
Genome-Wide Identification and Evolution Analysis of the Gibberellin Oxidase Gene Family in Six Gramineae Crops

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Treatments
2.2. Identification of GAox Genes
2.3. Phylogenetic Tree Construction of GAox Genes
2.4. Homology Analysis of GAox Genes
2.5. Distribution and Structure of GAox Genes
2.6. Cis-Elements and Expression Analyses of GAox Genes
2.7. RNA Extraction and qRT-PCR Analysis
3. Results
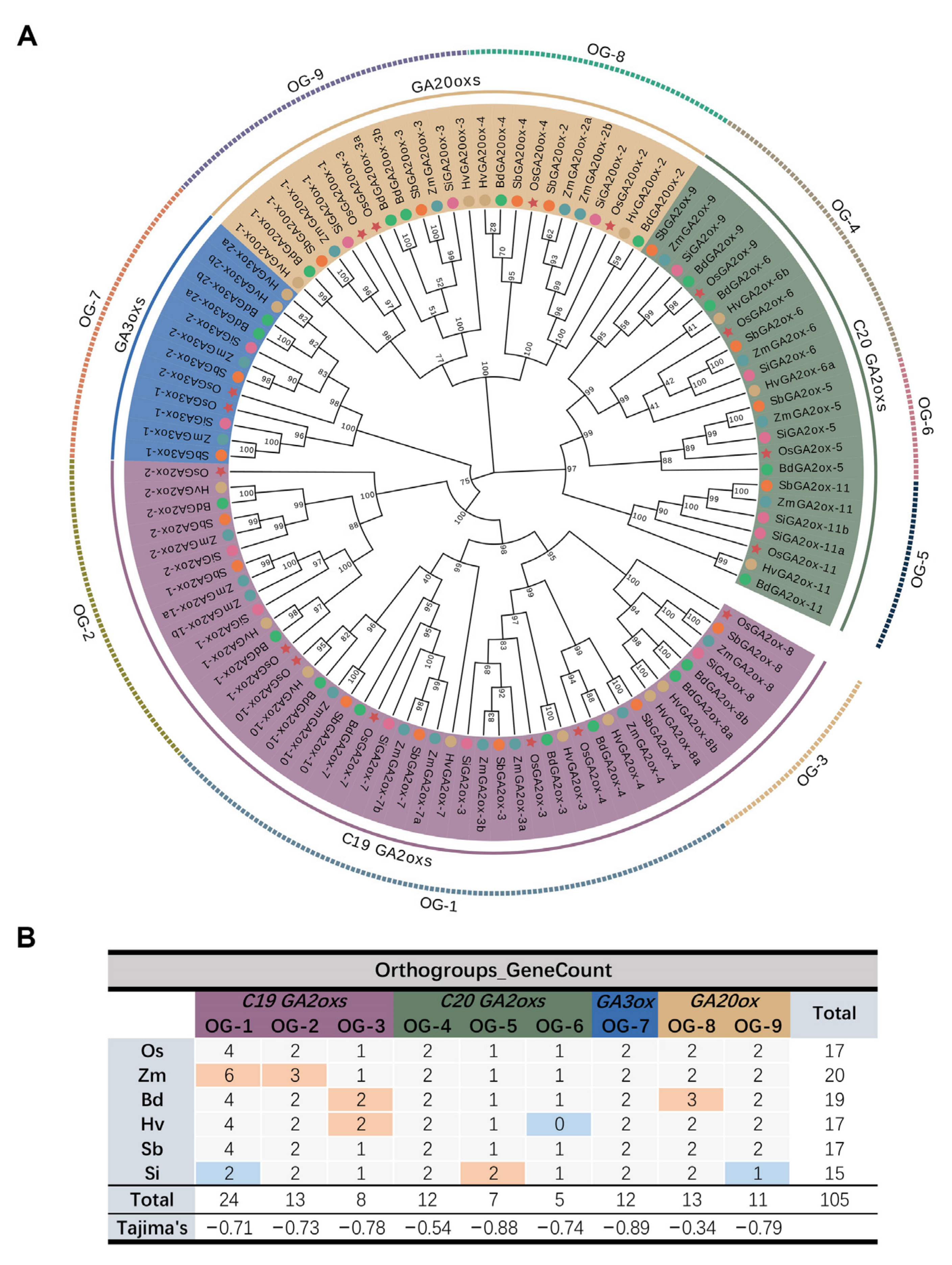
3.1. Identification and OG Analysis of GAoxs in Six Gramineae
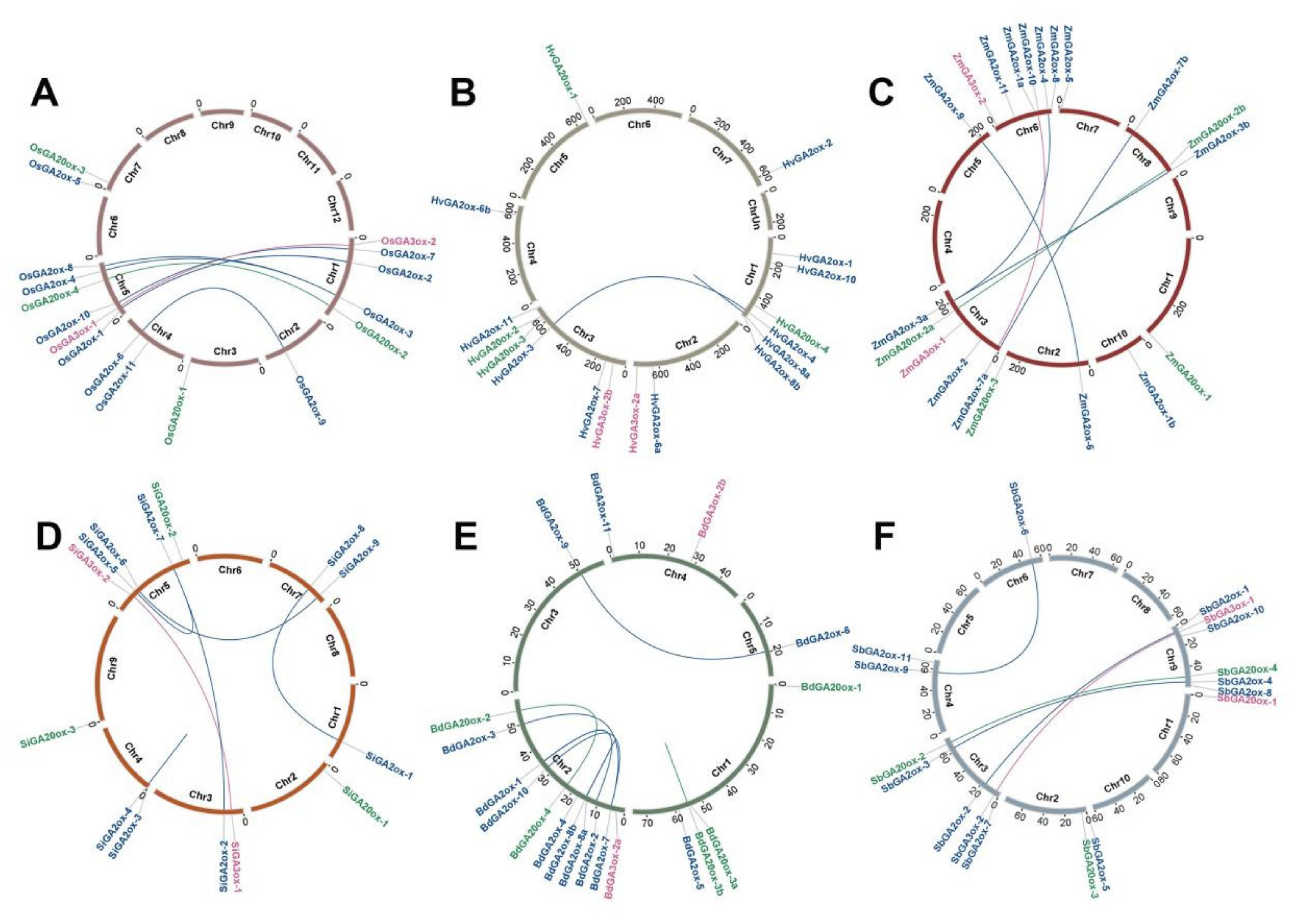
3.2. Chromosome Localization, Gene Duplication Events, and Selective Forces Analysis
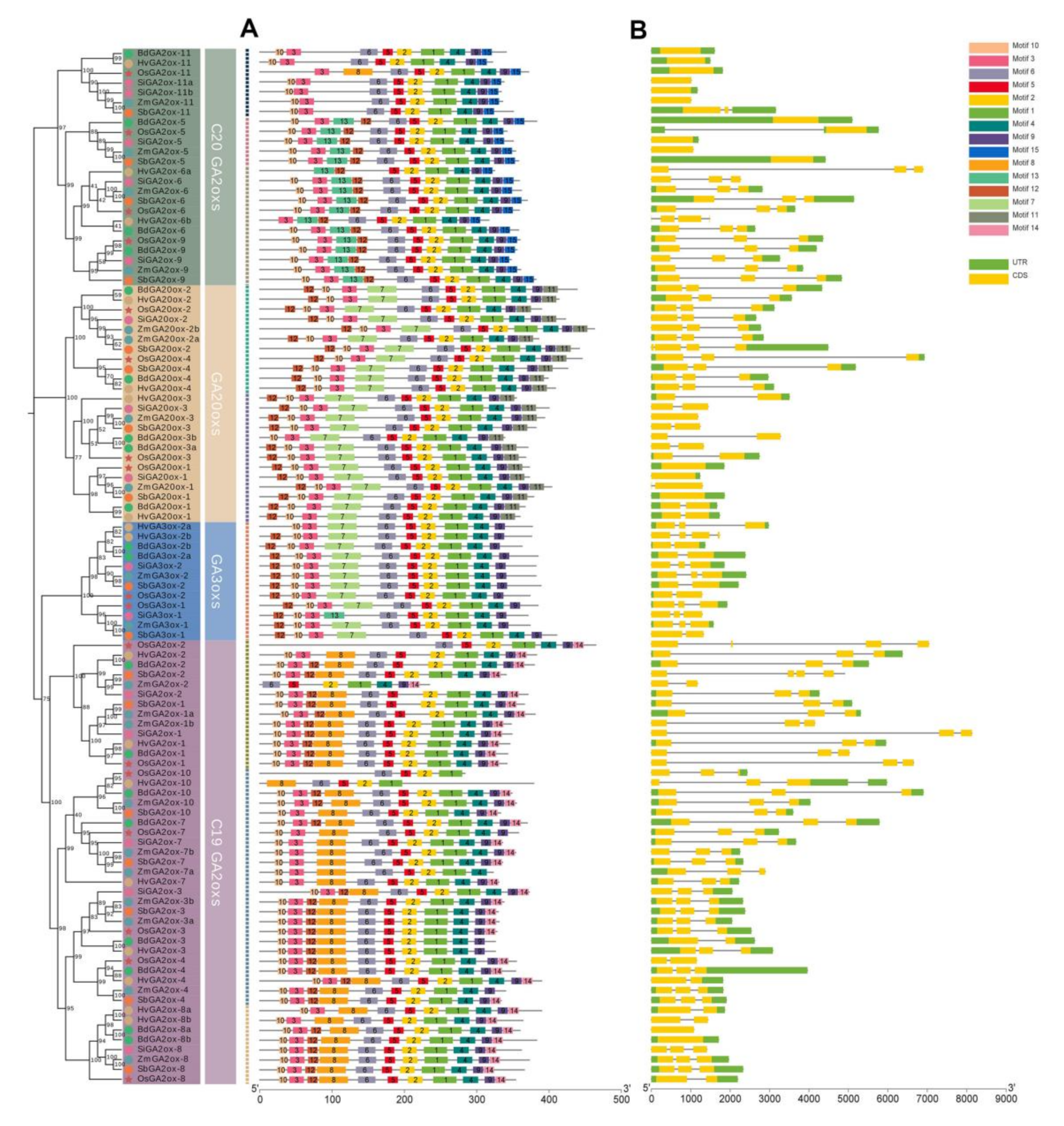
3.3. Gene Structure and Conserved Motif Analysis
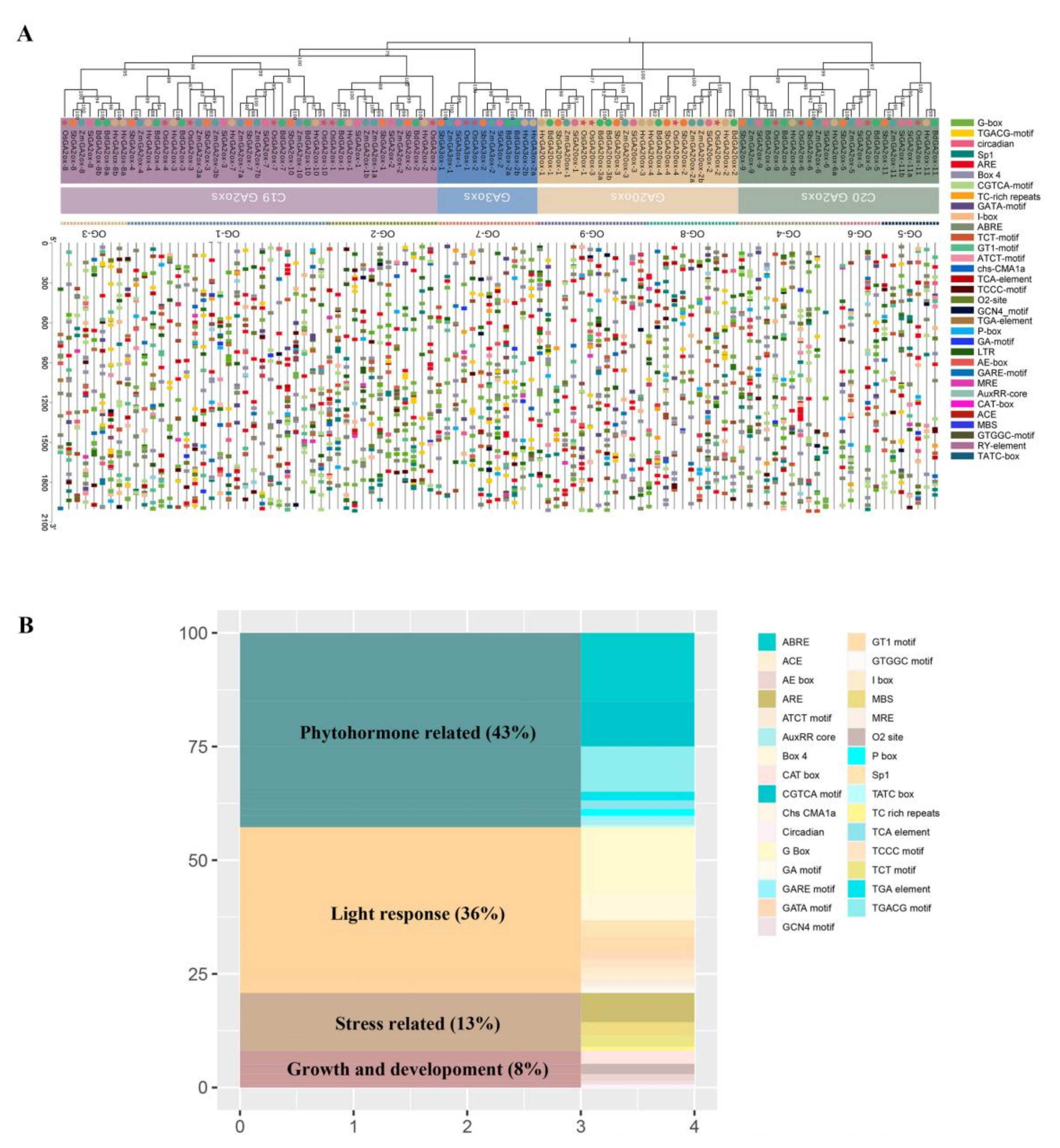
3.4. Cis-Elements Analysis of GAoxs
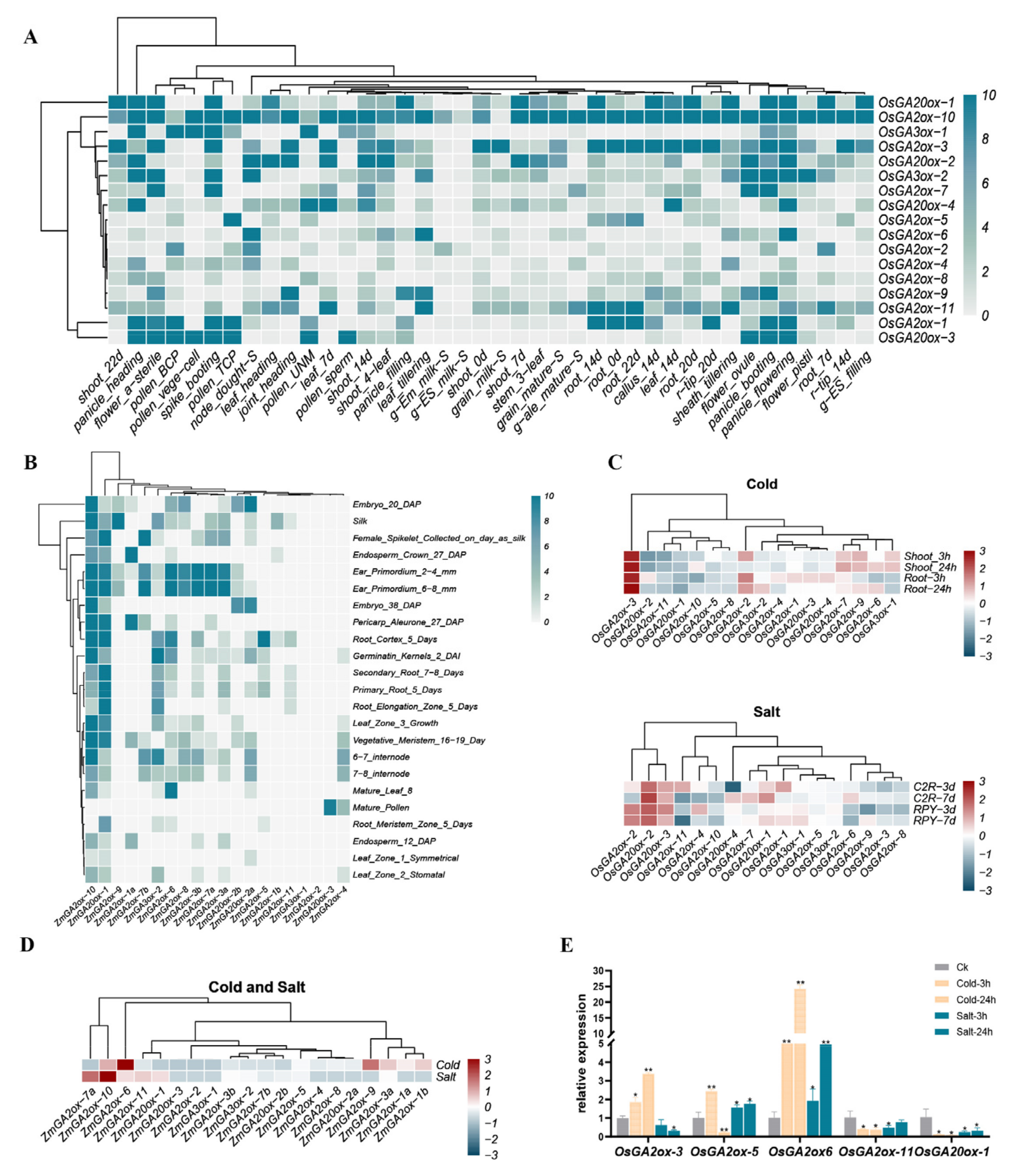
3.5. Tissue-Specific Expression of GAox Genes
3.6. GAox Genes Responded to Cold or Salt Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yamaguchi, S.; Kamiya, Y. Gibberellins and Light-Stimulated Seed Germination. J. Plant Growth Regul. 2001, 20, 369–376. [Google Scholar] [CrossRef]
- Weller, J.L.; Ross, J.J.; Reid, J.B. Gibberellins and Phytochrome Regulation of Stem Elongation in Pea. Planta 1994, 192, 489–496. [Google Scholar] [CrossRef]
- Appleford, N.E.J.; Lenton, J.R. Gibberellins and Leaf Expansion in near-Isogenic Wheat Lines Containing Rht1 and Rht3 Dwarfing Alleles. Planta 1991, 183, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Achard, P.; Genschik, P. Releasing the brakes of plant growth: How GAs shutdown DELLA proteins. J. Exp. Bot. 2008, 60, 1085–1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukazawa, J.; Ohashi, Y.; Takahashi, R.; Nakai, K.; Takahashi, Y. DELLA degradation by gibberellin promotes flowering via GAF1-TPR-dependent repression of floral repressors in Arabidopsis. Plant Cell 2021, 33, 2258–2272. [Google Scholar] [CrossRef] [PubMed]
- Weiss, D.; Van Der Luit, A.; Knegt, E.; Vermeer, E.; Mol, J.; Kooter, J.M. Identification of Endogenous Gibberellins in Petunia Flowers (Induction of Anthocyanin Biosynthetic Gene Expression and the Antagonistic Effect of Abscisic Acid). Plant Physiol. 1995, 107, 695–702. [Google Scholar] [CrossRef] [Green Version]
- Gou, J.; Strauss, S.H.; Tsai, C.J.; Fang, K.; Chen, Y.; Jiang, X.; Busov, V.B. Gibberellins regulate lateral root formation in Populus through interactions with auxin and other hormones. Plant Cell 2010, 22, 623–639. [Google Scholar] [CrossRef] [Green Version]
- Arnaud, N.; Girin, T.; Sorefan, K.; Fuentes, S.; Wood, T.A.; Lawrenson, T.; Sablowski, R.; Ostergaard, L. Gibberellins control fruit patterning in Arabidopsis thaliana. Genes Dev. 2010, 24, 2127–2132. [Google Scholar] [CrossRef] [Green Version]
- Kawaide, H. Biochemical and molecular analyses of gibberellin biosynthesis in fungi. Biosci. Biotechnol. Biochem. 2006, 70, 583–590. [Google Scholar] [CrossRef]
- Yamaguchi, S. Gibberellin metabolism and its regulation. Annu. Rev. Plant Biol. 2008, 59, 225–251. [Google Scholar] [CrossRef]
- Daviere, J.M.; Achard, P. Gibberellin signaling in plants. Development 2013, 140, 1147–1151. [Google Scholar] [CrossRef] [Green Version]
- Olszewski, N.; Sun, T.P.; Gubler, F. Gibberellin signaling: Biosynthesis, catabolism, and response pathways. Plant Cell 2002, 14, S61–S80. [Google Scholar] [CrossRef] [Green Version]
- Rieu, I.; Eriksson, S.; Powers, S.J.; Gong, F.; Griffiths, J.; Woolley, L.; Benlloch, R.; Nilsson, O.; Thomas, S.G.; Hedden, P.; et al. Genetic analysis reveals that C19-GA 2-oxidation is a major gibberellin inactivation pathway in Arabidopsis. Plant Cell 2008, 20, 2420–2436. [Google Scholar] [CrossRef] [Green Version]
- Magome, H.; Nomura, T.; Hanada, A.; Takeda-Kamiya, N.; Ohnishi, T.; Shinma, Y.; Katsumata, T.; Kawaide, H.; Kamiya, Y.; Yamaguchi, S. CYP714B1 and CYP714B2 encode gibberellin 13-oxidases that reduce gibberellin activity in rice. Proc. Natl. Acad. Sci. USA 2013, 110, 1947–1952. [Google Scholar] [CrossRef] [Green Version]
- Hedden, P.; Phillips, A.L. Gibberellin metabolism: New insights revealed by the genes. Trends Plant. Sci. 2000, 5, 523–530. [Google Scholar] [CrossRef]
- Griggs, D.L.; Hedden, P.; Lazarus, C.M. Partial purification of two gibberellin 2β-hydroxylases from cotyledons of Phaseolus vulgaris. Phytochemistry 1991, 30, 2507–2512. [Google Scholar] [CrossRef]
- Murase, K.; Hirano, Y.; Sun, T.P.; Hakoshima, T. Gibberellin-induced DELLA recognition by the gibberellin receptor GID1. Nature 2008, 456, 459–463. [Google Scholar] [CrossRef]
- Wuddineh, W.A.; Mazarei, M.; Zhang, J.; Poovaiah, C.R.; Mann, D.; Ziebell, A.; Sykes, R.W.; Davis, M.F.; Udvardi, M.K.; Stewart, C.N. Identification and overexpression of gibberellin 2-oxidase (GA2ox) in switchgrass (Panicum virgatum L.) for improved plant architecture and reduced biomass recalcitrance. Plant. Biotechnol. J. 2015, 13, 636–647. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.J.; Zeevaart, J.A. Molecular Cloning of GA 2-Oxidase3 from Spinach and Its Ectopic Expression in Nicotiana sylvestris. Plant Physiol. 2005, 138, 243–254. [Google Scholar] [CrossRef] [Green Version]
- Lange, T.; Krämer, C.; Pimenta Lange, M.J. The Class III Gibberellin 2-Oxidases AtGA2ox9 and AtGA2ox10 Contribute to Cold Stress Tolerance and Fertility. Plant Physiol. 2020, 184, 478–486. [Google Scholar] [CrossRef]
- Han, F.; Zhu, B. Evolutionary analysis of three gibberellin oxidase genes in rice, Arabidopsis, and soybean. Gene 2011, 473, 23–35. [Google Scholar] [CrossRef]
- Dong, F.; Fan, S.; Ma, X.; Meng, Y.; Zhang, D. Genome-wide Identification and Expression Analysis of GA2ox, GA3ox and GA20ox in Apple. Acta Hortic. Sin. 2018, 10, 680. [Google Scholar] [CrossRef] [Green Version]
- Hu, L.; Wang, P.; Hao, Z.; Lu, Y.; Xue, G.; Cao, Z.; Qu, H.; Cheng, T.; Shi, J.; Chen, J. Gibberellin Oxidase Gene Family in L. chinense: Genome-Wide Identification and Gene Expression Analysis. Int. J. Mol. Sci. 2021, 22, 7167. [Google Scholar] [CrossRef]
- Sakamoto, T.; Miura, K.; Itoh, H.; Tatsumi, T.; Ueguchi-Tanaka, M.; Ishiyama, K.; Kobayashi, M.; Agrawal, G.K.; Takeda, S.; Abe, K.; et al. An overview of gibberellin metabolism enzyme genes and their related mutants in rice. Plant. Physiol. 2004, 134, 1642–1653. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, A.; Ashikari, M.; Ueguchi-Tanaka, M.; Itoh, H.; Nishimura, A.; Swapan, D.; Ishiyama, K.; Saito, T.; Kobayashi, M.; Khush, G.S.; et al. Green revolution: A mutant gibberellin-synthesis gene in rice. Nature 2002, 416, 701–702. [Google Scholar] [CrossRef]
- Chi, S.; Mei, Z.; Duan, J.; Chen, H.; Feng, H.; Cai, W.; Gloria, M. OsGA2ox5, a Gibberellin Metabolism Enzyme, Is Involved in Plant Growth, the Root Gravity Response and Salt Stress. PLoS ONE 2014, 9, e87110. [Google Scholar] [CrossRef]
- Lv, S.F.; Yu, D.Y.; Sun, Q.Q.; Jiang, J. Activation of gibberellin 20-oxidase 2 undermines auxin-dependent root and root hair growth in NaCl-stressed Arabidopsis seedlings. Plant Growth Regul. 2018, 84, 225–236. [Google Scholar] [CrossRef]
- Xue, Q.; Liu, J.H.; Zhao, W.; Chen, X.; Peng, Y. Gibberellin 20-Oxidase Gene OsGA20ox3 Regulates Plant Stature and Disease Development in Rice. Mol. Plant-Microbe Interact. MPMI 2013, 26, 227–239. [Google Scholar] [CrossRef] [Green Version]
- Asano, K.; Yamasaki, M.; Takuno, S.; Miura, K.; Katagiri, S.; Ito, T.; Doi, K.; Wu, J.; Ebana, K.; Matsumoto, T.; et al. Artificial selection for a green revolution gene during japonica rice domestication. Proc. Natl. Acad. Sci. USA 2011, 108, 11034–11039. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Li, M.W.; Wong, F.L.; Luk, C.Y.; Chung, C.Y.; Yung, W.S.; Wang, Z.; Xie, M.; Song, S.; Chung, G.; et al. Increased copy number of gibberellin 2-oxidase 8 genes reduced trailing growth and shoot length during soybean domestication. Plant J. Cell Mol. Biol. 2021, 107, 1739–1755. [Google Scholar] [CrossRef]
- Jian, S.; Guo, B.; Song, F.; Peng, H.; Yao, Y.; Zhang, Y.; Sun, Q.; Ni, Z. Genome-wide identification of gibberellins metabolic enzyme genes and expression profiling analysis during seed germination in maize. Gene 2011, 482, 34–42. [Google Scholar] [CrossRef]
- Pearce, S.; Huttly, A.K.; Prosser, I.M.; Li, Y.D.; Vaughan, S.P.; Gallova, B.; Patil, A.; Coghill, J.A.; Dubcovsky, J.; Hedden, P.; et al. Heterologous expression and transcript analysis of gibberellin biosynthetic genes of grasses reveals novel functionality in the GA3ox family. BMC Plant Biol. 2015, 15, 130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.L.; Sun, J.Z.; Ali, S.S.; Gao, L.; Ni, X.N.; Li, X.; Wu, Y.F.; Jiang, J.X. Identification and expression analysis of Sorghum bicolor gibberellin oxidase genes with varied gibberellin levels involved in regulation of stem biomass. Ind. Crop. Prod. 2020, 145, 111951. [Google Scholar] [CrossRef]
- Kong, W.; Zhong, H.; Deng, X.; Gautam, M.; Gong, Z.; Zhang, Y.; Zhao, G.; Liu, C.; Li, Y. Evolutionary Analysis of GH3 Genes in Six Oryza Species/Subspecies and Their Expression under Salinity Stress in Oryza sativa ssp. japonica. Plants 2019, 8, 30. [Google Scholar] [CrossRef] [Green Version]
- Kong, W.; Sun, T.; Zhang, C.; Qiang, Y.; Li, Y. Micro-Evolution Analysis Reveals Diverged Patterns of Polyol Transporters in Seven Gramineae Crops. Front. Genet. 2020, 11, 565. [Google Scholar] [CrossRef]
- Kong, W.; Zhang, Y.; Deng, X.; Li, S.; Zhang, C.; Li, Y. Comparative Genomic and Transcriptomic Analysis Suggests the Evolutionary Dynamic of GH3 Genes in Gramineae Crops. Front. Plant. Sci. 2019, 10, 1297. [Google Scholar] [CrossRef] [Green Version]
- Zhong, H.; Kong, W.; Gong, Z.; Fang, X.; Deng, X.; Liu, C.; Li, Y. Evolutionary Analyses Reveal Diverged Patterns of SQUAMOSA Promoter Binding Protein-Like (SPL) Gene Family in Oryza Genus. Front. Plant. Sci. 2019, 10, 565. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Kozlov, A.M.; Darriba, D.; Flouri, T.; Morel, B.; Stamatakis, A. RAxML-NG: A fast, scalable and user-friendly tool for maximum likelihood phylogenetic inference. Bioinformatics 2019, 35, 4453–4455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subramanian, B.; Gao, S.; Lercher, M.J.; Hu, S.; Chen, W.H. Evolview v3: A webserver for visualization, annotation, and management of phylogenetic trees. Nucleic Acids Res. 2019, 47, W270–W275. [Google Scholar] [CrossRef] [PubMed]
- Emms, D.M.; Kelly, S. OrthoFinder: Solving fundamental biases in whole genome comparisons dramatically improves orthogroup inference accuracy. Genome Biol. 2015, 16, 157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef] [PubMed]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Tang, H.; Debarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.H.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, W.; Gong, Z.; Zhong, H.; Zhang, Y.; Zhao, G.; Gautam, M.; Deng, X.; Liu, C.; Zhang, C.; Li, Y. Expansion and Evolutionary Patterns of Glycosyltransferase Family 8 in Gramineae Crop Genomes and Their Expression under Salt and Cold Stresses in Oryza sativa ssp. japonica. Biomolecules 2019, 9, 18. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant. 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Peng, H.; Wang, K.; Chen, Z.; Cao, Y.; Gao, Q.; Li, Y.; Li, X.; Lu, H.; Du, H.; Lu, M.; et al. MBKbase for rice: An integrated omics knowledgebase for molecular breeding in rice. Nucleic Acids Res. 2020, 48, D1085–D1092. [Google Scholar] [CrossRef] [Green Version]
- Gui, S.; Yang, L.; Li, J.; Luo, J.; Xu, X.; Yuan, J.; Chen, L.; Li, W.; Yang, X.; Wu, S.; et al. ZEAMAP, a Comprehensive Database Adapted to the Maize Multi-Omics Era. iScience 2020, 23, 101241. [Google Scholar] [CrossRef]
- Kong, W.; Sun, T.; Zhang, C.; Deng, X.; Li, Y. Comparative Transcriptome Analysis Reveals the Mechanisms Underlying Differences in Salt Tolerance Between indica and japonica Rice at Seedling Stage. Front. Plant. Sci. 2021, 12, 725436. [Google Scholar] [CrossRef]
- Lo, S.F.; Yang, S.Y.; Chen, K.T.; Hsing, Y.I.; Zeevaart, J.A.; Chen, L.J.; Yu, S.M. A novel class of gibberellin 2-oxidases control semidwarfism, tillering, and root development in rice. Plant. Cell 2008, 20, 2603–2618. [Google Scholar] [CrossRef] [Green Version]
- Cannon, S.B.; Mitra, A.; Baumgarten, A.; Young, N.D.; May, G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant. Biol. 2004, 4, 10. [Google Scholar] [CrossRef] [Green Version]
- Ye, H.; Feng, J.; Zhang, L.; Zhang, J.; Mispan, M.S.; Cao, Z.; Beighley, D.H.; Yang, J.; Gu, X.Y. Map-Based Cloning of Seed Dormancy1-2 Identified a Gibberellin Synthesis Gene Regulating the Development of Endosperm-Imposed Dormancy in Rice. Plant. Physiol 2015, 169, 2152–2165. [Google Scholar] [CrossRef] [Green Version]
- Vikram, P.; Kadam, S.; Singh, B.P.; Lee, Y.J.; Pal, J.K.; Singh, S.; Singh, O.N.; Swamy, B.P.M.; Thiyagarajan, K.; Singh, S.; et al. Genetic Diversity Analysis Reveals Importance of Green Revolution Gene (Sd1 Locus) for Drought Tolerance in Rice. Agr. Res. 2016, 5, 1–12. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, Y.; Mi, X.F.; Shan, J.X.; Li, X.M.; Xu, J.L.; Lin, H.X. The QTL GNP1 Encodes GA20ox1, Which Increases Grain Number and Yield by Increasing Cytokinin Activity in Rice Panicle Meristems. PLoS Genet. 2016, 12, e1006386. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Underhill, S.J. Breadfruit (Artocarpus altilis) gibberellin 2-oxidase genes in stem elongation and abiotic stress response. Plant. Physiol. Biochem. PPB 2016, 98, 81–88. [Google Scholar] [CrossRef]
- Hedden, P. The genes of the Green Revolution. Trends Genet. TIG 2003, 19, 5–9. [Google Scholar] [CrossRef]
- Swigonová, Z.; Lai, J.; Ma, J.; Ramakrishna, W.; Llaca, V.; Bennetzen, J.L.; Messing, J. Close split of sorghum and maize genome progenitors. Genome Res. 2004, 14, 1916–1923. [Google Scholar] [CrossRef] [Green Version]
- Vidal, A.M.; Gisbert, C.; Talón, M.; Primo-Millo, E.; López-Díaz, I.; García-Martínez, J.L. The ectopic overexpression of a citrus gibberellin 20-oxidase enhances the non-13-hydroxylation pathway of gibberellin biosynthesis and induces an extremely elongated phenotype in tobacco. Physiol. Plant 2001, 112, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Pan, C.; Tian, K.; Ban, Q.; Wang, L.; Sun, Q.; He, Y.; Yang, Y.; Pan, Y.; Li, Y.; Jiang, J.; et al. Genome-Wide Analysis of the Biosynthesis and Deactivation of Gibberellin-Dioxygenases Gene Family in Camellia sinensis (L.) O. Kuntze. Genes 2017, 8, 235. [Google Scholar] [CrossRef] [PubMed]
- Lange, T.; Hedden, P.; Graebe, J.E. Expression cloning of a gibberellin 20-oxidase, a multifunctional enzyme involved in gibberellin biosynthesis. Proc. Natl. Acad. Sci. USA 1994, 91, 8552–8556. [Google Scholar] [CrossRef] [Green Version]
- Schomburg, F.M.; Bizzell, C.M.; Lee, D.J.; Zeevaart, J.A.; Amasino, R.M. Overexpression of a novel class of gibberellin 2-oxidases decreases gibberellin levels and creates dwarf plants. Plant Cell 2003, 15, 151–163. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Cai, W.J.; Yin, X.; Yang, D.; Dong, T.; Feng, Y.Q.; Wu, Y. Two SLENDER AND CRINKLY LEAF dioxygenases play an essential role in rice shoot development. J. Exp. Bot. 2020, 71, 1387–1401. [Google Scholar] [CrossRef]
- Sun, H.; Pang, B.; Yan, J.; Wang, T.; Wang, L.; Chen, C.; Li, Q.; Ren, Z. Comprehensive Analysis of Cucumber Gibberellin Oxidase Family Genes and Functional Characterization of CsGA20ox1 in Root Development in Arabidopsis. Int. J. Mol. Sci. 2018, 19, 3135. [Google Scholar] [CrossRef] [Green Version]
- Lo, S.F.; Ho, T.D.; Liu, Y.L.; Jiang, M.J.; Hsieh, K.T.; Chen, K.T.; Yu, L.C.; Lee, M.H.; Chen, C.Y.; Huang, T.P.; et al. Ectopic expression of specific GA2 oxidase mutants promotes yield and stress tolerance in rice. Plant. Biotechnol. J. 2017, 15, 850–864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magome, H.; Yamaguchi, S.; Hanada, A.; Kamiya, Y.; Oda, K. The DDF1 transcriptional activator upregulates expression of a gibberellin-deactivating gene, GA2ox7, under high-salinity stress in Arabidopsis. Plant J. Cell Mol. Biol. 2008, 56, 613–626. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Seq-1 | Seq-2 | Ka | Ks | Ka_Ks | Duplication Type | Divergence Time (Mya) |
|---|---|---|---|---|---|---|
| BdGA20ox-3a | BdGA20ox-3b | 0.03 | 0.08 | 0.37 | TD | 4.24 |
| ZmGA20ox-2a | ZmGA20ox-2b | 0.03 | 0.09 | 0.31 | WGD/SD | 5.04 |
| ZmGA2ox-3a | ZmGA2ox-3b | 0.04 | 0.10 | 0.45 | WGD/SD | 5.40 |
| ZmGA2ox-7a | ZmGA2ox-7b | 0.04 | 0.10 | 0.39 | WGD/SD | 5.43 |
| HvGA2ox-4 | HvGA2ox-3 | 0.06 | 0.21 | 0.31 | WGD/SD | 11.38 |
| SiGA2ox-11a | SiGA2ox-11b | 0.20 | 0.25 | 0.83 | TD | 13.53 |
| BdGA2ox-8a | BdGA2ox-8b | 0.14 | 0.35 | 0.40 | TD | 19.12 |
| OsGA20ox-2 | OsGA20ox-4 | 0.22 | 0.36 | 0.61 | WGD/SD | 19.82 |
| SiGA3ox-1 | SiGA3ox-2 | 0.32 | 0.37 | 0.87 | WGD/SD | 20.42 |
| SbGA2ox-3 | SbGA2ox-4 | 0.18 | 0.41 | 0.44 | WGD/SD | 22.47 |
| ZmGA2ox-3a | ZmGA2ox-4 | 0.19 | 0.44 | 0.43 | WGD/SD | 24.40 |
| SiGA2ox-9 | SiGA2ox-6 | 0.19 | 0.45 | 0.42 | WGD/SD | 24.52 |
| BdGA2ox-7 | BdGA2ox-10 | 0.20 | 0.47 | 0.43 | WGD/SD | 25.73 |
| SiGA2ox-8 | SiGA2ox-3 | 0.33 | 0.47 | 0.71 | WGD/SD | 25.83 |
| HvGA2ox-8a | HvGA2ox-8b | 0.20 | 0.49 | 0.42 | TD | 26.84 |
| SbGA3ox-2 | SbGA3ox-1 | 0.34 | 0.51 | 0.68 | WGD/SD | 27.76 |
| BdGA20ox-4 | BdGA20ox-2 | 0.26 | 0.51 | 0.50 | WGD/SD | 27.86 |
| OsGA3ox-2 | OsGA3ox-1 | 0.33 | 0.51 | 0.64 | WGD/SD | 28.00 |
| ZmGA3ox-2 | ZmGA3ox-1 | 0.33 | 0.53 | 0.63 | WGD/SD | 28.87 |
| OsGA2ox-3 | OsGA2ox-4 | 0.26 | 0.53 | 0.49 | WGD/SD | 29.37 |
| ZmGA2ox-6 | ZmGA2ox-9 | 0.20 | 0.54 | 0.38 | WGD/SD | 29.43 |
| OsGA2ox-9 | OsGA2ox-6 | 0.21 | 0.54 | 0.39 | WGD/SD | 29.45 |
| SiGA2ox-7 | SiGA2ox-3 | 0.36 | 0.54 | 0.67 | WGD/SD | 29.55 |
| SbGA20ox-2 | SbGA20ox-4 | 0.27 | 0.55 | 0.50 | WGD/SD | 30.28 |
| BdGA2ox-9 | BdGA2ox-6 | 0.20 | 0.58 | 0.34 | WGD/SD | 32.10 |
| OsGA2ox-3 | OsGA2ox-8 | 0.31 | 0.59 | 0.53 | WGD/SD | 32.26 |
| SbGA2ox-9 | SbGA2ox-6 | 0.21 | 0.59 | 0.36 | WGD/SD | 32.35 |
| BdGA2ox-7 | BdGA2ox-3 | 0.41 | 0.61 | 0.67 | WGD/SD | 33.41 |
| BdGA2ox-2 | BdGA2ox-1 | 0.24 | 0.71 | 0.34 | WGD/SD | 39.18 |
| SbGA2ox-2 | SbGA2ox-1 | 0.23 | 0.78 | 0.29 | WGD/SD | 42.60 |
| OsGA2ox-2 | OsGA2ox-1 | 0.49 | 0.99 | 0.50 | WGD/SD | 54.19 |
| OsGA2ox-7 | OsGA2ox-10 | 0.46 | 1.06 | 0.43 | WGD/SD | 58.07 |
| SiGA2ox-2 | SiGA2ox-1 | 0.23 | 1.21 | 0.19 | WGD/SD | 66.43 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, C.; Nie, X.; Kong, W.; Deng, X.; Sun, T.; Liu, X.; Li, Y. Genome-Wide Identification and Evolution Analysis of the Gibberellin Oxidase Gene Family in Six Gramineae Crops. Genes 2022, 13, 863. https://doi.org/10.3390/genes13050863
Zhang C, Nie X, Kong W, Deng X, Sun T, Liu X, Li Y. Genome-Wide Identification and Evolution Analysis of the Gibberellin Oxidase Gene Family in Six Gramineae Crops. Genes. 2022; 13(5):863. https://doi.org/10.3390/genes13050863
Chicago/Turabian StyleZhang, Chenhao, Xin Nie, Weilong Kong, Xiaoxiao Deng, Tong Sun, Xuhui Liu, and Yangsheng Li. 2022. "Genome-Wide Identification and Evolution Analysis of the Gibberellin Oxidase Gene Family in Six Gramineae Crops" Genes 13, no. 5: 863. https://doi.org/10.3390/genes13050863