


Microarray Analysis Reveals Changes in tRNA-Derived Small RNAs (tsRNAs) Expression in Mice with Septic Cardiomyopathy

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials & Methods
2.1. Animals and Experimental Groups
2.2. Cell Culture
2.3. Cell Transfection Experiments
2.4. Cell Viability
2.5. Lactate Dehydrogenase Assay
2.6. Array Hybridization
2.7. Data Analysis
2.8. RNA Extraction
2.9. RT-qPCR Validation
2.10. Target Gene Prediction
2.11. Gene Ontology (GO) Annotation and Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathway Enrichment Analyses
2.12. Statistical Analysis
3. Results
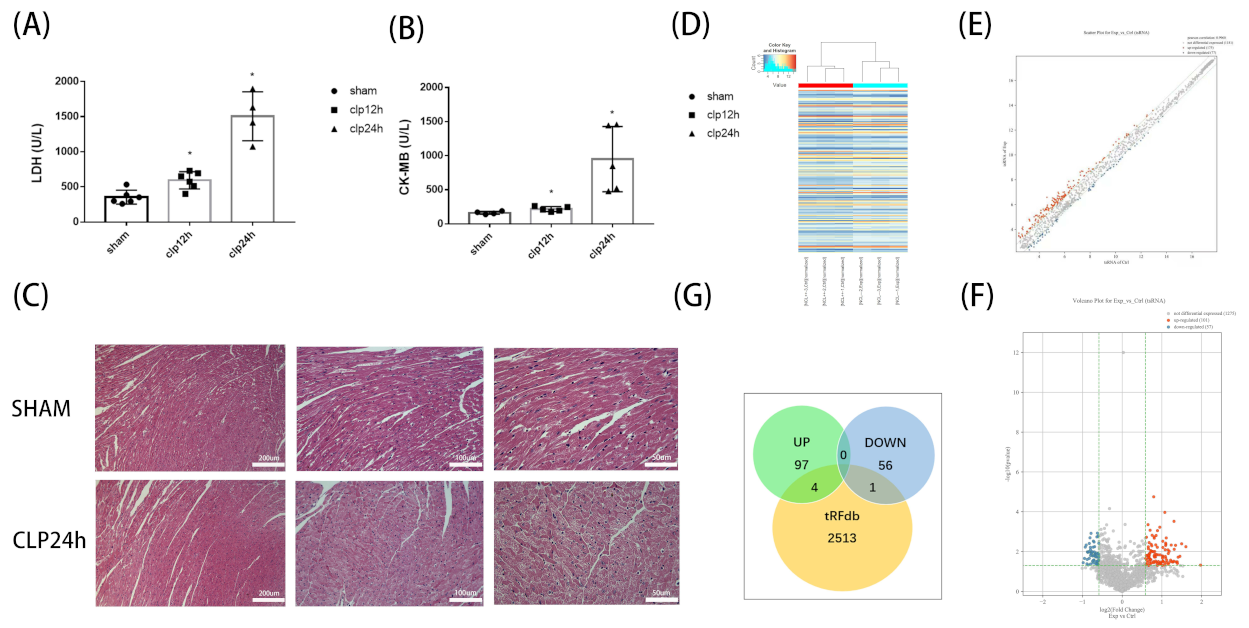
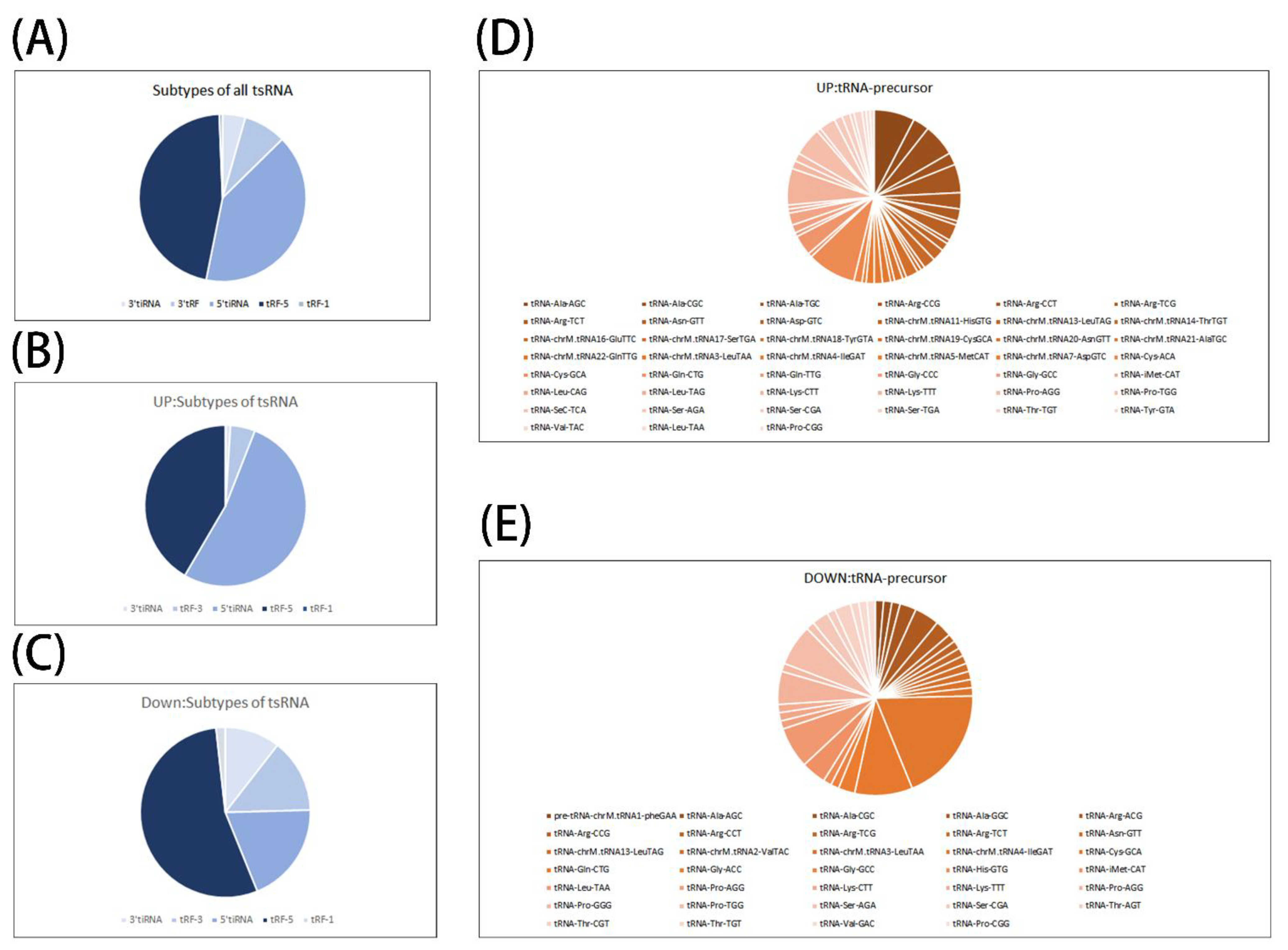
3.1. Alterations of tsRNA Expression Profiles Induced by Septic Cardiomyopathy
3.2. Preliminary Analysis of Microarray Results
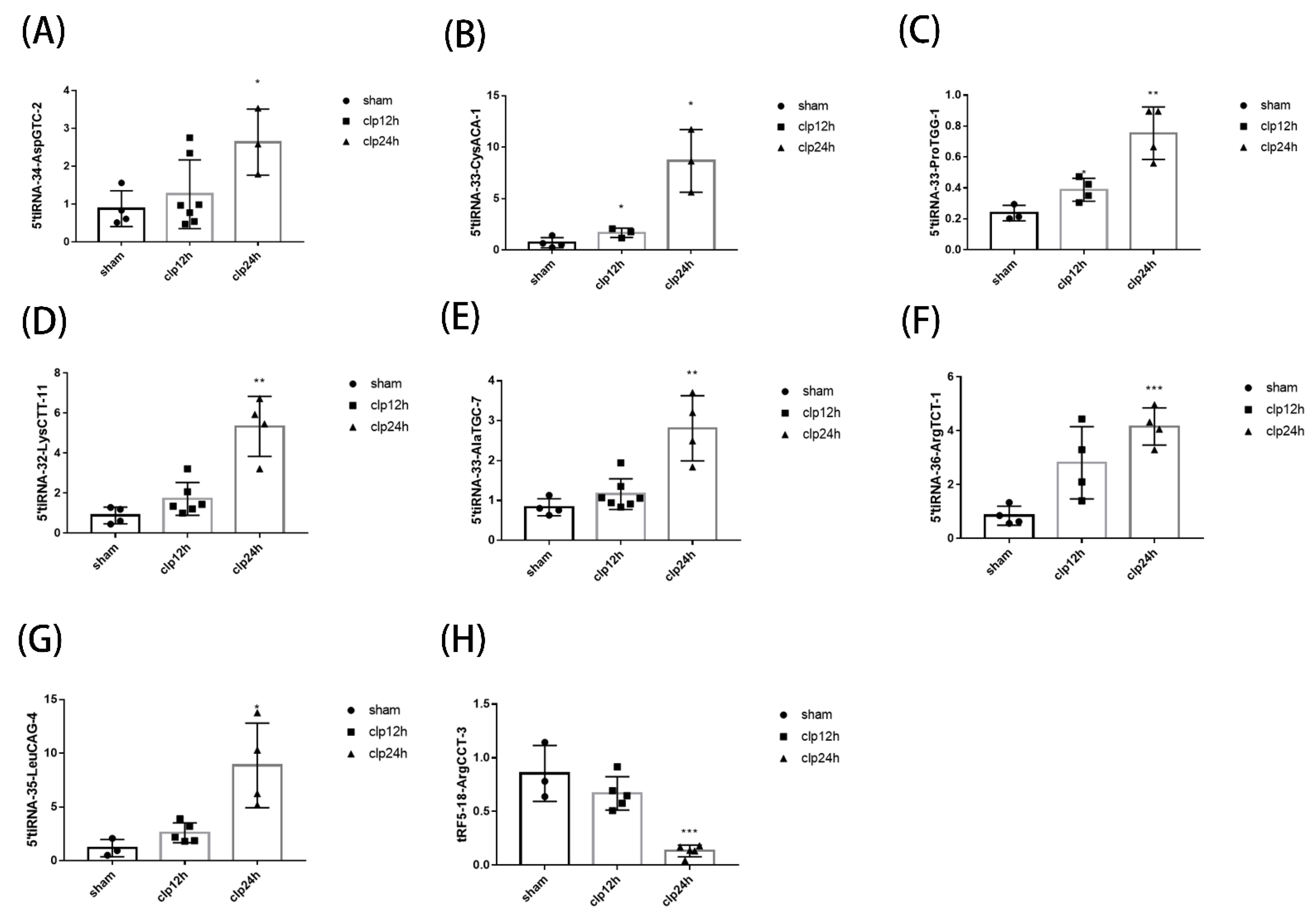
3.3. Identification of tsRNA & qPCR Verification
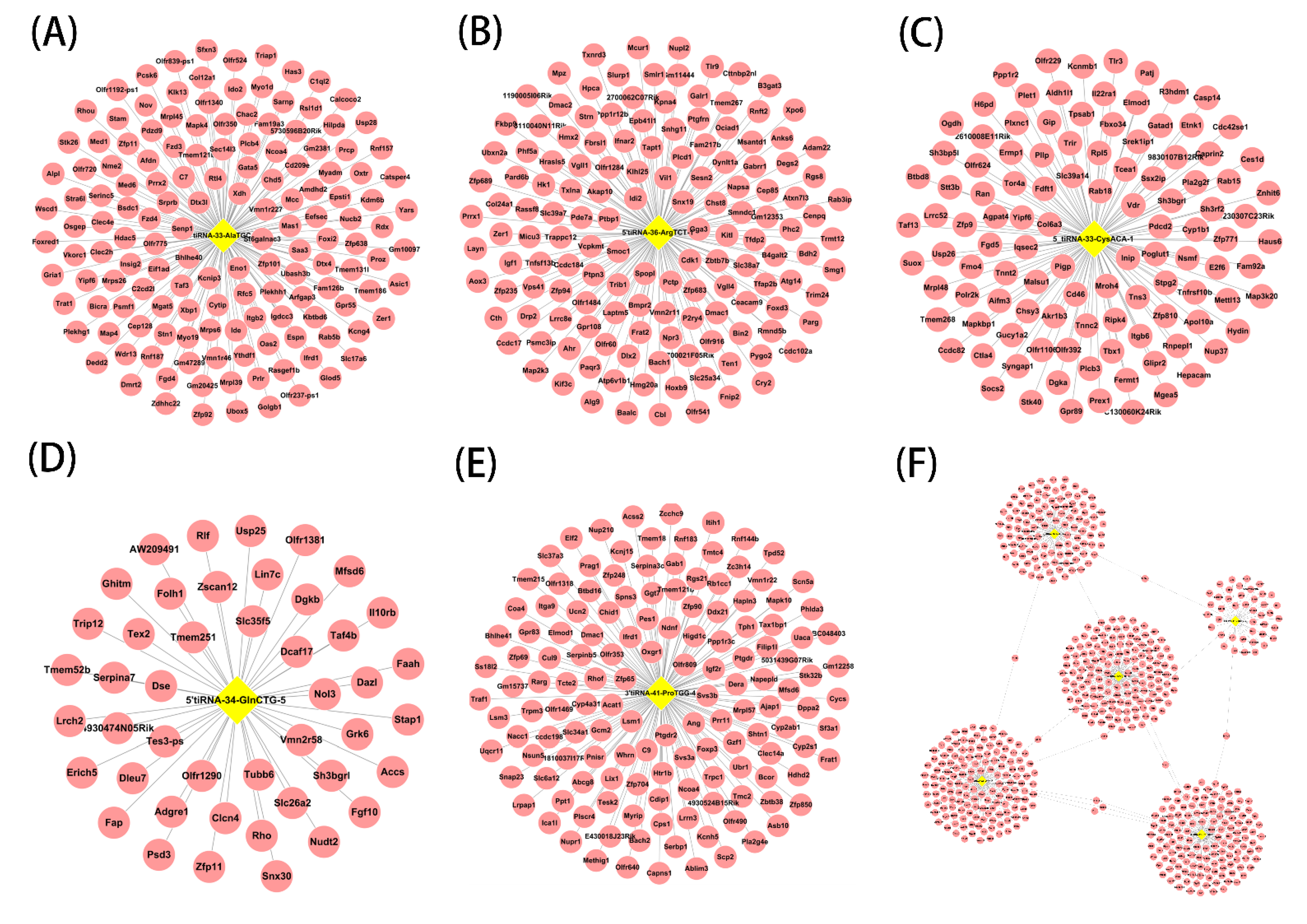
3.4. Targets of Septic Cardiomyopathy-Related tsRNAs
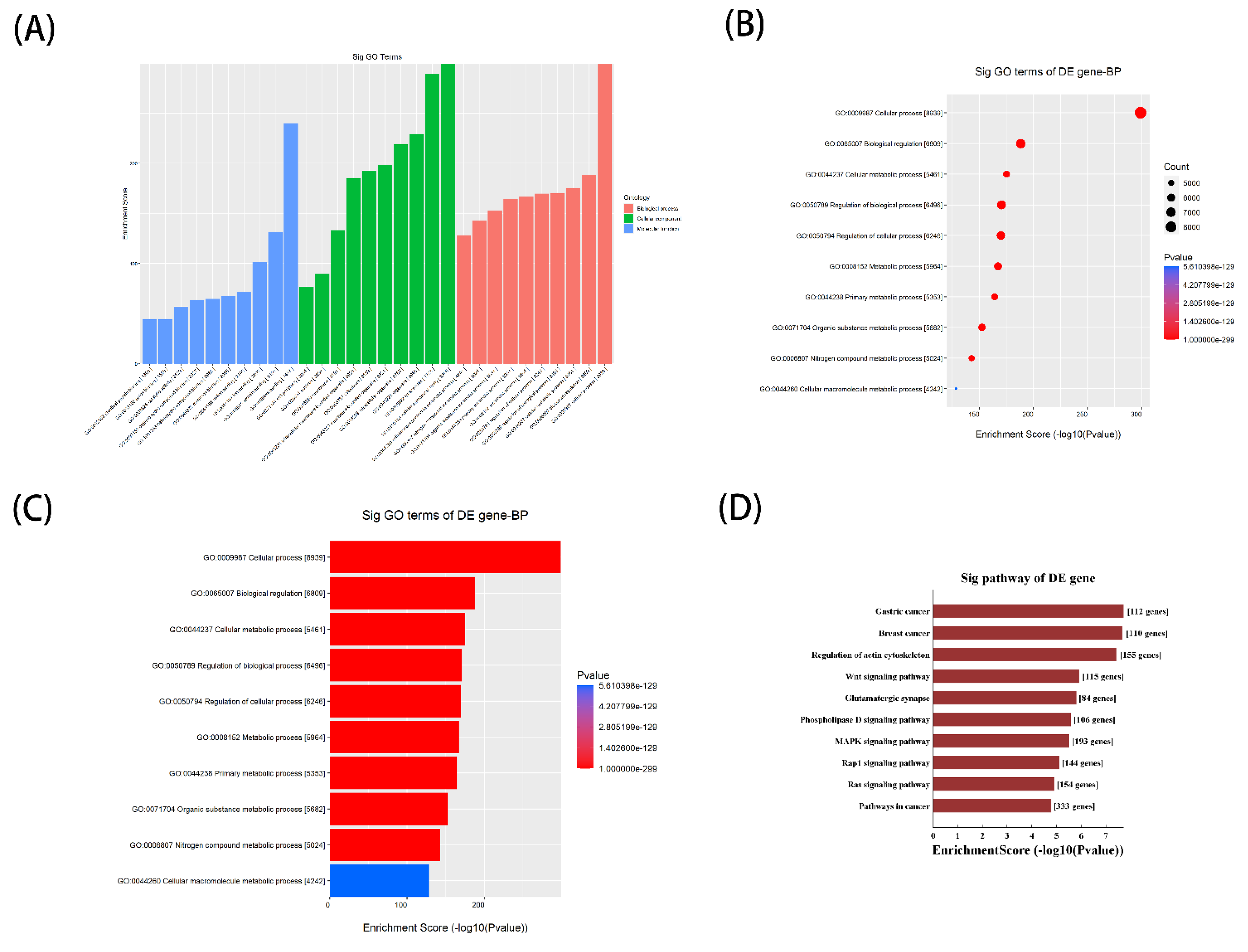
3.5. Potential function of Septic Cardiomyopathy-Related tsRNAs Revealed by GO and KEGG Analysis
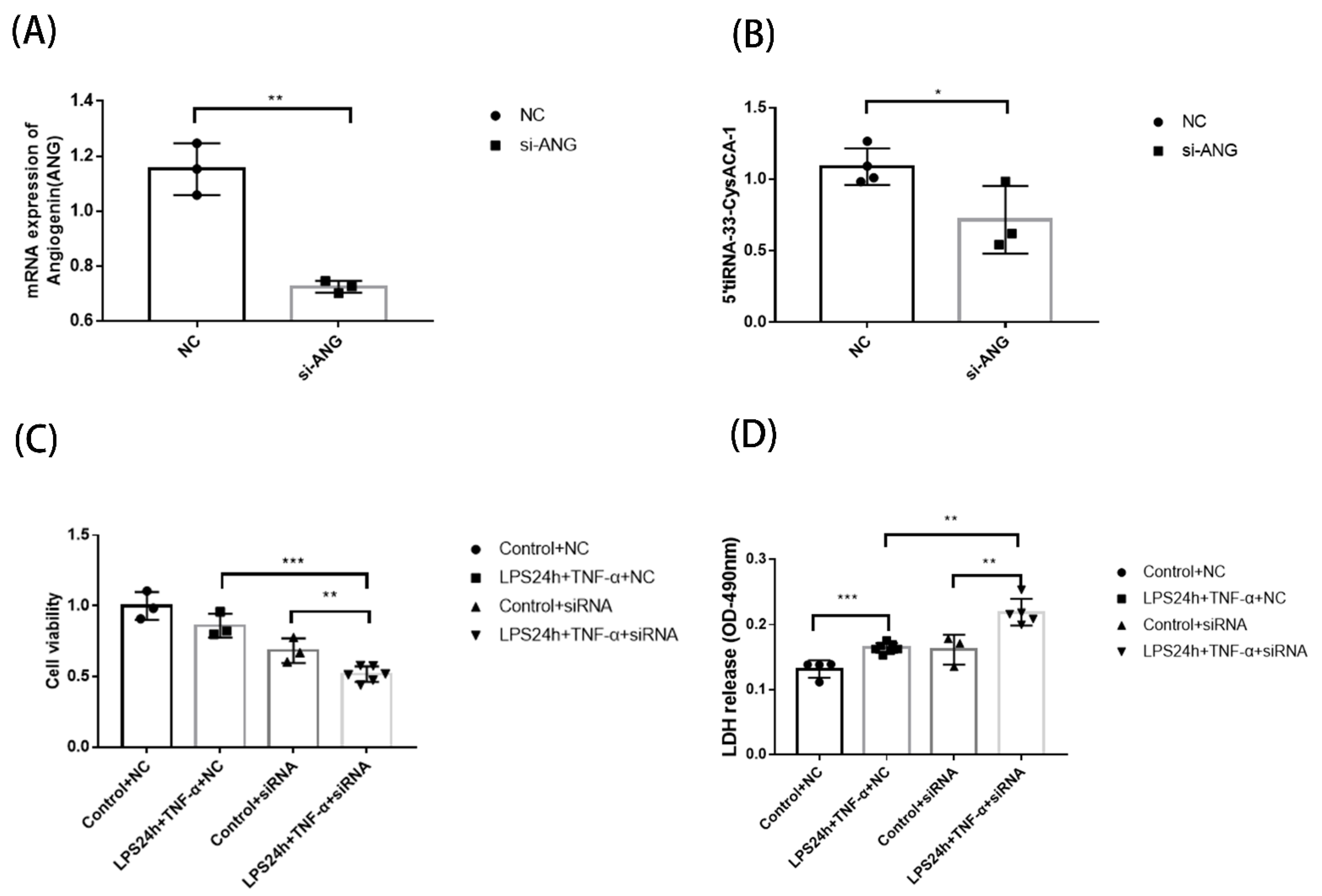
3.6. Function Detection of Septic Cardiomyopathy-Related tsRNAs after ANG Interference
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singer, M.; Deutschman, C.S.; Seymour, C.W.; Shankar-Hari, M.; Annane, D.; Bauer, M.; Bellomo, R.; Bernard, G.R.; Chiche, J.D.; Coopersmith, C.M.; et al. The Third International Consensus Definitions for Sepsis and Septic Shock (Sepsis-3). JAMA 2016, 315, 801–810. [Google Scholar] [CrossRef] [PubMed]
- L’Heureux, M.; Sternberg, M.; Brath, L.; Turlington, J.; Kashiouris, M.G. Sepsis-Induced Cardiomyopathy: A Comprehensive Review. Curr. Cardiol. Rep. 2020, 22, 35. [Google Scholar] [CrossRef]
- Lv, X.; Wang, H. Pathophysiology of sepsis-induced myocardial dysfunction. Mil. Med. Res. 2016, 3, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beesley, S.J.; Weber, G.; Sarge, T.; Nikravan, S.; Grissom, C.K.; Lanspa, M.J.; Shahul, S.; Brown, S.M. Septic Cardiomyopathy. Crit. Care Med. 2018, 46, 625–634. [Google Scholar] [CrossRef]
- Rudd, K.E.; Johnson, S.C.; Agesa, K.M.; Shackelford, K.A.; Tsoi, D.; Kievlan, D.R.; Colombara, D.V.; Ikuta, K.S.; Kissoon, N.; Finfer, S.; et al. Global, regional, and national sepsis incidence and mortality, 1990–2017: Analysis for the Global Burden of Disease Study. Lancet 2020, 395, 200–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, L.; Moore, B.; Soni, A. National Inpatient Hospital Costs: The Most Expensive Conditions by Payer, 2017. 2020. In Healthcare Cost and Utilization Project (HCUP) Statistical Briefs [Internet]; Agency for Healthcare Research and Quality (US): Rockville, MD, USA, 2006; Statistical Brief #261. [Google Scholar]
- Brown, G.L.; Lee, S. Amino-acid-transfer ribonucleic acid: Structure and function. Br. Med. Bull. 1965, 21, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Luo, J.; Zhou, H.; Liao, J.Y.; Ma, L.M.; Chen, Y.Q.; Qu, L.H. Stress-induced tRNA-derived RNAs: A novel class of small RNAs in the primitive eukaryote Giardia lamblia. Nucleic Acids Res. 2008, 36, 6048–6055. [Google Scholar] [CrossRef] [Green Version]
- Jin, F.; Yang, L.; Wang, W.; Yuan, N.; Zhan, S.; Yang, P.; Chen, X.; Ma, T.; Wang, Y. A novel class of tsRNA signatures as biomarkers for diagnosis and prognosis of pancreatic cancer. Mol. Cancer 2021, 20, 95. [Google Scholar] [CrossRef]
- Ha, M.; Kim, V.N. Regulation of microRNA biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef]
- Fett, J.W.; Strydom, D.J.; Lobb, R.R.; Alderman, E.M.; Bethune, J.L.; Riordan, J.F.; Vallee, B.L. Isolation and characterization of angiogenin, an angiogenic protein from human carcinoma cells. Biochemistry 1985, 24, 5480–5486. [Google Scholar] [CrossRef]
- Sheng, J.; Xu, Z. Three decades of research on angiogenin: A review and perspective. Acta Biochim. Biophys. Sin. 2016, 48, 399–410. [Google Scholar] [CrossRef] [Green Version]
- Fu, H.; Feng, J.; Liu, Q.; Sun, F.; Tie, Y.; Zhu, J.; Xing, R.; Sun, Z.; Zheng, X. Stress induces tRNA cleavage by angiogenin in mammalian cells. FEBS Lett. 2009, 583, 437–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamasaki, S.; Ivanov, P.; Hu, G.F.; Anderson, P. Angiogenin cleaves tRNA and promotes stress-induced translational repression. J. Cell Biol. 2009, 185, 35–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, X.; Xie, Y.; Zhang, S.; Song, X.; Xiao, B.; Yan, Z. tRNA-derived fragments: Mechanisms underlying their regulation of gene expression and potential applications as therapeutic targets in cancers and virus infections. Theranostics 2021, 11, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Tao, E.W.; Cheng, W.Y.; Li, W.L.; Yu, J.; Gao, Q.Y. tiRNAs: A novel class of small noncoding RNAs that helps cells respond to stressors and plays roles in cancer progression. J. Cell Physiol. 2020, 235, 683–690. [Google Scholar] [CrossRef]
- Wen, J.T.; Huang, Z.H.; Li, Q.H.; Chen, X.; Qin, H.L.; Zhao, Y. Research progress on the tsRNA classification, function, and application in gynecological malignant tumors. Cell Death Discov. 2021, 7, 388. [Google Scholar] [CrossRef]
- Wang, C.; Yuan, W.; Hu, A.; Lin, J.; Xia, Z.; Yang, C.F.; Li, Y.; Zhang, Z. Dexmedetomidine alleviated sepsis-induced myocardial ferroptosis and septic heart injury. Mol. Med. Rep. 2020, 22, 175–184. [Google Scholar] [CrossRef]
- Cao, L.; Wang, H.; Wang, F.; Xu, D.; Liu, F.; Liu, C. Aβ-induced senescent retinal pigment epithelial cells create a proinflammatory microenvironment in AMD. Investig. Ophthalmol. Vis. Sci. 2013, 54, 3738–3750. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.; Yu, X.; Zhu, L.; Li, T.; Yan, Z.; Guo, J. Transfer RNA-derived fragments and tRNA halves: Biogenesis, biological functions and their roles in diseases. J. Mol. Med. 2018, 96, 1167–1176. [Google Scholar] [CrossRef]
- Lewis, B.P.; Shih, I.H.; Jones-Rhoades, M.W.; Bartel, D.P.; Burge, C.B. Prediction of mammalian microRNA targets. Cell 2003, 115, 787–798. [Google Scholar] [CrossRef]
- Nam, J.W.; Rissland, O.S.; Koppstein, D.; Abreu-Goodger, C.; Jan, C.H.; Agarwal, V.; Yildirim, M.A.; Rodriguez, A.; Bartel, D.P. Global analyses of the effect of different cellular contexts on microRNA targeting. Mol. Cell. 2014, 53, 1031–1043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riffo-Campos, Á.L.; Riquelme, I.; Brebi-Mieville, P. Tools for Sequence-Based miRNA Target Prediction: What to Choose? Int. J. Mol. Sci. 2016, 17, 1987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enright, A.J.; John, B.; Gaul, U.; Tuschl, T.; Sander, C.; Marks, D.S. MicroRNA targets in Drosophila. Genome Biol. 2003, 5, R1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peterson, S.M.; Thompson, J.A.; Ufkin, M.L.; Sathyanarayana, P.; Liaw, L.; Congdon, C.B. Common features of microRNA target prediction tools. Front. Genet. 2014, 5, 23. [Google Scholar] [CrossRef] [Green Version]
- Brandl, K.; Tomisato, W.; Li, X.; Neppl, C.; Pirie, E.; Falk, W.; Xia, Y.; Moresco, E.M.; Baccala, R.; Theofilopoulos, A.N.; et al. Yip1 domain family, member 6 (Yipf6) mutation induces spontaneous intestinal inflammation in mice. Proc. Natl. Acad. Sci. USA 2012, 109, 12650–12655. [Google Scholar] [CrossRef] [Green Version]
- Quiles Del Rey, M.; Mancias, J.D. NCOA4-Mediated Ferritinophagy: A Potential Link to Neurodegeneration. Front. Neurosci. 2019, 13, 238. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Zhang, Y.; Liu, J.; Kang, R.; Klionsky, D.J.; Tang, D. Mitochondrial DNA stress triggers autophagy-dependent ferroptotic death. Autophagy 2021, 17, 948–960. [Google Scholar] [CrossRef]
- George, S.J. Wnt pathway: A new role in regulation of inflammation. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 400–402. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Li, X.; Zhang, X.; Li, Z.; Wang, L.; Sun, Y.; Liu, Z.; Ma, X. Elevated levels of plasma TNF-α are associated with microvascular endothelial dysfunction in patients with sepsis through activating the NF-κB and p38 mitogen-activated protein kinase in endothelial cells. Shock 2014, 41, 275–281. [Google Scholar] [CrossRef]
- Gotts, J.E.; Matthay, M.A. Sepsis: Pathophysiology and clinical management. BMJ 2016, 353, i1585. [Google Scholar] [CrossRef]
- Yang, H.; Zhang, Z. Sepsis-induced myocardial dysfunction: The role of mitochondrial dysfunction. Inflamm. Res. 2021, 70, 379–387. [Google Scholar] [CrossRef] [PubMed]
- Rittirsch, D.; Huber-Lang, M.S.; Flierl, M.A.; Ward, P.A. Immunodesign of experimental sepsis by cecal ligation and puncture. Nat. Protoc. 2009, 4, 31–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.S.; Shibata, Y.; Malhotra, A.; Dutta, A. A novel class of small RNAs: tRNA-derived RNA fragments (tRFs). Genes Dev. 2009, 15, 2639–2649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, L.; Gan, M.; Tan, Z.; Jiang, D.; Jiang, Y.; Li, M.; Wang, J.; Li, X.; Zhang, S.; Zhu, L. A Novel Class of tRNA-Derived Small Non-Coding RNAs Respond to Myocardial Hypertrophy and Contribute to Intergenerational Inheritance. Biomolecules 2018, 16, 54. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.Y.; Li, P.F.; Li, Z.Q.; Tang, T.; Liu, W.; Wang, Y. Altered Expression of Transfer-RNA-Derived Small RNAs in Human With Rheumatic Heart Disease. Front. Cardiovasc. Med. 2021, 8, 716716. [Google Scholar] [CrossRef]
- Wang, J.; Han, B.; Yi, Y.; Wang, Y.; Zhang, L.; Jia, H.; Lv, J.; Yang, X.; Jiang, D.; Zhang, J. Expression profiles and functional analysis of plasma tRNA-derived small RNAs in children with fulminant myocarditis. Epigenomics 2021, 13, 1057–1075. [Google Scholar] [CrossRef]
- Liu, W.; Liu, Y.; Pan, Z.; Zhang, X.; Qin, Y.; Chen, X.; Li, M.; Chen, X.; Zheng, Q.; Liu, X.; et al. Systematic Analysis of tRNA-Derived Small RNAs Discloses New Therapeutic Targets of Caloric Restriction in Myocardial Ischemic Rats. Front. Cell Dev. Biol. 2020, 8, 568116. [Google Scholar] [CrossRef]
- He, X.; Yang, Y.; Wang, Q.; Wang, J.; Li, S.; Li, C.; Zong, T.; Li, X.; Zhang, Y.; Zou, Y.; et al. Expression profiles and potential roles of transfer RNA-derived small RNAs in atherosclerosis. J. Cell Mol. Med. 2021, 5, 7052–7065. [Google Scholar] [CrossRef]
- Sun, H.; Tong, Z.; Fang, Y.; Jiang, B.; Liang, P.; Tang, Y.; Li, Y.; Wu, Y.; Xiao, X. Nucleolin protects against doxorubicin-induced cardiotoxicity via upregulating microRNA-21. J. Cell Physiol. 2018, 233, 9516–9525. [Google Scholar] [CrossRef]
- Green, J.A.; Ansari, M.Y.; Ball, H.C.; Haqqi, T.M. tRNA-derived fragments (tRFs) regulate post-transcriptional gene expression via AGO-dependent mechanism in IL-1β stimulated chondrocytes. Osteoarthr. Cartil. 2020, 28, 1102–1110. [Google Scholar] [CrossRef]
- Fu, X.; He, X.; Yang, Y.; Jiang, S.; Wang, S.; Peng, X.; Tang, G.; Zong, T.; Li, X.; Zhang, Y.; et al. Identification of transfer RNA-derived fragments and their potential roles in aortic dissection. Genomics 2021, 113, 3039–3049. [Google Scholar] [CrossRef] [PubMed]
- Sen, P.; Gupta, K.; Kumari, A.; Singh, G.; Pandey, S.; Singh, R. Wnt/β-Catenin Antagonist Pyrvinium Exerts Cardioprotective Effects in Polymicrobial Sepsis Model by Attenuating Calcium Dyshomeostasis and Mitochondrial Dysfunction. Cardiovasc. Toxicol. 2021, 21, 517–532. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wang, L.; Xie, W.; Hu, S.; Zhou, H.; Zhu, P.; Zhu, H. Melatonin attenuates ER stress and mitochondrial damage in septic cardiomyopathy: A new mechanism involving BAP31 upregulation and MAPK-ERK pathway. J. Cell Physiol. 2020, 235, 2847–2856. [Google Scholar] [CrossRef] [PubMed]
- Czech, A.; Wende, S.; Mörl, M.; Pan, T.; Ignatova, Z. Reversible and rapid transfer-RNA deactivation as a mechanism of translational repression in stress. PLoS Genet. 2013, 9, e1003767. [Google Scholar] [CrossRef] [Green Version]
- Honda, S.; Loher, P.; Shigematsu, M.; Palazzo, J.P.; Suzuki, R.; Imoto, I.; Rigoutsos, I.; Kirino, Y. Sex hormone-dependent tRNA halves enhance cell proliferation in breast and prostate cancers. Proc. Natl. Acad. Sci. USA 2015, 21, E3816–E3825. [Google Scholar] [CrossRef] [Green Version]
- Schaefer, M.; Pollex, T.; Hanna, K.; Tuorto, F.; Meusburger, M.; Helm, M.; Lyko, F. RNA methylation by Dnmt2 protects transfer RNAs against stress-induced cleavage. Genes Dev. 2010, 24, 1590–1595. [Google Scholar] [CrossRef] [Green Version]
- Saikia, M.; Krokowski, D.; Guan, B.J.; Ivanov, P.; Parisien, M.; Hu, G.F.; Anderson, P.; Pan, T.; Hatzoglou, M. Genome-wide identification and quantitative analysis of cleaved tRNA fragments induced by cellular stress. J. Biol. Chem. 2012, 287, 42708–42725. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, L.; Tang, Y.; Yin, L.; Lin, X.; Luo, Z.; Wang, S.; Li, J.; Liang, P.; Jiang, B. Microarray Analysis Reveals Changes in tRNA-Derived Small RNAs (tsRNAs) Expression in Mice with Septic Cardiomyopathy. Genes 2022, 13, 2258. https://doi.org/10.3390/genes13122258
Yuan L, Tang Y, Yin L, Lin X, Luo Z, Wang S, Li J, Liang P, Jiang B. Microarray Analysis Reveals Changes in tRNA-Derived Small RNAs (tsRNAs) Expression in Mice with Septic Cardiomyopathy. Genes. 2022; 13(12):2258. https://doi.org/10.3390/genes13122258
Chicago/Turabian StyleYuan, Ludong, Yuting Tang, Leijing Yin, Xiaofang Lin, Zhengyang Luo, Shuxin Wang, Jing Li, Pengfei Liang, and Bimei Jiang. 2022. "Microarray Analysis Reveals Changes in tRNA-Derived Small RNAs (tsRNAs) Expression in Mice with Septic Cardiomyopathy" Genes 13, no. 12: 2258. https://doi.org/10.3390/genes13122258