
Third-Generation Vaccines: Features of Nucleic Acid Vaccines and Strategies to Improve Their Efficiency

 , , ,
, , ,  , , and
, , and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
1. Introduction
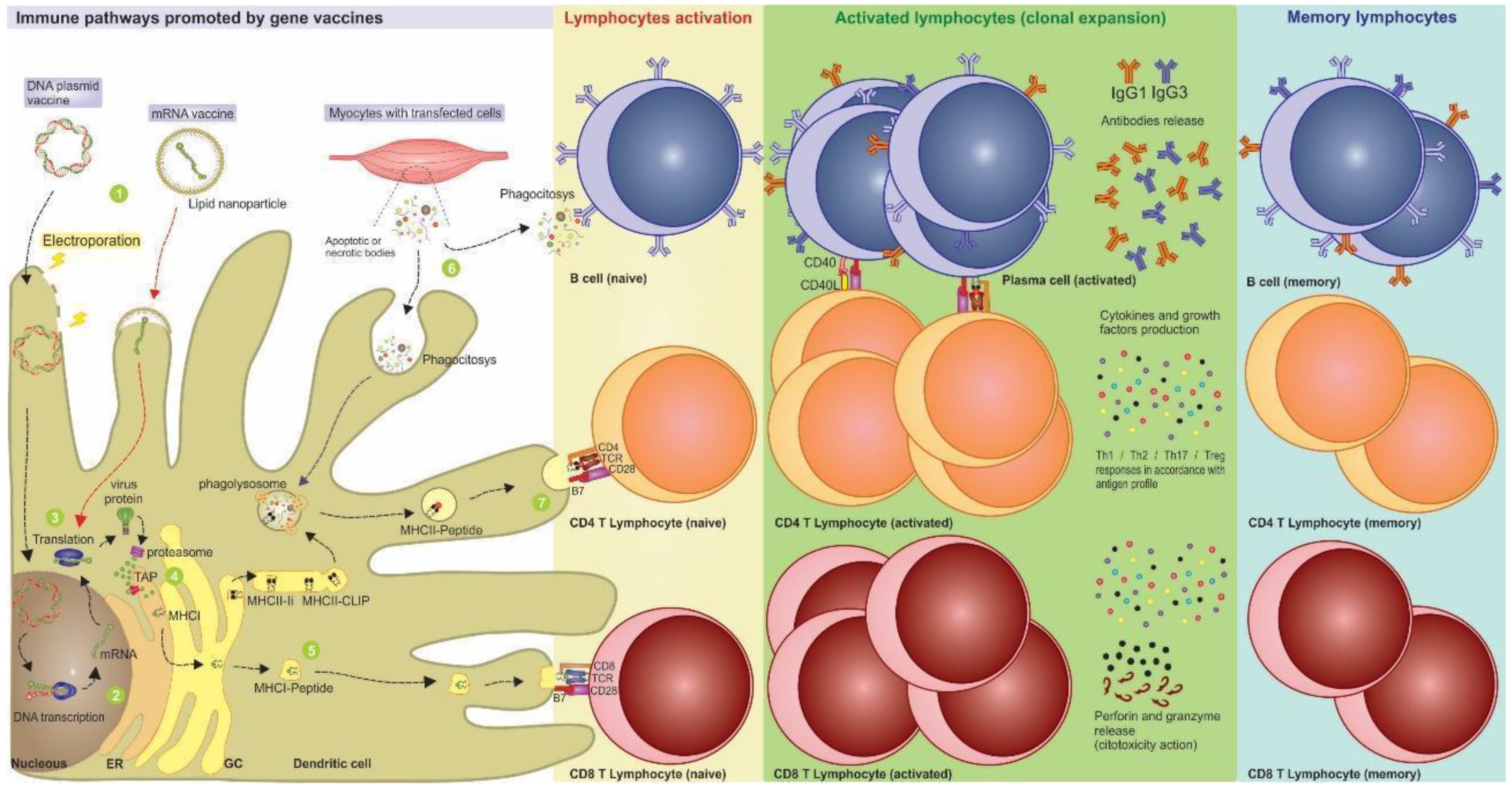
2. New Technologies: Gene-based Vaccines
3. Nucleic Acid Vaccines Allow Better Immune Response Directing
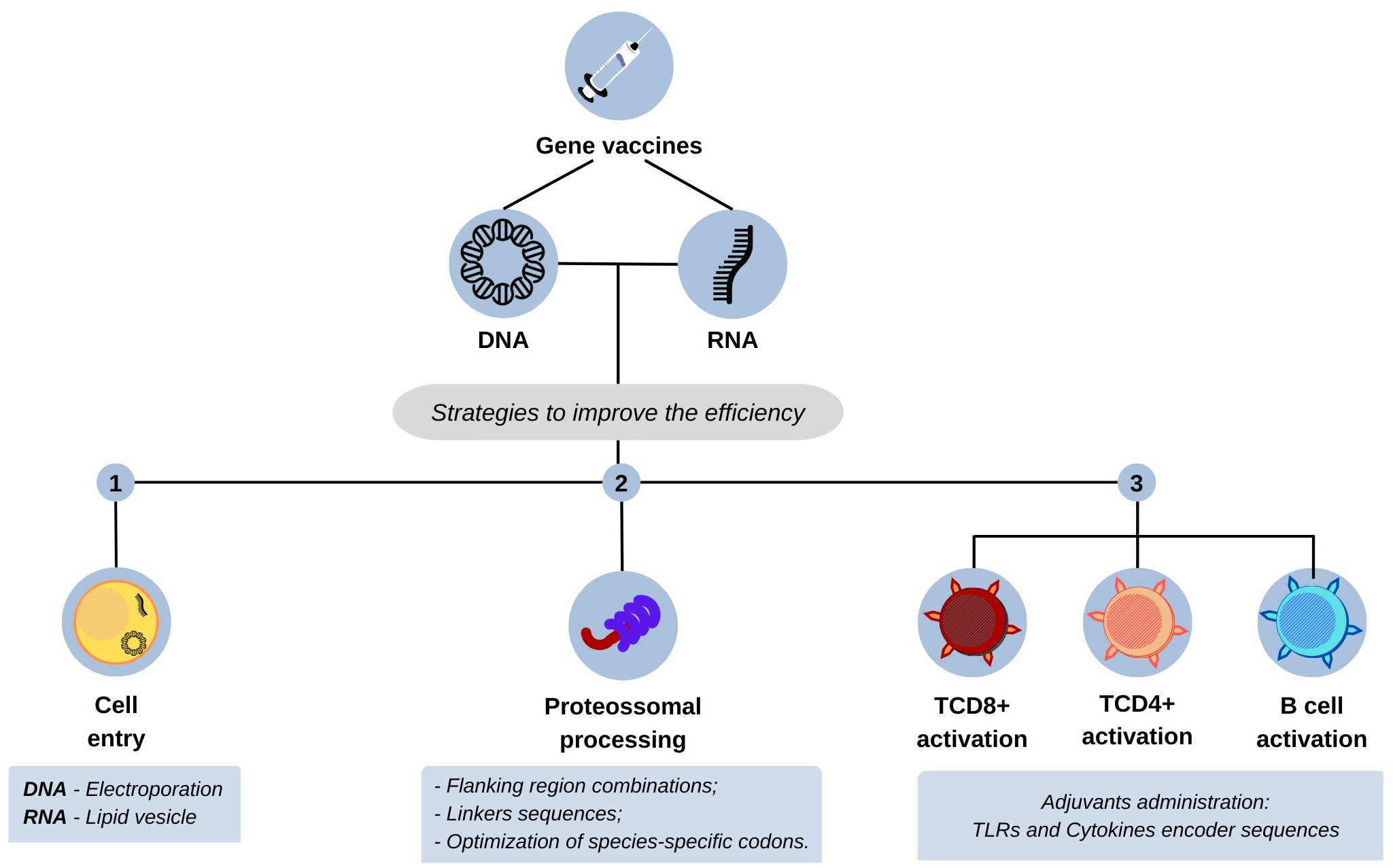
4. Strategies to Improve the Efficiency of Nucleic Acid Vaccines
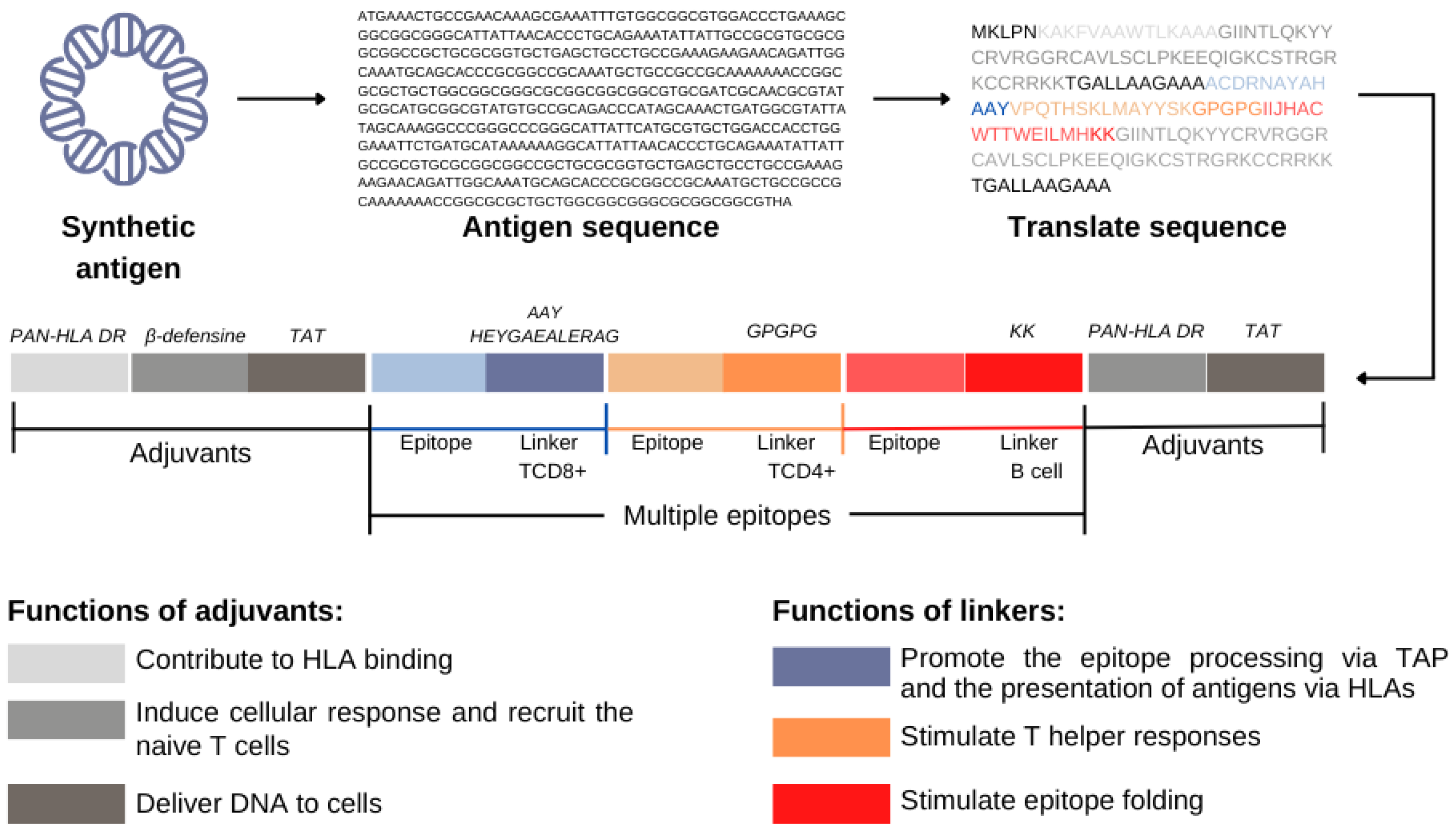
4.1. Enhancement of Presentation Efficacy of Epitopes to the MHC System
4.2. Increasing Vaccine Immunogenicity
4.3. Vaccine Transfection Efficacy Enhancement
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Domínguez-Andrés, J.; van Crevel, R.; Divangahi, M.; Netea, M.G. Designing the Next Generation of Vaccines: Relevance for Future Pandemics. mBio 2020, 11, e02616–e02620. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Naghavi, M.; Allen, C.; Barber, R.M.; Bhutta, Z.A.; Carter, A.; Casey, D.C.; Charlson, F.J.; Chen, A.Z.; Coates, M.M.; et al. Global, Regional, and National Life Expectancy, All-Cause Mortality, and Cause-Specific Mortality for 249 Causes of Death, 1980–2015: A Systematic Analysis for the Global Burden of Disease Study 2015. Lancet 2016, 388, 1459–1544. [Google Scholar] [CrossRef] [Green Version]
- Plotkin, S.A. Vaccines: Past, Present and Future. Nat. Med. 2005, 11, S5–S11. [Google Scholar] [CrossRef] [PubMed]
- Francis, M.J. Recent Advances in Vaccine Technologies. Vet. Clin. North Am. Small Anim. Pract. 2018, 48, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Flanagan, K.L.; MacIntyre, C.R.; McIntyre, P.B.; Nelson, M.R. SARS-CoV-2 Vaccines: Where Are We Now? J. Allergy Clin. Immunol. Pract. 2021, 9, 3535–3543. [Google Scholar] [CrossRef] [PubMed]
- Rappuoli, R.; De Gregorio, E.; Del Giudice, G.; Phogat, S.; Pecetta, S.; Pizza, M.; Hanon, E. Vaccinology in the Post−COVID-19 Era. Proc. Natl. Acad. Sci. USA 2021, 118, e2020368118. [Google Scholar] [CrossRef] [PubMed]
- González-Romo, F.; Picazo, J.J. El desarrollo de nuevas vacunas. Enferm. Infecc. Y Microbiol. Clínica 2015, 33, 557–568. [Google Scholar] [CrossRef]
- Carter, C.; Houser, K.V.; Yamshchikov, G.V.; Bellamy, A.R.; May, J.; Enama, M.E.; Sarwar, U.; Larkin, B.; Bailer, R.T.; Koup, R.; et al. Safety and Immunogenicity of Investigational Seasonal Influenza Hemagglutinin DNA Vaccine Followed by Trivalent Inactivated Vaccine Administered Intradermally or Intramuscularly in Healthy Adults: An Open-Label Randomized Phase 1 Clinical Trial. PLoS ONE 2019, 14, e0222178. [Google Scholar] [CrossRef]
- Barbier, A.J.; Jiang, A.Y.; Zhang, P.; Wooster, R.; Anderson, D.G. The Clinical Progress of MRNA Vaccines and Immunotherapies. Nat. Biotechnol. 2022, 40, 840–854. [Google Scholar] [CrossRef]
- Tombácz, I.; Weissman, D.; Pardi, N. Vaccination with Messenger RNA: A Promising Alternative to DNA Vaccination. In DNA Vaccines; Sousa, Â., Ed.; Methods in Molecular Biology; Springer: New York, NY, USA, 2021; Volume 2197, pp. 13–31. ISBN 978-1-07-160871-5. [Google Scholar]
- Wadhwa, A.; Aljabbari, A.; Lokras, A.; Foged, C.; Thakur, A. Opportunities and Challenges in the Delivery of MRNA-Based Vaccines. Pharmaceutics 2020, 12, 102. [Google Scholar] [CrossRef]
- Sheets, R.L.; Stein, J.; Manetz, T.S.; Duffy, C.; Nason, M.; Andrews, C.; Kong, W.-P.; Nabel, G.J.; Gomez, P.L. Biodistribution of DNA Plasmid Vaccines against HIV-1, Ebola, Severe Acute Respiratory Syndrome, or West Nile Virus Is Similar, without Integration, despite Differing Plasmid Backbones or Gene Inserts. Toxicol. Sci. 2006, 91, 610–619. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.-C.E.; Livingstone, A.M. Cutting Edge: CD4+ T Cell Help Can Be Essential for Primary CD8 + T Cell Responses In Vivo. J. Immunol. 2003, 171, 6339–6343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.-H.; Danishmalik, S.N.; Sin, J.-I. DNA Vaccines, Electroporation and Their Applications in Cancer Treatment. Hum. Vaccines Immunother. 2015, 11, 1889–1900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, S.; Tang, X.; Chen, Y.; Chen, K.; Fan, N.; Xiao, W.; Zheng, Q.; Li, G.; Teng, Y.; Wu, M.; et al. MRNA-Based Therapeutics: Powerful and Versatile Tools to Combat Diseases. Signal Transduct. Target. Ther. 2022, 7, 166. [Google Scholar] [CrossRef] [PubMed]
- Baden, L.R.; El Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B.; et al. Efficacy and Safety of the MRNA-1273 SARS-CoV-2 Vaccine. N. Engl. J. Med. 2021, 384, 403–416. [Google Scholar] [CrossRef]
- Jackson, L.A.; Anderson, E.J.; Rouphael, N.G.; Roberts, P.C.; Makhene, M.; Coler, R.N.; McCullough, M.P.; Chappell, J.D.; Denison, M.R.; Stevens, L.J.; et al. An MRNA Vaccine against SARS-CoV-2—Preliminary Report. N. Engl. J. Med. 2020, 383, 1920–1931. [Google Scholar] [CrossRef]
- Rittig, S.M.; Haentschel, M.; Weimer, K.J.; Heine, A.; Muller, M.R.; Brugger, W.; Horger, M.S.; Maksimovic, O.; Stenzl, A.; Hoerr, I.; et al. Intradermal Vaccinations With RNA Coding for TAA Generate CD8+ and CD4+ Immune Responses and Induce Clinical Benefit in Vaccinated Patients. Mol. Ther. 2011, 19, 990–999. [Google Scholar] [CrossRef]
- Cafri, G.; Gartner, J.J.; Zaks, T.; Hopson, K.; Levin, N.; Paria, B.C.; Parkhurst, M.R.; Yossef, R.; Lowery, F.J.; Jafferji, M.S.; et al. MRNA Vaccine–Induced Neoantigen-Specific T Cell Immunity in Patients with Gastrointestinal Cancer. J. Clin. Investig. 2020, 130, 5976–5988. [Google Scholar] [CrossRef]
- Trimble, C.L.; Morrow, M.P.; Kraynyak, K.A.; Shen, X.; Dallas, M.; Yan, J.; Edwards, L.; Parker, R.L.; Denny, L.; Giffear, M.; et al. Safety, Efficacy, and Immunogenicity of VGX-3100, a Therapeutic Synthetic DNA Vaccine Targeting Human Papillomavirus 16 and 18 E6 and E7 Proteins for Cervical Intraepithelial Neoplasia 2/3: A Randomised, Double-Blind, Placebo-Controlled Phase 2b Trial. Lancet 2015, 386, 2078–2088. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.J.; Jin, H.-T.; Hur, S.-Y.; Yang, H.G.; Seo, Y.B.; Hong, S.R.; Lee, C.-W.; Kim, S.; Woo, J.-W.; Park, K.S.; et al. Clearance of Persistent HPV Infection and Cervical Lesion by Therapeutic DNA Vaccine in CIN3 Patients. Nat. Commun. 2014, 5, 5317. [Google Scholar] [CrossRef]
- Houseley, J.; Tollervey, D. The Many Pathways of RNA Degradation. Cell 2009, 136, 763–776. [Google Scholar] [CrossRef] [Green Version]
- Khobragade, A.; Bhate, S.; Ramaiah, V.; Deshpande, S.; Giri, K.; Phophle, H.; Supe, P.; Godara, I.; Revanna, R.; Nagarkar, R.; et al. Efficacy, Safety, and Immunogenicity of the DNA SARS-CoV-2 Vaccine (ZyCoV-D): The Interim Efficacy Results of a Phase 3, Randomised, Double-Blind, Placebo-Controlled Study in India. Lancet 2022, 399, 1313–1321. [Google Scholar] [CrossRef] [PubMed]
- van de Wall, S.; Ljungberg, K.; Ip, P.P.; Boerma, A.; Knudsen, M.L.; Nijman, H.W.; Liljeström, P.; Daemen, T. Potent Therapeutic Efficacy of an Alphavirus Replicon DNA Vaccine Expressing Human Papilloma Virus E6 and E7 Antigens. OncoImmunology 2018, 7, e1487913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ljungberg, K.; Liljeström, P. Self-Replicating Alphavirus RNA Vaccines. Expert Rev. Vaccines 2015, 14, 177–194. [Google Scholar] [CrossRef] [PubMed]
- Le, T.P.; Coonan, K.M.; Hedstrom, R.C.; Charoenvit, Y.; Sedegah, M.; Epstein, J.E.; Kumar, S.; Wang, R.; Doolan, D.L.; Maguire, J.D.; et al. Safety, Tolerability and Humoral Immune Responses after Intramuscular Administration of a Malaria DNA Vaccine to Healthy Adult Volunteers. Vaccine 2000, 18, 1893–1901. [Google Scholar] [CrossRef]
- Palucka, K.; Banchereau, J. Cancer Immunotherapy via Dendritic Cells. Nat. Rev. Cancer 2012, 12, 265–277. [Google Scholar] [CrossRef]
- Chahal, J.S.; Khan, O.F.; Cooper, C.L.; McPartlan, J.S.; Tsosie, J.K.; Tilley, L.D.; Sidik, S.M.; Lourido, S.; Langer, R.; Bavari, S.; et al. Dendrimer-RNA Nanoparticles Generate Protective Immunity against Lethal Ebola, H1N1 Influenza, and Toxoplasma Gondii Challenges with a Single Dose. Proc. Natl. Acad. Sci. USA 2016, 113, E4133–E4142. [Google Scholar] [CrossRef] [Green Version]
- Stitz, L.; Vogel, A.; Schnee, M.; Voss, D.; Rauch, S.; Mutzke, T.; Ketterer, T.; Kramps, T.; Petsch, B. A Thermostable Messenger RNA Based Vaccine against Rabies. PLoS Negl. Trop. Dis. 2017, 11, e0006108. [Google Scholar] [CrossRef] [Green Version]
- Liu A Comparison of Plasmid DNA and MRNA as Vaccine Technologies. Vaccines 2019, 7, 37. [CrossRef] [Green Version]
- Hollister, K.; Chen, Y.; Wang, S.; Wu, H.; Mondal, A.; Clegg, N.; Lu, S.; Dent, A. The Role of Follicular Helper T Cells and the Germinal Center in HIV-1 Gp120 DNA Prime and Gp120 Protein Boost Vaccination. Hum. Vaccines Immunother. 2014, 10, 1985–1992. [Google Scholar] [CrossRef]
- Holdsworth, S.R.; Kitching, A.R.; Tipping, P.G. Th1 and Th2 T Helper Cell Subsets Affect Patterns of Injury and Outcomes in Glomerulonephritis. Kidney Int. 1999, 55, 1198–1216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delgado, M.F.; Coviello, S.; Monsalvo, A.C.; Melendi, G.A.; Hernandez, J.Z.; Batalle, J.P.; Diaz, L.; Trento, A.; Chang, H.-Y.; Mitzner, W.; et al. Lack of Antibody Affinity Maturation Due to Poor Toll-like Receptor Stimulation Leads to Enhanced Respiratory Syncytial Virus Disease. Nat. Med. 2009, 15, 34–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, W.S.; Wheatley, A.K.; Kent, S.J.; DeKosky, B.J. Antibody-Dependent Enhancement and SARS-CoV-2 Vaccines and Therapies. Nat. Microbiol. 2020, 5, 1185–1191. [Google Scholar] [CrossRef] [PubMed]
- Suschak, J.J.; Williams, J.A.; Schmaljohn, C.S. Advancements in DNA Vaccine Vectors, Non-Mechanical Delivery Methods, and Molecular Adjuvants to Increase Immunogenicity. Hum. Vaccines Immunother. 2017, 13, 2837–2848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Invenção, M.C.D.V.; Melo, A.R.d.S.; de Macêdo, L.S.; da Costa Neves, T.S.P.; de Melo, C.M.L.; Cordeiro, M.N.; de Aragão Batista, M.V.; de Freitas, A.C. Development of Synthetic Antigen Vaccines for COVID-19. Hum. Vaccines Immunother. 2021, 17, 3855–3870. [Google Scholar] [CrossRef]
- Sanami, S.; Azadegan-Dehkordi, F.; Rafieian-Kopaei, M.; Salehi, M.; Ghasemi-Dehnoo, M.; Mahooti, M.; Alizadeh, M.; Bagheri, N. Design of a Multi-Epitope Vaccine against Cervical Cancer Using Immunoinformatics Approaches. Sci. Rep. 2021, 11, 12397. [Google Scholar] [CrossRef]
- Sanami, S.; Rafieian-Kopaei, M.; Dehkordi, K.A.; Pazoki-Toroudi, H.; Azadegan-Dehkordi, F.; Mobini, G.-R.; Alizadeh, M.; Nezhad, M.S.; Ghasemi-Dehnoo, M.; Bagheri, N. In Silico Design of a Multi-Epitope Vaccine against HPV16/18. BMC Bioinform. 2022, 23, 311. [Google Scholar] [CrossRef]
- Kumar, A.; Sahu, U.; Kumari, P.; Dixit, A.; Khare, P. Designing of Multi-Epitope Chimeric Vaccine Using Immunoinformatic Platform by Targeting Oncogenic Strain HPV 16 and 18 against Cervical Cancer. Sci. Rep. 2022, 12, 9521. [Google Scholar] [CrossRef]
- Frelin, L.; Ahlén, G.; Alheim, M.; Weiland, O.; Barnfield, C.; Liljeström, P.; Sällberg, M. Codon Optimization and MRNA Amplification Effectively Enhances the Immunogenicity of the Hepatitis C Virus Nonstructural 3/4A Gene. Gene Ther. 2004, 11, 522–533. [Google Scholar] [CrossRef] [Green Version]
- Ramakrishna, L.; Anand, K.K.; Mohankumar, K.M.; Ranga, U. Codon Optimization of the Tat Antigen of Human Immunodeficiency Virus Type 1 Generates Strong Immune Responses in Mice Following Genetic Immunization. J. Virol. 2004, 78, 9174–9189. [Google Scholar] [CrossRef]
- Livingston, B.D.; Newman, M.; Crimi, C.; McKinney, D.; Chesnut, R.; Sette, A. Optimization of Epitope Processing Enhances Immunogenicity of Multiepitope DNA Vaccines. Vaccine 2001, 19, 4652–4660. [Google Scholar] [CrossRef] [PubMed]
- Yano, A.; Onozuka, A.; Asahiozaki, Y.; Imai, S.; Hanada, N.; Miwa, Y.; Nisizawa, T. An Ingenious Design for Peptide Vaccines. Vaccine 2005, 23, 2322–2326. [Google Scholar] [CrossRef] [PubMed]
- Livingston, B.; Crimi, C.; Newman, M.; Higashimoto, Y.; Appella, E.; Sidney, J.; Sette, A. A Rational Strategy to Design Multiepitope Immunogens Based on Multiple Th Lymphocyte Epitopes. J. Immunol. 2002, 168, 5499–5506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dolenc, I.; Seemüller, E.; Baumeister, W. Decelerated Degradation of Short Peptides by the 20S Proteasome. FEBS Lett. 1998, 434, 357–361. [Google Scholar] [CrossRef] [Green Version]
- Wei, C.-J.; Boyington, J.C.; Dai, K.; Houser, K.V.; Pearce, M.B.; Kong, W.-P.; Yang, Z.; Tumpey, T.M.; Nabel, G.J. Cross-Neutralization of 1918 and 2009 Influenza Viruses: Role of Glycans in Viral Evolution and Vaccine Design. Sci. Transl. Med. 2010, 2, 24ra21. [Google Scholar] [CrossRef] [Green Version]
- Kutzler, M.A.; Weiner, D.B. DNA Vaccines: Ready for Prime Time? Nat. Rev. Genet. 2008, 9, 776–788. [Google Scholar] [CrossRef]
- Eusébio, D.; Neves, A.R.; Costa, D.; Biswas, S.; Alves, G.; Cui, Z.; Sousa, Â. Methods to Improve the Immunogenicity of Plasmid DNA Vaccines. Drug Discov. Today 2021, 26, 2575–2592. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, C.; Zhang, L.; Li, J.; Huang, Z.; Lu, S. The Relative Immunogenicity of DNA Vaccines Delivered by the Intramuscular Needle Injection, Electroporation and Gene Gun Methods. Vaccine 2008, 26, 2100–2110. [Google Scholar] [CrossRef] [Green Version]
- Newton, K.; Dixit, V.M. Signaling in Innate Immunity and Inflammation. Cold Spring Harb. Perspect. Biol. 2012, 4, a006049. [Google Scholar] [CrossRef] [Green Version]
- Pone, E.J.; Zan, H.; Zhang, J.; Al-Qahtani, A.; Xu, Z.; Casali, P. Toll-Like Receptors and B-Cell Receptors Synergize to Induce Immunoglobulin Class-Switch DNA Recombination: Relevance to Microbial Antibody Responses. Crit. Rev. Immunol. 2010, 30, 1–29. [Google Scholar] [CrossRef]
- Ramakrishna, V.; Vasilakos, J.P.; Tario, J.D.; Berger, M.A.; Wallace, P.K.; Keler, T. Toll-like Receptor Activation Enhances Cell-Mediated Immunity Induced by an Antibody Vaccine Targeting Human Dendritic Cells. J. Transl. Med. 2007, 5, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roman, M.; Martin-Orozco, E.; Goodman, J.S.; Nguyen, M.-D.; Sato, Y.; Ronaghy, A.; Kornbluth, R.S.; Richman, D.D.; Carson, D.A.; Raz, E. Immunostimulatory DNA Sequences Function as T Helper-1-Promoting Adjuvants. Nat. Med. 1997, 3, 849–854. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Yamane, H.; Paul, W.E. Differentiation of Effector CD4 T Cell Populations. Annu. Rev. Immunol. 2010, 28, 445–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Longhi, M.P.; Trumpfheller, C.; Idoyaga, J.; Caskey, M.; Matos, I.; Kluger, C.; Salazar, A.M.; Colonna, M.; Steinman, R.M. Dendritic Cells Require a Systemic Type I Interferon Response to Mature and Induce CD4+ Th1 Immunity with Poly IC as Adjuvant. J. Exp. Med. 2009, 206, 1589–1602. [Google Scholar] [CrossRef]
- Fourcade, J.; Kudela, P.; Andrade Filho, P.A.; Janjic, B.; Land, S.R.; Sander, C.; Krieg, A.; Donnenberg, A.; Shen, H.; Kirkwood, J.M.; et al. Immunization With Analog Peptide in Combination With CpG and Montanide Expands Tumor Antigen-Specific CD8+ T Cells in Melanoma Patients. J. Immunother. 2008, 31, 781–791. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Wang, H.; Zheng, X.; Xue, X.; Wang, B.; Wu, H.; Zhang, K.; Fan, S.; Wang, T.; Li, N.; et al. CpG/Poly (I:C) Mixed Adjuvant Priming Enhances the Immunogenicity of a DNA Vaccine against Eastern Equine Encephalitis Virus in Mice. Int. Immunopharmacol. 2014, 19, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Sajadian, A.; Tabarraei, A.; Soleimanjahi, H.; Fotouhi, F.; Gorji, A.; Ghaemi, A. Comparing the Effect of Toll-like Receptor Agonist Adjuvants on the Efficiency of a DNA Vaccine. Arch. Virol. 2014, 159, 1951–1960. [Google Scholar] [CrossRef] [Green Version]
- Öhlschläger, P.; Spies, E.; Alvarez, G.; Quetting, M.; Groettrup, M. The Combination of TLR-9 Adjuvantation and Electroporation-Mediated Delivery Enhances in Vivo Antitumor Responses after Vaccination with HPV-16 E7 Encoding DNA. Int. J. Cancer 2011, 128, 473–481. [Google Scholar] [CrossRef] [Green Version]
- Sabino, E.C.; Buss, L.F.; Carvalho, M.P.S.; Prete, C.A.; Crispim, M.A.E.; Fraiji, N.A.; Pereira, R.H.M.; Parag, K.V.; da Silva Peixoto, P.; Kraemer, M.U.G.; et al. Resurgence of COVID-19 in Manaus, Brazil, despite High Seroprevalence. Lancet 2021, 397, 452–455. [Google Scholar] [CrossRef]
- Salem, M.L.; Kadima, A.N.; Cole, D.J.; Gillanders, W.E. Defining the Antigen-Specific T-Cell Response to Vaccination and Poly(I:C)/TLR3 Signaling: Evidence of Enhanced Primary and Memory CD8 T-Cell Responses and Antitumor Immunity. J. Immunother. 2005, 28, 220–228. [Google Scholar] [CrossRef]
- Hirao, L.A.; Wu, L.; Khan, A.S.; Hokey, D.A.; Yan, J.; Dai, A.; Betts, M.R.; Draghia-Akli, R.; Weiner, D.B. Combined Effects of IL-12 and Electroporation Enhances the Potency of DNA Vaccination in Macaques. Vaccine 2008, 26, 3112–3120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henry, C.J.; Ornelles, D.A.; Mitchell, L.M.; Brzoza-Lewis, K.L.; Hiltbold, E.M. IL-12 Produced by Dendritic Cells Augments CD8 + T Cell Activation through the Production of the Chemokines CCL1 and CCL17. J. Immunol. 2008, 181, 8576–8584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsieh, C.-S.; Macatonia, S.E.; Tripp, C.S.; Wolf, S.F.; O’Garra, A.; Murphy, K.M. Development of TH1 CD4+ T Cells Through IL-12 Produced by Listeria -Induced Macrophages. Science 1993, 260, 547–549. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Petrovsky, N. Molecular Adjuvants for DNA Vaccines. Curr. Issues Mol. Biol. 2017, 22, 17–40. [Google Scholar] [CrossRef]
- Hobernik, D.; Bros, M. DNA Vaccines—How Far From Clinical Use? IJMS 2018, 19, 3605. [Google Scholar] [CrossRef] [Green Version]
- Lapuente, D.; Stab, V.; Storcksdieck genannt Bonsmann, M.; Maaske, A.; Köster, M.; Xiao, H.; Ehrhardt, C.; Tenbusch, M. Innate Signalling Molecules as Genetic Adjuvants Do Not Alter the Efficacy of a DNA-Based Influenza A Vaccine. PLoS ONE 2020, 15, e0231138. [Google Scholar] [CrossRef]
- Xu, R.; Megati, S.; Roopchand, V.; Luckay, A.; Masood, A.; Garcia-Hand, D.; Rosati, M.; Weiner, D.B.; Felber, B.K.; Pavlakis, G.N.; et al. Comparative Ability of Various Plasmid-Based Cytokines and Chemokines to Adjuvant the Activity of HIV Plasmid DNA Vaccines. Vaccine 2008, 26, 4819–4829. [Google Scholar] [CrossRef]
- Kumari, R.; Kole, S.; Soman, P.; Rathore, G.; Tripathi, G.; Makesh, M.; Rajendran, K.V.; Bedekar, M.K. Bicistronic DNA Vaccine against Edwardsiella Tarda Infection in Labeo Rohita: Construction and Comparative Evaluation of Its Protective Efficacy against Monocistronic DNA Vaccine. Aquaculture 2018, 485, 201–209. [Google Scholar] [CrossRef]
- Kang, S.-M.; Compans, R.W. Host Responses from Innate to Adaptive Immunity after Vaccination: Molecular and Cellular Events. Mol. Cells 2009, 27, 5–14. [Google Scholar] [CrossRef]
- Zuckerman, J.N. The Importance of Injecting Vaccines into Muscle. BMJ 2000, 321, 1237–1238. [Google Scholar] [CrossRef]
- McCluskie, M.J.; Millan, C.L.B.; Gramzinski, R.A.; Robinson, H.L.; Santoro, J.C.; Fuller, J.T.; Widera, G.; Haynes, J.R.; Purcell, R.H.; Davis, H.L. Route and Method of Delivery of DNA Vaccine Influence Immune Responses in Mice and Non-Human Primates. Mol. Med. 1999, 5, 287–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pardi, N.; Parkhouse, K.; Kirkpatrick, E.; McMahon, M.; Zost, S.J.; Mui, B.L.; Tam, Y.K.; Karikó, K.; Barbosa, C.J.; Madden, T.D.; et al. Nucleoside-Modified MRNA Immunization Elicits Influenza Virus Hemagglutinin Stalk-Specific Antibodies. Nat. Commun. 2018, 9, 3361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Midoux, P.; Pichon, C. Lipid-Based MRNA Vaccine Delivery Systems. Expert Rev. Vaccines 2015, 14, 221–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petkov, S.; Starodubova, E.; Latanova, A.; Kilpeläinen, A.; Latyshev, O.; Svirskis, S.; Wahren, B.; Chiodi, F.; Gordeychuk, I.; Isaguliants, M. DNA Immunization Site Determines the Level of Gene Expression and the Magnitude, but Not the Type of the Induced Immune Response. PLoS ONE 2018, 13, e0197902. [Google Scholar] [CrossRef]
- Wagner, E.; Kircheis, R.; Walker, G.F. Targeted Nucleic Acid Delivery into Tumors: New Avenues for Cancer Therapy. Biomed. Pharmacother. 2004, 58, 152–161. [Google Scholar] [CrossRef]
- Cappelletti, M.; Zampaglione, I.; Rizzuto, G.; Ciliberto, G.; Monica, N.L.; Fattori, E. Gene Electro-Transfer Improves Transduction by Modifying the Fate of Intramuscular DNA. J. Gene Med. 2003, 5, 324–332. [Google Scholar] [CrossRef]
- Schultheis, K.; Smith, T.R.F.; Kiosses, W.B.; Kraynyak, K.A.; Wong, A.; Oh, J.; Broderick, K.E. Delineating the Cellular Mechanisms Associated with Skin Electroporation. Hum. Gene Ther. Methods 2018, 29, 177–188. [Google Scholar] [CrossRef]
- Vasan, S.; Hurley, A.; Schlesinger, S.J.; Hannaman, D.; Gardiner, D.F.; Dugin, D.P.; Boente-Carrera, M.; Vittorino, R.; Caskey, M.; Andersen, J.; et al. In Vivo Electroporation Enhances the Immunogenicity of an HIV-1 DNA Vaccine Candidate in Healthy Volunteers. PLoS ONE 2011, 6, e19252. [Google Scholar] [CrossRef] [Green Version]
- Todorova, B.; Adam, L.; Culina, S.; Boisgard, R.; Martinon, F.; Cosma, A.; Ustav, M.; Kortulewski, T.; Le Grand, R.; Chapon, C. Electroporation as a Vaccine Delivery System and a Natural Adjuvant to Intradermal Administration of Plasmid DNA in Macaques. Sci. Rep. 2017, 7, 4122. [Google Scholar] [CrossRef] [Green Version]
- Low, L.; Mander, A.; McCann, K.; Dearnaley, D.; Tjelle, T.; Mathiesen, I.; Stevenson, F.; Ottensmeier, C.H. DNA Vaccination with Electroporation Induces Increased Antibody Responses in Patients with Prostate Cancer. Hum. Gene Ther. 2009, 20, 1269–1278. [Google Scholar] [CrossRef]
- Diken, M.; Kreiter, S.; Selmi, A.; Britten, C.M.; Huber, C.; Türeci, Ö.; Sahin, U. Selective Uptake of Naked Vaccine RNA by Dendritic Cells Is Driven by Macropinocytosis and Abrogated upon DC Maturation. Gene Therapy 2011, 18, 702–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miao, L.; Li, L.; Huang, Y.; Delcassian, D.; Chahal, J.; Han, J.; Shi, Y.; Sadtler, K.; Gao, W.; Lin, J.; et al. Delivery of MRNA Vaccines with Heterocyclic Lipids Increases Anti-Tumor Efficacy by STING-Mediated Immune Cell Activation. Nat. Biotechnol. 2019, 37, 1174–1185. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Melo, A.R.d.S.; de Macêdo, L.S.; Invenção, M.d.C.V.; de Moura, I.A.; da Gama, M.A.T.M.; de Melo, C.M.L.; Silva, A.J.D.; Batista, M.V.d.A.; Freitas, A.C.d. Third-Generation Vaccines: Features of Nucleic Acid Vaccines and Strategies to Improve Their Efficiency. Genes 2022, 13, 2287. https://doi.org/10.3390/genes13122287
Melo ARdS, de Macêdo LS, Invenção MdCV, de Moura IA, da Gama MATM, de Melo CML, Silva AJD, Batista MVdA, Freitas ACd. Third-Generation Vaccines: Features of Nucleic Acid Vaccines and Strategies to Improve Their Efficiency. Genes. 2022; 13(12):2287. https://doi.org/10.3390/genes13122287
Chicago/Turabian StyleMelo, Alanne Rayssa da Silva, Larissa Silva de Macêdo, Maria da Conceição Viana Invenção, Ingrid Andrêssa de Moura, Marco Antonio Turiah Machado da Gama, Cristiane Moutinho Lagos de Melo, Anna Jéssica Duarte Silva, Marcus Vinicius de Aragão Batista, and Antonio Carlos de Freitas. 2022. "Third-Generation Vaccines: Features of Nucleic Acid Vaccines and Strategies to Improve Their Efficiency" Genes 13, no. 12: 2287. https://doi.org/10.3390/genes13122287