
Complex Changes in the Efficiency of the Expression of Many Genes in Monogenic Diseases, Mucopolysaccharidoses, May Arise from Significant Disturbances in the Levels of Factors Involved in the Gene Expression Regulation Processes



Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Culture Conditions
2.2. Transcriptomics
2.3. Western Blotting
2.4. Statistical Analyses
3. Results
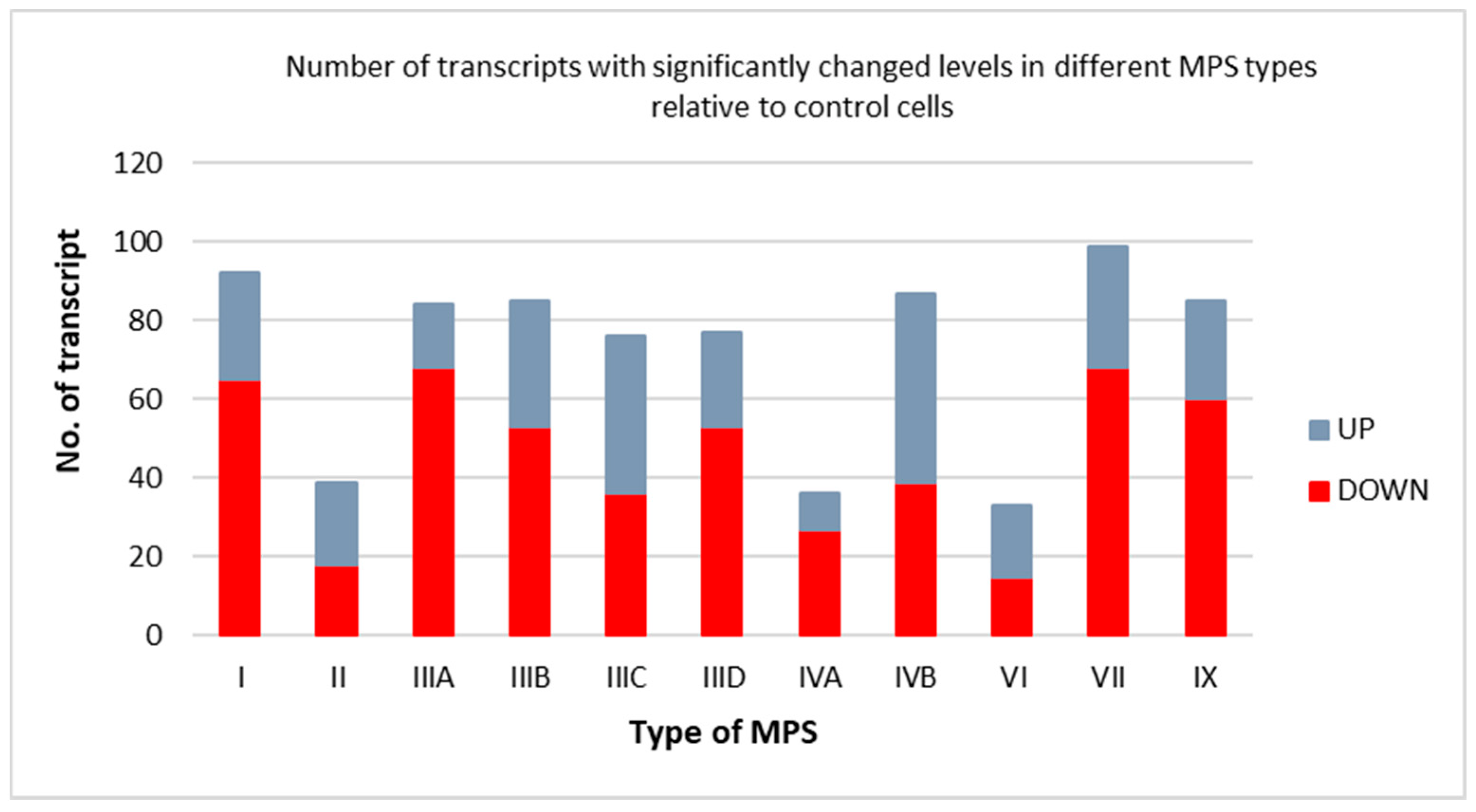
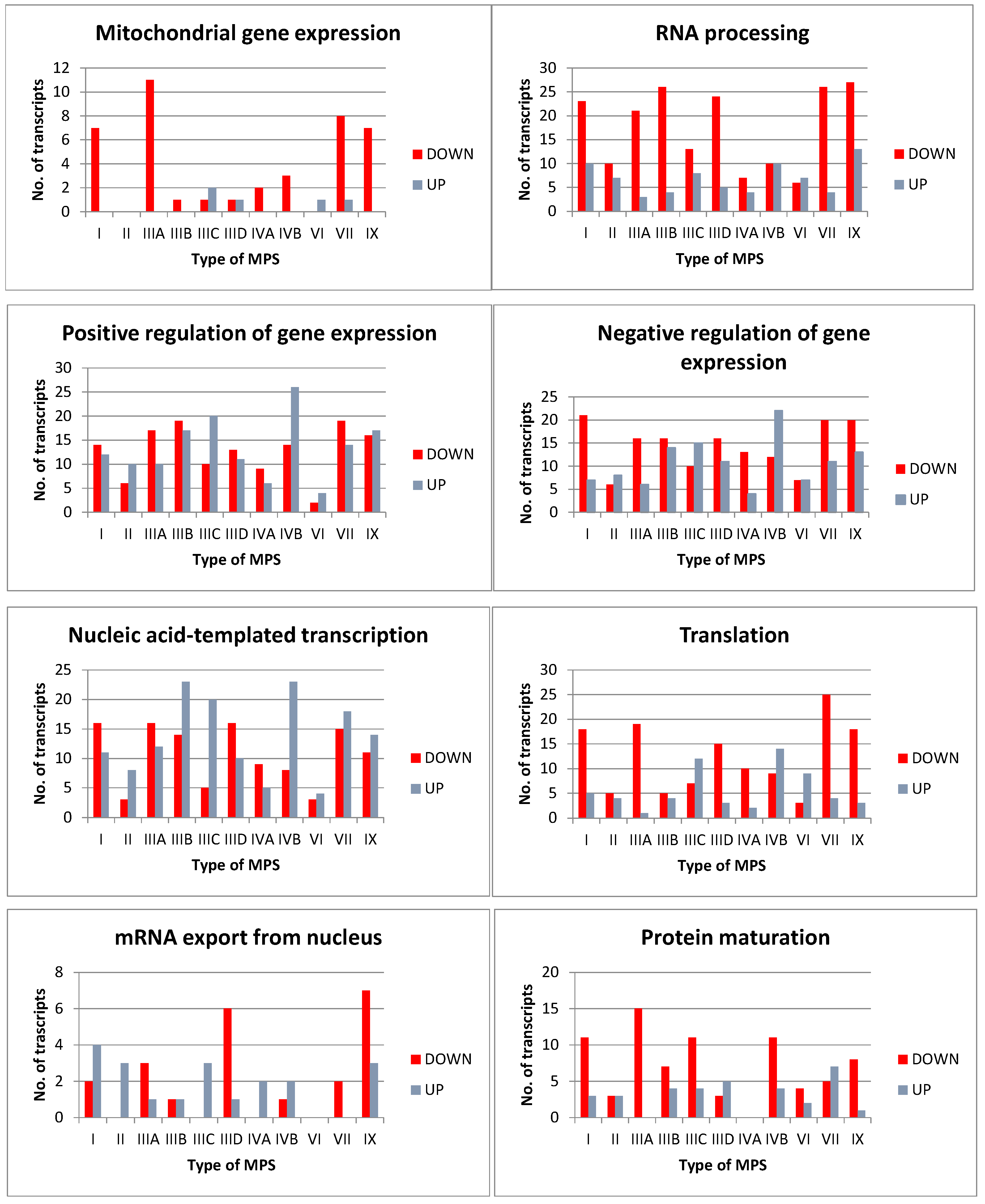
3.1. Transcriptomic Analyses
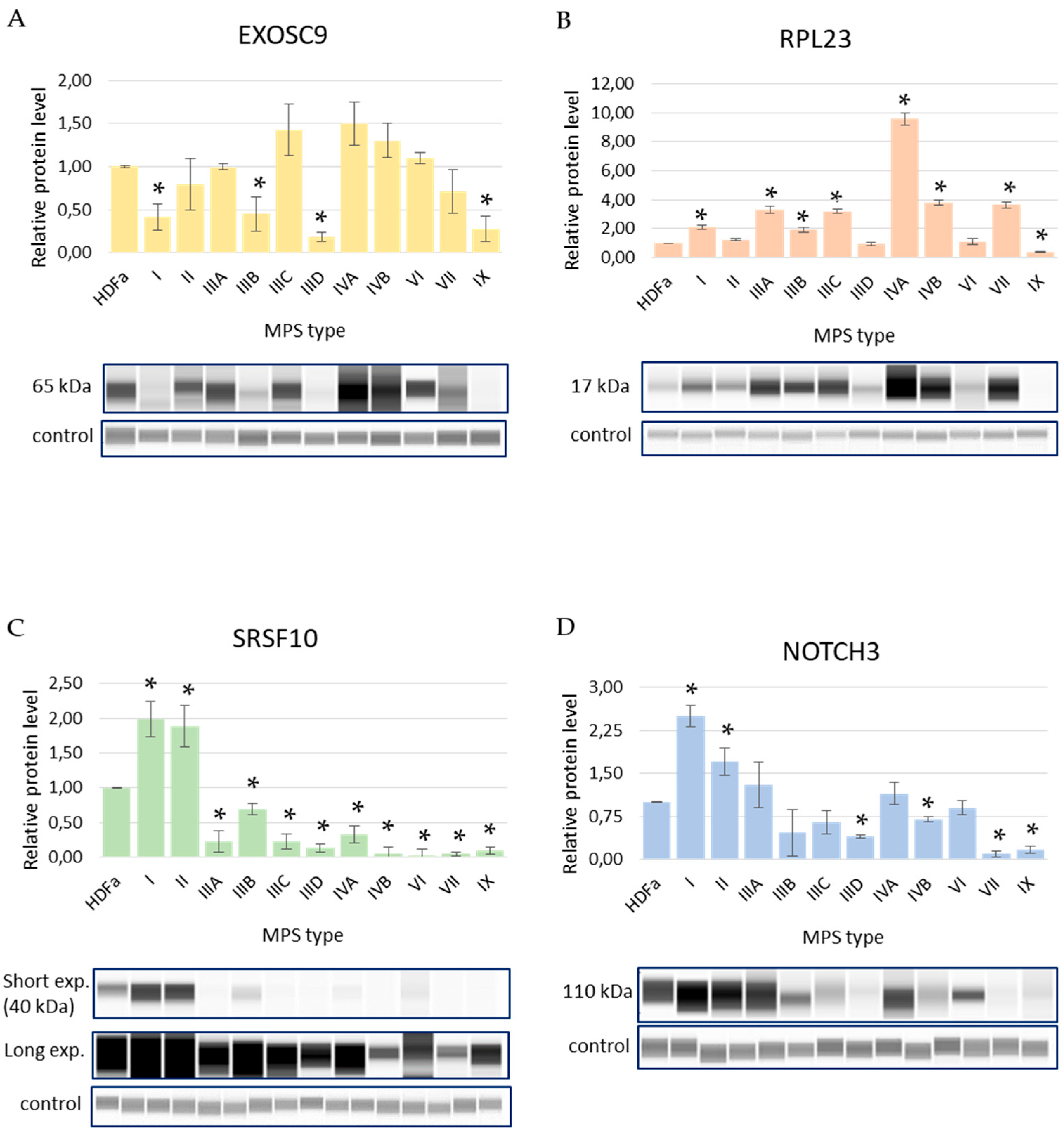
3.2. Levels of Selected Proteins in MPS Cells
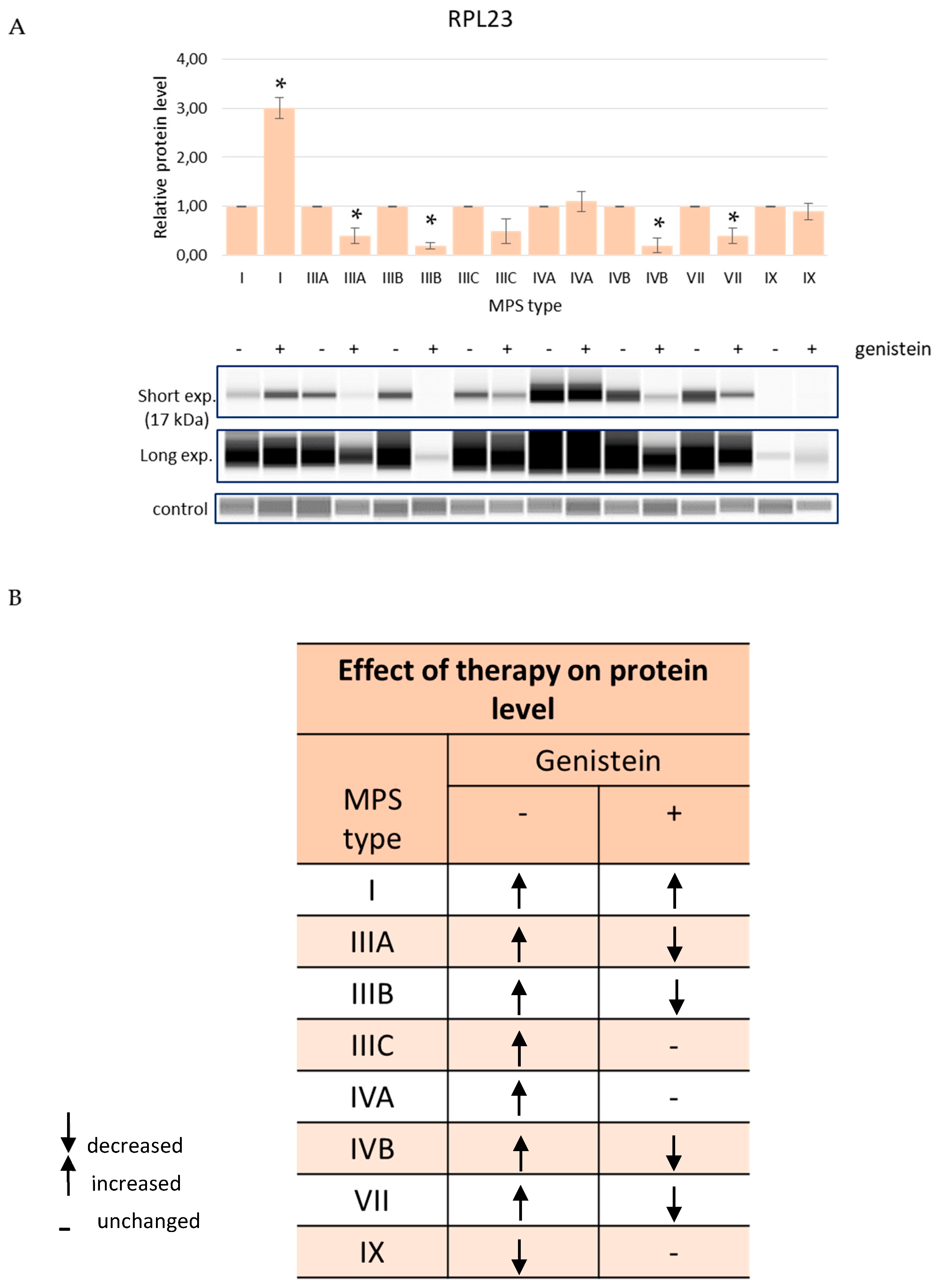
3.3. Effects of the Reduction in the GAG Levels in MPS Cells on the Abundance of the Selected Proteins
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Amberger, J.S.; Bocchini, C.A.; Schiettecatte, F.; Scott, A.F.; Hamosh, A. OMIM.org: Online Mendelian Inheritance in Man (OMIM®), an online catalog of human genes and genetic disorders. Nucleic Acids Res. 2015, 43, D789–D798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boycott, K.M.; Rath, A.; Chong, J.X.; Hartley, T.; Alkuraya, F.S.; Baynam, G.; Brookes, A.J.; Brudno, M.; Carracedo, A.; den Dunnen, J.T.; et al. International Cooperation to Enable the Diagnosis of All Rare Genetic Diseases. Am. J. Hum. Genet. 2017, 100, 695–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niemi, M.E.K. Common genetic variants contribute to risk of rare severe neurodevelopmental disorders. Nature 2018, 562, 268–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, R.; Shi, L.; Hakenberg, J.; Naughton, B.; Sklar, P.; Zhang, J.; Zhou, H.; Tian, L.; Prakash, O.; Lemire, M.; et al. Analysis of 589,306 genomes identifies individuals resilient to severe Mendelian childhood diseases. Nat. Biotechnol. 2016, 34, 531–538. [Google Scholar] [CrossRef]
- Gaffke, L.; Pierzynowska, K.; Podlacha, M.; Hoinkis, D.; Rintz, E.; Brokowska, J.; Cyske, Z.; Wegrzyn, G. Underestimated aspect of mucopolysaccharidosis pathogenesis: Global changes in cellular processes revealed by transcriptomic studies. Int. J. Mol. Sci. 2020, 21, 1204. [Google Scholar] [CrossRef] [Green Version]
- Çelik, B.; Tomatsu, S.C.; Tomatsu, S.; Khan, S.A. Epidemiology of mucopolysaccharidoses update. Diagnostics 2021, 11, 273. [Google Scholar] [CrossRef]
- Fecarotta, S.; Tarallo, A.; Damiano, C.; Minopoli, N.; Parenti, G. Pathogenesis of mucopolysaccharidoses, an update. Int. J. Mol. Sci. 2020, 21, 2515. [Google Scholar] [CrossRef] [Green Version]
- Tomatsu, S.; Lavery, C.; Giugliani, R.; Harmatz, P.; Scarpa, M.; Wegrzyn, G.; Orii, T. (Eds.) Mucopolysaccharidoses Update; Nova Science Publishers: Hauppauge, NY, USA, 2018. [Google Scholar]
- McBride, K.L.; Flanigan, K.M. Update in the mucopolysaccharidoses. Semin. Pediatr. Neurol. 2021, 37, 100874. [Google Scholar] [CrossRef]
- Taylor, M.; Khan, S.; Stapleton, M.; Wang, J.; Chen, J.; Wynn, R.; Yabe, H.; Chinen, Y.; Boelens, J.J.; Mason, R.W.; et al. Hematopoietic Stem Cell Transplantation for mucopolysaccharidoses: Past, present, and future. Biol. Blood Marrow Transplant. 2019, 25, e226–e246. [Google Scholar] [CrossRef]
- Chen, H.H.; Sawamoto, K.; Mason, R.W.; Kobayashi, H.; Yamaguchi, S.; Suzuki, Y.; Orii, K.; Orii, T.; Tomatsu, S. Enzyme replacement therapy for mucopolysaccharidoses; past, present, and future. J. Hum. Genet. 2019, 64, 1153–1171. [Google Scholar] [CrossRef]
- Parini, R.; Deodato, F. Intravenous enzyme replacement therapy in mucopolysaccharidoses: Clinical effectiveness and limitations. Int. J. Mol. Sci. 2020, 21, 2975. [Google Scholar] [CrossRef]
- Ghosh, A.; Rust, S.; Langford-Smith, K.; Weisberg, D.; Canal, M.; Breen, C.; Hepburn, M.; Tylee, K.; Vaz, F.M.; Vail, A.; et al. High dose genistein in Sanfilippo syndrome: A randomised controlled trial. J. Inherit. Metab. Dis. 2021, 44, 1248–1262. [Google Scholar] [CrossRef]
- Nan, H.; Park, C.; Maeng, S. Mucopolysaccharidoses I and II: Brief review of therapeutic options and supportive/palliative therapies. Biomed. Res. Int. 2020, 2020, 2408402. [Google Scholar] [CrossRef]
- Losada Díaz, J.C.; Cepeda Del Castillo, J.; Rodriguez-López, E.A.; Alméciga-Díaz, C.J. Advances in the development of pharmacological chaperones for the mucopolysaccharidoses. Int. J. Mol. Sci. 2019, 21, 232. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Lin, J.; Leung, W.T.; Wang, L. A basic understanding of mucopolysaccharidosis: Incidence, clinical features, diagnosis, and management. Intractable Rare Dis. Res. 2020, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Gaffke, L.; Pierzynowska, K.; Podlacha, M.; Brokowska, J.; Węgrzyn, G. Changes in cellular processes occurring in mucopolysaccharidoses as underestimated pathomechanisms of these diseases. Cell Biol. Int. 2021, 45, 498–506. [Google Scholar] [CrossRef]
- Rintz, E.; Gaffke, L.; Podlacha, M.; Brokowska, J.; Cyske, Z.; Węgrzyn, G.; Pierzynowska, K. Transcriptomic changes related to cellular processes with particular emphasis on cell activation in lysosomal storage diseases from the group of mucopolysaccharidoses. Int. J. Mol. Sci. 2020, 21, 3194. [Google Scholar] [CrossRef]
- Gaffke, L.; Pierzynowska, K.; Krzelowska, K.; Piotrowska, E.; Węgrzyn, G. Changes in expressions of genes involved in the regulation of cellular processes in mucopolysaccharidoses as assessed by fibroblast culture-based transcriptomic analyses. Metab. Brain Dis. 2020, 35, 1353–1360. [Google Scholar] [CrossRef]
- Pierzynowska, K.; Gaffke, L.; Jankowska, E.; Rintz, E.; Witkowska, J.; Wieczerzak, E.; Podlacha, M.; Węgrzyn, G. Proteasome composition and activity changes in cultured fibroblasts derived from mucopolysaccharidoses patients and their modulation by genistein. Front. Cell Dev. Biol. 2020, 8, 540726. [Google Scholar] [CrossRef]
- Brokowska, J.; Pierzynowska, K.; Gaffke, L.; Rintz, E.; Węgrzyn, G. Expression of genes involved in apoptosis is dysregulated in mucopolysaccharidoses as revealed by pilot transcriptomic analyses. Cell Biol. Int. 2021, 45, 549–557. [Google Scholar] [CrossRef]
- Gaffke, L.; Pierzynowska, K.; Rintz, E.; Cyske, Z.; Giecewicz, I.; Węgrzyn, G. Gene expression-related changes in morphologies of organelles and cellular component organization in mucopolysaccharidoses. Int. J. Mol. Sci. 2021, 22, 2766. [Google Scholar] [CrossRef] [PubMed]
- Gaffke, L.; Szczudło, Z.; Podlacha, M.; Cyske, Z.; Rintz, E.; Mantej, J.; Krzelowska, K.; Węgrzyn, G.; Pierzynowska, K. Impaired ion homeostasis as a possible associate factor in mucopolysaccharidosis pathogenesis: Transcriptomic, cellular and animal studies. Metab. Brain Dis. 2022, 37, 299–310. [Google Scholar] [CrossRef] [PubMed]
- Pierzynowska, K.; Gaffke, L.; Węgrzyn, G. Transcriptomic analyses suggest that mucopolysaccharidosis patients may be less susceptible to COVID-19. FEBS Lett. 2020, 594, 3363–3370. [Google Scholar] [CrossRef] [PubMed]
- Pierzynowska, K.; Gaffke, L.; Podlacha, M.; Węgrzyn, G. Genetic base of behavioral disorders in mucopolysaccharidoses: Transcriptomic studies. Int. J. Mol. Sci. 2020, 21, 1156. [Google Scholar] [CrossRef] [Green Version]
- De Pasquale, V.; Costanzo, M.; Siciliano, R.A.; Mazzeo, M.F.; Pistorio, V.; Bianchi, L.; Marchese, E.; Ruoppolo, M.; Pavone, L.M.; Caterino, M. Proteomic analysis of mucopolysaccharidosis IIIB mouse brain. Biomolecules 2020, 10, 355. [Google Scholar] [CrossRef] [Green Version]
- De Pasquale, V.; Caterino, M.; Costanzo, M.; Fedele, R.; Ruoppolo, M.; Pavone, L.M. Targeted metabolomic analysis of a mucopolysaccharidosis IIIB mouse model reveals an imbalance of branched-chain amino acid and fatty acid metabolism. Int. J. Mol. Sci. 2020, 21, 4211. [Google Scholar] [CrossRef]
- Jakóbkiewicz-Banecka, J.; Piotrowska, E.; Narajczyk, M.; Barańska, S.; Wegrzyn, G. Genistein-mediated inhibition of glycosaminoglycan synthesis, which corrects storage in cells of patients suffering from mucopolysaccharidoses, acts by influencing an epidermal growth factor-dependent pathway. J. Biomed. Sci. 2009, 16, 26. [Google Scholar] [CrossRef] [Green Version]
- Fasken, M.B.; Morton, D.J.; Kuiper, E.G.; Jones, S.K.; Leung, S.W.; Corbett, A.H. The RNA exosome and human disease. Methods Mol. Biol. 2020, 2062, 3–33. [Google Scholar]
- Yang, M.; Zhou, Y.; Deng, H.; Zhou, H.; Cheng, S.; Zhang, D.; Chen, Y.; Chen, J. Ribosomal protein L23 drives the metastasis of hepatocellular carcinoma via upregulating MMP9. Front. Oncol. 2021, 11, 779748. [Google Scholar] [CrossRef]
- Shkreta, L.; Delannoy, A.; Salvetti, A.; Chabot, B. SRSF10: An atypical splicing regulator with critical roles in stress response, organ development, and viral replication. RNA 2021, 27, 1302–1317. [Google Scholar] [CrossRef]
- Schoemaker, D.; Arboleda-Velasquez, J.F. Notch3 signaling and aggregation as targets for the treatment of CADASIL and other NOTCH3-associated small-vessel diseases. Am. J. Pathol. 2021, 191, 1856–1870. [Google Scholar] [CrossRef]
- Moskot, M.; Gabig-Cimińska, M.; Jakóbkiewicz-Banecka, J.; Węsierska, M.; Bocheńska, K.; Węgrzyn, G. Cell cycle is disturbed in mucopolysaccharidosis type II fibroblasts, and can be improved by genistein. Gene 2016, 585, 100–103. [Google Scholar] [CrossRef]
- Arfi, A.; Richard, M.; Gandolphe, C.; Scherman, D. Storage correction in cells of patients suffering from mucopolysaccharidoses types IIIA and VII after treatment with genistein and other isoflavones. J. Inherit. Metab. Dis. 2010, 33, 61–67. [Google Scholar] [CrossRef]
- Friso, A.; Tomanin, R.; Salvalaio, M.; Scarpa, M. Genistein reduces glycosaminoglycan levels in a mouse model of mucopolysaccharidosis type II. Br. J. Pharmacol. 2010, 159, 1082–1091. [Google Scholar] [CrossRef] [Green Version]
- Otomo, T.; Hossain, M.A.; Ozono, K.; Sakai, N. Genistein reduces heparan sulfate accumulation in human mucolipidosis II skin fibroblasts. Mol. Genet. Metab. 2012, 105, 266–269. [Google Scholar] [CrossRef]
- Fumić, B.; Jablan, J.; Cinčić, D.; Zovko Končić, M.; Jug, M. Cyclodextrin encapsulation of daidzein and genistein by grinding: Implication on the glycosaminoglycan accumulation in mucopolysaccharidosis type II and III fibroblasts. J. Microencapsul. 2018, 35, 1–12. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Transcript 1 | Transcripts with Especially Changed Levels (log2FC > 2) in Different MPS Types Relative to Control Cells 2 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| I | II | IIIA | IIIB | IIIC | IIID | IVA | IVB | VI | VII | IX | |
| AEBP1 | ↓ | ||||||||||
| AKT1 | ↓ | ||||||||||
| CDKN1A | ↓ | ↓ | ↓ | ||||||||
| DEK | ↓ | ||||||||||
| DHCR24 | ↓ | ||||||||||
| F3 | ↓ | ||||||||||
| EXOSC9 | ↓ | ↓ | |||||||||
| HNRNPF | ↓ | ||||||||||
| MME (tr. 1) | ↓ | ||||||||||
| MME (tr. 2) | ↓ | ||||||||||
| MME (tr. 3) | ↓ | ||||||||||
| MME (tr. 4) | ↓ | ↓ | |||||||||
| RPL23 | ↓ | ↓ | ↓ | ↓ | ↓ | ↓ | ↓ | ||||
| NUP88 | ↓ | ||||||||||
| RPL10 (tr. 1) | ↓ | ↓ | ↓ | ||||||||
| RPL10 (tr. 2) | ↓ | ||||||||||
| HOXC9 | ↓ | ||||||||||
| SNAPC1 | ↓ | ||||||||||
| RPP25 | ↓ | ↓ | |||||||||
| EIF1AX | ↓ | ||||||||||
| COL4A2 (tr. 1) | ↓ | ↑ | |||||||||
| COL4A2 (tr. 2) | ↑ | ||||||||||
| FLNA | ↑ | ||||||||||
| COMP | ↑ | ↑ | |||||||||
| CPE | ↑ | ↑ | |||||||||
| CSDC2 | ↑ | ||||||||||
| GATA2 | ↑ | ↑ | ↑ | ||||||||
| NOTCH3 | ↑ | ↑ | ↑ | ↑ | ↑ | ↑ | |||||
| POLDIP3 | ↑ | ||||||||||
| RPLP2 | ↑ | ↑ | |||||||||
| SCG5 | ↑ | ||||||||||
| SPOCD1 (tr. 1) | ↑ | ||||||||||
| SPOCD1 (tr. 2) | ↑ | ||||||||||
| CEBPD | ↑ | ||||||||||
| Transcript | Log2FC of Levels of Selected Transcripts in Different MPS Types vs. HDFa Cells | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| I | II | IIIA | IIIB | IIIC | IIID | IVA | IVB | VI | VII | IX | |
| EXOSC9 | −2.32 | −1.18 | −1.02 | −1.69 | −2.33 | −1.99 | −1.72 | −1.10 | −1.58 | −0.48 | −1.60 |
| RPL23 | −0.08 | −2.71 | −0.18 | −4.15 | −4.24 | −4.16 | −4.09 | −3.72 | −3.07 | −3.85 | 0.10 |
| SRSF10 | −1.08 | −0.74 | −0.69 | −0.74 | −0.69 | −0.66 | −0.81 | −0.72 | −0.56 | −0.54 | −0.93 |
| NOTCH3 | 2.42 | 0.97 | 1.93 | 3.66 | 2.56 | 2.93 | 2.70 | 1.97 | 2.01 | 3.32 | 2.70 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cyske, Z.; Gaffke, L.; Pierzynowska, K.; Węgrzyn, G. Complex Changes in the Efficiency of the Expression of Many Genes in Monogenic Diseases, Mucopolysaccharidoses, May Arise from Significant Disturbances in the Levels of Factors Involved in the Gene Expression Regulation Processes. Genes 2022, 13, 593. https://doi.org/10.3390/genes13040593
Cyske Z, Gaffke L, Pierzynowska K, Węgrzyn G. Complex Changes in the Efficiency of the Expression of Many Genes in Monogenic Diseases, Mucopolysaccharidoses, May Arise from Significant Disturbances in the Levels of Factors Involved in the Gene Expression Regulation Processes. Genes. 2022; 13(4):593. https://doi.org/10.3390/genes13040593
Chicago/Turabian StyleCyske, Zuzanna, Lidia Gaffke, Karolina Pierzynowska, and Grzegorz Węgrzyn. 2022. "Complex Changes in the Efficiency of the Expression of Many Genes in Monogenic Diseases, Mucopolysaccharidoses, May Arise from Significant Disturbances in the Levels of Factors Involved in the Gene Expression Regulation Processes" Genes 13, no. 4: 593. https://doi.org/10.3390/genes13040593