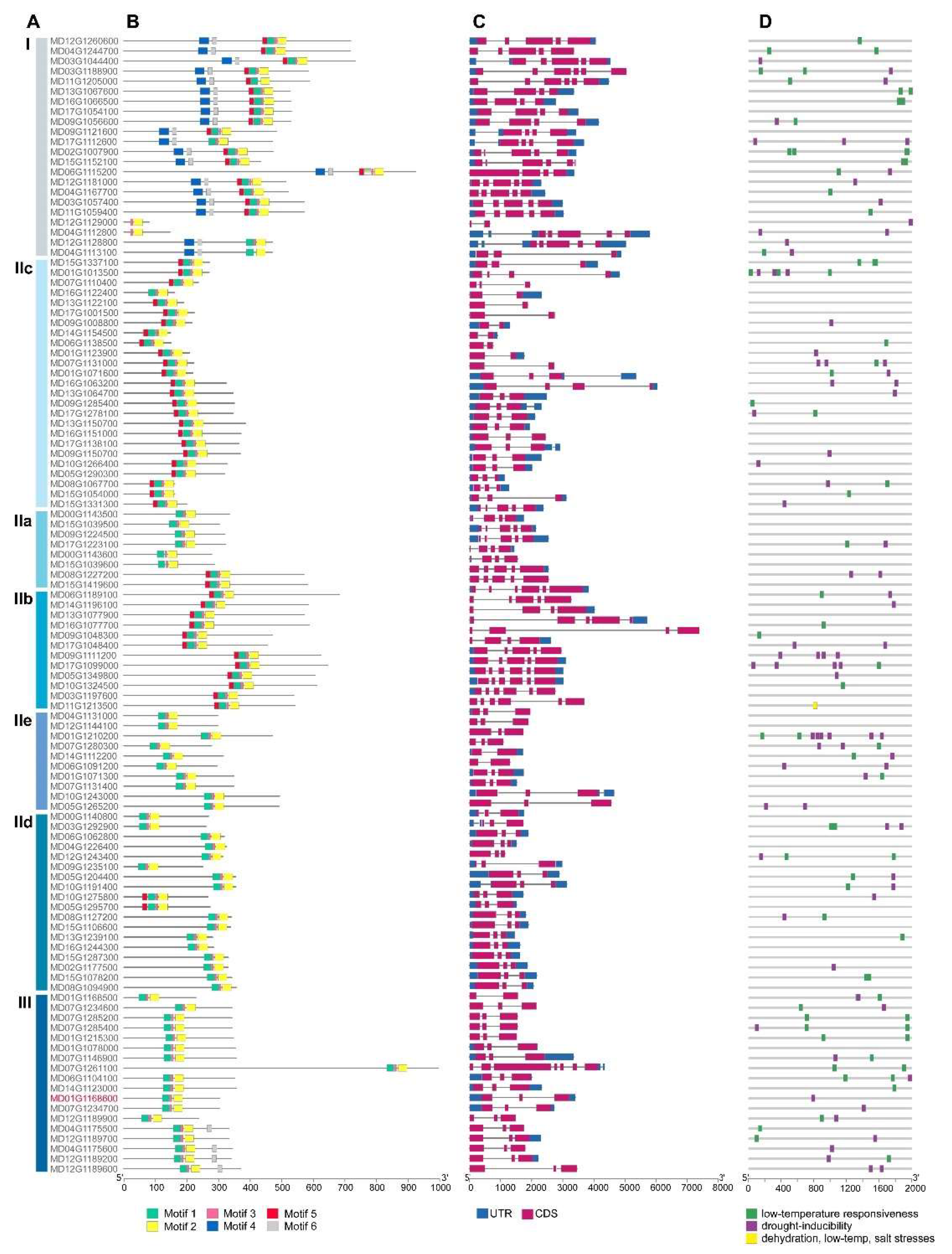
3.3. Structural Analyses of the WRKY Sequences
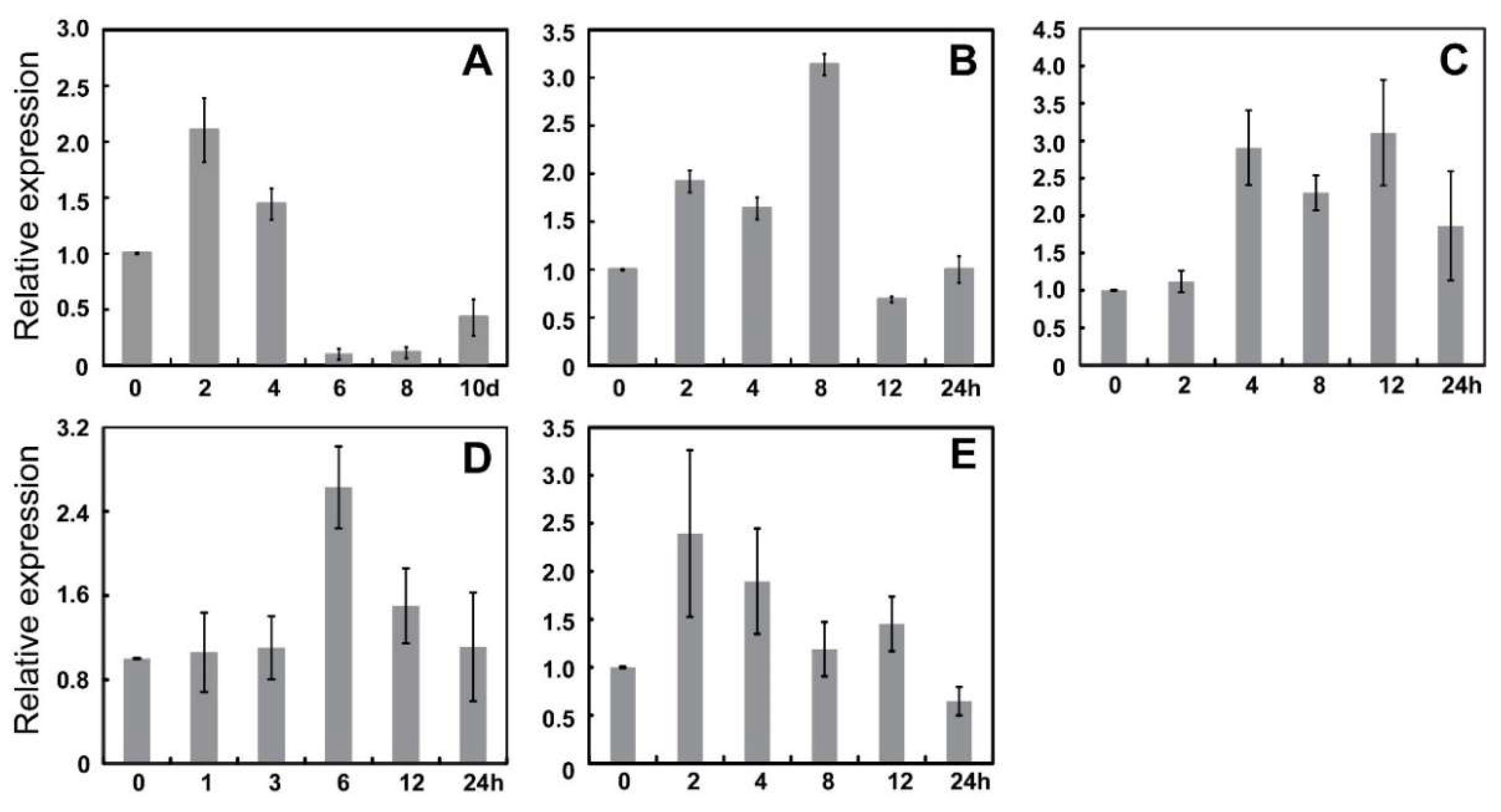
3.4. Expression Profile of MdWRKY Genes
3.5. Subcellular Localization of MdWRKYs Proteins
3.6. MdWRKY70L Enhanced Drought and Salt Stress Tolerance in Transgenic Tobacco Plants
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shepherd, T.; Griffiths, D.W. The effects of stress on plant cuticular waxes. New Phytol. 2006, 171, 469–499. [Google Scholar] [CrossRef]
- Zhang, A.; Liu, M.; Gu, W.; Chen, Z.; Gu, Y.; Pei, L.; Tian, R. Effect of drought on photosynthesis, total antioxidant capacity, bioactive component accumulation, and the transcriptome of Atractylodes lancea. BMC Plant Biol. 2021, 21, 293. [Google Scholar] [CrossRef] [PubMed]
- Khaleghnezhad, V.; Yousefi, A.R.; Tavakoli, A.; Farajmand, B.; Mastinu, A. Concentrations-dependent effect of exogenous abscisic acid on photosynthesis, growth and phenolic content of Dracocephalum moldavica L. under drought stress. Planta 2021, 253, 127. [Google Scholar] [CrossRef] [PubMed]
- Huo, L.; Guo, Z.; Jia, X.; Sun, X.; Wang, P.; Gong, X.; Ma, F. Increased autophagic activity in roots caused by overexpression of the autophagy-related gene MdATG10 in apple enhances salt tolerance. Plant Sci. 2020, 294, 110444. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Ma, Z.; Hu, L.; Huang, K.; Zhang, M.; Zhang, S.; Jiang, W.; Wu, T.; Du, X. Involvement of rice transcription factor OsERF19 in response to ABA and salt stress responses. Plant Physiol. Biochem. 2021, 167, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Gupta, B.; Huang, B. Mechanism of salinity tolerance in plants: Physiological, biochemical, and molecular characterization. Int. J. Genom. 2014, 2014, 701596. [Google Scholar] [CrossRef] [PubMed]
- Deinlein, U.; Stephan, A.B.; Horie, T.; Luo, W.; Schroeder, J.I. Plant salt-tolerance mechanisms. Trends Plant Sci. 2014, 19, 371–379. [Google Scholar] [CrossRef] [Green Version]
- Wani, S.H.; Anand, S.; Singh, B.; Bohra, A.; Joshi, R. WRKY transcription factors and plant defense responses: Latest discoveries and future prospects. Plant Cell Rep. 2021, 40, 1071–1085. [Google Scholar] [CrossRef]
- Wang, L.; Zhu, W.; Fang, L.; Sun, X.; Su, L.; Liang, Z.; Wang, N.; Londo, J.P.; Li, S.; Xin, H. Genome-wide identification of WRKY family genes and their response to cold stress in Vitis vinifera. BMC Plant Biol. 2014, 14, 103. [Google Scholar] [CrossRef] [Green Version]
- Rushton, P.J.; Somssich, I.E.; Ringler, P.; Shen, Q.J. WRKY transcription factors. Trends Plant Sci. 2010, 15, 247–258. [Google Scholar] [CrossRef]
- Eulgem, T.; Rushton, P.J.; Robatzek, S.; Somssich, I.E. The WRKY superfamily of plant transcription factors. Trends Plant Sci. 2000, 5, 199–206. [Google Scholar] [CrossRef]
- Li, H.L.; Zhang, L.B.; Guo, D.; Li, C.Z.; Peng, S.Q. Identification and expression profiles of the WRKY transcription factor family in Ricinus communis. Gene 2012, 503, 248–253. [Google Scholar] [CrossRef] [PubMed]
- Xiong, W.; Xu, X.; Zhang, L.; Wu, P.; Chen, Y.; Li, M.; Jiang, H.; Wu, G. Genome-wide analysis of the WRKY gene family in physic nut (Jatropha curcas L.). Gene 2013, 524, 124–132. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Yang, J.; Li, G.; Sun, Z.; Hu, S.; Chen, Y.; Guo, W.; Hou, H. Genome-wide identification and comparative analysis of the WRKY gene family in aquatic plants and their response to abiotic stresses in giant duckweed (Spirodela polyrhiza). Genomics 2021, 113, 1761–1777. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Pang, S.; Lu, Z.; Jin, B. Function and mechanism of WRKY transcription factors in abiotic stress responses of plants. Plants 2020, 9, 1515. [Google Scholar] [CrossRef] [PubMed]
- Dong, Q.; Zheng, W.; Duan, D.; Huang, D.; Wang, Q.; Liu, C.; Li, C.; Gong, X.; Li, C.; Mao, K.; et al. MdWRKY30, a group IIa WRKY gene from apple, confers tolerance to salinity and osmotic stresses in transgenic apple callus and Arabidopsis seedlings. Plant Sci. 2020, 299, 110611. [Google Scholar] [CrossRef] [PubMed]
- Kuki, Y.; Ohno, R.; Yoshida, K.; Takumi, S. Heterologous expression of wheat WRKY transcription factor genes transcriptionally activated in hybrid necrosis strains alters abiotic and biotic stress tolerance in transgenic Arabidopsis. Plant Physiol. Biochem. 2020, 150, 71–79. [Google Scholar] [CrossRef]
- Lv, B.; Wu, Q.; Wang, A.; Li, Q.; Dong, Q.; Yang, J.; Zhao, H.; Wang, X.; Chen, H.; Li, C. A WRKY transcription factor, FtWRKY46, from Tartary buckwheat improves salt tolerance in transgenic Arabidopsis thaliana. Plant Physiol. Biochem. 2020, 147, 43–53. [Google Scholar] [CrossRef]
- Zhu, H.; Zhou, Y.; Zhai, H.; He, S.; Zhao, N.; Liu, Q. A novel sweet potato WRKY transcription factor, IbWRKY2, positively regulates drought and salt tolerance in transgenic Arabidopsis. Biomolecules 2020, 10, 506. [Google Scholar] [CrossRef] [Green Version]
- Kang, G.; Yan, D.; Chen, X.; Li, Y.; Yang, L.; Zeng, R. Molecular characterization and functional analysis of a novel WRKY transcription factor HbWRKY83 possibly involved in rubber production of Hevea brasiliensis. Plant Physiol. Biochem. 2020, 155, 483–493. [Google Scholar] [CrossRef]
- Gao, Y.F.; Liu, J.K.; Yang, F.M.; Zhang, G.Y.; Wang, D.; Zhang, L.; Ou, Y.B.; Yao, Y.A. The WRKY transcription factor WRKY8 promotes resistance to pathogen infection and mediates drought and salt stress tolerance in Solanum lycopersicum. Physiol. Plant. 2020, 168, 98–117. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Jiang, Y.; Guo, Y.; Huang, J.; Zhou, M.; Tang, Y.; Sui, J.; Wang, J.; Qiao, L. A novel salt inducible WRKY transcription factor gene, AhWRKY75, confers salt tolerance in transgenic peanut. Plant Physiol. Biochem. 2021, 160, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Yao, Q.Y.; Xia, E.H.; Liu, F.H.; Gao, L.Z. Genome-wide identification and comparative expression analysis reveal a rapid expansion and functional divergence of duplicated genes in the WRKY gene family of cabbage, Brassica oleracea var. capitata. Gene 2015, 557, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Zou, X.H.; Dong, C.; Liu, H.T.; Gao, Q.H. Genome-wide characterization and expression analysis of WRKY family genes during development and resistance to Colletotrichum fructicola in cultivated strawberry (Fragaria×ananassa Duch.). J. Integr. Agric. 2022, 21, 1658–1672. [Google Scholar] [CrossRef]
- Meng, D.; Li, Y.; Bai, Y.; Li, M.; Cheng, L. Genome-wide identification and characterization of WRKY transcriptional factor family in apple and analysis of their responses to waterlogging and drought stress. Plant Physiol. Biochem. 2016, 103, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Gong, X.; Zhang, J.; Hu, J.; Wang, W.; Wu, H.; Zhang, Q.; Liu, J.H. FcWRKY70, a WRKY protein of Fortunella crassifolia, functions in drought tolerance and modulates putrescine synthesis by regulating arginine decarboxylase gene. Plant Cell Environ. 2015, 38, 2248–2262. [Google Scholar] [CrossRef]
- Horsch, R. A simple and general method for transferring genes into plants. Science 1985, 227, 1229–1231. [Google Scholar] [CrossRef]
- Liang, B.; Li, C.; Ma, C.; Wei, Z.; Wang, Q.; Huang, D.; Chen, Q.; Li, C.; Ma, F. Dopamine alleviates nutrient deficiency-induced stress in Malus hupehensis. Plant Physiol. Biochem. 2017, 119, 346–359. [Google Scholar] [CrossRef]
- Chen, X.; Li, C.; Wang, H.; Guo, Z.J. WRKY transcription factors: Evolution, binding, and action. Phytopathol. Res. 2019, 1, 13. [Google Scholar] [CrossRef]
- Eulgem, T.; Somssich, I.E. Networks of WRKY transcription factors in defense signaling. Curr. Opin. Plant Biol. 2007, 10, 366–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ross, C.A.; Yue, L.; Shen, Q.J. The WRKY gene family in rice (Oryza sativa). J. Integr. Plant Biol. 2010, 49, 827–842. [Google Scholar] [CrossRef]
- Pandey, S.P.; Somssich, I.E. The role of WRKY transcription factors in plant immunity. Plant Physiol. 2009, 150, 1648–1655. [Google Scholar] [CrossRef] [Green Version]
- Yuan, H.; Guo, W.; Zhao, L.; Yu, Y.; Chen, S.; Tao, L.; Cheng, L.; Kang, Q.; Song, X.; Wu, J.; et al. Genome-wide identification and expression analysis of the WRKY transcription factor family in flax (Linum usitatissimum L.). BMC Genom. 2021, 22, 375. [Google Scholar] [CrossRef]
- Liu, J.; Wang, X.; Chen, Y.; Liu, Y.; Wu, Y.; Ren, S.; Li, L. Identification, evolution and expression analysis of WRKY gene family in Eucommia ulmoides. Genomics 2021, 113, 3294–3309. [Google Scholar] [CrossRef]
- Chen, Y.; Jing, X.; Wang, S.; Wang, J.; Zhang, S.; Shi, Q. Genome-wide analysis of WRKY transcription factor family in melon (cucumis melo L.) and their response to powdery mildew. Plant Mol. Biol. Rep. 2021, 39, 686–699. [Google Scholar] [CrossRef]
- Zheng, J.; Zhang, Z.; Tong, T.; Fang, Y.; Zhang, X. Genome-wide identification of WRKY gene family and expression analysis under abiotic stress in barley. Agronomy 2021, 11, 521. [Google Scholar] [CrossRef]
- Qu, R.; Cao, Y.; Tang, X.; Sun, L.; Wei, L.; Wang, K. Identification and expression analysis of the WRKY gene family in Isatis indigotica. Gene 2021, 783, 145561. [Google Scholar] [CrossRef]
- Lee, F.C.; Yeap, W.C.; Appleton, D.R.; Ho, C.L.; Kulaveerasingam, H. Identification of drought responsive Elaeis guineensis WRKY transcription factors with sensitivity to other abiotic stresses and hormone treatments. BMC Genom. 2022, 26, 164. [Google Scholar] [CrossRef]
- Finatto, T.; Viana, V.E.; Woyann, L.G.; Busanello, C.; Maia, L.; Oliveira, A.C. Can WRKY transcription factors help plants to overcome environmental challenges? Genet. Mol. Biol. 2018, 41, 533–544. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Song, Y.; Li, S.; Zhang, L.; Zou, C.; Yu, D. The role of WRKY transcription factors in plant abiotic stresses. Biochim. Biophys. Acta 2012, 1819, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Chen, Q.; Chen, W.; Liu, X.; Xia, Y.; Guo, Q.; Jing, D.; Liang, G. A WRKY transcription factor, EjWRKY17, from Eriobotrya japonica enhances drought tolerance in transgenic Arabidopsis. Int. J. Mol. Sci. 2021, 22, 5593. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Tang, Y.; Zhou, C.; Zhang, L.; Lv, J. A wheat WRKY transcription factor TaWRKY46 enhances tolerance to osmotic stress in transgenic Arabidopsis plants. Int. J. Mol. Sci. 2020, 21, 1321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.; Zhang, G.Y.; Chen, S.Y. Structure and regulation of plant transcription factors. Chin. Sci. Bull. 2000, 45, 1465–1474. [Google Scholar]
- Wang, R.; Wang, R.; Liu, M.; Yuan, W.; Zhao, Z.; Liu, X.; Peng, Y.; Yang, X.; Sun, Y.; Tang, W. Nucleocytoplasmic trafficking and turnover mechanisms of BRASSINAZOLE RESRSTANT 1 in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2021, 118, e2101838118. [Google Scholar] [CrossRef]
- Trofimov, K.; Ivanov, R.; Eutebach, M.; Acaroglu, B.; Mohr, I.; Bauer, P.; Brumbarova, T. Mobility and localization of the iron deficiency-induced transcription factor bHLH039 change in the presence of FIT. Plant Direct 2019, 3, e00190. [Google Scholar] [CrossRef] [Green Version]
- Monteiro de Almeida, S.D.; Jordão do Amaral, O.D.; Del-Bem, L.E.; Bronze Dos Santos, E.; Silva, R.J.S.; Gramacho, K.P.; Vincentz, M.; Micheli, F. Genome-wide identification and characterization of cacao WRKY transcription factors and analysis of their expression in response to witches’ broom disease. PLoS ONE 2017, 12, e0187346. [Google Scholar]
- Tang, Y.; Guo, J.; Zhang, T.; Bai, S.; He, K.; Wang, Z. Genome-wide analysis of WRKY gene family and the dynamic responses of key WRKY genes involved in Ostrinia furnacalis attack in Zea mays. Int. J. Mol. Sci. 2021, 22, 13045. [Google Scholar] [CrossRef]
- Yue, M.L.; Jiang, L.Y.; Liu, Y.; Li, L.; Liu, Y.Q.; Chen, Q.; Tang, H.R. Cloning, subcellular localization and expression characterization of strawberry FaWRKY31. Acta Hortic. Sin. 2019, 46, 1947–1959. [Google Scholar]
- Chu, X.; Wang, C.; Chen, X.; Lu, W.; Li, H.; Wang, X.; Hao, L.; Guo, X. The cotton WRKY gene GhWRKY41 positively regulates salt and drought stress tolerance in transgenic Nicotiana benthamiana. PLoS ONE 2016, 11, e0157026. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Nolan, T.M.; Ye, H.; Zhang, M.; Tong, H.; Xin, P.; Chu, J.; Chu, C.; Li, Z.; Yin, Y. Arabidopsis WRKY46, WRKY54, and WRKY70 transcription factors are involved in brassinosteroid-regulated plant growth and drought Responses. Plant Cell 2017, 29, 1425–1439. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Zhou, K.; Liu, Y.; Yang, S.; Ma, F. Overexpression of MdMIPS1 enhances salt tolerance by improving osmosis, ion balance, and antioxidant activity in transgenic apple. Plant Sci. 2020, 301, 110654. [Google Scholar] [CrossRef] [PubMed]
- Golldack, D.; Li, C.; Mohan, H.; Probst, N. Tolerance to drought and salt stress in plants: Unraveling the signaling networks. Front. Plant Sci. 2014, 5, 151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.; Chi, X.; Guo, F.; Jin, X.; Luo, H.; Hawar, A.; Chen, Y.; Feng, K.; Wang, B.; Qi, J.; et al. SbWRKY30 enhances the drought tolerance of plants and regulates a drought stress-responsive gene, SbRD19, in sorghum. J. Plant Physiol. 2020, 246–247, 153142. [Google Scholar] [CrossRef] [PubMed]
- Abu-Ghosh, S.; Iluz, D.; Dubinsky, Z.; Miller, G. Exogenous abscisic acid confers salinity tolerance in Chlamydomonas reinhardtii during its life cycle. J. Phycol. 2021, 57, 1323–1334. [Google Scholar] [CrossRef] [PubMed]
- Gai, Z.; Wang, Y.; Ding, Y.; Qian, W.; Qiu, C.; Xie, H.; Sun, L.; Jiang, Z.; Ma, Q.; Wang, L.; et al. Exogenous abscisic acid induces the lipid and flavonoid metabolism of tea plants under drought stress. Sci. Rep. 2020, 10, 12275. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Li, C.; He, X.; Zhang, X.; Zhu, L. ABA signaling is negatively regulated by GbWRKY1 through JAZ1 and ABI1 to affect salt and drought tolerance. Plant Cell Rep. 2020, 39, 181–194. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Deyholos, M.K. Functional characterization of Arabidopsis NaCl-inducible WRKY25 and WRKY33 transcription factors in abiotic stresses. Plant Mol. Biol. 2009, 69, 91–105. [Google Scholar] [CrossRef] [PubMed]
- Dai, W.; Wang, M.; Gong, X.; Liu, J. The transcription factor FcWRKY40 of Fortunella crassifolia functions positively in salt tolerance through modulation of ion homeostasis and proline biosynthesis by directly regulating SOS2 and P5CS1 homologs. New Phytol. 2018, 219, 972–989. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Genome No. | Chromosome No. | CDS(bp) | Amino Acid | MW (kDa) | Ip | Subcellular localization | Duplications |
|---|---|---|---|---|---|---|---|---|
| I | MD03G1057400 | Chr03:4579079.4582074 | 1716 | 571 | 62.5 | 7.02 | Nuclear | WGD/segmental |
| MD11G1059400 | Chr11:5068503.5071526 | 1719 | 572 | 62.9 | 6.77 | Nuclear | WGD/segmental | |
| MD12G1181000 | Chr12:26084482.26086791 | 1539 | 512 | 56.7 | 6.73 | Nuclear | WGD/segmental | |
| MD04G1167700 | Chr04:25792146.25794577 | 1563 | 520 | 57.7 | 7.10 | Nuclear | WGD/segmental | |
| MD06G1115200 | Chr06:25425907.25429273 | 2775 | 924 | 102.8 | 5.33 | Nuclear | proximal | |
| MD12G1260600 | Chr12:32647306.32651367 | 2157 | 718 | 79.0 | 6.40 | Nuclear | WGD/segmental | |
| MD04G1244700 | Chr04:32067852.32071212 | 2154 | 717 | 78.5 | 5.99 | Nuclear | WGD/segmental | |
| MD12G1128800 | Chr12:20397217.20403016 | 1413 | 470 | 51.5 | 9.00 | Nuclear | WGD/segmental | |
| MD04G1113100 | Chr04:19846116.19851149 | 1413 | 470 | 51.5 | 8.93 | Nuclear | proximal | |
| MD03G1188900 | Chr03:25924164.25929514 | 1755 | 584 | 63.9 | 5.97 | Nuclear | WGD/segmental | |
| MD11G1205000 | Chr11:29914448.29918933 | 1767 | 588 | 64.0 | 5.86 | Nuclear | WGD/segmental | |
| MD03G1044400 | Chr03:3511777.3516305 | 2199 | 732 | 79.5 | 5.96 | Nuclear | dispersed | |
| MD13G1067600 | Chr13:4637048.4640408 | 1581 | 526 | 57.3 | 7.26 | Nuclear | WGD/segmental | |
| MD16G1066500 | Chr16:4642014.4644789 | 1587 | 528 | 57.2 | 8.38 | Nuclear | WGD/segmental | |
| MD17G1054100 | Chr17:4198534.4202033 | 1593 | 530 | 57.4 | 8.39 | Nuclear | WGD/segmental | |
| MD09G1056600 | Chr09:3741354.3745516 | 1587 | 528 | 57.6 | 8.62 | Nuclear | WGD/segmental | |
| MD09G1121600 | Chr09:9379281.9382725 | 1455 | 484 | 53.2 | 5.93 | Nuclear | WGD/segmental | |
| MD17G1112600 | Chr17:9643152.9646837 | 1416 | 471 | 51.7 | 6.64 | Nuclear | WGD/segmental | |
| MD02G1007900 | Chr02:496443.499890 | 1422 | 473 | 52.0 | 8.82 | Nuclear | WGD/segmental | |
| MD15G1152100 | Chr15:11267047.11270466 | 1305 | 434 | 48.1 | 7.27 | Nuclear | WGD/segmental | |
| IIa | MD15G1039600 | Chr15:2798151.2799702 | 861 | 286 | 32.1 | 8.15 | Nuclear | tandem |
| MD00G1143600 | Chr00:31240990.31242443 | 837 | 278 | 31.3 | 8.85 | Nuclear | tandem | |
| MD15G1039500 | Chr15:2783151.2784898 | 909 | 302 | 33.6 | 7.10 | Nuclear | WGD/segmental | |
| MD00G1143500 | Chr00:31227045.31229422 | 1005 | 334 | 37.0 | 7.00 | Nuclear | tandem | |
| MD17G1223100 | Chr17:27209201.27211741 | 966 | 321 | 35.7 | 8.20 | Nuclear | WGD/segmental | |
| MD09G1224500 | Chr09:27404228.27406362 | 963 | 320 | 35.3 | 7.59 | Nuclear | WGD/segmental | |
| MD04G1112800 | Chr04:19827951.19828792 | 441 | 146 | 16.7 | 9.60 | Extracellular | WGD/segmental | |
| MD12G1129000 | Chr12:30451746.30454869 | 243 | 80 | 9.3 | 9.85 | Cytoplasmic/ Nuclear/ Mitochondrial | proximal | |
| IIb | MD17G1099000 | Chr17:8402127.8405223 | 1938 | 645 | 70.0 | 6.42 | Nuclear | WGD/segmental |
| MD09G1111200 | Chr09:8267211.8270166 | 1878 | 625 | 68.2 | 6.33 | Nuclear | WGD/segmental | |
| MD05G1349800 | Chr05:46759691.46762704 | 1821 | 606 | 65.3 | 7.67 | Nuclear | WGD/segmental | |
| MD11G1213500 | Chr11:31232215.31235905 | 1626 | 541 | 59.0 | 6.45 | Nuclear | WGD/segmental | |
| MD03G1197600 | Chr03:27003520.27006281 | 1617 | 538 | 59.0 | 6.42 | Nuclear | WGD/segmental | |
| MD15G1419600 | Chr15:52086817.52089361 | 1749 | 582 | 64.2 | 5.10 | Nuclear | WGD/segmental | |
| MD08G1227200 | Chr08:29356105..29358646 | 1713 | 570 | 62.312 | 5.35 | Nuclear | WGD/segmental | |
| MD14G1196100 | Chr14:28655099.28658370 | 1755 | 584 | 63.3 | 6.48 | Nuclear | WGD/segmental | |
| MD06G1189100 | Chr06:32608598.32612425 | 2052 | 683 | 73.5 | 7.23 | Nuclear | WGD/segmental | |
| MD16G1077700 | Chr16:5438696.5444408 | 1764 | 587 | 64.1 | 6.43 | Nuclear | WGD/segmental | |
| MD13G1077900 | Chr13:5478407.5482438 | 1716 | 571 | 61.9 | 5.95 | Nuclear | WGD/segmental | |
| MD17G1048400 | Chr17:3528939.3531555 | 1368 | 455 | 50.0 | 7.59 | Nuclear | WGD/segmental | |
| MD09G1048300 | Chr09:3166930.3174316 | 1413 | 470 | 51.2 | 7.61 | Nuclear | WGD/segmental | |
| MD10G1324500 | Chr10:40512430.40515449 | 1836 | 611 | 65.7 | 6.61 | Nuclear | WGD/segmental | |
| IIc | MD15G1054000 | Chr15:3683177.3684465 | 486 | 161 | 18.2 | 5.65 | Nuclear | WGD/segmental |
| MD08G1067700 | Chr08:5387244.5388373 | 486 | 161 | 18.2 | 8.53 | Nuclear | WGD/segmental | |
| MD15G1331300 | Chr15:36715532.36718649 | 600 | 199 | 22.3 | 5.86 | Nuclear | dispersed | |
| MD07G1110400 | Chr07:12683567.12688395 | 711 | 236 | 26.8 | 8.17 | Nuclear | dispersed | |
| MD01G1013500 | Chr01:6515683.6519808 | 813 | 270 | 30.4 | 8.93 | Nuclear | WGD/segmental | |
| MD15G1337100 | Chr15:37891029.37895909 | 816 | 271 | 30.2 | 8.93 | Nuclear | WGD/segmental | |
| MD05G1290300 | Chr05:42212192.42214208 | 960 | 319 | 35.1 | 6.60 | Nuclear | WGD/segmental | |
| MD10G1266400 | Chr10:35935449.35937762 | 984 | 327 | 36.0 | 6.46 | Nuclear | WGD/segmental | |
| MD09G1150700 | Chr09:11892767.11895678 | 1110 | 369 | 41.4 | 7.75 | Nuclear | WGD/segmental | |
| MD17G1138100 | Chr17:12392128..12394598 | 1098 | 365 | 41.2 | 6.76 | Nuclear | WGD/segmental | |
| MD13G1064700 | Chr13:4465565.4471604 | 1041 | 346 | 37.7 | 6.59 | Nuclear | WGD/segmental | |
| MD16G1063200 | Chr16:4485285.4490649 | 975 | 324 | 35.5 | 6.60 | Nuclear | WGD/segmental | |
| MD16G1151000 | Chr16:11906129.11908067 | 1116 | 371 | 41.1 | 5.60 | Nuclear | WGD/segmental | |
| MD13G1150700 | Chr13:11807823.11809934 | 1158 | 385 | 42.7 | 6.11 | Nuclear | WGD/segmental | |
| MD17G1278100 | Chr17:33812003.33814316 | 1041 | 346 | 38.2 | 6.27 | Nuclear | WGD/segmental | |
| MD09G1285400 | Chr09:36364455.36366937 | 1053 | 350 | 38.4 | 5.85 | Nuclear | WGD/segmental | |
| MD01G1123900 | Chr01:23690319.23691083 | 627 | 208 | 23.6 | 8.91 | Nuclear | dispersed | |
| MD01G1071600 | Chr01:17669166.17671888 | 657 | 218 | 24.6 | 9.36 | Nuclear | WGD/segmental | |
| MD07G1131000 | Chr07:18748505..18750267 | 669 | 222 | 24.9 | 9.36 | Nuclear | WGD/segmental | |
| MD06G1138500 | Chr06:28356819.28357724 | 453 | 150 | 17.1 | 9.56 | Nuclear | WGD/segmental | |
| MD14G1154500 | Chr14:24914608.24915904 | 447 | 148 | 17.1 | 9.59 | Extracellular | WGD/segmental | |
| MD16G1122400 | Chr16:8821613.8823564 | 486 | 161 | 18.0 | 9.08 | Nuclear | WGD/segmental | |
| MD13G1122100 | Chr13:8993050.8995368 | 573 | 190 | 21.7 | 9.74 | Nuclear | WGD/segmental | |
| MD09G1008800 | Chr09:616602.619344 | 651 | 216 | 24.8 | 9.14 | Nuclear | WGD/segmental | |
| MD17G1001500 | Chr17:91553.93431 | 672 | 223 | 25.6 | 9.14 | Nuclear | WGD/segmental | |
| IId | MD15G1078200 | Chr15:5334685.5336858 | 1029 | 342 | 37.2 | 9.26 | Nuclear | WGD/segmental |
| MD06G1062800 | Chr06:10770799.10772691 | 957 | 318 | 35.9 | 9.58 | Nuclear | dispersed | |
| MD10G1191400 | Chr10:28819078.28822211 | 1068 | 355 | 40.0 | 9.68 | Nuclear | WGD/segmental | |
| MD05G1204400 | Chr05:33402565.33405458 | 1065 | 354 | 40.0 | 9.70 | Nuclear | WGD/segmental | |
| MD02G1177500 | Chr02:15663260.15665127 | 993 | 330 | 36.0 | 9.65 | Nuclear | WGD/segmental | |
| MD08G1094900 | Chr08:7968389.7970451 | 1071 | 356 | 38.7 | 9.41 | Nuclear | WGD/segmental | |
| MD15G1287300 | Chr15:26365198.26366819 | 996 | 331 | 36.2 | 9.54 | Nuclear | WGD/segmental | |
| MD12G1243400 | Chr12:31414877.31416020 | 942 | 313 | 35.3 | 9.96 | Nuclear | WGD/segmental | |
| MD04G1226400 | Chr04:30603205.30604718 | 978 | 325 | 36.6 | 9.77 | Nuclear | WGD/segmental | |
| MD15G1106600 | Chr15:7467131.7469028 | 1017 | 338 | 36.7 | 9.46 | Nuclear | WGD/segmental | |
| MD08G1127200 | Chr08:11928202.11930014 | 1026 | 341 | 37.0 | 9.32 | Nuclear | WGD/segmental | |
| MD16G1244300 | Chr16:26579453.26581081 | 855 | 284 | 30.9 | 9.87 | Nuclear | WGD/segmental | |
| MD13G1239100 | Chr13:24320203.24321656 | 846 | 281 | 30.7 | 9.99 | Nuclear | WGD/segmental | |
| IIe | MD04G1131000 | Chr04:21800375.21802338 | 894 | 297 | 33.4 | 5.72 | Nuclear | WGD/segmental |
| MD12G1144100 | Chr12:22324583.22326472 | 894 | 297 | 33.1 | 5.94 | Nuclear | WGD/segmental | |
| MD03G1292900 | Chr03:36975267.36977012 | 783 | 260 | 30.3 | 5.17 | Nuclear | WGD/segmental | |
| MD00G1140800 | Chr00:30743431.30745181 | 807 | 268 | 30.8 | 4.81 | Nuclear | WGD/segmental | |
| MD05G1295700 | Chr05:43023949.43025466 | 822 | 273 | 30.2 | 5.80 | Nuclear | WGD/segmental | |
| MD10G1275800 | Chr10:36698263.36699996 | 801 | 266 | 29.5 | 5.10 | Nuclear | WGD/segmental | |
| MD09G1235100 | Chr09:29539556.29542546 | 753 | 250 | 27.4 | 5.47 | Nuclear | WGD/segmental | |
| MD05G1265200 | Chr05:40011060.40015621 | 1479 | 492 | 53.8 | 5.85 | Nuclear | WGD/segmental | |
| MD10G1243000 | Chr10:33776504.33781157 | 1482 | 493 | 53.2 | 6.05 | Nuclear | WGD/segmental | |
| MD06G1091200 | Chr06:21994255.21995556 | 888 | 295 | 33.6 | 5.49 | Nuclear | WGD/segmental | |
| MD14G1112200 | Chr14:18037604.18039324 | 948 | 315 | 35.6 | 5.05 | Nuclear | WGD/segmental | |
| MD07G1131400 | Chr07:18815423.18816949 | 1047 | 348 | 37.7 | 6.75 | Nuclear | WGD/segmental | |
| MD01G1071300 | Chr01:17602924.17604669 | 1050 | 349 | 37.7 | 8.16 | Nuclear | WGD/segmental | |
| MD07G1280300 | Chr07:34424687.34425782 | 837 | 278 | 31.1 | 7.10 | Nuclear | WGD/segmental | |
| MD01G1210200 | Chr01:30413680.30415409 | 1413 | 470 | 51.8 | 5.08 | Nuclear | WGD/segmental | |
| III | MD01G1168500 | Chr01:27285229.27286785 | 687 | 228 | 25.7 | 7.72 | Cytoplasmic/ Nuclear | WGD/segmental |
| MD01G1215300 | Chr01:30851473.30853003 | 1044 | 347 | 38.7 | 5.93 | Nuclear | WGD/segmental | |
| MD07G1234600 | Chr07:30813251.30815403 | 1029 | 342 | 37.4 | 5.48 | Nuclear | WGD/segmental | |
| MD06G1104100 | Chr06:24218568.24220572 | 1056 | 351 | 39.4 | 5.72 | Nuclear | WGD/segmental | |
| MD07G1285200 | Chr07:34745168.34746713 | 1029 | 342 | 38.0 | 5.50 | Nuclear | WGD/segmental | |
| MD07G1285400 | Chr07:34761880.34763425 | 1029 | 342 | 38.0 | 5.50 | Nuclear | proximal | |
| MD07G1261100 | Chr07:32679484.32683843 | 2988 | 995 | 113.2 | 8.64 | Nuclear | dispersed | |
| MD14G1123000 | Chr14:19736610.19738943 | 1068 | 355 | 39.9 | 5.66 | Nuclear | WGD/segmental | |
| MD01G1078000 | Chr01:18445739.18447924 | 1062 | 353 | 39.2 | 5.30 | Nuclear | WGD/segmental | |
| MD07G1146900 | Chr07:21450190.21453542 | 1071 | 356 | 39.7 | 5.48 | Nuclear | WGD/segmental | |
| MD01G1168600 | Chr01:27287662.27291066 | 912 | 303 | 34.2 | 5.71 | Nuclear | WGD/segmental | |
| MD12G1189200 | Chr12:27086797.27089009 | 1020 | 339 | 37.7 | 6.41 | Nuclear | WGD/segmental | |
| MD04G1175600 | Chr04:26669452.26671247 | 1035 | 344 | 38.2 | 6.01 | Nuclear | WGD/segmental | |
| MD04G1175500 | Chr04:26653551.26655300 | 1002 | 333 | 37.2 | 5.38 | Nuclear | WGD/segmental | |
| MD12G1189900 | Chr12:27224756.27226237 | 714 | 237 | 27.3 | 8.26 | Extracellular | tandem | |
| MD07G1234700 | Chr07:30816225.30818950 | 909 | 302 | 34.0 | 6.38 | Nuclear | WGD/segmental | |
| MD12G1189700 | Chr12:27213135.27215427 | 999 | 332 | 37.2 | 5.77 | Nuclear | tandem | |
| MD12G1189600 | Chr12:27190989.27194447 | 1110 | 369 | 40.6 | 5.69 | Nuclear | tandem |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qin, Y.; Yu, H.; Cheng, S.; Liu, Z.; Yu, C.; Zhang, X.; Su, X.; Huang, J.; Shi, S.; Zou, Y.; et al. Genome-Wide Analysis of the WRKY Gene Family in Malus domestica and the Role of MdWRKY70L in Response to Drought and Salt Stresses. Genes 2022, 13, 1068. https://doi.org/10.3390/genes13061068
Qin Y, Yu H, Cheng S, Liu Z, Yu C, Zhang X, Su X, Huang J, Shi S, Zou Y, et al. Genome-Wide Analysis of the WRKY Gene Family in Malus domestica and the Role of MdWRKY70L in Response to Drought and Salt Stresses. Genes. 2022; 13(6):1068. https://doi.org/10.3390/genes13061068
Chicago/Turabian StyleQin, Ying, Haixia Yu, Siyuan Cheng, Zhu Liu, Cheng Yu, Xiaoli Zhang, Xinjian Su, Jingwen Huang, Shuting Shi, Yangjun Zou, and et al. 2022. "Genome-Wide Analysis of the WRKY Gene Family in Malus domestica and the Role of MdWRKY70L in Response to Drought and Salt Stresses" Genes 13, no. 6: 1068. https://doi.org/10.3390/genes13061068